
Abstract
The African turquoise killifish, Nothobranchius furzeri, has emerged as an important vertebrate model for aging research due to its naturally short lifespan and hallmarks of aging. However, these characteristics create challenges for colony management, such as high breeder turnover, rapid generational change, and the risk of inbreeding or unintended selection in closed laboratory populations. Since new individuals cannot be introduced from the wild or purchased from commercial laboratory animal breeders, maintaining genetically stable laboratory stocks requires carefully controlled breeding strategies. Here, we describe an innovative colony management approach termed dynamic population breeding (DPB) and evaluated its performance in two commonly used laboratory lines with different lifespans, GRZ-D and MZCS-08/122. DPB integrates several key principles: the maintenance of overlapping breeding age cohorts, harem-based breeding groups, continuous monitoring of clutch quality and quantity, and the controlled use of embryonic diapause for flexible embryo storage and synchronized hatching. Embryos from multiple breeding groups were pooled, stored in diapause stage II, and hatched at defined time points to generate new cohorts while avoiding sibling-only populations. Using this approach, we systematically analyzed reproductive performance, embryo quality, hatching success, and survival in laboratory populations. Fertilization rates and clutch sizes remained within stable ranges across the reproductive period, although an age-dependent decline in fertilization efficiency was observed in both lines. Quality control at the clutch level allowed early identification and exclusion of low-performing breeding groups. Importantly, implementation of DPB reduced variability between cohorts and improved early-life survival of offspring, particularly during the first weeks after hatching. Overall, DPB provides a practical framework for maintaining stable and robust killifish colonies while minimizing unintended selection and inbreeding. By integrating diapause biology with structured breeding management, this strategy enhances reproducibility and sustainability of N. furzeri populations used in aging research.
Introduction
The life cycle of the African turquoise killifish Nothobranchius furzeri (N. furzeri) is perfectly adapted to the transient nature of its habitat and consists of distinct phases. During the rainy season, the fish hatch and undergo rapid growth until they reach sexual maturity. 1 While mating, females lay eggs that are resistant to desiccation. During embryogenesis, development can enter three defined stages of diapause, 2 developmentally programmed arrests that enable long-term survival under harsh conditions. 3 These diapausing embryos endure the dry season protected within the substrate, resume development and hatch with the onset of the next rainy season. 4 For our research at the Leibniz Institute on Aging—Fritz Lipmann Institute (FLI), we maintain different lines, such as GRZ-D 5 and MZCS-08/122 6 that are distinguished by their different geographic origins, coloration patterns, and life spans. 1 Each of these lines carries unique genetic traits, which are invaluable for scientific studies. The maintenance of these stocks through captive reproduction is crucial, as no commercial providers are available. While their short lifespan and accelerated aging is advantageous for scientific research,7–10 it creates significant challenges for husbandry, requiring an increased level of care and precision from facility managers and staff. The rapid life cycle: hatching, maturation, reproduction, and senescence occurs in a matter of weeks or few months 11 and requires much more intensive colony management 12 compared with longer-lived species, like zebrafish. This necessitates frequent monitoring and immediate responses to changes in environment and health, 13 as even small deviations in temperature, 14 or food intake15–18 can have an outsized impact on their lifespan and reproduction. The high turnover rate of breeding animals further complicates facility operations. Since short-lived species quickly transit from juveniles to reproductive adults, space management must be dynamic, with tanks and equipment rapidly reassigned between cohorts. To ensure that these fish receive appropriate care throughout all life stages, specialized training for facility staff and continuous refinement of husbandry protocols is required.19–21 Suboptimal conditions can lead to reduced oocyte production and higher rates of morbidity and mortality. Additionally, a key challenge lies in preserving the genetic profile of the fish lines while avoiding both selective pressures22,23 and inbreeding depression.24–26 Both outcomes would compromise the comparability of data across generations. The use of breeding programs reduces the need for excessive animal numbers and maintains healthier, more robust populations27,28 and researchers can generate consistent data while upholding the highest standards of animal care. To address this, we developed the innovative breeding strategy of dynamic population breeding (DPB) that takes advantage of diapause, timed hatching, and the combination of breeding groups from different breeding generations.
Materials and Methods
Housing of N. furzeri
All fish were maintained at the FLI according to German Animal Welfare Law under license §11–003798, approved by the local authorities (Zweckverband Veterinar- und Lebensmittelüberwachungsamt Jena-Saale-Holzland). Husbandry conditions were the following: Water temperature 26°C, conductivity 0.5–0.8 mS, pH ∼7.3, photoperiod 12 h light/12 h dark including dawn and dusk simulation, LED lighting. Fish were fed from hatching with Artemia nauplii (Ocean Nutrition, Essen, Belgium) twice daily and weaned to live bloodworms (Chironomidae) at 4–6 weeks post hatching (wph) twice a day. Health monitoring protocols are detailed in 13 . All group-housed fish were supplied with certified contaminant-screened blue polycarbonate igloos, retreats, huts, and tents (Bio-Serv, Flemington) as enrichment.
Selection of breeders and breeding of N. furzeri
N. furzeri were group-housed in recirculating fish racks (Aqua Schwarz, Göttingen, Germany) in 10 L tanks at ∼1 fish/L (max. 10 fish per tank). At 3–4 weeks post-hatching, males developed coloration and were separated from females to allow females to reach a size suitable for oocyte production. At 8–10 wph, harem-based matings (1 male and up to 4 females) were established in 10 L tanks. Only healthy fish with normal behavior, intact fins, and no signs of disease or deformities (e.g., operculum malformations) were used as breeders. 13 Group housing continued throughout the reproductive period to support continuous spawning. Spawning occurred in small (12 cm × 8 cm) dark blue pipette boxes filled with fine white sand. Boxes were placed in tanks for 4 days, followed by a 3-day resting period; this cycle was repeated throughout the breeding period. Breeding groups showing persistently low performance in egg production were dissolved, whereas in consistently low fertilization despite adequate egg production, males were replaced by new ones, when possible.
Dynamic population breeding
Before 2022, no standardized breeding strategy was in place at the FLI. In beginning of 2022, we developed the structured breeding strategy of DPB (Fig. 1): One breeding population consists of three distinct age cohorts, and each age cohort includes up to five harem-based breeding groups (Fig. 1A). Accordingly, each breeding population comprises up to 75 fish, with 15 males and 60 females. Each age cohort has a defined breeding period starting at 8–10 wph, corresponding to reaching sexual maturity and breeding age, and continues until the average line specific survival rate drops to around 60%. Embryos produced by the current breeding population are pooled and stored in the diapause II stage (DII, somite stage) (Fig. 1B).
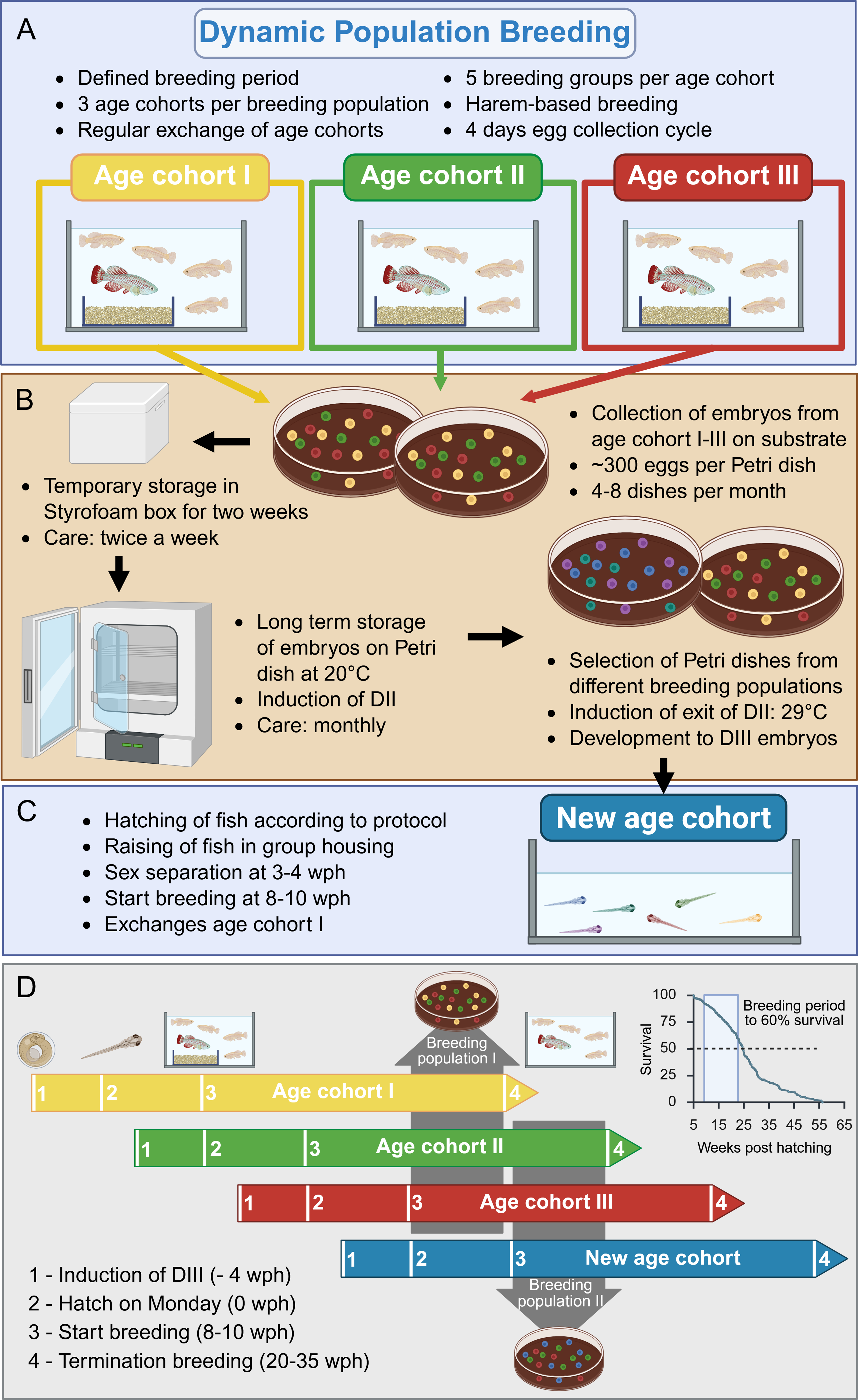
Dynamic population breeding scheme.
To maintain dynamic continuous breeding, a new age cohort is hatched in time (Fig. 1C). Each age cohort consists of fish hatched on the same day, but according to the breeding strategy, these fish are unlikely to be siblings, since each cohort receives contributions from multiple previous breeding groups. The oldest age cohort is replaced, when breeding maturity is reached in the new age cohort, marking the start of the new breeding population (Fig. 1D).
Embryo collection and storage
Clutches from all breeding groups were collected on the same day. Sand boxes were removed from tanks, and each clutch was separated from the sand using a sieve. After quality assessment, clutches were pooled, and viable embryos were transferred to Petri dishes containing slightly moistened, not autoclaved coconut substrate (Dragon Coco-Ground, Zoo Zajac, Duisburg, Germany) either manually or using the EggSorter (Bionomous, Villaz-Saint-Pierre, Swizerland). Each month, 4–8 Petri dishes with ∼300 embryos per dish were collected and labeled with the month and year of collection.
To avoid repeated disturbance of long-term stored embryos and to induce DII (Fig. 3C), we kept Petri dishes with newly collected embryos at room temperature (20°C) in a Styrofoam box for 2 weeks and inspected them twice a week to remove dead embryos. They were then transferred to incubators (20°C, Memmert IPP30, Bremen, Germany) to maintain DII. During storage, DII embryos were checked monthly, the substrate was moistened at the edge of the Petri dish using a pipette with autoclaved water. Embryos exiting DII or dead ones were removed. Under these conditions, embryos could be stored for 1–2 years.
Hatching process
Petri dishes with embryos in DII from different collection time points were retrieved from the incubator, and dead embryos and those that had exited DII were removed. To induce further development, Petri dishes were transferred to a 29°C incubator (Fig. 1B). Embryos completed development within 2–4 weeks and reached diapause III (DIII, ready-to-hatch stage; Fig. 3C).
Hatching was performed on Mondays. Up to 100 embryos were placed in a hatching box (12 cm x 8 cm) with 50 mL of autoclaved, cooled (8–10°C) water, supplemented with a few crumbs of oxygen powder (JBL OxyTabs, JBL, Neuhofen, Germany) and freshly hatched Artemia nauplii (SEPART Artemia Cysts, Ocean Nutrition, Essen, Belgium) under additional light. The water level did not exceed 1 cm and was renewed twice a day. Hatching occurred within minutes to several hours and was terminated after 30 h; remaining embryos were discarded.
Up to 25 hatchlings were transferred to fresh hatching boxes with 50 mL room-temperature supplemented water. Hatchlings were fed with freshly hatched Artemia nauplii twice daily and transferred to recirculating racks at 4–5 days post-hatching (dph). Feeding success was indicated by an orange abdomen. During the first 4 days, water was exchanged twice daily to maintain stable quality. Water levels in tanks were increased slowly.
Monitoring of clutch quality and clutch quantity
Reproductive performance was monitored by collecting clutches from each harem-based breeding group every 4 days and transferring them to Petri dishes. Embryo quality was assessed under a stereomicroscope based on morphology: viable embryos showed intact chorion, turgid yolk, visible lipid droplets, and a defined perivitelline space, whereas nonviable embryos or unfertilized oocytes displayed early structural degradation such as a ruptured or collapsed chorion, increased opacity, and a collapsed or disintegrating yolk. Clutch size from each breeding group was scored semi-quantitatively: 1 = low, 2 = normal, 3 = high, to standardize spawning assessment and detect impairments. Exact oocyte number per female per day were determined from photographs of individual clutches. During storage, dishes with embryos were evaluated weekly under the stereomicroscope to assess embryo number, quality, and developmental stage. Dead embryos and those reaching DIII were removed, and embryo counts were documented based on images.
Survival and survival curve analysis
Survival curve analysis was performed using the lines GRZ-D and MZCS-08/122 that hatched between 2020 and 2024. The analysis included group-housed animals that were designated as broodstock or used for organ collection. Fish that were euthanized upon reaching a predetermined humane endpoint, 13 as well as those that died peracutely, were included in the survival analysis. Fish that were euthanized for other purposes were considered censored cases. The Kaplan–Meier survival functions were estimated using GraphPad Prism (Dotmatics, Boston, MA) and were plotted from 5 wph. All animals were tracked in Tick@lab (a-tune, Darmstadt, Germany) from hatching on and survival data for 4 dph and 5 wph were calculated for corresponding hatches between 2020 and 2024.
Hatching test
Embryos of the MZCS-08/122 line were put to 29°C for 3 weeks until they reached DIII-stage. A total of 300 embryos were collected, and each week for 10 weeks, 10 embryos were selected for hatching. Fish that hatched within 30 h were defined as successfully hatched. Embryos not hatched within 30 h were classified as “not hatched”. Survival of hatchlings was monitored for 30 days.
Results
At the FLI, multiple N. furzeri lines are maintained to address aging-related research questions. Here, we systematically analyzed breeding performance, clutch size, and lifespan for two commonly used lines, the short-lived GRZ-D and the longer-lived MZCS-08/122. For both lines, breeding groups were established between 8 and 10 wph. To minimize unintentional selection for increased lifespan, mating was discontinued once average line specific survival declined to approximately 60%, corresponding to ∼20 wph for GRZ-D and ∼35 wph for MZCS-08/122. This strategy ensured that breeding occurred within a defined and comparable life phase for each line. Following the establishment of breeding groups according to the DPB scheme, we assessed each clutch using a combination of quantitative and qualitative parameters. For quantitative evaluation, clutch quality was determined by calculating the proportion of viable embryos at the day of collection, based on counts of viable embryos and nonviable embryos or unfertilized oocytes (Fig. 2D, indicated by arrows). To maintain stable clutch quality, we defined a fertilization rate threshold of 40%, and clutches falling below this threshold were classified as low quality and excluded from further processing. In the GRZ-D line, average fertilization rates ranged from 47% to 72% across the breeding period from 8 to 20 wph (Fig. 2A), resulting in an overall fertilization rate of 62.5%. Notably, we observed a decline in fertilization success after 14 wph. Fertilization rates decreasing from approximately 65% before 14 wph to 57% thereafter, indicating an age-dependent decline in fertilization efficiency. In MZCS-08/122, average fertilization rates varied between 36% and 91% over a broader breeding window from 9 to 35 wph (Fig. 2B). The overall fertilization rate across the entire breeding period was 71%. Similar to GRZ-D, we detected an age-associated decline in clutch quality, with fertilization rates dropping from approximately 80% to 56% at around 26 wph, suggesting that reduced fertilization efficiency emerges at later ages in this longer-lived line.
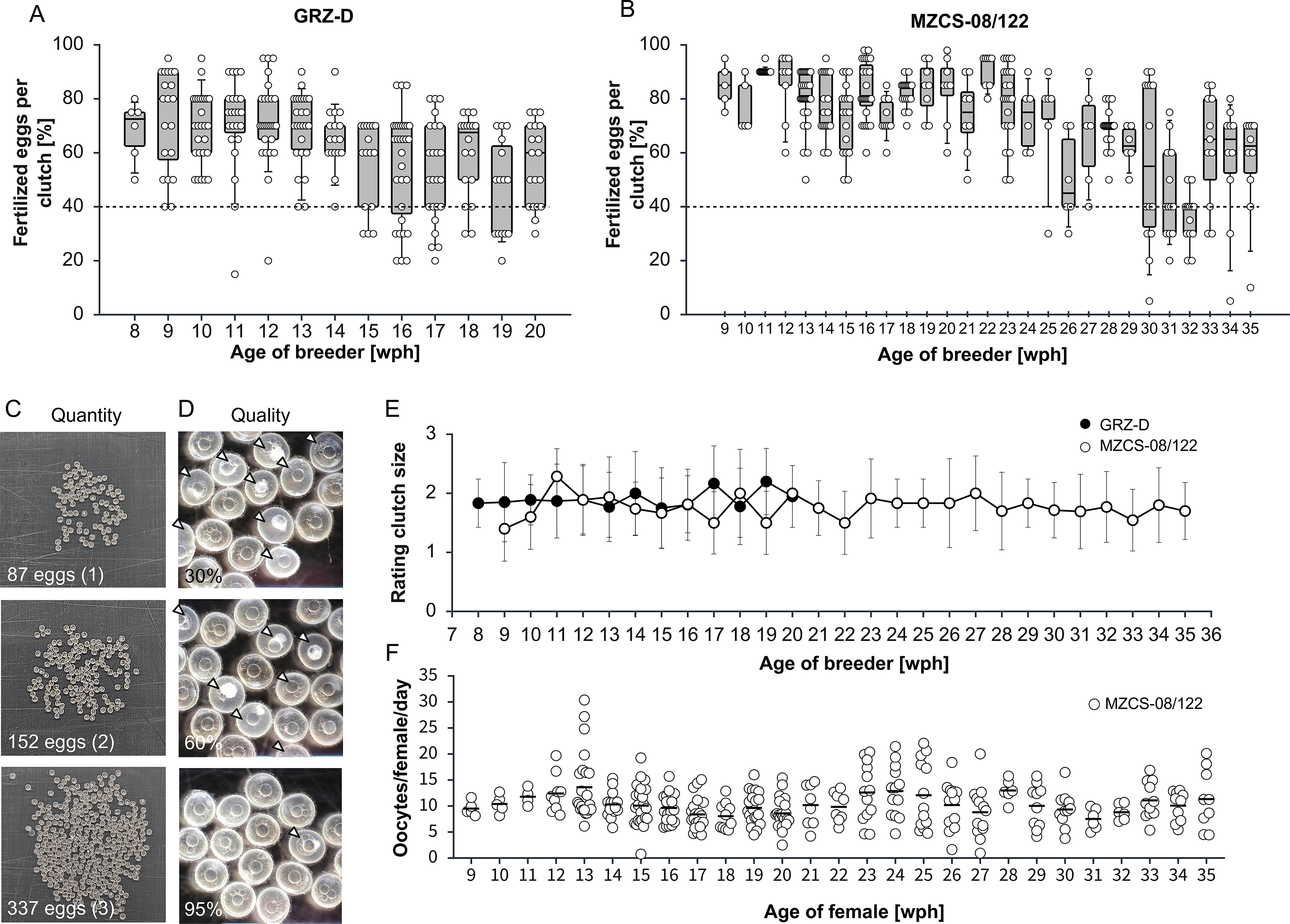
Breeder performance
To assess clutch quantity, we applied a semi-quantitative scoring system. Analyzing clutches of breeding groups, 130–250 eggs were considered normal and assigned a score of “2,” clutches with fewer eggs were rated as “1,” and clutches exceeding the average egg number were rated as “3.” Representative examples of these clutch size categories are shown in Figure 2C. Clutch size was recorded throughout the entire reproductive phase for GRZ-D and MZCS-08/122 breeding groups (Fig. 2E). In GRZ-D, mean clutch ratings ranged from 1.8 to 2.5 across the breeding period, whereas MZCS-08/122 showed slightly lower values, ranging from 1.4 to 2.3. Breeding groups that exhibited a low fertilization rate or reduced clutch size over 3 consecutive weeks were excluded from further breeding. This on-site evaluation of clutch quality and quantity enabled effective assessment of reproductive performance, allowing identification and replacement of poorly performing breeding animals the group.
A more detailed analysis of reproductive fitness was performed for MZCS-08/122 by determining oocyte production of females in relation to age (Fig. 2F). Clutches were collected from 4 days and normalized by the number of females per breeding group to calculate mean oocytes per female per day. Across the breeding period, average production ranged from 9 to 14 oocytes per female per day. In individual cases, peaks were observed, with values reaching 20–25 oocytes per female per day. Assessing the quantity and quality of clutches provides a quantitative basis for estimating embryo output and for calculating the number of breeding groups required to achieve defined embryo numbers.
Clutches collected on the same sampling date from different age cohorts that passed the predefined quality criteria were pooled and transferred onto coconut fiber substrate. To minimize secondary loss due to microbial contamination, we removed nonviable embryos (Fig. 3A, left image). If plates were not regularly cleaned, fungal growth would spread rapidly and affect all embryos on the plate, leading to complete loss of the clutch (Fig. 3A, right image). During the first weeks after collection, nonviable embryos were consistently observed on the plates. The highest incidence occurred within the first week post-collection, amounting to 7% (SD = 2.6%) for MZCS-08/122 and 12.5% (SD = 6.8%) for GRZ-D, indicating that early post-collection mortality represents the major source of embryo loss. After 4 weeks post collection, all remaining embryos had entered DII. Embryos in DII were characterized by a segmented body with clearly defined somites, which were only faintly visible on the coconut substrate, and by the presence of a prominent, spherical lipid droplet within the yolk (Fig. 3D). After 4 weeks post collection embryo numbers remained stable and mortality rates were consistently very low (<0.2%). Plates were subsequently monitored monthly. After approximately 5 months of storage, we observed first embryos exiting DII, indicated by the onset of eye pigmentation (black eye stage), and this fraction increased rapidly (Fig. 3C). Between 7 and 8 months of storage, approximately 60% in MZCS-08/122 and 10% in GRZ-D of embryos had reached DIII stage. Embryos in DIII were readily identifiable by large, golden eyes and a distinct bright ring surrounding the pupil (Fig. 3D).
To investigate hatch success after entering DIII, embryos in DII were incubated at 29°C to reach DIII synchronously and weekly hatching assays were performed over a 10-week period. Following initiation of the hatching protocol, embryos were monitored and classified as hatching within 12 h or within 30 h (Fig. 3E). During the first week after reaching DIII, not all embryos successfully hatched. The highest hatching rates were observed between 2 and 4 weeks after entry into DIII. From week 5 after entering DIII onwards, hatching increasingly occurred within 30 h or failed. When analyzed across the entire 10-week period, we did not detect a statistically significant difference in hatching timing or overall hatching success (p > 0.05, ANOVA). In contrast, analysis of post-hatching survival revealed a clear temporal effect. Fish hatched within 1–5 weeks after reaching DIII showed significantly higher survival during the first 30 dph compared with fish hatched between weeks 6 and 10 after reaching DIII (p = 0.0178; Fig. 3F), suggesting that prolonged maintenance in DIII negatively affects early post-hatching viability rather than hatching efficiency.

Embryo care and survival.
To evaluate the robustness of subsequent generations, early juvenile survival was assessed at 4 dph and 5 wph for GRZ-D and MZCS-08/122 hatches generated in the years before and after the establishment of DPB. For GRZ-D, prior to DPB implementation, survival at 4 dph was highly variable across individual hatches (Fig. 4A), with mean survival rates ranging from 90.4% (SD = 16.6%) to 75.7% (SD = 25.4%). Following the establishment of DPB, median survival at 4 dph increased to 93.1% 3 years later, accompanied by a marked reduction in variability with SD = 10.8%. At 5 wph, survival rates before DPB implementation were low, averaging 60.2% (SD = 25.5%) and 53.4% (SD = 27.4%) across two pre-DPB years (Fig. 4C). After implementation of DPB, mean survival at 5 wph increased to 78.8% 3 years later, with substantially reduced variability of SD = 11.9%. These results indicate that DPB and controlled synchronization of development through regulated DII exit stabilizes early juvenile survival in the shorter-lived GRZ-D line.
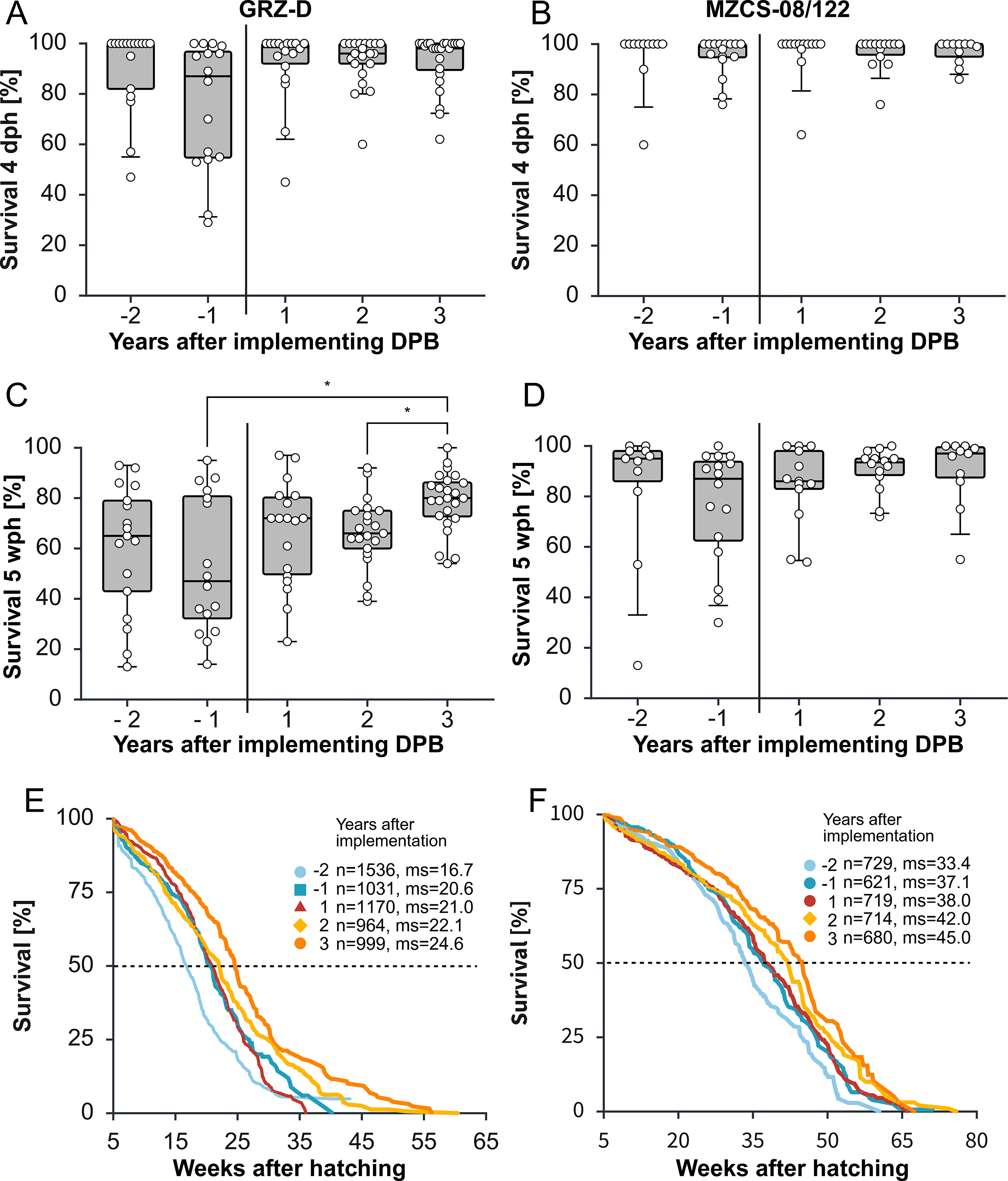
Early hatching survival and life span analysis.
The longer-lived MZCS-08/122 line exhibited consistently high survival at 4 dph, with rates ranging from 95.0% (SD = 7.7%) to 96.8% (SD = 5.0%) throughout the observation period (Fig. 4B). In contrast, survival at 5 wph showed greater variability (Fig. 4D). In the 2 years preceding DPB implementation, mean survival was 83.4% (SD = 27.0%) and 76.3% (SD = 22.8%). Three years after DPB implementation, mean survival increased to 90.4% (SD = 14.1%). Although this increase was not statistically significant, the lowest survival rates observed within each year improved notably, suggesting enhanced robustness of juveniles during the early growth phase in subsequent generations.
The Kaplan–Meier survival analysis demonstrated an increase in early-life survival following the implementation of DPB in both the GRZ-D and MZCS-08/122 lines (Fig. 4E and F). In GRZ-D, mean survival increased progressively from 16.7 weeks 2 years before DPB to 24.6 weeks 3 years after its introduction. A similar pattern was observed in MZCS-08/122, where mean survival rose from 33.4 weeks 2 years before implementation of DPB to 45.0 weeks 3 years post implementing DPB. Notably, these increases were gradual and consistent across consecutive cohorts, suggesting that the improvements were not attributable to year-to-year environmental variation.
Overall, DPB reduced inter-cohort variability and improved early-life survival across subsequent generations. At the same time, the strategy enabled controlled breeding within defined life phases while minimizing unintended selection pressures.
Discussion
Maintaining genetic consistency across generations is a central challenge in the long-term management of N. furzeri laboratory colonies. Due to the lack of commercial providers and the species’ short generation time, laboratory colonies are particularly vulnerable to genetic drift, inbreeding, and unintentional selection.
A strategy to preserve genetic consistency across generations is the maintenance of multiple parallel breeding lines derived from the same ancestral population. This approach reduces the impact of genetic drift and stochastic breeding failures by distributing risk across independent sub-colonies.29,30 Parallel lines require a substantial increase in tank numbers, and associated infrastructure, which rapidly exceeds available housing capacity in facilities primarily designed to support experimental work. Despite its genetic advantages, implementation of the parallel-line model is limited by the spatial constraints of routine laboratory colony management.
Breeding schemes in N. furzeri husbandry usually rely on single, large-scale hatching events. Such approaches require the simultaneous availability of large numbers of ready-to-hatch embryos and impose a narrow temporal window in which sufficient breeders must be obtained. If the hatch fails, the entire breeding cycle may be compromised. Smaller, planned hatchings distributed over time provide enhanced resilience and flexibility. By maintaining multiple, partially overlapping cohorts, breeding is not dependent on the success of a single hatch. From a genetic standpoint, spreading the breeding effort over time helps conserve a more diverse gene pool, as selection is not constrained to a single group of animals.
The timing of breeding termination represents another critical parameter in preserving population viability and avoiding unintended selective pressures. In our system, mating was stopped at 60% survival to balance reproductive output with genetic stability. Extending breeding beyond this point could increase the likelihood of preferentially propagating long-lived individuals or other correlated phenotypes, potentially shifting trait distributions in subsequent generations. This consideration is particularly important in short-lived species such as N. furzeri, where lifespan variation is tightly linked to reproductive timing and fitness. Future investigations could assess how adjustments to this survival threshold influence population dynamics and trait distributions, thereby refining breeding protocols further.
Male turquoise killifish exhibit an age-dependent decline in sperm quality, fertilization success, and embryo survival, indicating reproductive senescence even in this naturally short-lived model. 31 Broader reviews confirm that reproductive senescence is widespread across ray-finned fishes, with declines in key reproductive traits linked to chronological age. 32 Reproductive decline with age does not constitute an absolute barrier to effective breeding. Our data reveal that although fertilization success declines with breeder age, fertile individuals remain usable when combined with structured quality assessments. Systematic exclusion of clutches that failed early quality criteria effectively compensated for age-related declines, allowing prolonged use of breeders without compromising reproducibility as previously reported.33,34
Embryonic diapause in N. furzeri represents a natural adaptation that decouples reproduction from immediate hatching and provides important advantages for colony management. By controlling diapause exit under defined incubation conditions, embryo development and hatching can be synchronized without compromising survival. Embryos should be handled as little as possible, since repeated light exposure during inspections may stimulate development. 19 Integrating diapause biology into breeding protocols not only reflects the natural ecology of killifish 35 but also enhances resource efficiency in long-term colony management.
In N. furzeri, survival during the first post-hatching stages reflects the cumulative outcome of processes occurring both before and after fertilization, including embryogenesis, developmental competence, and the ability of hatchlings to withstand handling and environmental stressors. Our application of DPB reduced variability in survival between hatches and increased average survival at key developmental milestones. This improvement is likely linked to more consistent clutch quality assessment and the structured use of overlapping cohorts, which also reduces the likelihood of sibling-based breeding within a cohort. The data indicate that DPB is an effective management strategy to enhances survival by stabilizing reproductive output.
Despite its advantages, DPB also has limitations. Managing overlapping cohorts requires dedicated planning, infrastructure, and sustained labor investment. Regular clutch assessment and embryo handling demand technical expertise, and inconsistent handling may introduce inadvertent selection biases. Moreover, while DPB reduces variability, it cannot eliminate intrinsic biological variation arising from maternal, 36 paternal 37 and environmental effects 38 on embryo development. The comparison of survival before and after DPB implementation is observational across calendar years, during which staff experience, protocol refinements, and other unmeasured improvements could have co-varied with DPB. Although the consistent improvement across cohorts argues against random year-to-year variation, a contribution from concurrent husbandry improvements cannot be excluded.
In summary, DPB provides a structured framework for the long-term management of N. furzeri colonies by combining controlled breeding cohorts with diapause-based embryo storage. This approach supports stable reproductive output while minimizing variability between generations. By integrating quality control at the clutch level and synchronized cohort replacement, DPB improves colony consistency and reproducibility. Complementing DPB, cryopreservation 39 offers additional advantages for long-term colony management. It ensures genetic stability over multiple generations, reduces husbandry demands by eliminating the need for continuous maintenance, and provides a reliable backup against colony loss. Together, these strategies create a synergistic framework that balances short-term population management with long-term genetic security, supporting reproducible and sustainable N. furzeri research.
Authors’ Contributions
B.H.: Conceptualization, methodology, formal analysis, investigation, writing—original draft preparation; U.N.: Conceptualization, methodology, formal analysis, investigation, writing—original draft preparation; J.H.: Methodology, formal analysis, writing—review and editing; C.P.: Methodology, formal analysis, writing—review and editing, S.G.: Methodology, formal analysis, writing—review and editing; M.N.: Methodology, writing—review and editing.
Ethical Considerations
All animal experiments were performed in accordance with the German Animal Welfare Act (TierSchG) and relevant national regulations. The Housing and Husbandry of N. furzeri was approved by the competent regional authority Zweckverband Veterinar- und Lebensmittelüberwachungsamt Jena-Saale-Holzland, Germany (approval no. §11–003798) on August 29, 2017.
Footnotes
Acknowledgments
The authors thank the animal technicians of the Animal Facility Fish at the Leibniz Institute on Aging—Fritz Lipmann Institute Jena for their passionate work in fish caring every day. The authors thank Anne Süßenguth for her work as veterinarian, caring about the health of our fish.
Disclosure Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
The FLI is a member of the Leibniz Association and is financially supported by the Federal Government of Germany and the State of Thuringia.