
Abstract
Amelogenin has been widely used in clinical practice for periodontal bone regeneration. However, the precise mechanism underlying its osteogenic effects remains incompletely understood. In this study, we hypothesized that amelogenin enhances periodontal bone regeneration by facilitating the migration and homing of bone marrow mesenchymal stem cells (BMMSCs). BMMSCs were used to evaluate the cell migration promoting ability of amelogenin by the Transwell test. Immunofluorescence was performed to assess the beta-catenin nuclear translocation following amelogenin treatment. To investigate amelogenin-induced cell homing in vivo, we established a green fluorescent protein (GFP)-labeled bone marrow transplantation model using BALB/c mice transgenic for GFP. The migratory effects of amelogenin were examined in this model, with Wnt3a, a Wnt/β-catenin pathway activator, serving as a positive control. Subsequently, cell homing and bone regeneration were evaluated through a fluorescence microscope, micro-CT, hematoxylin and eosin (H&E), and Masson staining. In vitro Transwell assays demonstrated that amelogenin significantly enhanced BMMSC migration, with effects comparable with Wnt3a, a canonical Wnt/β-catenin pathway activator. Immunofluorescence analysis revealed pronounced nuclear translocation of β-catenin in BMMSCs following a 24-h amelogenin treatment. Notably, these effects were abolished by a Wnt/β-catenin pathway inhibitor, confirming the pathway’s involvement. In GFP-labeled bone marrow-transplanted mice, amelogenin treatment significantly increased GFP+ cell recruitment to the bone defect site, mirroring the effects of Wnt3a. Micro-CT and histological (H&E) analyses further demonstrated that both amelogenin and Wnt3a accelerated bone regeneration compared with untreated controls. Crucially, this regenerative effect was suppressed upon Wnt/β-catenin pathway inhibition, reinforcing the mechanistic link between amelogenin and β-catenin-mediated osteogenesis. Amelogenin and Wnt3a promoted periodontal bone regeneration both in vitro and in vivo by enhancing BMMSC migration through Wnt/β-catenin signaling activation.
Introduction
Periodontal diseases represent a global health challenge, affecting up to 90% of the worldwide population. 1 These conditions lead to progressive alveolar bone resorption and connective tissue destruction, ultimately becoming the primary cause of tooth loss in adults. Despite significant advances in dental medicine, complete regeneration of lost periodontal tissues remains an unmet clinical need. Current therapeutic strategies for periodontal regeneration include guided bone regeneration surgery, 2 bone graft, 3 and stem cell treatment. 4 Among these approaches, stem cell treatment has attracted considerable attention due to its potential to regenerate multiple tissue components and restore the complex architecture of periodontal structures. 5 However, the scarcity of endogenous periodontal stem cells limits their regenerative capacity, highlighting the need for alternative approaches that enhance host cell recruitment. Conventional stem cell transplantation faces several challenges, including limited cell sources, ethical concerns, and iatrogenic damage to donor sites. 6 In response to these limitations, cytokine-mediated stem cell homing has emerged as a promising alternative.
Enamel matrix derivative (EMD) has demonstrated significant clinical efficacy in periodontal regeneration therapy, with numerous studies documenting its osteogenic properties. 7 The therapeutic effects of EMD are mediated through multiple mechanisms: upregulation of osteoprotegerin mRNA expression, suppression of osteoclast differentiation and activity, and enhancement of alkaline phosphatase activity. 8 Furthermore, EMD stimulates transforming growth factor-beta1 (TGF-β1) production in osteoblasts, thereby facilitating bone formation. 9 Notably, amelogenins constitute over 95% of EMD’s protein content, with purified amelogenins recapitulating the biological effects of complete EMD. 10 Amelogenin plays a crucial role in the formation of tooth enamel. 11 Previous study has shown that amelogenin is the main effective component of EMD in periodontal regeneration. 12 For that reason, it is important to have a more profound understanding of the mechanism of amelogenin-induced periodontal bone regeneration. Research on amelogenin in craniofacial regeneration dates back to the 1990s. 13 Early investigations primarily centered on its capacity to induce cementum formation. 14,15 Viswanathan et al. 16 demonstrated via in vitro studies that amelogenin regulates the expression of cementum-associated genes (e.g., Bone Sialoprotein (BSP), type I collagen) in a dose-dependent manner, thereby establishing a foundation for subsequent investigations into craniofacial tissue regeneration. These findings provided the theoretical framework for this study.
Current research has expanded our understanding of amelogenin’s regenerative potential, revealing its ability to modulate osteogenic responses, promote cementoblast differentiation in periodontal defects, and facilitate periodontal fiber reattachment. 17 However, the precise molecular mechanisms underlying amelogenin-induced bone regeneration remain incompletely characterized. Unlike the mechanism of Wnt3a agonists, amelogenin binds to LRP5/6 receptors through its LRAP fragment, stabilizes β-catenin, and promotes its nuclear translocation, thereby activating TCF/LEF transcription factors. This process synergizes with the BMP-2 pathway, increasing Smad1/5/8 phosphorylation levels by 1.8-fold. 18 In this study, we attempt to observe the effect of amelogenin inducing bone marrow mesenchymal stem cells (BMMSCs) migration and elucidate the involvement of the Wnt/β-catenin pathway in this process.
The Wnt protein family comprises evolutionarily conserved secreted glycoproteins that play essential roles throughout development—from early zygotic cleavage to adult tissue homeostasis. 19 These proteins regulate fundamental cellular processes, including proliferation, differentiation, and migration, through highly conserved signaling pathways. 20 Emerging evidence suggests that amelogenin modulates cell behavior via Wnt signaling. Amelogenin has been demonstrated to enhance Periodontal Ligament (PDL) migration in vitro through Wnt pathway activation. 21 Further study showed that beta-catenin would be highly phosphorylated when human mesenchymal stem cells were incubated with amelogenin. 22 Based on these findings, we hypothesized that amelogenin could induce BMMSC migration and proliferation by triggering the Wnt/β-catenin pathway during the periodontal regeneration progress. This study aims to establish amelogenin-induced BMMSC migration as a key mechanism in periodontal regeneration and characterize Wnt/β-catenin pathway activation during this process. Wnt3a serves as a pathway-specific positive control to validate Wnt/β-catenin involvement and provide comparative signaling benchmarks. This design does not imply functional equivalence between Wnt3a and amelogenin, which operates through distinct mechanisms.
Materials and Methods
Cell culture and identification
All animal experiments were approved by the Animal Care and Experimentation Committee of Xi’an Jiaotong University (Xi’an, China). C57BL/6N mice (4–6 weeks, 20 g, male or female) were obtained from Xi’an Jiaotong University Laboratorial Animal Center. The primary BMMSCs were isolated from C57 mice according to previously published procedures. 23 Briefly, muscle and other soft tissue were carefully removed from C57 mice femurs and tibiae to prevent myogenic precursor contamination. Both ends of the bones were cut off with bone scissors. The bone marrow plugs were hydrostatically expelled from the bones by insertion of 2 mL syringes filled with complete Dulbecco’s Modified Eagle’s Medium (DMEM). Then the bone marrow cells were centrifuged and resuspended into a 10 cm Petri dish with 10 mL growth medium at 37°C in a humidified 5% CO2 incubator. Three days later, the medium was changed and unattached cells were discarded. Then the medium was changed every other day. Passages three to five were used in the following experiments. Flow cytometry was performed on passage three cells to examine the purification of BMMSCs. CD29 and CD90 and hematopoietic cell markers CD34 and CD45 24 were analyzed during FACS analysis.
Differentiation ability was tested with adipogenic, osteogenic, and cartilage differentiation medium on passage three BMMSCs. For adipogenic differentiation, BMMSCs were seeded at a density of 5 × 104 cells/mL in 6-well plates cultured with adipogenic medium (DMEM, 10% Fetal Bovine Serum (FBS), 1 µmol/L dexamethasone, 0.5 mmol/L methyl isobutyl xanthine, and 10 µg/mL insulin; Sigma, Saint Louis, Missouri, USA), which was changed for every 3 days. Adipogenic differentiation was observed 2 weeks later with Oil Red O staining (Sigma, Saint Louis, Missouri, USA).
The same method was used to induce osteogenic and cartilage differentiation using osteogenic medium (0.01 µmol/L dexamethasone, 10 mmol/L glycerophosphate, and 50 mmol/L L-ascorbic acid-2-phosphate, Sigma, Saint Louis, Missouri, USA) and cartilage differentiation medium (0.1 µmol/L dexamethasone, 1mmmol/L sodium pyruvate, 50 mmol/L 2-phosphate ascorbic acid, and 1% ITS + 2, Sigma, Saint Louis, Missouri, USA), respectively. In the osteogenic differentiation test, mineralized hydroxyapatite extracellular matrix was observed 3 weeks after incubation stained by 40 mmol/L Alizarin Red S (Sigma, Saint Louis, Missouri, USA). The cartilage differentiation was observed by 0.5% Toluidine Blue O staining (Sigma, Saint Louis, Missouri, USA).
Transwell migration assay
Twenty-four-well cell culture plates and 6.5 mm COL collagen-coated Transwell Insert (8 µm Pore size, Corning, New York, USA) were used in Transwell migration assays. BMMSCs were trypsinized and resuspended at a density of 5 × 104 cells/mL in serum-free DMEM and transferred to the upper chamber. The lower chambers were filled with different concentrations of Wnt3a (100 ng/mL; PeproTech, Rocky Hill, USA), amelogenin (100 ng/mL, Fitzgerald, Acton, Massachusetts, USA), or amelogenin with 100 ng/mL Wnt pathway inhibitor DKK-1(PeproTech, Rocky Hill, USA). The chambers were removed from plates after a 24-h incubation. The remaining cells on the upper surface of the filter were wiped away with cotton swabs. The lower surface of the Transwell chambers was stained with 0.1% hematoxylin for 30 min. Cell counting was performed in six random high-power fields (×200). The average number of cell counting stands for the cell migration ability of different media. All treatments were assessed in triplicate. Wnt3a (100 ng/mL) served solely as a positive control for Wnt/β-catenin pathway activation.
Immunofluorescence analysis
Immunofluorescence was used to assay the activation of Wnt/β-catenin pathway triggered by amelogenin. Beta-catenin protein expression was upregulated and transported into the nucleus, which represents the activation of Wnt/β-catenin pathway. Passage three BMMSCs were treated with amelogenin (100 ng/mL) or amelogenin with DKK-1, respectively, for 24 h. Then, cells were fixed in 4% paraformaldehyde for 10 min at room temperature and blocked with 3% BSA in PBS for 30 min. After that, cells were immersed in a primary antibody (diluted at 1:50, beta-catenin, Cell Signaling Technology, Danvers, Massachusetts, USA) and incubated at 4°C overnight. After three washes in PBS, a secondary antibody was added (1:200 dilution, CY3-conjugated goat anti-rabbit, Santa Cruz, California, USA) and incubated for 1 h at room temperature. The cell nucleus was counterstained with 4’,6-diamidino-2-phenylindole (DAPI) at 1:5,000 dilutions in PBS. The result was observed with a fluorescence microscope (DMi8, Lecia, German).
Green fluorescent protein BMMSC transplantation mouse model
Previous research has shown that BMMSCs have the potential to migrate to bone wound sites and participate in bone regeneration. 25 To prove that BMMSCs were recruited by amelogenin through the Wnt/β-catenin pathway in periodontal bone regeneration, a green fluorescent protein (GFP) bone marrow transplantation C57 mouse model was created. The radiation and transplant procedures followed previously published research. 26 Briefly, the bone marrow cells were extracted from GFP transgenic BALB/c mice. The cell suspension was filtrated to remove large bone marrow tissue to prevent blood clots. Then, the GFP bone marrow cells were centrifuged and resuspended in natural saline. Forty C57 mice were irradiated by the 60 Co Gammacell 1000 Irradiator (Atomic Energy, Canada) with a sublethal dosage of 9.5 Gy for total body irradiation. Finally, the cell suspension containing 2 × 106 cells was transplanted into irradiated C57 mice through tail vein injection to create the GFP bone marrow transplantation C57 mouse model. The donor GFP BALB/c mice were purchased from Nanjing Biomedical Research Institute of Nanjing University. The recipients were 6-week-old C57 mice. Flow cytometry was used to examine the transplant result 4 weeks after GFP bone marrow cell transplantation. Tail vein blood, 0.5 mL, was taken from recipients. Red blood cells were removed using 2 mL Red Blood Cell Lysis Buffer (Solarbio, Beijing). After centrifugation, the supernatant was discarded, and the pellet was washed twice with PBS before being resuspended for flow cytometry analysis to determine the proportion of green fluorescent cells.
Fabrication of amelogenin hydrogel
Previous studies have shown the periodontal regeneration capability of amelogenin. 27,28 However, the protein solution could not stably attach to periodontal defects and would be rapidly cleared out by the metabolism system. 29,30 To achieve long-term treatment and controlled release of amelogenin, a chitosan-based hydrogel was chosen as the drug loading system. Previous research used this hydrogel for wound healing, tissue engineering, and cell scaffolding, showing ideal biocompatibility and degradation duration. 31 –33 The fabrication process of this chitosan-based hydrogel is briefly described as follows: 150 mg chitosan was dissolved in 0.1 mmol/L acetic acid and mixed with 9 mL glycerol phosphate (GP, 1.3M) in ice under vigorous stirring for 2 h. The loading drugs (amelogenin or Wnt3a) were mixed into 20 mg/mL hydroxyethyl cellulose and added into 4 mL chitosan-GP solution. The concentration of Wnt3a or amelogenin was adjusted to 0.6 mg/mL or 6 mg/mL for further use. After 2 h, a transparent chitosan-based hydrogel loaded with amelogenin or Wnt3a protein was formed.
Creation of periodontal bone defect model
Artificial bone defects were created on the GFP bone marrow transplantation C57 mouse model. All operations were performed under anesthesia as previously published research. 34 Briefly, after general anesthesia, 36 mice were selected from the GFP bone marrow transplantation C57 mouse models with their heads and limbs fixed. The gingiva was carefully incised and lifted to reveal mandible bone. After that, a bone defect (diameter: 1 mm depth: 2 mm) was created with a dental microdrill, 2 mm above the lower edge of the mandible. The gingival and the upper edge of mandibular were kept intact. For experimental groups, hydrogels containing amelogenin or Wnt3a were injected into the bone defect area and fixed with tightly sutured gingival tissue.
Grouping of in vivo experiment
The bone defect GFP bone marrow transplantation C57 mouse models were randomly divided into three groups. Each group contains 12 mice, with their right mandibles set as an experimental group which will be implanted with hydrogels carrying amelogenin or Wnt3a (Groups A & B,). The left side of their mandible bones was set as a self-control group, which will be injected with hydrogel only (Group C). The third group was a blank-control group (Group O) whose mandible bone wounds were treated with natural saline only. Our pilot experiment has proven the cell homing effect of amelogenin, at the concentration of 0.6 mg/mL, and reached its maximum effect at 6 mg/mL. Accordingly, two subgroups (each containing six mice) were set for Groups A and B, respectively. The mice in Group A1 were treated with 0.6 mg/mL amelogenin, while Group A2 was treated with 6 mg/mL amelogenin. Group B1 was treated with 0.6 mg/mL Wnt3a, while 6 mg/mL Wnt3a was used on Group B2 mice. The bone defect mouse model was executed at days 5 and 30 after the injection, and the mandible bone was harvested for further observation.
Micro-CT and histological observation
The prementioned mandibular bone tissue was harvested and fixed in 4% paraformaldehyde for 24 h. Then micro-CT (scan resolution 12 μm, tube voltages 70KV, tube current 114μA, exposure time 200 ms) was used to examine bone recovery of the wound sites. Artificial bone defect areas were set as the region of interest. Bone tissue regeneration was evaluated by bone volume/total volume (BV/TV), bone surface (BS)/BV, and trabecular number.
The samples were then decalcified and dehydrated through a serial alcohol gradient and embedded in paraffin wax blocks before immunostaining. Five-micrometer-thick bone tissue sections were dewaxed in xylene, rehydrated through decreasing ethanol concentrations, and washed in PBS. The cell nucleus was stained with DAPI (Sigma, Saint Louis, Missouri, USA) at a 1:5,000 dilution in PBS. Finally, a fluorescence microscope was used to examine the GFP BMMSCs homing at bone defect sites. Hematoxylin and eosin (H&E) and Masson staining was also used to observe tissue recovery.
Statistical analyses
The data are shown as mean ± standard deviation. Student’s t-tests were used for two independent-sample comparisons. A P value of <0.05 was considered statistically significant. All statistical analyses were performed in GraphPad version 8.0.
Results
Culture and identification of BMMSCs
BMMSCs were extracted from C57BL/6 mice and identified according to a previously reported protocol. The isolated cells were cultured and expanded to the third passage for characterization. Flow cytometry showed that these cells highly express stem cell markers, CD29 and CD90, and do not express hematopoietic cell markers, CD34 and CD45 (Fig. 1A). The adipogenic, osteogenic, and cartilage differentiation abilities were judged in Fig. 1B. These phenotypic and functional assessments collectively verified the successful isolation and characterization of mouse BMMSCs with typical mesenchymal stem cell properties.
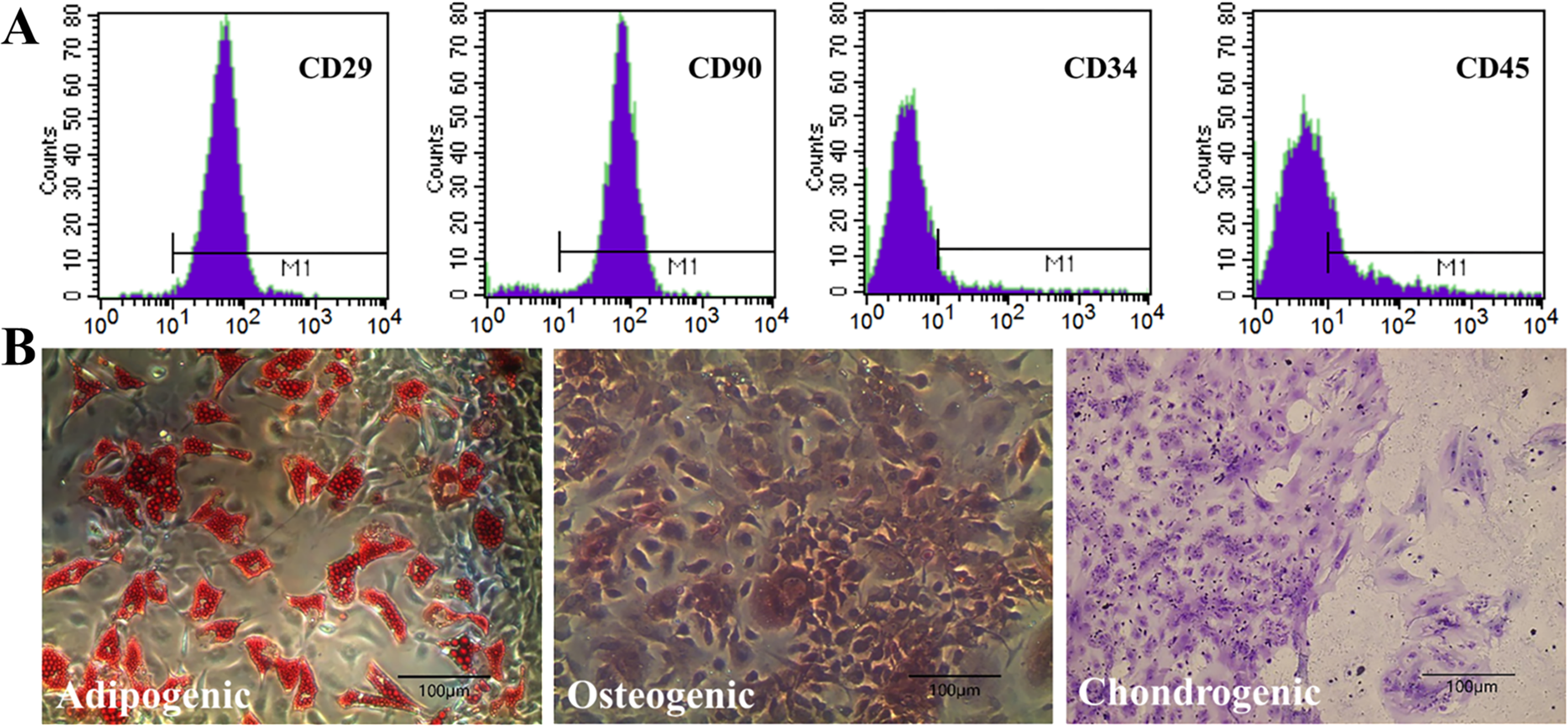
The identification of BMMSCs.
Transwell migration assay
Transwell results showed that the presence of Wnt3a and amelogenin significantly increased cell migration (P < 0.01) (Fig. 2A, B, D, E). Furthermore, amelogenin shows a concentration-dependent effect on cell migration (Fig. 2F) and reached a maximum effect at approximately 50 ng/mL. On the contrary, the Wnt pathway inhibitor, DKK-1, effectively suppressed amelogenin -induced migration (P < 0.01) (Fig. 2C), which further proved the connection between amelogenin-induced cell migration and Wnt/β-catenin pathway.

Transwell migration assay. BMMSCs were treated with amelogenin
Immunofluorescence analysis
Immunofluorescence was used to identify the changes in beta-catenin localization and expression when BMMSCs were cultivated with amelogenin. The results showed that beta-catenin was detected in the cytoplasm and peripheral sites of intercellular connections in the control group. However, after exposure to 100 ng/mL amelogenin for 24 h, beta-catenin was predominantly identified in the nucleus of cells, which was colorized by nuclear staining DAPI (Fig. 3C, D). For comparison, Wnt-pathway blocker DKK-1 downregulated beta-catenin expression and nuclear translocation in BMMSCs, as shown in Figure 3G, H. These results strongly support our hypothesis that amelogenin triggered Wnt-pathway in BMMSCs and induced beta-catenin nuclear translocation, and such effect could be blocked by DKK-1.
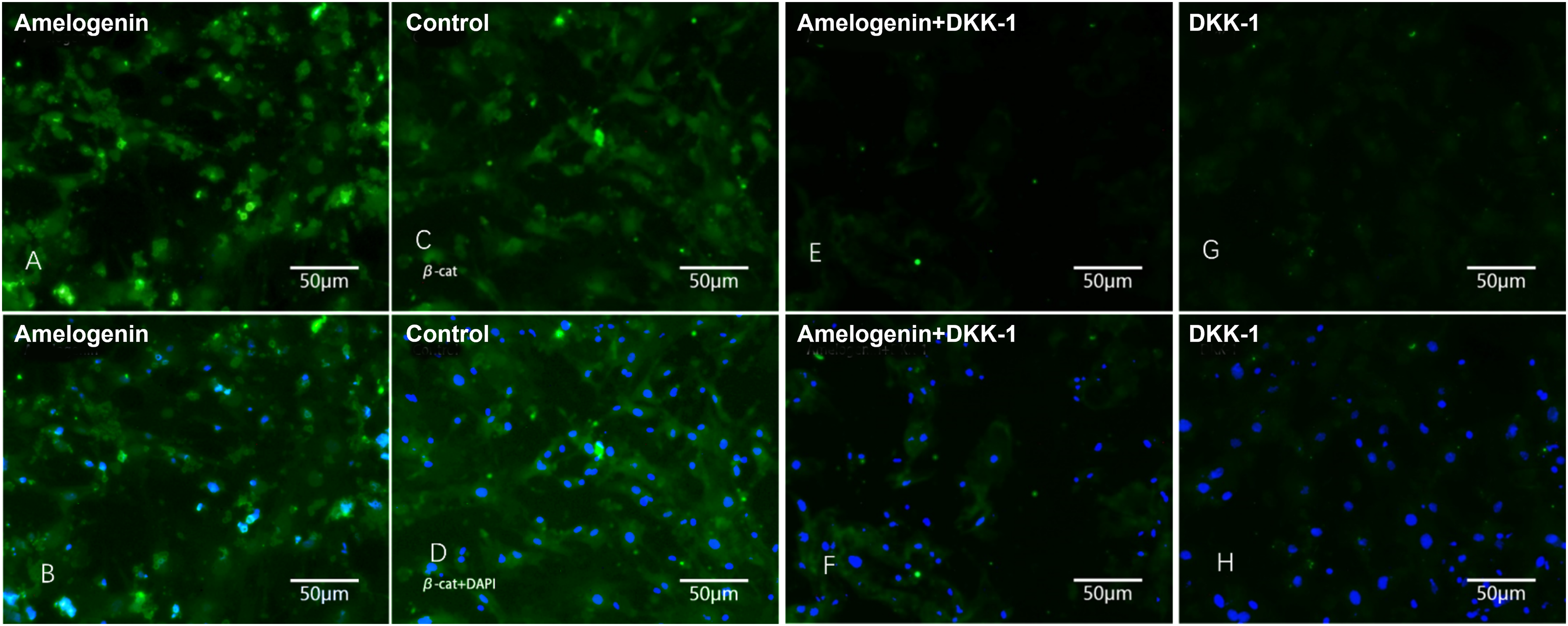
Immunofluorescence assay of bone marrow mesenchymal stem cells (BMMSCs). Cells were treated with amelogenin
Cell homing experiment
GFP-labeled BMMSC transplantation
The study used a myeloablative conditioning regimen in C57BL/6 mice, followed by transplantation of GFP-labeled bone marrow cells via tail vein injection. Flow cytometry conducted at 4 weeks post-transplantation revealed robust engraftment, with donor cell chimerism reaching 96% in the experimental group. In contrast, control animals exhibited minimal background fluorescence (Fig. 4A). These findings confirm the successful establishment of the GFP-labeled bone marrow transplantation model.
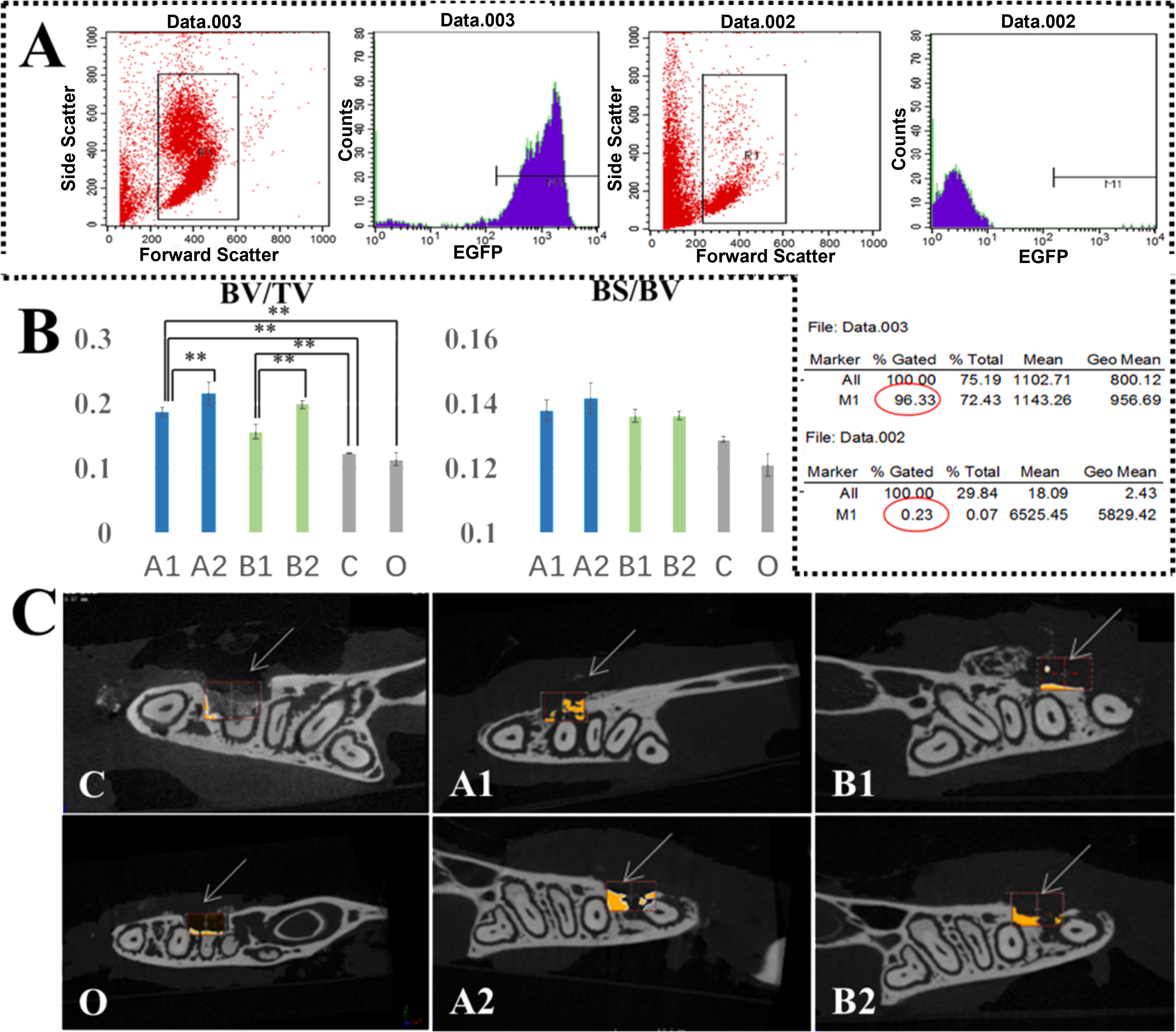
Periodontal bone regeneration assay.
Periodontal bone regeneration assay
The mandible samples were harvested 30 days postsurgery. Micro-CT, histological observation, and immunohistochemical observation were used to assay cell homing and tissue healing. Micro-CT showed significant bone regeneration (marked in orange) in the periodontal defect area with amelogenin (Group A) or Wnt3a (Group B) hydrogels, while the control (Group C) and blank (Group O) groups exhibited persistent bone defects at 30 days after hydrogel injection (Fig. 4C). Moreover, higher concentration formulations (6 μg/μL A2 and B2) demonstrated enhanced bone regeneration compared with their lower concentration counterparts (0.6 μg/μL A1 & B1) in both treatment groups, as shown in Fig. 4C.
Bone regeneration was quantitatively assessed at day 30 using BV/TV and BS/BV measurements. The result showed that, compared with the control group and blank control group, BV/TV was significantly increased in mice treated with amelogenin and Wnt3a, whether in higher or lower concentration groups (P < 0.01). Notably, the high-concentration groups (A2 and B2, 6 μg/μL) exhibited superior bone formation versus their low-concentration counterparts (A1 and B1, 0.6 μg/μL) (P < 0.01), indicating a dose-dependent therapeutic effect. While the amelogenin group showed marginally higher BS/BV values than Wnt3a-treated specimens, this difference lacked statistical significance (P > 0.05), suggesting comparable bone quality across treatment groups (Fig. 4B). These findings collectively demonstrate that both amelogenin and Wnt3a effectively promote bone regeneration in a concentration-dependent manner, with no significant difference in the structural quality of newly formed bone tissue.
Histological assessment
H&E and Masson staining was used to evaluate tissue healing. The result shows that, 30 days after injection, newly grown bone tissue (red arrow) was found at the periodontal defect area in amelogenin and Wnt3a groups (Fig. 5A, B), while control groups exhibited persistent defects with incomplete healing (Fig. 5C). Masson staining provided further characterization of bone maturation, with newly formed bone matrix staining blue (red arrow) contrasting with the red-stained mature bone (Fig. 5D–F). These histological findings corroborate the micro-CT results, confirming enhanced bone regeneration following either amelogenin or Wnt3a administration.

Histological evaluation of periodontal bone regeneration using Hematoxylin and eosin (H&E) and Masson staining.
Cellular assessment
Fluorescence microscope was used to observe cell homing to the periodontal defect area after bone marrow transplantation was performed. While all groups exhibited some GFP+ cells in the surgical area at 5 days post-transplantation (Fig. 6), quantitative analysis demonstrated significantly enhanced homing in both amelogenin (A1 and A2)- and Wnt3a-treated (B1 and B2) groups compared with controls. Notably, the 6 μg/μL concentration groups (A1 and B1) showed superior BMMSC recruitment compared with their 0.6 μg/μL counterparts (A2 and B2), consistent with our in vitro Transwell assays demonstrating concentration-dependent migratory effects (data not shown). These findings establish a clear dose–response relationship for both therapeutic agents in promoting stem cell homing to periodontal defects.
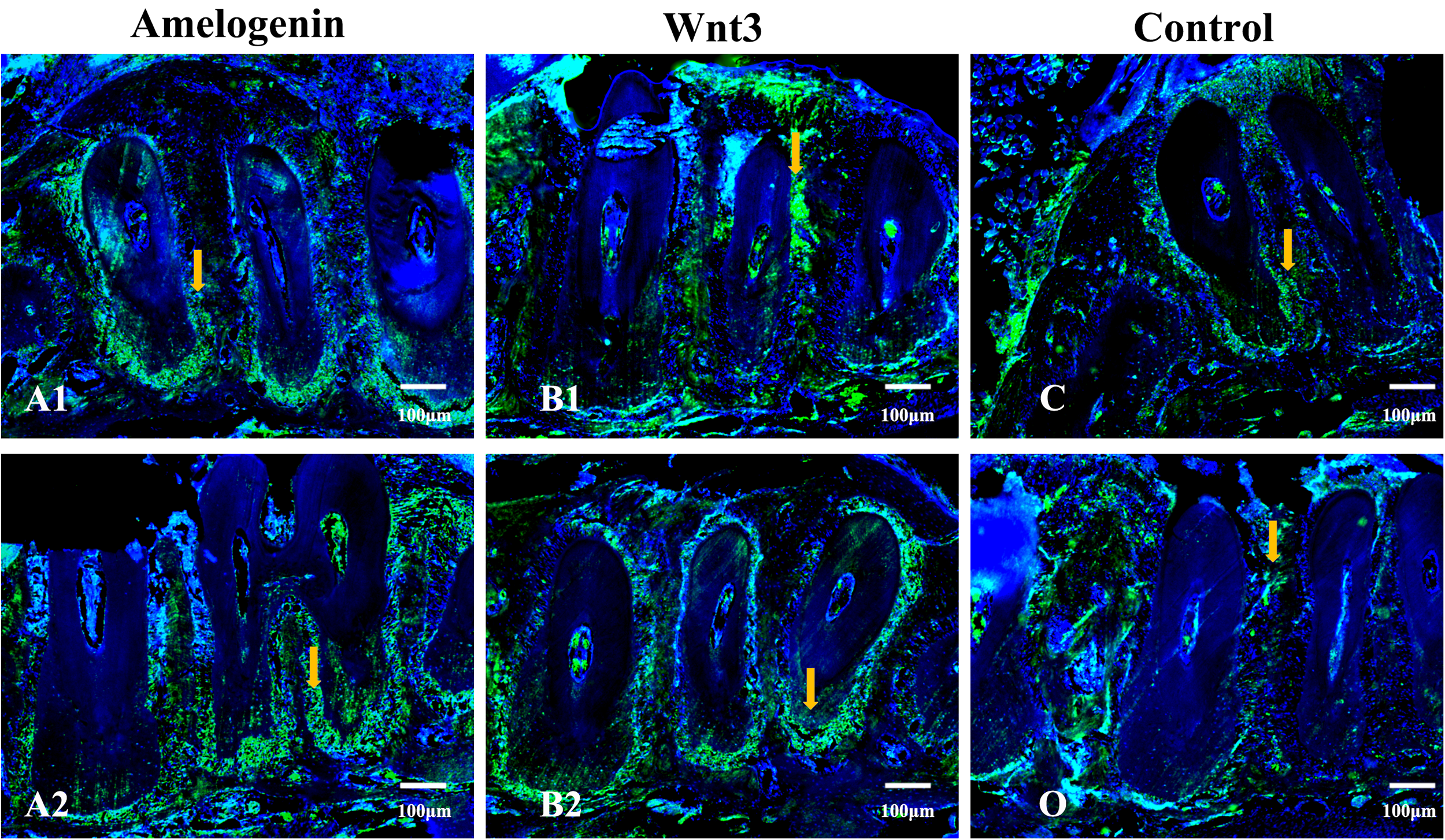
Fluorescence microscopic examination of bone samples in the Amelogenin, Wnt3a, and Control group mice at 5 days of bone marrow transplantation. More GFP+ transplanted BMMSCs (white arrow) were observed in the Amelogenin and Wnt3a groups. Cell nucleus was marked in blue fluorescence. (
Discussion
Periodontal disease represents a global health challenge, affecting over 50% of the world’s population and frequently leading to tooth loss through progressive bone destruction. 1 Despite numerous attempts to develop effective regeneration strategies, current therapeutic outcomes remain unsatisfactory. Among emerging approaches, stem cell-based therapies have shown particular promise for periodontal tissue regeneration. Conventional methods relying on stem cell transplantation involve isolating, expanding, and seeding cells onto scaffolds before surgical implantation into defect sites. Various cell sources, including BMMSCs, periodontal ligament stem cells, dental follicle cells, adipose-derived stem cells, and apical papilla stem cells, have been investigated for this purpose. However, several critical limitations hinder the clinical translation of these approaches. Unlike life-threatening cardiac or neural defects, periodontal conditions present unique challenges as elective procedures. The high cost, high risk, or high technology required from dentists could hinder the clinical application of stem cells for periodontal regeneration. Technically, it is hard to harvest periodontal stem cells without the extraction of teeth. Stem cells from other tissues, such as bone marrow, still require extra surgery, which will cause damage to healthy tissue. Besides, the number of translated stem cells may decrease due to apoptosis or migration. Furthermore, allogeneic stem cells may cause potential immunological or even ethical risks. These collective limitations underscore the need for alternative regenerative strategies that can overcome the inherent constraints of traditional stem cell transplantation approaches.
The cell homing strategy presents a promising alternative approach, leveraging the body’s innate capacity to recruit circulating stem cells to target tissues. The concept now also refers to stem cells migrating to the given region of the anatomy. Stem cell homing was studied in many researches about the regeneration of the cardiac nerve. It was also introduced into dental and facial regeneration. The tooth-shaped scaffolds were put under the skin of rats, to achieve periodontal regeneration. Although the source of the cells was not explored, it still proved the possibility of a cell homing strategy for periodontal regeneration. Stem cell translation strategy faces some challenges because of the specialty of periodontal defects. However, just because of other specialties, the cell homing strategy is more suitable for periodontal regeneration, technically and economically. For example, the volume of periodontal tissue is much smaller compared with the skin, heart, and joints. Fewer stem cells are needed for regeneration, which makes it easier to gain periodontal regeneration by inducing stem cell homing. These researches may provide a potential therapeutic idea and method for periodontal defects.
Many cytokines have been used to achieve stem cell homing such as TGF-β, 35 Concentrated Growth Factor (CGF), 36 and amelogenin. 6 Of these, amelogenin has emerged as particularly promising, showing clinically significant periodontal regeneration outcomes. As the primary component of enamel matrix proteins, amelogenin has been widely used in periodontal therapy, although its precise mechanistic action remains incompletely understood. 14 Existing studies have documented amelogenin’s regenerative capacity across multiple periodontal tissues, demonstrating its ability to promote the simultaneous regeneration of PDL, alveolar bone, and cementum. 37
The mechanistic basis of amelogenin-induced periodontal regeneration has been extensively investigated through multiple research avenues. Current evidence reveals that amelogenin exerts complex biological effects through two primary mechanisms: osteogenic promotion 15 and inflammatory modulation. 38 Further study shows that multiple signal pathways were involved in this process, such as ERK 39 or autocrine/paracrine 40 pathways. However, these findings fail to fully account for the clinically observed phenomenon of stem cell accumulation in amelogenin-treated defect sites, suggesting the existence of additional, uncharacterized mechanisms governing stem cell recruitment and proliferation.
Extensive research has established Wnt3a as a potent inducer of mesenchymal stem cell migration and homing, 41 –43 with emerging evidence implicating the Wnt/β-catenin pathway in amelogenin-mediated periodontal regeneration. 44 This aligns with Wnt signaling’s fundamental role in developmental processes, including cell migration, proliferation, and differentiation. 45 Our experimental data demonstrate that amelogenin treatment significantly enhances β-catenin nuclear translocation, an effect completely abolished by the Wnt inhibitor DKK-1. Comparative analyses revealed that amelogenin and Wnt3a exhibit comparable efficacy in promoting BMMSC migration and periodontal regeneration, as evidenced by both in vitro Transwell assays and in vivo GFP-labeled cell tracking in C57 mouse models. While both compounds displayed concentration-dependent homing effects and similar regenerative outcomes, their mechanisms differ substantially. While Wnt3a confirmed pathway involvement, amelogenin’s effects are mechanistically distinct. Amelogenin engages GRP78 receptors and suppresses β-catenin degradation, offering a localized therapeutic strategy for periodontal regeneration. Future studies may use amelogenin-specific inhibitors or gene-edited models to further dissect its independent mechanisms. It is reasonable to believe that amelogenin achieves periodontal regeneration through the Wnt pathway. While our data support amelogenin’s involvement in Wnt signaling, the precise mechanism governing its promotion of stem cell migration—whether mediated directly through Wnt3a or alternative pathways such as MSX (a key regulator of craniofacial morphogenesis and cell motility)—remains experimentally unresolved. This represents a key limitation of our study. Although we have demonstrated amelogenin’s potent homing effect and its association with β-catenin stabilization within the broader Wnt signaling context, the specific upstream triggers and potential parallel signaling pathways (e.g., MSX-related cascades) activated by amelogenin-GRP78 interactions to drive cellular migration require further investigation. As proposed, future studies using targeted inhibitors or genetic models will be essential for elucidating these mechanistic details.
In conclusion, we demonstrated that amelogenin could induce bone marrow mesenchymal stem cells homing as Wnt3, which is crucial in periodontal bone regeneration. However, this study underscores amelogenin’s unique role in periodontal regeneration, independent of Wnt3a. Its indirect signaling mechanisms highlight potential clinical advantages for targeted tissue repair. These findings give us a further understanding of amelogenin-induced periodontal regeneration and suggest a potential therapeutic target for future clinical application.
Footnotes
Acknowledgments
The authors extend their sincere gratitude to all staff members of the Orthodontics Department, College of Stomatology, Xi’an Jiaotong University, for their invaluable technical and logistical support throughout this study.
Data Availability Statement
Data will be made available on request.
Author Disclosure Statement
All authors declare no conflicts of interest.
Funding Information
This work was financially supported by the following grants: National Natural Science Foundation of China (Grant Nos. 81200822, 81371694).