
Abstract
Hawai‘i and other island communities around the globe are experiencing high exposures to micro- and nanoplastics (MNPs). Widespread use of various types of plastics and the action of wave physics coupled with abundant silica creates a constant production of MNPs. The community health effects of exposure to MNPs are not yet fully understood. The aquatic vertebrate model zebrafish, Danio rerio, was utilized to evaluate the developmental and behavioral effects of nanoplastic (NP) exposure as well as visual confirmation of plastic accumulation after embryonic and larval exposure. Eye and yolk size, heart rate, angle of development, and early larval locomotion behavior were quantified after early NP exposure lasting 3 or 5 days. In this work, a digital laboratory environment is used as the context for research trainees spanning high school to graduate and medical school in Hawai‘i to explore health outcomes of developmental NP exposures. Presence of MNPs in the early developmental environment led to changes in growth and behavior of exposed fish. Data analysis showed significant effects of exposures on changes in the rate of development and yolk consumption on day 2 and reduced body length and increased locomotion on day 6. Nanoplastic exposure also affected overall body length, but this effect was not significant. These data support the potential dangers of MNP exposure and demonstrate their persistent detrimental effects on development and behavior in young zebrafish. Accordingly, our work contributes toward shifting focus toward understanding longer impacts on health, growth, and behaviors and potentially the reduction of harm in developing vertebrates.
Introduction
Home to over 1 million people, Hawai‘i remains one of the world’s top tourist destinations due to its majestic mountain landscape, idyllic beaches, deep history, and “Aloha” spirit. However, Hawai‘i and its people are highly vulnerable to micro- and nanoplastic (MNP) pollutants due to the effects on marine life and Hawai‘i’s high seafood consumption. Contributors to ocean plastic debris include agriculture, fishing industries, and rivers flowing through large cities to the ocean,1–7 all of which exist in Hawai‘i. Islands are particularly significant in the production of MNPs. The churn of silica in beaches with recurring waves drives the process of macro-level plastics being ground into ultrafine particles, the waves then act as a distributor of MNPs worldwide.8,9
As MNPs emerge as pressing public health and environmental concerns, research into their effects has become a global priority. These synthetic polymers recognized as anthropogenic pollutants are defined as plastic fragments measuring less than 5 mm.10,11 When their size falls within the 1 nm to 1 μm range, they are classified as nanoplastics (NPs). 12 The widespread use of plastics, driven by their lightweight and durable nature, has resulted in global plastic pollution and consumption. 13 Evaluation of means of exposure, tissues at risk, and developmental and behavioral effects of MNPs is critical in addressing ongoing health concerns worldwide.
Background
The physicochemical properties of MNPs, including their hydrophobicity, high porosity, and large surface area, facilitate their interaction with environmental contaminants. These particles concentrate and interact with hydrophobic organic contaminants, heavy metals, polycyclic aromatic hydrocarbons, pharmaceuticals, pesticides, other nanoparticles, and organic halogens, thereby intensifying the potential for toxic exposure in living organisms. 14 Due to their persistence as organic pollutants, MNPs lead to consequences of bioaccumulation and biomagnification.
Several studies have identified MNPs in various human tissues including the brain, liver, kidney, colon, lungs, heart, placenta, and testes.15–22 One study analyzing colectomy samples detected microplastics (MPs) in all specimens with an average of 331 particles per individual sample. 17 Alarmingly, MNPs have also been found to be selectively retained in the human brain. 15 Despite these findings, there remains a gap in research about the health risks linked with MNP exposure and accumulation, particularly with stress and anxiety responses.
With the high prevalence of MNPs already found in human and animal tissues, it has become imperative to determine how they are traced in respective tissues, their impacts on physical and mental health, and if they require immediate policy intervention. However, if intervention is required, determination of impacts in humans could delay progress as we await studies across the human lifespan.
On the contrary, MNPs found in and near waterbodies have been increasingly shown to affect early development, growth, and behavior in aquatic organisms, particularly during sensitive periods and organogenesis. Previous studies in developmental teleost models have reported developmental delays, altered heart rate, and increased inflammation and oxidative stress23–26 but there is no consensus on mortality and survival rates. 27 Behavioral alterations including changes in larval locomotor activity and impaired predator avoidance and disrupted shoaling in adult fish have also been reported after exposure to MNPs.23,24,28,29 Together, these findings indicate that exposure in aquatic species has physiological and behavioral consequences, and additional research is needed to fully understand the longer-term impact of plastics-contaminated environments on early and later development.
Danio rerio (zebrafish) are often used to study the effects of bioaccumulation and toxicity of environmental contaminants on humans because they share a 70% genetic homology. 30 They are also utilized due to their small size, ease of breeding, high fecundity, short life cycle, and low maintenance. Paired with their transparency, availability of transgenic lines, high-throughput sequencing, and genetic similarities to humans, zebrafish allow effective investigation of MNP accumulation and toxicity. 31
Behavioral responses to various biotic and abiotic stressors have been studied in larval and adult zebrafish previously and are well established. For example, Clark et al. 32 explain that stress assays in zebrafish typically involve acute or chronic exposure to controlled stressors and determination of their effects. These stressors include physical challenges such as handling, capture, net chasing, and other disturbances; social stressors, including dominant and submissive interactions with other fish and overcrowding or isolation; and environmental stressors like water temperature and quality measures.
Two primary stressor response measures include physiological and behavioral. Physiological stress measures typically involve quantification of cortisol levels. The behavioral stress measurement consists of analysis of changing behavior, such as changes in preference of place, locomotion style, and speed or even social interaction with other fish. Lee et al. 33 have previously outlined a reproducible behavioral assay to describe the stress response of larval zebrafish to environmental stressors via the animal’s locomotion used in this study.
Herein, we describe an analysis of the effects of NPs on the development and behavior of zebrafish in an effort to demonstrate their potential effects on animal (and human) health, driven by NMPs. This work is the product of an online laboratory including high schoolers, undergraduate, and medical and graduate students in Hawai‘i and an investigative team from Minnesota. 34 All imaging and video data were collected via a streaming laboratory connection and analyzed by the student researchers. The experimental groups and timeline are shown as the schematic representation of procedures in Figure 1.

Simplified schematic diagram of experimental setup and timeline. Top of the image
Methods
NPs solution preparation
To ensure standardization and replication of the plastics treatments, we used commercially made and available plastics that are guaranteed to be of the same-sized particles. 1% Fluorescent polystyrene nanoplastics (PS-NPs) with a diameter of approximately 0.044 μm or 44 nm (cat. #FSDG001) were purchased from Bangs Laboratories Inc. 10 ppm PS-NP embryo–water solution was freshly prepared in a glass container and stored out of light until unavoidable exposure during treatment (Fig. 1A). Treatment schedule, dosage, and stages were based on our own earlier preliminary work and published literature available at the time.23–26
Zebrafish husbandry and experimental setup
Adult Danio rerio (wild-type, Mayo Recessive Free) stock fish were kept in laboratory aquaria with a 28°C temperature and a recirculation system with a 14:10 light–dark cycle. Diet consisting of brine shrimp and dry feed was given three times a day. Two males and two females were kept overnight in a breeding tank, and embryos were collected the next morning immediately after spawning started with light onset. Zebrafish husbandry, handling, and experimental procedures followed standard practices and guidelines from the Institutional Animal Care and Use Committee (IACUC) at the Mayo Clinic (A34513-13-R22).
Fertilized eggs were collected and kept in a standard petri dish containing embryo medium solution for cleaning and sorting (Fig. 1A). At approximately 6 h post fertilization (hpf), once embryos were counted and cleaned, they were randomly assigned to treatment groups (three treatment groups and a fourth untreated control group). At this time, 50 embryos were put in each of the four experimental petri dishes, followed by the addition of 10 ppm PS-NP solution for the appropriate exposure times as stated below and giving a gentle swirl to allow some mixing within the media. The evaluation of effects of plastics exposure led to the selection of three levels of treatments (Fig. 1B): throughout 5 days of embryogenesis and larval growth, during the first 3 days of rapid early development only, during 3 days of late development only when most organogenesis is finished but larval growth continues (Kimmel et al.
35
). The 200 embryos were thus assigned to one of the following four groups (n = 50 for each):
Group 1: Treated 0–5 days post fertilization (dpf). Group 2: Treated 0–3 dpf. Group 3: Treated 3–5 dpf. Group 4: Control (no exposure to PS-NPs).
During the exposure protocol, all four embryos (treated and untreated) in petri dishes were kept in an incubator (30°C temperature) with a 12:12 light–dark cycle (lights on at 8:00), made possible by a strip of LED light connected to an automated programmable switch. All fish in the four groups were checked daily, particularly noting survival rate, hatching time, and any potential abnormalities. For each petri dish, 50% water changes were gently done (embryo water with appropriate NPs-treatment mixture) using new pipettes (to prevent cross-contamination). We expected ∼60%–70% embryos survival rate, typical of our facility, and any dead embryos and debris were removed to consistently maintain optimal water perimeters and to prevent fungal growth. The selection of embryos/larvae from each group for monitoring and assessments below was blinded when possible, and randomized and counterbalanced as appropriate across the various procedures.
As the subjects were in the larval stages during the entirety of our experiment, we were unable to determine sex but estimated it to be approximately 50:50, based on previous observation in the facility. Following exposure treatment, at the end of day 5, survival rates were noted, and zebrafish from each group were moved to new 48-well plates (one fish per well) for behavioral analysis.
Physiological assessment
Measurements of eye size, yolk size, heart rate, and angle of development were recorded at 24-h intervals from 24 to 96 hpf for Groups 1 and 4 (n = 20 for each, randomly selected, Fig. 1C) following well-established guidelines (as described by Kimmel et al. 35 ). Following hatching, tricaine was used as an anesthetic for heart rate measurements, which were carried out by counting each beat over a 15-s period in 20 randomly selected embryos from each of the two groups. A Zeiss microscope, equipped with green fluorescent protein filters, was used to screen the fish for fluorescence as an indicator of NP presence. At 3 dpf, one fish from each of the treatment and control groups was randomly selected for verification of the presence of fluorescence using light-sheet microscopy. Another set of five fish (one from each of the four groups) was selected for light-sheet microscopy done at 5 dpf.
Imaging of NPs exposure
Light-sheet microscopy was used on fish taken at 5 dpf to determine the presence/absence of NPs inside zebrafish tissues by an observer blind to the treatment group, following previously published studies. 36 Prior to imaging, all fish (untreated and NPs-treated) were rinsed twice to remove any remaining particles from the surface in the treated group. The larvae were then placed in a solution of tricaine and 1% agar. Brightfield images and z-stack maximum intensity projections were collected to determine the presence and location of the NPs in the fish.
Image analysis using ImageJ
The fluorescent images of NPs buildup were processed with the open-source image-processing software ImageJ/Fiji (Version 1.54h, developed by the National Institutes of Health). First, raw files to be examined were all opened at once and were adjusted simultaneously to allow visualization of the outline and other anatomical landmarks of the fish in all conditions. Built-in tools in the software (e.g., Adjust > Brightness/Contrast, Adjust > Window/Level, Propagate to all other open images option) were used to set the same brightness and contrast levels in the different images from the four groups and to standardize the fluorescence across the images. Also, Analyze > Set Scale and Analyze > Tools > Scale Bar functions and a scale reference image were used to globally apply the same set scale to all images and overlay the scale bar in the saved .png images.
Behavioral response to light stimulus
To assess reactions to a light stimulus, all zebrafish that survived to 5 dpf (n = 30 for each group) were placed in new 48-well plates (one fish per well) and acclimated overnight in the incubator (Fig. 1D). We observed 6 dpf fish using a Kestrel high-throughput parallel live-imaging system in the multi-camera array microscope (Romana Optics). After a 30-min acclimatization period in darkness, fish were video-recorded using the Kestrel imaging system and MCAM software in darkness for 27 min and exposed to a 2-min 3000 lux burst of white light between minutes 10–12. To measure response behaviors, data were compared during the first 10 min (baseline) before the light stimulus, during the 2-min light period (light exposure), and 17 min thereafter in the post-light dark period (post-light). Afterward, all larvae were euthanized according to the IACUC guidelines.
Data and statistical analyses
Statistical analyses were performed using IBM SPSS Statistics version 25 and graphed using GraphPad Prism version 10. Main effects of treatment and group differences were considered significant at an alpha level of 0.05, and 0.05 < p < 0.1 are also reported as potential trends that may prompt further investigation. Chi-square test was done to compare the survival rates at 5 dpf of the three treated groups of fish with their untreated control sibling group. Hypothesis testing was done using generalized linear models, followed by post-hoc tests if there was a significant main effect of treatment or any interactions between factors. Larval behavior videos were tracked using offline automated procedures in the Kestrel imaging system, and total distance moved per minute, average distance moved, as well as fish body lengths were quantified. Behavioral (distance and speed) and physiological (eye size, yolk size, heart rate, and angle of development) data were analyzed for main effects of NP treatment (between-subject factor) and time (minutes for behavioral data, days for physiological variables) using repeated-measures analyses of variance (ANOVAs). Any significant main effects or interactions were followed by post-hoc testing (t-tests or one-way ANOVAs) to determine directionality. Group differences in mean distance and body lengths were determined using univariate ANOVAs.
Results
PS-NPs effects on development
Survival rates did not differ among groups of fish treated with commercially available 1% PS-NPs (60% for treated 0–5 dpf, 62% for treated 0–3 dpf, and 58% for treated 3–5 dpf) compared with control (66%, chi-square tests, all p > 0.05).
Both control and treated groups showed an increase in eye size over time (Fig. 2A). When treated zebrafish larvae were compared with control fish, repeated-measures ANOVA showed a significant main effect of NPs, F(1, 18) = 15.526, p < 0.001, and a significant day × treatment interaction, F(3, 54) = 5.088, p = 0.008. With post-hoc one-way ANOVAs, we noted that the eye size was marginally different between treated and control fish on 2 dpf, F(1, 39) = 3.237, p = 0.08, and 3 dpf, F(1, 39) = 3.249, p = 0.079, but not on 1 and 4 dpf (p > 0.05 for both).
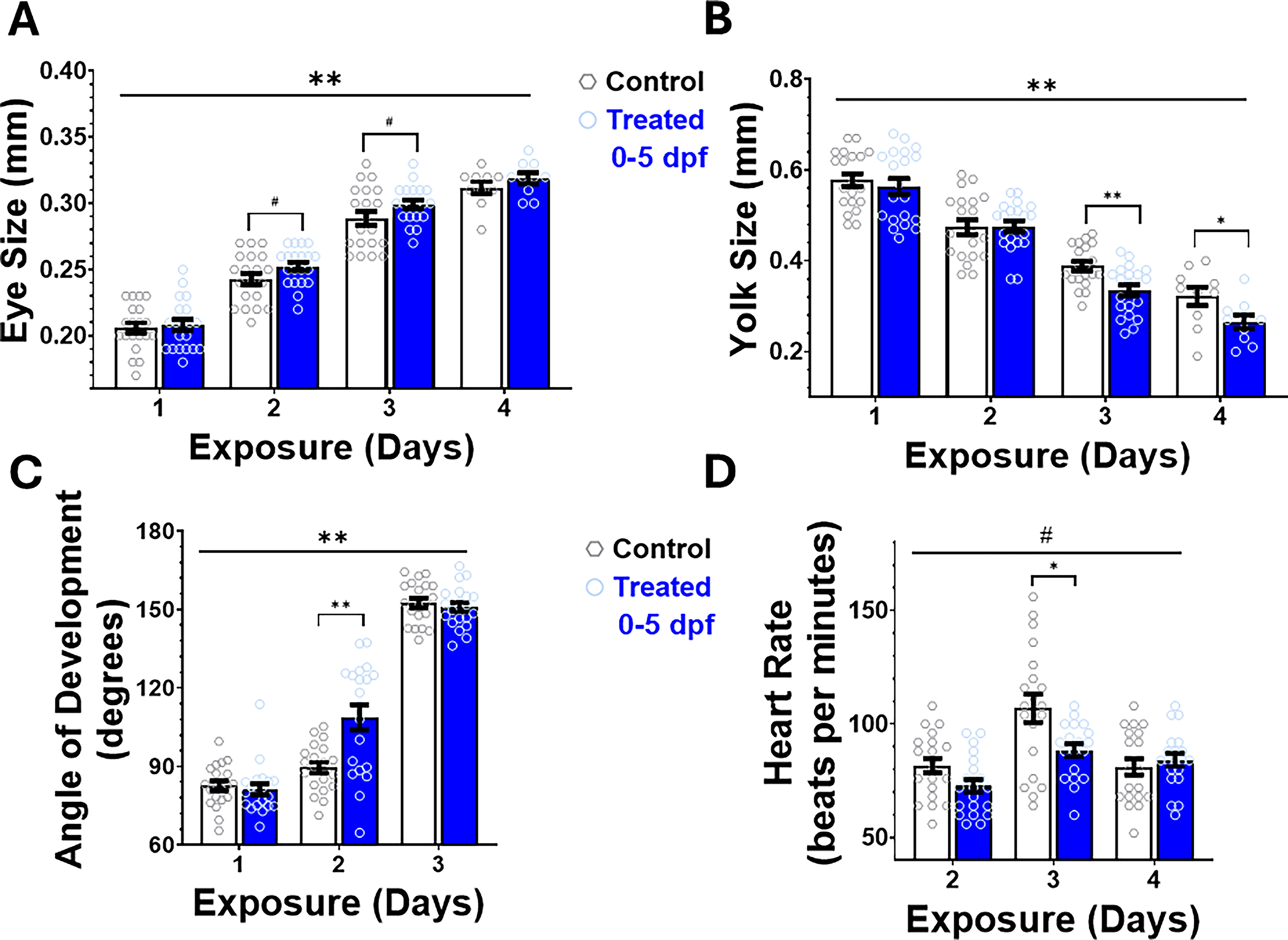
Physiological assessment of embryonic development. Daily evaluation of embryonic development for randomly selected fish from Group 1 (Treated 0–5 dpf, n = 20) and Group 4 (Control, n = 20) using a stereoscope are shown. Although all fish had an increase in eye size across development
Yolk size reduced over time in both groups (Fig. 2B). Treated fish had decreased yolk size relative to the control over the 4 exposure days. A repeated-measures ANOVA showed a significant effect of treatment, F(1, 18) = 10.609, p = 0.004, but no day × treatment interaction (p = 0.109). Post-hoc one-way ANOVAs showed significant group differences on 3 dpf, F(1, 38) = 11.45, p = 0.002, and 4 dpf, F(1, 38) = 4.90, p = 0.04, but not on earlier days (p > 0.05 for both 1 and 2 dpf).
Zebrafish larvae treated with PS-NPs had similar developmental angles as the control on days 1 and 3, and exceeded the control on day 2. A repeated-measures ANOVA showed a significant effect of treatment, F(1, 38) = 8.627, p = 0.006, and a significant day × treatment interaction, F(2, 76) = 9.397, p = 0.001. As depicted in Figure 2C, in post-hoc one-way ANOVAs, we observed that the treated fish had significantly larger angles on day 2, F(1, 38) = 13.98, p < 0.001, compared with control fish, but not on days 1 and 3 (p > 0.05 for both days).
Both groups experience their peak heart rate at 3 dpf, as illustrated in Figure 2D. Using repeated-measures ANOVA, we observed a significant day × treatment interaction, F(2, 74) = 6.492, p = 0.005, but the main effects of treatment did not reach statistical significance, F(1, 37) = 3.552, p = 0.067. Post-hoc one-way ANOVAs indicated that heart rate was significantly different between treated and control fish on 2 dpf, F(1, 39) = 4.239, p = 0.046, and 3 dpf, F(1, 38) = 7.165, p = 0.011, but not on 4 dpf (p > 0.05).
PS-NPs uptake
Representative light-sheet fluorescence microscopy images of 5 dpf larvae are shown in Figure 3 including the control (untreated) zebrafish larvae and fish exposed to a 10-ppm concentration of 44 nm sized PS-NPS for 0–3 dpf, 3–5 dpf, and 0–5 dpf. NPs fluorescent signal was not detected in the untreated control and the group treated for 0–3 dpf. Fluorescence was present in fish treated during 3–5 dpf and 0–5 dpf, with the latter showing the strongest signal. Fluorescence was present along the anterior and posterior lateral lines, the anterior macula, the gut, and the kidneys. Thus, the accumulation of PS-NPs within the treated larvae was higher, as the time of exposure continued, with the greatest concentration of PS-NPs visible in the group that was exposed from 0 to 5 dpf.
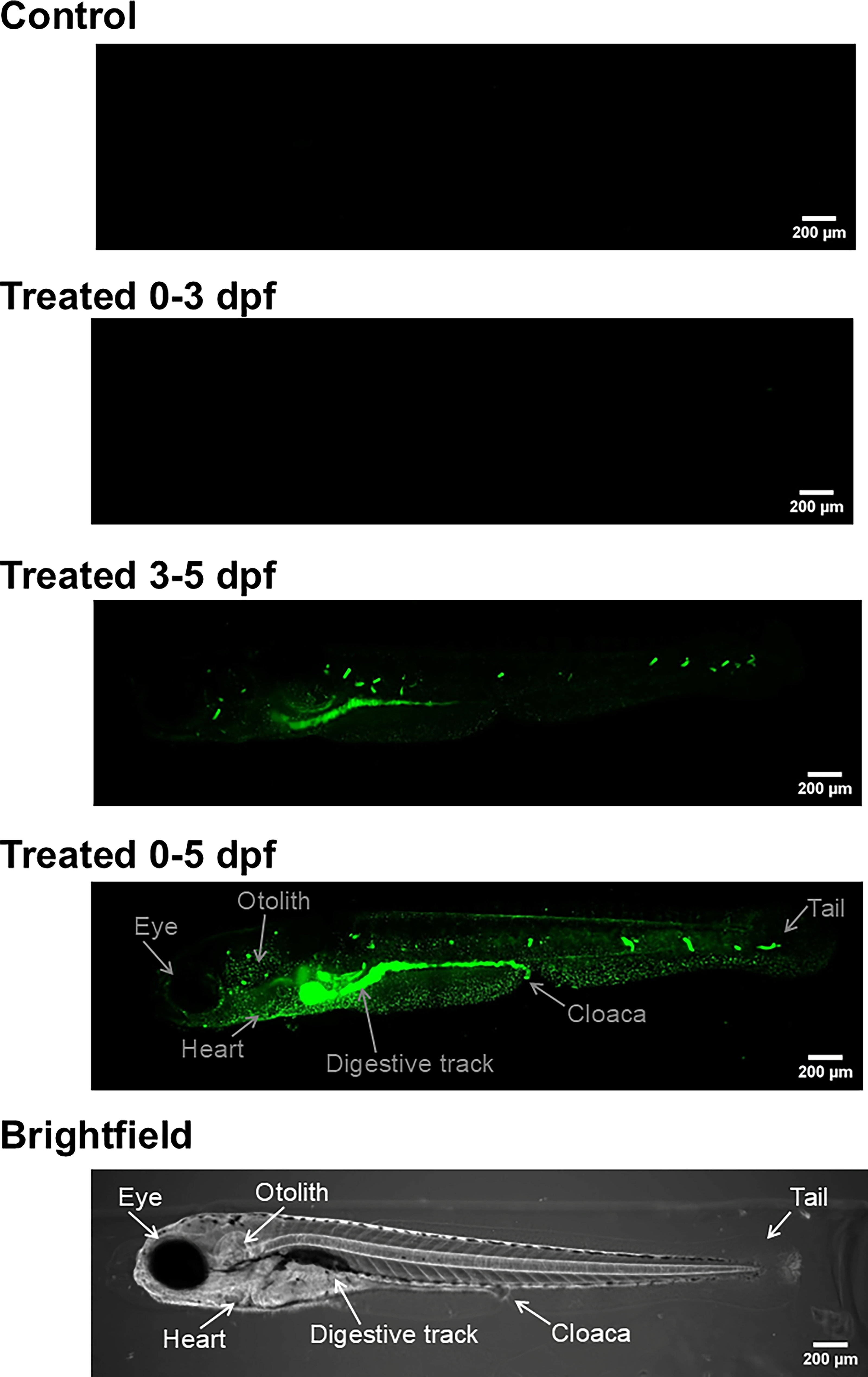
Microscopy. Representative light-sheet microscopy images of 5 dpf zebrafish larvae treated with a 10-ppm concentration of fluorescent polystyrene nanoplastics (44 nm) for control (untreated), and treated 0–3 dpf, treated 3–5 dpf, and treated 0–5 dpf. There was no fluorescent signal present in either the control or the group treated 0–3 dpf, indicating absence of nanoplastics. Fluorescent signal was present in both 3–5 dpf and 0–5 dpf treated fish, with the highest presence of fluorescence in the 0–5 dpf treated fish. Main anatomical structures (i.e., eye, otolith, heart, digestive tract, cloaca, and tail) are indicated on the Treated 0–5 dpf and brightfield images.
Automated behavioral observations
Automated behavioral tracking quantified distances moved by the treated and control group larvae during the 27 min of observation. As shown in the time plot in Figure 4A, all fish exhibited hypolocomotion in response to the light stimulus (minutes 10–12). When we compared the three PS-NP-treated fish with the untreated control fish, all plastic-exposed groups had slightly higher locomotion than control fish during baseline and post-light periods. With a repeated-measures ANOVA, we observed no significant effect of treatment, F(3, 119) = 1.160, p = 0.328, or minute × treatment interaction, F(78, 3094) = 1.460, p = 0.071.
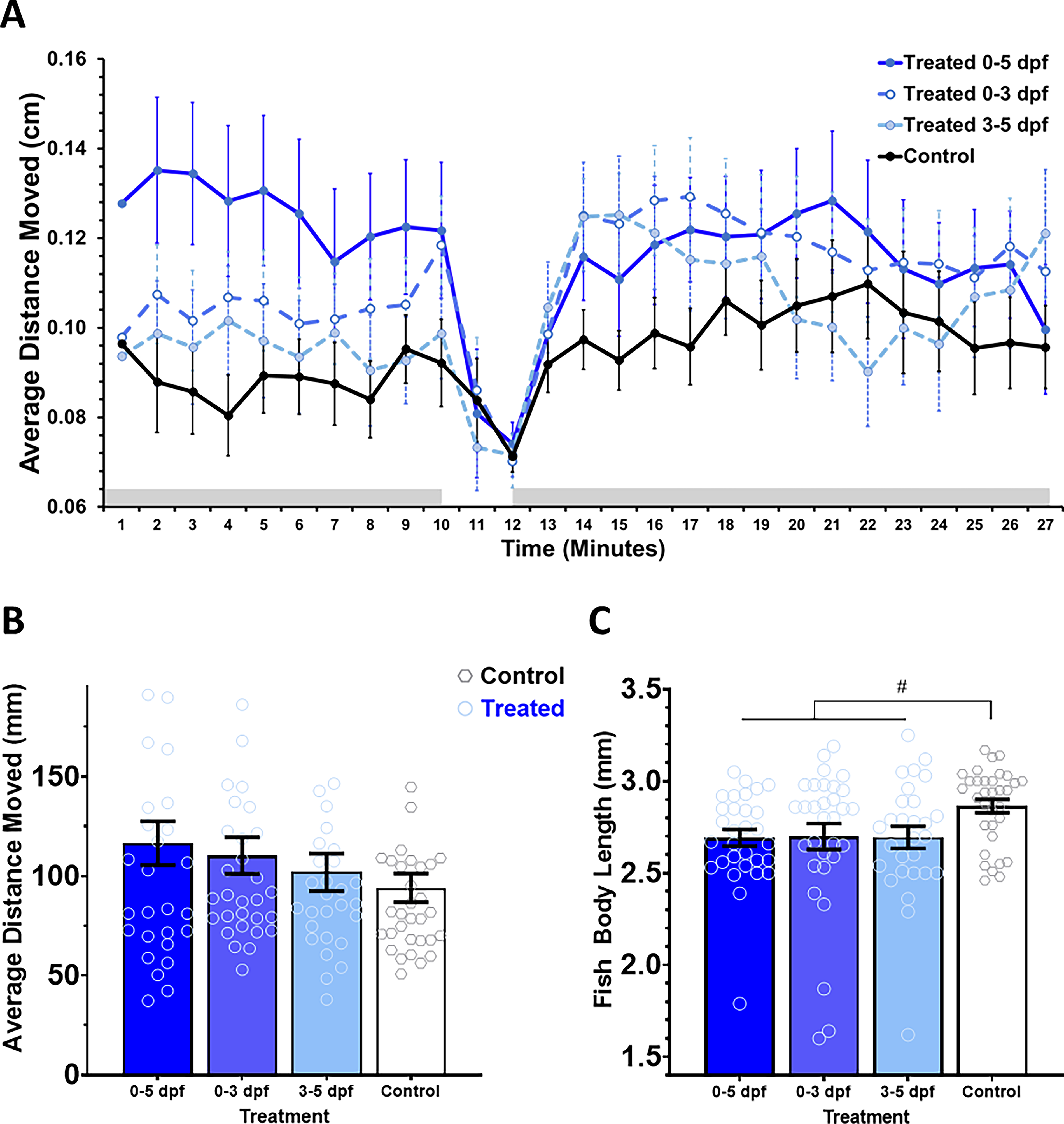
Locomotion and behavioral response to light stimulus.
We also compared group means for average distance moved per minute (Fig. 4B) and fish body length (Fig. 4C). Based on a univariate ANOVA, we noted that there was no significant effect of PS-NPs on average movement, F(3, 119) = 1.160, p = 0.328. Although all plastics-exposed fish had decreased body lengths, compared with control fish, a univariate ANOVA indicated a marginally significant effect of treatment, F(3, 118) = 2.603, p = 0.05.
Discussion
To estimate potential effects of NMPs on human health, we analyzed the effects of NPs on the development and behavior of larval zebrafish using a student-driven research training approach. The greater eye size seen in the treatment group, compared with the control fish, could be the phenotypic manifestations of increased NP exposure, as compromised structural integrity of the eyes following embryonic MPs exposure has been described before. 25 Recently, additional phenotypic alterations such as tail malformation and deformities of the yolk sac were also reported as outcomes of increased MNP exposure in zebrafish embryos.27,37 Such occurrences have been linked to oxidative stress and genotoxicity, with both subjects serving as the basis for the primary hypothesis on how MNPs affect development.23–26,36,37 Current data suggest that degradation products released by MNPs during late development may be linked to previously established transcriptional changes in genes related to notochord and bone development.25–29,37 Although several studies have provided evidence to support this hypothesis, more research is needed to fully understand the mechanism behind the effect of MPs on progressive stages during zebrafish development.
The smaller yolk size observed in treated zebrafish may be the result of increased metabolic activity and higher embryonic nutrient uptake stemming from the toxicological properties of NPs. This aligns with the findings that PS-NPs significantly increased food consumption in zebrafish larvae. 38 Expedited yolk absorption may potentially deplete yolk prematurely and lead to a perceived starvation environment. 39 PS-MPs at 1 and 3 μm have been found to accumulate in zebrafish yolks and to be associated with yolk deformities. 37 NPs may also lead to abnormal heart rate due to disruption in metabolic, cardiac, and endocrine pathways. Past studies have found both bradycardia and tachycardia in zebrafish larvae after exposure to PS-NPs.23,37,40 The treatment group’s deceased heart activity might indicate toxicity or physiological stress. Although the exact mechanisms are not fully understood, some researchers propose that increased heart rate may arise from the hypoxic conditions resulting from the adhesion of MPs to the chorion and MP uptake in the yolk.36,37 Inhibited heart rate is suspected to be linked to sarcomere disturbances. 37 However, further research is needed to more conclusively determine the implicated pathways involved.
These findings may provide additional insight into the effects MNPs may have on human health due to developmental exposure during critical and sensitive periods. A recent study located MPs inside the human placenta, 41 and similar results have since been replicated including one study conducted in Hawai‘i.22,42 MPs have been found to cross the maternal–fetal interface, raising concern that they may affect nutrient exchange during pregnancy.41,42 These studies align with the disruption in yolk usage seen in our results, underscoring the potential risks MNPs may have on early fetal development. Additionally, MNPs in human studies suggest that exposure may be linked to cardiovascular disease. Patients containing MNPs in plaque buildup were found to be at higher risk of heart attack, stroke, and death. 20 Previous in vivo studies have also found the effects of MNPs on heart rate to have been inconclusive, suggesting that symptoms may be dependent on size, dose concentration, length of exposure, and plastic type. 43
The observed increase in MNP concentration in zebrafish with longer exposure times could also be linked to the increase in developmental and behavioral results seen in subsequent analyses. Although a comprehensive understanding of the mechanism on a molecular level remains largely uncertain, several studies aimed at exploring the effects of exposure on growth and development have highlighted key aspects,24–26,44 which may explain the trends observed in this study. Whole-body transcriptomics uncovered significant changes in larval gene expression within 48 h of exposure to MPs. 44 These mechanisms may underlie the reduction in body length and other developmental disparities observed in the plastic-treated zebrafish groups in our study. Although transcriptional level changes at a critical time during larval development have been examined, the possibility of long-term effects warrants further exploration using cross-sectional and longer-term studies.
The absence of fluorescent NPs in 0–3 dpf larvae (no developed mouth) suggests that the mouth may be a major pathway in the intake and subsequent distribution of NPs, consistent with previously shown accumulation in the digestive track.23,24 Fluorescence indicative of NP presence was only observed in the 3–5 and 0–5 dpf treated fish, time periods in which a developed mouth was present. As shown in Figure 3, the highest concentration of NP was found in the intestine, further supporting oral ingestion as the primary entry point for NPs. High NP levels in the intestine may also potentially link to previously reported disruptions in the gut microbiome and gut–brain axis.45–47
Although there was no observed significant effect of treatment on locomotion (Fig. 4), all plastics-exposed groups showed slightly higher locomotion than control fish, which may demonstrate the ability of MNPs to cause changes in swimming. A past study found that adult zebrafish became hyperactive after MP exposure and observed a shortage in energy-supplying substances such as glucose after MP exposure. 48 PS-NPs also led to disruptions in glucose homeostasis that coincided with increased cortisol secretion and hyperactivity. 49 Thus, the slightly higher locomotion of the plastics-exposed groups may suggest disruptions in energy metabolism.
Interestingly, previously reported observation of lack of effect on survival and mortality rates23,25 was replicated in our work as well. Our findings also suggest that NPs may inhibit growth in zebrafish larvae, leading to a reduction in body length. This finding is supported by another study, which found NPs (100 nm) to reduce body length and decrease heart rate at a 200 and 400 mg/L dosage after 48 h in zebrafish embryos. 40 Together, these consistent effects further highlight the feasibility of using developing zebrafish for future research of plastic contamination and pollution, 34 as the contrasting sensitivity without decreasing survival allows optimal utility not possible in many other models.
Overall, our results suggest that the presence of MNPs in the early developmental environment may lead to changes in growth, behavior, and overall health of fish and other animals. Our work aligns well with existing literature, and together, these studies highlight that more thorough and long-term investigations must be done in both animal models and in humans to clarify underlying mechanisms and long-term consequences. MNPs represent a relatively new challenge to our understanding of environmental toxicities, means of exposure, and the risks of single-use plastics. As data from more studies emerge, it is essential that a comprehensive risk assessment is performed at the global level, including in geographically unique locations such as Hawai‘i, which are particularly at risk from MP pollution.
Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors’ Contributions
K.S.: Conceptualization, investigation, methodology, data collection, data curation, writing—original draft. T.H., L.I., C.L., K.M., Z.O., V.O., and G.Q.: Conceptualization, data curation, validation, writing—original draft. K.M.W.: Review and editing, project administration, funding acquisition. C.P.: Conceptualization, methodology, writing—original draft, writing—review and editing, supervision, project administration, funding acquisition. S.S.: Methodology, data collection, formal data analysis, validation, writing—original draft; writing—review and editing; Supervision, Project Administration. All authors reviewed the finalized manuscript and gave approval for publication.
Footnotes
Funding Information
This work was supported by a grant from the National Institutes of Health and support from the