
Abstract
This is the second of two articles about fiber-type composition of the human jaw muscles. It reviews the functional relationship of hybrid fibers and the adaptive properties of jaw-muscle fibers. In addition, to explain inter-individual variation in fiber-type composition, we discuss these adaptive properties in relation to environmental stimuli or perturbations. The fiber-type composition of the human jaw muscles is very different from that of limb and trunk muscles. Apart from the presence of the usual type I, IIA, and IIX myosin heavy-chains (MyHC), human jaw-muscle fibers contain MyHCs that are typical for developing or cardiac muscle. In addition, much more frequently than in limb and trunk muscles, jaw-muscle fibers are hybrid, i.e., they contain more than one type of MyHC isoform. Since these fibers have contractile properties that differ from those of pure fibers, this relatively large quantity of hybrid fibers provides a mechanism that produces a very fine gradation of force and movement. The presence of hybrid fibers might also reflect the adaptive capacity of jaw-muscle fibers. The capacity for adaptation also explains the observed large inter-individual variability in fiber-type composition. Besides local influences, like the amount of muscle activation and/or stretch, more general influences, like aging and gender, also play a role in the composition of fiber types.
INTRODUCTION
The variability in fiber-type composition in the same muscle in different individuals is large (Korfage et al., 2000, 2003a). Systemic differences between and among individuals, like age, gender, and/or hormones, can lead to differently adapted muscle fibers (see below). In addition, local activation- and stretch-related influences might contribute to variation in fiber-type expression (see Korfage et al., 2005). These latter influences also include behavioral differences—for instance, food type (hardness), bruxism, the use of artificial prostheses, etc.
The present article will address: (1) the role of hybrid fibers in the jaw muscles, (2) possible factors that explain the inter-individual variation in fiber-type composition, (3) differences between jaw muscles and limb/trunk muscles, and (4) differences in fiber-type composition of jaw muscles in different animals. The origin and functional significance of fiber-type diversity in the jaw muscles have been discussed in an accompanying article (Korfage et al., 2005).
(1) HYBRID FIBERS IN THE JAW MUSCLES
One of the remarkable features of the jaw muscles is the abundance of hybrid fibers, particularly in the jaw-closers (Fig. 1). These fibers have contractile properties that are intermediate between those of the myosin heavy-chain (MyHC) isoforms they express (Larsson and Moss, 1993; Bottinelli et al., 1996; Widrick et al., 1996). Hybrid fibers that express both MyHC-I and -IIA, for instance, will be faster than pure MyHC-I fibers but slower than pure MyHC-IIA fibers. Gel electrophoresis of single fibers (Fig. 2) allowed for the measurement of the relative proportion of each MyHC isoform (Korfage and Van Eijden, 2003b). The proportions of the individual MyHC isoforms within hybrid fibers varied highly. Although the actual contractile properties of the jaw-muscle fibers remain to be investigated, this variability suggests a continuous range of contractile properties among the jaw-muscle fibers.
Many hybrid fibers contain MyHC-cardiac α and MyHC-fetal, isoforms that are normally not found in limb and trunk muscles. The contraction velocity of MyHC-cardiac α lies between the velocities of MyHC-I and MyHC-IIA fibers (Kwa et al., 1995; Sciote and Kentish, 1996; Galler et al., 2002); the contraction velocity of MyHC-fetal seems to be low (D’Antona et al., 2003).
Functional Significance of Hybrid Fibers
The occurrence of hybrid fibers in considerable numbers is probably due to specific functional demands. Hybrid fibers could increase the capacity of the jaw muscles to produce a large variety of motor tasks, since they have contractile properties which lie between those of pure fibers. The greater the number of different hybrid fibers, the more a continuum exists in contractile properties which could contribute to a precise modulation of mandibular position and force. Whereas hybrid fibers represent a potentially unique design at the molecular motor level, motor unit recruitment schemes are also capable of providing smooth transitions throughout the contractile spectrum (see ‘size principle’, Korfage et al., 2005). The large number of hybrid fibers, as seen in the jaw muscles, raises two important issues with respect to the concept of motor units in human jaw muscles. First, it is obvious that, in these muscles, the concept of ‘one motoneuron, one MyHC isoform’ is not correct. Indeed, for motor units of the rabbit masseter muscle, it has been demonstrated that a single motor unit can have muscle fibers with different MyHCs (Kwa et al., 1995). Second, the large number of different hybrid fibers and their variable MyHC contents (see above) suggests that the relatively simple classification of motor units into three distinct categories—namely, S (slow, fatigue-resistant), FR (fast, fatigue-resistant), and FF (fast, fatigable) fibers (Barnard et al., 1971; Burke et al., 1971; Peter et al., 1972)—fails to hold for jaw muscles. It also indicates that human jaw-muscle motor units display a continuous range of properties with respect to contractile speed and fatigability, features which have also been demonstrated for the rabbit masseter (Turkawski et al., 1998; Van Eijden and Turkawski, 2001; Kwa et al., 2003).
Why are There So Many Hybrid Fibers in the Jaw Muscles?
Except for their functional significance, the presence of hybrid fibers could also reflect the adaptive capacity of jaw-muscle fibers. Muscle fibers have an inherent ability to alter their contractile properties to optimize the efficiency and economy of energy usage. They can change their phenotype in response to several influences by switching different isoform genes on or off. In mammals, the conversion of fiber types normally follows a strict order, from MyHC-I → -IIA → -IIX → -IIB, or vice versa (Schiaffino and Reggiani, 1994). During these transformations, the fibers will change from a pure fiber type, that expresses only one MyHC isoform, into another pure fiber type, via hybrid fiber types that express the old as well as the new MyHC isoforms. It might therefore be concluded that the jaw muscles are continuously switching from one fiber type to another. Some reports question this strict order of MyHC transformation. For instance, under certain conditions, like space flight (Talmadge et al., 1996), or hindlimb suspension, alone (Stevens et al., 1996) or in combination with hyperthyroidism (Caiozzo et al., 1998), some hybrid fibers were found in limb muscles that express MyHC-I and -IIX but not MyHC-IIA. This particular hybrid fiber type is also found in the jaw-closing muscles (Korfage and Van Eijden, 1999, 2000, 2003b). Thus, jaw-closing muscle fibers also seem to diverge from this strict order of MyHC transformations.
The MyHC expression within a fiber is not necessarily homogeneous. Muscle fibers contain many nuclei, and, therefore, some of these nuclei might express a different MyHC isoform, one not expressed in other parts of the muscle fiber. This was noticed in fibers of denervated rat limb muscles (Schiaffino et al., 1988) in which MyHC-fetal was regionally expressed in some fibers. Also, detailed investigation of longitudinally cut fibers in limb muscles from very old people indicated that some fibers switch their fiber type along the length of the fiber, or contain areas or nuclear domains in which the MyHC expression was different from that in other parts of the fiber (Andersen, 2003). An examination of serial sections of muscle fibers for a considerable length could confirm whether this is also true for the jaw muscles. The functional implication of this particular diversity is not known.
A possible explanation for the many hybrid fibers in the jaw muscles could be due to a difference in innervation as compared with limb and trunk muscles, namely, branchial vs. spinal (Stål et al., 1987). However, this explanation is questionable, because it has been shown that several muscles which are innervated by branchial nerves, like the digastric and the mylohyoid, contain only a few hybrid fibers that express MyHC-fetal and/or -cardiac α (Korfage and Van Eijden, 2000).
The many hybrid fibers in adult jaw muscles could also be regarded as a characteristic of development (Butler-Browne et al., 1988). During development, and also during regeneration, fibers can change their phenotype, during which hybrid fibers arise. During these processes, the MyHC-fetal gene is, temporarily, up-regulated. Still, this does not explain why these muscles continue to appear as developing muscles.
We would like to suggest another possible explanation. There is evidence that the membranes of normal muscle fibers are occasionally ruptured by stretch and/or stimulation (Petrof et al., 1993), which stimulates satellite cells to repair the damage. During the regeneration process, fibers transiently start to express MyHC-fetal and MyHC-I (Yang et al., 1997). The continued expression of these fiber types in adult jaw muscles might indicate a longer time period for jaw muscles to heal. Indeed, the masseter is known to heal badly after an injury (Pavlath et al., 1998), due to a lower proportion of satellite fibers in this muscle. Furthermore, these satellite cells might be different from satellite cells found in limb and trunk muscles with respect to their genetic make-up, as is the case with the muscle fibers. This is reflected in the expression of transcription factors, like Pax-3, myf5, and MyoD, which are needed to express specific muscle proteins during development and during activation of satellite cells. For instance, a genetic difference was noticed after ablation of the Pax-3 transcription factor in mice which led to an absence in the formation of muscle fibers in the body, but not in the jaw muscles (Tajbakhsh et al., 1997). Also, the timescale for the expression of transcription factors can be different. For instance, in chick somites, the interval between the onset of myf5 and the expression of MyoD is only a few hours. In cranial muscles, however, this interval is 8–18 hours (Noden et al., 1999). Furthermore, the onset of MyHC expression is also delayed in the cranial muscles. Thus, it could be that the satellite cells of the jaw muscles and the jaw-muscle fibers are also slower in the production of MyHC isoforms. This might result in a longer presence of hybrid fibers, including those hybrid fibers that co-express MyHC-fetal.
Alternatively, hybrid fibers can be seen as a legitimate phenotype, or an allotype, as named by Hoh (1991), and not as a transitional phenotype. A fiber integrates several of the stimuli it receives—like neural activity, hormones, stretch, etc.—and produces a response to them that signals the expression of MyHC gene transcription. Variations in the exact nature of the stimuli, and the variation in responsiveness to these stimuli, can be different between jaw muscles and limb/trunk muscles.
Many hybrid fibers in the jaw muscles co-express MyHC-cardiac α (Bredman et al., 1991; Sciote et al., 1994; Korfage and Van Eijden, 1999, 2000). In the masseter of the rabbit, this particular fiber type is often co-expressed with either MyHC-I or MyHC-IIA, and it was thus thought that MyHC-cardiac α is an intermediate isoform between the slow MyHC-I and the fast MyHC-IIA isoforms (Hämäläinen and Pette, 1997; Peuker et al., 1998). Jaw-muscle fibers can use this MyHC isoform as a ‘fine-tuning’ link protein between the slow-type MyHC-I and the fast-type MyHC-IIA isoforms. It is still not fully known why this particular MyHC isoform is up-regulated in the jaw muscles.
(2) DIFFERENCES IN FIBER-TYPE COMPOSITION BETWEEN AND AMONG INDIVIDUALS
There are large differences in fiber-type compositions of the various human jaw muscles (Table). One of the major causes for these differences is a difference in methods used in the various studies to classify fiber types. In several studies, the ATPase histochemistry method was used. With this method, the hybrid fibers were classified as IIC or IM fibers, depending on the differences in their staining patterns. Furthermore, ATPase histochemistry does not stain fibers expressing MyHC-fetal and MyHC-cardiac α. Also, some of the earlier studies did not, or could not, make a distinction between type IIA and type IIX fibers.
The observed variability in fiber-type composition in the same muscle in different individuals is great (Korfage et al., 2000). Except for genetic influences, there are several explanations for this variability. Systemic differences between and among individuals can lead to differently adapted muscle fibers. For instance, fiber-type composition is related to age and gender (see below). In addition to adaptation brought about by systemic influences, there are also local, probably activation- and stretch-related, influences, which might be related to fiber-type plasticity and the large inter-individual variation in fiber-type expression. These latter influences include behavioral differences—for instance, food type (hardness), bruxism, the use of artificial prostheses, etc. Some possible explanations for differences in fiber-type composition due to adaptation are discussed below.
Aging
Aging leads to systemic changes and alterations in muscle use that have a marked influence on skeletal muscles (reviewed by McComas, 1998; Basu et al., 2002). In limb and trunk muscles, it coincides with a reduction of muscle cross-sectional area, synthesis rate for MyHC (Balagopal et al., 1997), muscle strength [estimated at about 10% per decade starting at the age of 50 years (Larsson et al., 1979)], and shortening velocities of fibers (Larsson et al., 1997). The muscle fatigability increases, and the fiber-type proportions and fiber cross-sectional areas change (Lexell, 1993). Furthermore, aging coincides with a loss of motoneurons (Larsson, 1998). In limb and trunk muscles, reduction of muscle force is mainly the result of a loss of fibers and, to a lesser extent, to a reduction in fiber size, especially among the fast type II fibers. The proportion of type I fibers remains more or less the same, while the proportion of type IIA fibers increases at the cost of the type IIX fibers (Monemi et al., 1999). It must be noted, though, that the properties of the MyHC isoforms may change during aging. It has been reported that many of the MyHC-I fibers in older subjects were either very slow or very fast (Krivickas et al., 2001). There is also an increase in the population of hybrid fibers in elderly people (Klitgaard et al., 1990), especially of hybrid fibers co-expressing MyHC-I and -IIA (Andersen, 2003).
In jaw muscles, aging coincides with a prolongation of muscle contraction time (Newton et al., 1993), a reduction of the reflex responses, both in number and in amplitude, and an increased latency of oral reflexes (Smith et al., 1991; Kossioni and Karkazis, 1994). The cross-sectional area of the muscles is reduced with age (Newton et al., 1987), primarily due to a decrease in the cross-sectional area of the fibers (Monemi et al., 1998).
The fiber-type composition of the jaw muscles also changes with age. In the masseter of elderly subjects, the proportion of pure type I fibers is decreased, while the proportion of pure type II fibers is increased (Eriksson and Thornell, 1983; Monemi et al., 1999). Furthermore, the proportion of hybrid fibers in the jaw-closing muscles of elderly subjects increases, particularly the proportion of fibers that co-express MyHC-fetal (Monemi et al., 1996). This might contribute to the relatively high proportion of MyHC-fetal-positive hybrid fibers found in the study by Korfage et al.(2000).
Hormones
Some of the changes during aging are related to a change in the level of hormones. For instance, the levels of circulating anabolic hormones—like thyroid hormone (Rubenstein et al., 1973), growth hormone, and insulin-like growth factor (IGF-I)—are reduced with age (Proctor et al., 1998).
Thyroid hormone is important for the maturation of muscle fibers during development. Hypothyroidism in the rat embryo (d’Albis et al., 1990) delays the MyHC isoform transition in all examined muscles, particularly in the sexually dimorphic muscles like the masseter (see next paragraph). Furthermore, it was noticed that hypothyroidism in adult rats caused an up-regulation of MyHC-fetal in the masseter but not in other muscles (Izumo et al., 1986). In adult humans, the concentration of thyroid hormone decreases with age (Rubenstein et al., 1973), which might induce the reported increase of MyHC-fetal in the jaw-closing muscles of elderly individuals.
Growth hormone is another hormone that might have an influence on the fiber-type composition (see Korfage et al., 2005).
Testosterone is a hormone that has a large influence on the jaw-muscle fibers. Castration of male adult rabbits has no significant effect on the fiber-type proportion of the masseter. However, castration of young adult rabbits results in a muscle fiber distribution similar to that in females. This can be reversed when these rabbits are briefly exposed to testosterone (Reader et al., 2001; English and Schwartz, 2002). Jaw-closing muscles were found to be sexually dimorphic in the masseter of the rabbit (English et al., 1998, 1999; Eason et al., 2000b), rat (Ramamani et al., 1999), and mouse (Eason et al., 2000a), and in the temporalis of the guinea pig (Lyons et al., 1986). Male rabbit masseters have a significantly larger proportion of fast fibers than do female rabbit masseters (English et al., 1998). It has been suggested that changes in motoneuron properties are (partly) responsible for the androgen-induced changes in muscle-fiber properties in the rabbit masseter (English and Widmer, 2003). In mice, the male masseter has twice as many fibers containing MyHC-IIB as does the female masseter, which in turn has twice as many fibers containing MyHC-IIA (Eason et al., 2000a). Furthermore, in the rabbit, by immunohistochemistry, it was possible to distinguish 4 different subtypes of MyHC-I: I1, I2, I3, and I4 (English et al., 1998, 1999; Eason et al., 2000b). This could be the result of post-translational modifications of the MyHC molecule (Pol-Rodriguez et al., 2001). The proportions of these individual subtypes differ between male and female rabbits. Subtypes of MyHC-I were also found to exist during development in limb and trunk muscles, but, in these muscles, were converted into one single MyHC-I isoform (Hughes et al., 1993).
In humans, differences between males and females in the proportions of fiber types of the jaw muscles have been described in young adults. It was found that the male masseter had a larger number of type II fibers, while the female masseter had a larger number of type I and IM fibers (Tuxen et al., 1999).
Food
A recent study (Kemsley et al., 2003) showed that there is a large variation in the way people activate their jaw muscles during so-called ‘free-style’ chewing. The redundancy of muscles and muscle portions provides several ways in which they can be activated to produce a certain jaw movement (Koolstra, 2002). This difference in activation is probably reflected in the large individual variation in fiber-type composition, as reported in many studies.
The hardness of daily food could also have an influence on the phenotype of jaw-muscle fibers. However, to our knowledge, no study has compared the influence of food hardness on the fiber-type composition and fiber cross-sectional areas of jaw muscles in humans. There are numerous studies on jaw muscles of animals that were fed either a hard or a soft diet. It has been demonstrated that long-term intake of an easily chewable, fine-grained diet results in degeneration of muscle fibers and muscle spindles in the masseter of mice (Maeda et al., 1987, 1990). Experiments with rats (Miehe et al., 1999; Saito et al., 2002) demonstrated that a soft diet, which requires less masticatory effort, results in a more MyHC-IIB-rich phenotype in the masseter muscle than does a hard diet. Type I fiber proportion and fiber cross-sectional area in the deep masseter did not differ between rats that were fed a normal and those fed a soft diet (Kiliaridis et al., 1988), although more type IIB and fewer type IIA fibers were noted in two regions of the deep masseter of rats that were fed a soft diet. Such differences were not seen in the digastric muscle of these animals.
The cross-sectional area of the slow fibers decreased significantly in the masseter of rabbits that were fed a soft diet for three months compared with those fed a diet of hard pellets (Langenbach et al., 2003). These two animal groups did not exhibit differences in fiber-type composition. The latter study focused only on differences between type I and type II fibers, and fibers that expressed MyHC-cardiac α. There could have been differences within the type II fiber group.
A more rigorous approach was made in a study (Maxwell et al., 1980) in which all molars and incisors were extracted in monkeys (Macaca mulatta). Four and a half years after the extraction, not only was there a significant decrease in the fiber cross-sectional area of the slow fibers in the masseter and temporalis muscles, but also there was a decrease in the proportion of slow fibers and an increase of fast, fatigable fibers in the posterior muscle part of the masseter. It is thus possible to see an adaptation of the fiber type a long time after the triggering event.
Adaptation Influences in Individuals Wearing Artificial Dentures
Generally, individuals with removable partial or complete dentures have a significant reduction in bite force and masticatory function (Fontijn-Tekamp et al., 2000). This might be reflected in the MyHC isoform proportion and the fiber cross-sectional areas of the jaw-muscle fibers. The possible changes are assumed to be similar to those induced by a soft diet. No studies have yet been performed that compared fiber-type proportion and cross-sectional area of edentulous and dentulous humans. It is likely that edentulous individuals have smaller muscle fiber cross-sectional areas, and that the fibers become faster.
Craniofacial Morphology
The variation in facial form is large among humans. Differences in skull shape are associated with differences in masticatory performance (Van Spronsen et al., 1992). For instance, individuals with an excessively large anterior face height, the so-called ‘long-face syndrome’, produce a significantly smaller maximum molar bite force than do those with a normal facial height (Proffit et al., 1983). This might be reflected in a difference in fiber-type composition. According to one study (Boyd et al., 1984), long-face individuals have a higher percentage of fast fibers in some areas of the masseter, but this is refuted in another study (Shaughnessy et al., 1989). This discrepancy in results can possibly be explained by the large inter-individual and intra-individual variations in fiber types of the jaw muscles (Korfage et al., 2000).
In animal studies, it is easier to investigate the relationship between craniofacial morphology and jaw-muscle fibers, for instance, by surgically manipulating the position of the mandible. After the vertical dimension of the jaws of guinea pigs was increased, their masseter muscles became slower and had a decreased ATPase activity (Paik et al., 1993). Protrusion of the mandible in rats led to an adaptation of the lateral pterygoid and the superficial masseter (Easton and Carlson, 1990). The lateral pterygoid had a significantly greater area occupied by type I fibers. The superficial masseter, in contrast, showed a significant increase in the area occupied by type IIA instead of type IIB fibers. It was concluded that both muscles became slower. A similar experiment in pigs led to an increase in the proportion of MyHC-I (and in mRNA for MyHC-I) in the anterior region of the masseter and in the posterior region of the temporalis (Gedrange et al., 2001). The fibers in these regions might experience more stretch.
(3) DIFFERENCES BETWEEN JAW MUSCLES AND LIMB/TRUNK MUSCLES
There are several differences between jaw muscles and limb and trunk muscles. First, the jaw muscles contain many hybrid fibers, in contrast to limb and trunk muscles. Many of these fibers co-express MyHC-fetal and/or MyHC-cardiac α. Second, there is a difference in the fiber diameter between the two muscle groups. Type II fibers are larger than type I fibers in limb and trunk muscles, while in jaw muscles the opposite is true (Polgar et al., 1973).
In addition to these differences, the question that can be raised is whether the MyHC-I isoform present in jaw muscles is the same as the one in limb and trunk muscles. For instance, the type I fibers in the masseter are reported to be slower than the same fiber type in limb and trunk muscles (Morris et al., 2001). Furthermore, the variability in contractile properties among the type I muscle fibers is larger in the masseter than in limb and trunk muscles. This could mean either that, in the jaw muscles, there are more MyHC-I isoforms or that there is a post-translational adaptation of MyHC-I—for instance, by glycation (Ramamurthy et al., 2001), deamination (Balagopal et al., 1997), or phosphorylation (English et al., 1998). It must be noted that, in the rabbit and guinea pig, monoclonal antibodies could identify at least 4 different subtypes of MyHC-I (English et al., 1998). In the temporalis and lateral pterygoid muscles (Korfage and Van Eijden, 1999, 2000), some MyHC type I fibers, which were named MyHC-la, had an antibody-staining pattern which suggested that there are more subtypes of MyHC-I. Whether there are indeed subtypes of MyHC-I expressed in human jaw muscles, or whether MyHC-I is post-translationally adapted, is not yet clear.
Differences in contractile properties between fibers of the same type in jaw muscles and limb and trunk muscles might also be related to a difference in the expression of the light-chain of the myosin protein (MyLC). In the human masseter, 4 different essential and regulatory MyLCs are expressed, namely, MyLC-1s, MyLC-2s, MyLC-1f, and MyLC-1emb/atrial (Soussi-Yanicostas et al., 1990; Stål et al., 1994). The latter MyLC isoform is transiently expressed in fetal limb and trunk muscles. However, it is always present in jaw muscles. Furthermore, in limb and trunk muscles, MyLC-2f and MyLC-3f are expressed, but they have not been found in the masseter.
In the rat, the large variability in contraction velocity of limb muscle fibers can mainly be accounted for by the essential MyLC composition of the fibers (Bottinelli et al., 1994; Bottinelli and Reggiani, 1995), in contrast to human limb muscles (Larsson and Moss, 1993). This might be due to the fact that MyHC-IIB, which is found in rat but not in human muscles, is more sensitive to the regulatory MyLC isoform than other MyHC isoforms. However, the unusually slow MyHC type I fibers found in the human masseter might be related to the expression of the MyLC-embryonic isoform (Sciote et al., 2003). More study is needed to solve the question as to how jaw muscles differ from limb and trunk muscles in relation to these factors.
Generally, fibers in the jaw muscles are smaller than fibers in limb and trunk muscles. Since smaller cross-sectional areas facilitate an increase in the exchange of O2 and nutrients, the resistance to fatigue might thus be improved, especially in the fast fibers. Consequently, the smaller muscle fibers may be advantageous for the jaw muscles.
The origin of the differences in fiber diameters between jaw muscles and limb and trunk muscles could be related to the homeobox-containing gene Engrailed-2 (En-2). The presence of this gene has been demonstrated in the masseter, temporalis, and the medial and lateral pterygoid muscles of the mouse (Degenhardt and Sassoon, 2001; Degenhardt et al., 2002), and in some branchial arch muscles of the zebrafish (Hatta et al., 1990). The reporter of this gene is specifically expressed in myoblasts in the first branchial arch and is maintained in adult jaw-closing muscles only. However, in the anterior digastric and the tensor tympani, which are also derived from the first branchial arch, just as the jaw-closing muscles, this reporter gene was not detected. In transgenic mice, in which all muscle fibers expressed En-2, all fibers decreased in cross-section. An over-expression of this En-2 in fast muscles coincided with a decrease of the fast fibers, and also with a shift in the metabolic properties of these fibers. The specific expression of En-2 in the jaws of normal, non-transgenic species can therefore play a role in specifying muscle-fiber characteristics that contribute to the physiologic properties of specific muscle groups (Degenhardt and Sassoon, 2001). Recently, the smaller fiber diameter of the fast-fiber types in humans was suggested to be related to a mutation in the MYH 16 gene (Stedman et al., 2004). All non-human primates investigated had a high level of MYH 16 protein predominantly expressed in the fast fibers of muscles that are derived from the embryonic first pharyngeal arch. The frameshift mutation of the human MYH 16 gene was said to result in an eight-fold reduction in the size of the type II fibers of the jaw muscles.
(4) DIFFERENCES IN THE FIBER-TYPE COMPOSITION OF THE JAW MUSCLES IN DIFFERENT ANIMALS
The previous discussion dealt with studies in which animals were used, since it would be unethical or impractical to conduct certain tests in humans. A comparison of human skeletal muscles with those of animals can be difficult. In general, large animals have more fibers that express MyHC-I than do small animals (Pellegrino et al., 2003). The result of the adaptation of muscle fibers to a new demand can, therefore, be quantitatively different among animals. For instance, in an animal in which the masseter contains predominantly fibers that express MyHC-IIX, there will, as a response to more activation and stretch, probably be a change in fiber types toward MyHC-IIA but not to MyHC-I (Korfage et al., 2005). An animal with a large proportion of MyHC-IIA fibers might convert its fibers to MyHC-I. Although many studies have investigated the fiber-type transformation in several species, they are difficult to compare quantitatively.
The general rule for limb and trunk muscles, that large animals have a higher proportion of slow fibers than do small animals, is also true for the jaw muscles. The masseter of the mouse, for instance, contains only fast fibers, while that of a large animal like the cow contains only slow fibers. The cow, which is a ruminant, is thus well-adapted for slow, long-term muscle exercises. The masseters of the cat, dog, and pig have a mixture of slow and fast fibers, although the fast-fiber types predominate (Tuxen and Kirkeby, 1990). A special fiber-type composition has been found in the masseter of the kangaroo. This species, which has more or less the same skull shape as sheep, and also eats the same type of food, has a masseter which predominantly expresses MyHC-cardiac α (Hoh et al., 2000). In contrast, the sheep’s masseter consists predominantly of MyHC-I fibers. This difference has been attributed to a need for a higher velocity for grinding food, because, in contrast to the sheep, the kangaroo does not ruminate.
In some animals, a special MyHC isoform, ‘superfast’ or MyHC-IIM, has been identified. This MyHC isoform is found in the jaw-closing muscles of the cat, the American opossum (Rowlerson et al., 1981; Sciote et al., 1995), five species of Carnivora, six species of primates, not including man (Rowlerson et al., 1983), and of the caiman and terrapin (Rowlerson, 1994). A more complete list of animals in which the jaw-closing muscles express MyHC-IIM has been reviewed by Hoh (2002). The maximum contraction velocity of MyHC-IIM fibers is higher than that of MyHC-IIB fibers. The occurrence of MyHC-IIM in animals is associated with an aggressive bite which is required for predatory or defense purposes (Rowlerson et al., 1983).
(5) FINAL REMARKS
The architectural and anatomical complex jaw muscles contain many fiber types that are normally not seen in large quantities in limb and trunk muscles. These fiber types differ in force production at various contraction velocities. The variation in fiber type enables the jaw muscles to produce an accurately controllable force over a wide range of contraction velocities. The dynamic nature of muscle fibers allows them to change their phenotype to optimize contractile function and energy uptake. This explains the variability in fiber-type proportions found between and among individuals, muscles, and muscle portions. The amount and pattern of activation and stretch probably have a large influence on the expression of the MyHC isoforms in the muscle fibers. Next to these influences, there are other internal and external influences which also can affect the fiber type.
These influences can have a different impact on jaw-muscle fibers than on those of limb and trunk muscles. However, the observed difference in MyHCs might also be related to a difference in genetic background. Although differences have been observed in the genes that are responsible for the development of jaw and other muscles, it is not yet known how these differences lead to the remarkable collection of fiber types that has been observed in the jaw muscles.
Fiber-type Composition (mean %) in Adult Human Jaw Muscles as Reported in the Literature
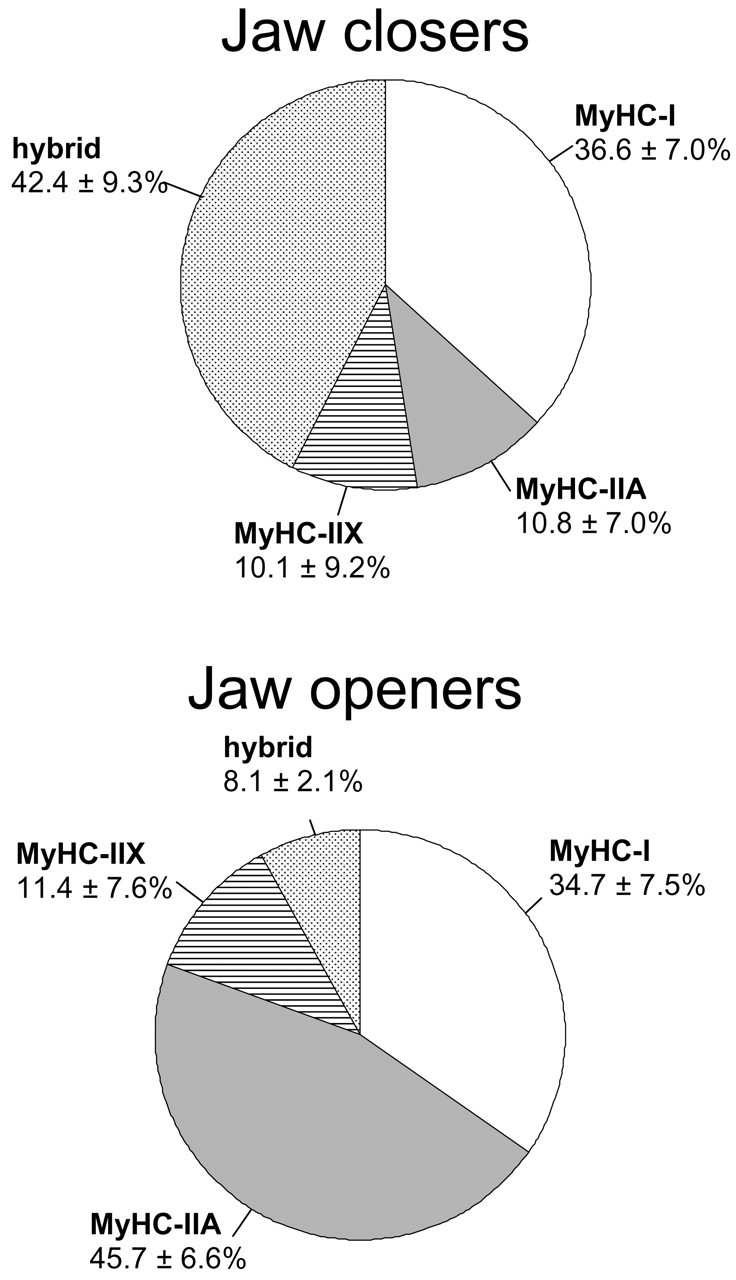
Distribution of pure and hybrid fiber types (mean ± SD) in the human jaw-closers (temporalis, masseter, pterygoids) and jaw-openers (mylohyoid, digastric, geniohyoid); SD values are a measure for inter-individual variability. Muscle samples (jaw-closers, 6 to 9 per muscle; jaw openers, 2 to 4 per muscle) were obtained from eight cadavers. Because the fiber-type distribution of the lateral pterygoid had more similarities with that of the jaw-closers than with that of the jaw-openers, this muscle was included among the group of jaw-closers. For more information, see Korfage et al.(2001). MyHC = myosin heavy-chain.
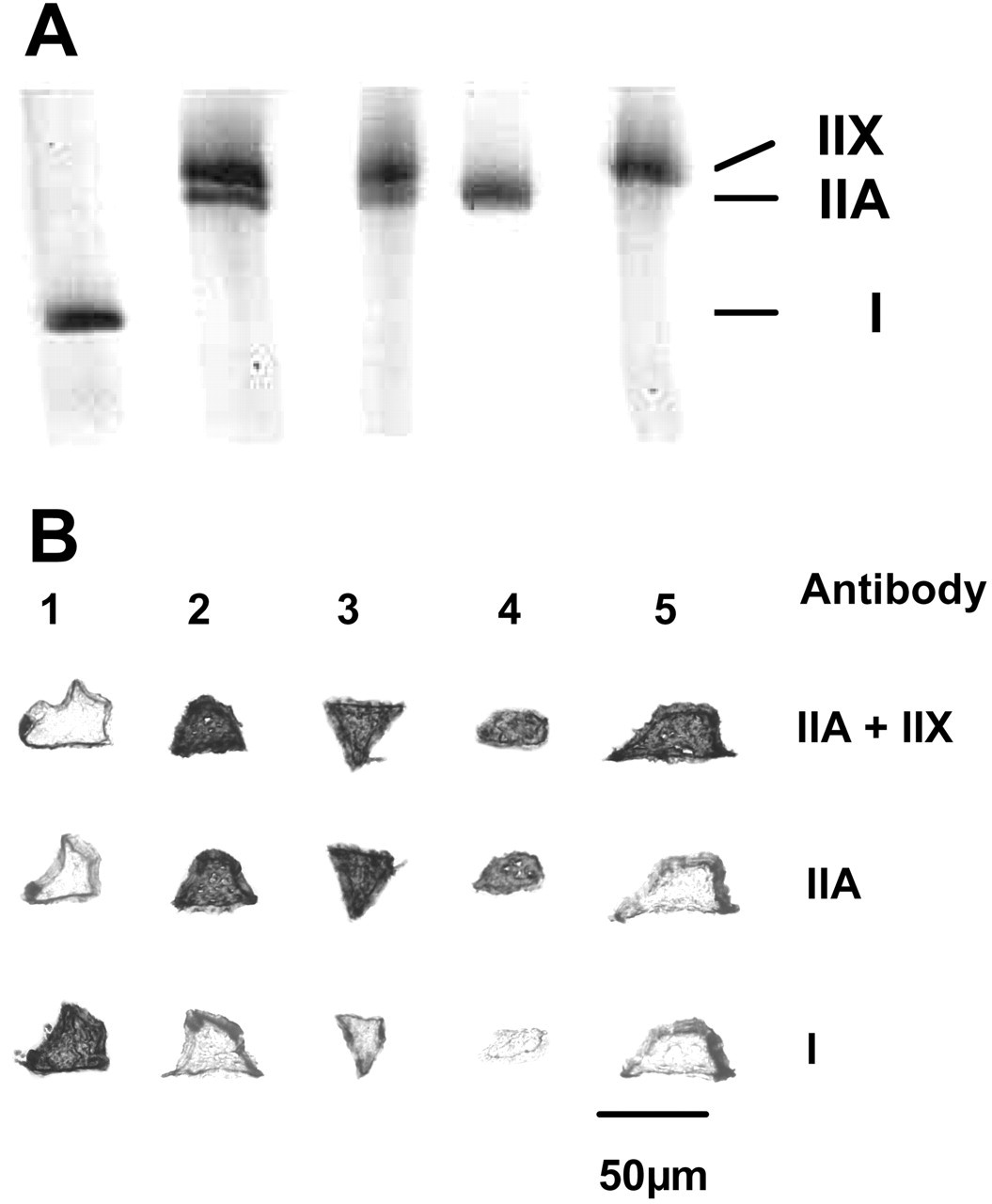
Electrophoretic
Footnotes
Notes
Acknowledgements
This research was supported by the Interuniversity Research School of Dentistry, through the Academic Center of Dentistry Amsterdam. We are grateful to Peter Brugman for technical assistance.