
Abstract
p21 is a cell cycle regulator that has been implicated in regeneration of tissues and in development of certain tumors. p21 inhibition also enhances bone regeneration after injury in p21−/− mice. To translate these findings to the clinic, we sought an FDA-approved p21 attenuator. UC2288, a derivative of sorafenib, selectively inhibits p21 independently of p53 and induces apoptosis in cancer cells. Given the central role of p21 in mesenchymal stem cell (MSC) proliferation and differentiation, its effects on MSCs merits investigation but remains unknown. Consequently, we hypothesized that UC2288 will improve the osteogenic potential of mesenchymal stem cells by suppressing p21. First, we examined the differential interaction of UC2288 with human bone marrow (BM) MSCs compared with breast cancer cells via viability assays. Increased cell death was observed in cancer cells, particularly at higher concentrations and with longer interaction times, whereas MSCs demonstrated lower cell death. Gene expression assay revealed upregulation of osteogenic genes, though the specific genes overexpressed varied depending on the culture medium. Interestingly, the culture medium also affected p21 expression, where p21 expression was upregulated in DMEM/F12 and downregulated in alpha-MEM as evidenced in gene and protein expression assays. Alizarin Red staining confirmed increased mineralization when UC2288 or UC2288+osteogenic factors were added. These findings indicate that UC2288 promotes osteogenesis in BM-MSCs in a concentration- and time-dependent manner. Further research is needed to optimize conditions for preclinical and clinical translation as an anabolic bone formation therapy.
Introduction
Bone fractures in young, healthy individuals with a noncomminuted fracture typically heal well. However, individuals who may have more complicated fractures, advanced age, or diseases like osteoporosis where the natural healing ability is disrupted may require external intervention. 1 Various methods are currently used to treat bone fractures commonly involving surgical interventions such as internal fixation and/or bone grafting. 2 Autologous bone grafting is the gold standard for enhancing bone repair in nonunion and delayed unions. 3 However, this method is associated with drawbacks that include availability of graft volume, donor site morbidities, and potential complications such as severe pain and formation of hematoma. 4 In recent years, the use of orthobiologics (biological substances to improve growth of bone and associated soft tissues) has gained popularity. Although orthobiologics have gained popularity, challenges remain regarding consistency, quality, and patient-specific responses. Furthermore, regulatory challenges and the cost to patients render this method prohibitive. 5 Gene therapy is another alternative where specific osteogenic or chondrogenic genes can be targeted to elicit favorable outcomes. 6 Genes involved in cell cycle regulation such as p53, p21, and p16 have been implicated in tissue regeneration in addition to the role they play in tumorigenesis. 7 Wang et al. demonstrated a role for p53 in regulating differentiation of osteoblasts and osteoclasts, where p53−/− mice displayed high bone mass and increased bone formation via increased expression of osterix (OSX). 8 Ding et al. demonstrated that in aged p16−/− mice, fracture healing was significantly accelerated and a concomitant increase in expression of osteogenic genes such as ColX, MMP13, and OSX was observed. 9 Premnath et al. demonstrated that compared with wildtype mice, in mice in which p21 is absent, bone formation, rate of bone formation, and bone mineral density are increased after injury. 10 They hypothesized that the increased bone formation capacity could be attributed to the presence of p21−/−-deficient MSCs at the site of injury where they act directly or release trophic factors that improve bone healing outcomes. Another study reported that loss of p21 increases the rate of osteoblast proliferation and differentiation. 11 These findings demonstrate that absence of p21 improves osteogenic differentiation and increases bone healing outcomes after injury. To translate these findings to the clinic, a method to inhibit p21 is required. Jiang et al. utilized siRNA to inhibit p21 expression in a skin wound healing model to improve healing outcomes. 12 While this study demonstrated effective knockdown of p21, translating siRNA therapies to humans would require suitable delivery mechanisms in addition to further research into intracellular processing as siRNAs leaves endosomal/lysosomal system to be returned to the surface. 13 A pharmacological alternative may be suitable to aid in knockdown of p21.
UC2288 is a small molecule that inhibits p21, a second-generation analog of sorafenib. Sorafenib, an FDA-approved drug, inhibits angiogenesis, suppresses tumor cell proliferation, and facilitates apoptosis in hepatocellular carcinoma. 14 UC2288 differs from sorafenib, as it does not affect levels of p-ERK protein or block the activity of Raf kinases. 15 UC2288 attenuates p21 protein at the transcription or posttranscription level and not via protein degradation. UC2288 has primarily been investigated in regard to cancer, including nasopharyngeal carcinoma 16 and neuroblastoma. 17 p21 acts based on its cellular context, its localization, and its posttranslational modifications. Therefore, it is hypothesized that while UC2288 increases apoptosis in cancer cells, it will affect differentiation in MSCs as inhibition of p21 has been shown to increase osteoblast differentiation. 11,18 To this end, in this study, we will evaluate the efficacy of UC2288 in reducing p21 levels in MSCs and assess osteogenic capacity of UC2288-treated MSCs.
Materials and Methods
Chemicals and reagents
UC2288 (purity ∼99%) was purchased from Abcam (Cambridge, UK) and dissolved in dimethyl sulfoxide, and the aliquots were stored at −20°C. Concentrations of 0.1 μM, 1 μM, 2.5 μM, 5 μM, and 10 μM were prepared by diluting required volumes directly in cell culture media.
Cell culture
Bone marrow-derived mesenchymal stem cells (BM-MSCs; American Type Culture Collection [ATCC] PCS-500-012, Lot number 81007237) were purchased from the ATCC, while MDA-MB-231 breast cancer cells (ATCC HTB-26) were generously provided by Dr. Qingsu Cheng. BM-MSCs were cultured in alpha-MEM (GIBCO) or DMEM/F12(GIBCO), and MDA-MB-231 cells were cultured in DMEM (GIBCO), all supplemented with 10% fetal bovine serum (GIBCO). Cells were maintained at 37°C with 5% CO2, with media changes every other day until reaching 80% confluency, at which point they were sub-cultured. BM-MSCs between passages 2–4 were used for experiments. These cells were pre-characterized for cell-specific markers like CD73, CD90, CD105, CD45, and CD19. Osteogenic media was prepared by supplementing the base media with 100 nM dexamethasone, 50 μg/mL l-ascorbic acid, and 10 mM β-glycerophosphate (henceforth referred to as OSTEO). 19 For control samples, only basal media was added.
Metabolic and cell viability assay
An MTT assay was performed to test if UC2288 is cytotoxic to BM-MSCs cells using 3-(4,5-dimethylthiazol-2-yl)−2,5-diphenyl-2H- tetrazolium bromide (MTT) (Thermo Fisher Scientific, MA). BM-MSCs cells were treated with UC2288 concentrations of 0.1 µM, 1 µM, 10 µM, 0.1 µM + OSTEO, and 1 µM + OSTEO. Media with no drug served as controls. All conditions were seeded in 96-well plates at 5,000 cells per well in triplicates at 37°C. Cells were tested at 48 h and 7 days. We used a 48-h time point to examine the short-term effects of UC2288 on cells and a 7-day time point to capture longer-term interactions, which also aligned with gene expression analysis. Samples were prepared as per manufacturer’s instructions, and the plates were read at 570 nm with a Varioskan LUX plate reader (Thermo Scientific, MA). Ordinary one-way ANOVA was performed to analyze significance. P < 0.05.
A LIVE/DEAD® Viability/Cytotoxicity kit (Fisher Scientific, Pittsburgh, PA) was used to assess the viability of BM-MSCs and MDA-MB-231 breast cancer cells cultured with no-drug, 2.5 μM, 5 μM, and 10 μM concentration of UC2288 at 7, 14, 21 days’ time point in triplicate. The cell viability was evaluated through visualization of cell morphology using calcein AM and ethidium homodimer-1 (EthD-1). The calcein binds to live cells to produce green fluorescence, while EthD-1 binds to dead cells and produces red fluorescence. The staining solution was prepared in D-PBS at a concentration of 1 µM calcein and 4 µM EthD-1. The media in the sample wells was aspirated and replaced with staining solution and incubated at room temperature for 30 min. The samples were washed with D-PBS, then imaged with the Echo revolve microscope using TXRED and FITC filters for dead and live cells, respectively, in Fluorescence Overlay Mode.
Osteogenic gene expression
To assess changes in gene expression, BM-MSCs were seeded at 5,000 cells/cm2 in a 24-well plate and treated with UC2288 at 0.1 µM, 1 µM, and 10 µM, as well as 0.1 µM + OSTEO and 1 µM + OSTEO in DMEM/F12. Based on trial-and-error optimization and other studies in our lab, it was determined that at 2.5 µM, the highest osteogenic gene expression was observed in DMEM/F12. Therefore, when testing alpha-MEM, cells were treated with 2.5 µM UC2288 and 2.5 µM + OSTEO in alpha-MEM for 7 days. No-drug treated cells served as controls. Total RNA extraction was performed with RNeasy Micro Kit (Qiagen, MD) as per manufacturer’s instructions. The collected total RNA was quantified using a NanoVue Spectrophotometer (GE Healthcare), and samples were stored at −80°C. Reverse transcription was performed with a High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific) using a MiniAmp Thermal Cycler (Thermo Scientific), samples were then stored at −20°C. To perform reverse transcription quantitative polymerase chain reaction (RT-qPCR), a mix of TaqMan Universal PCR Master Mix (Thermo Fisher Scientific) and gene of interest (TaqMan), which were 18s (endogenous control), p21 (CDKN1A), Sp7 (Osterix), RUNX2, were mixed with RNase-free water, and cDNA was prepared. 18s was chosen as the endogenous control, as it more stable under different conditions. 20 Samples were loaded on to a MicroAmp Optical 96-well Reaction Plate (Thermo Fisher Scientific) and processed in a QuantStudio 3 Real-Time PCR System (Thermo Fisher Scientific) with an initial denaturation at 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and annealing/extension at 60°C for 1 min. The target gene’s expression levels were normalized to endogenous 18s. Relative change in gene expression was calculated using 2−ΔΔCt with reference to the no-drug control samples. Results were averaged from triplicate samples. After the results were gathered, two-way ANOVA with Tukey post hoc was preformed to statistically analyze the outcomes. P < 0.05 was considered significant.
Histology
Alizarin Red staining was performed to assess the mineralization capacity of MSCs after interaction with UC2288 for 21 days. BM-MSCs were seeded at 5,000 cells/cm2 in a 24-well plate and treated with 2.5 µM UC2288 and 2.5 µM + OSTEO in alpha-MEM, with no-drug in alpha-MEM serving as controls. The wells were washed with D-PBS, fixed with cold 70% ethanol, then stained with 2% Alizarin Red stain solution.
Western blot
Total protein was isolated from cell lysate using RIPA lysis buffer (Thermo Scientific, 89900). Protein concentration was assessed using Pierce BCA Protein Assay Kit (Thermo Scientific, 23225). Proteins were loaded onto premade Novex™ Tricine Mini Protein Gels, 10% to 20%, 1.0 mm (Thermo Scientific, EC66252BOX). Next, proteins were transferred from the gel onto a 0.45-μm-pore-size nitrocellulose membrane (Cytiva Amersham, 45004002). The membrane was blocked with 5% bovine serum albumin (BSA) (Sigma-Aldrich, A8022) in 1× TBST (20 mM Tris, pH 7.6, 150 mM NaCl, 0.05% Tween-20). Primary and secondary antibodies were diluted in 5% BSA/1× TBST. Protein blot bands were imaged using Odyssey Sa Imager (LiCOREbio), and densitometry of western blot bands performed using Image J. Primary antibodies: GAPDH (Abcam, ab9485), phosphorylated p21 (p-p21; Abcam, ab47300), osteocalcin (Cell Signaling Technology, 59757) and p21 (Cell Signaling Technology, 2947). Secondary antibodies: goat anti-Rabbit (Abcam, ab175773).
Results
MTT assay was conducted to evaluate the metabolic activity and viability of BM-MSCs treated with different concentrations of UC2288 for 48 h (Fig. 1A) and 7 days (Fig. 1B). At 48 h, cell viability remained stable across all conditions, with no significant reduction observed, indicating that short-term exposure to UC2288, even at 10 µM, does not have a substantial cytotoxic effect on BM-MSCs. However, after 7 days, a significant decrease in cell viability was observed at 10 µM UC2288, suggesting a time-dependent cytotoxic effect at higher concentrations. Further, 0.1 µM UC2288 maintained the highest viability, while 1 µM UC2288 showed a slight but not statistically significant reduction. The presence of osteogenic media did not appear to alter the effects of UC2288, as viability trends remained similar in both osteogenic and nonosteogenic conditions. These results suggest that while lower concentrations of UC2288 (0.1–1 µM) are well tolerated, prolonged exposure to higher concentrations (10 µM) leads to a significant decline in BM-MSC viability.
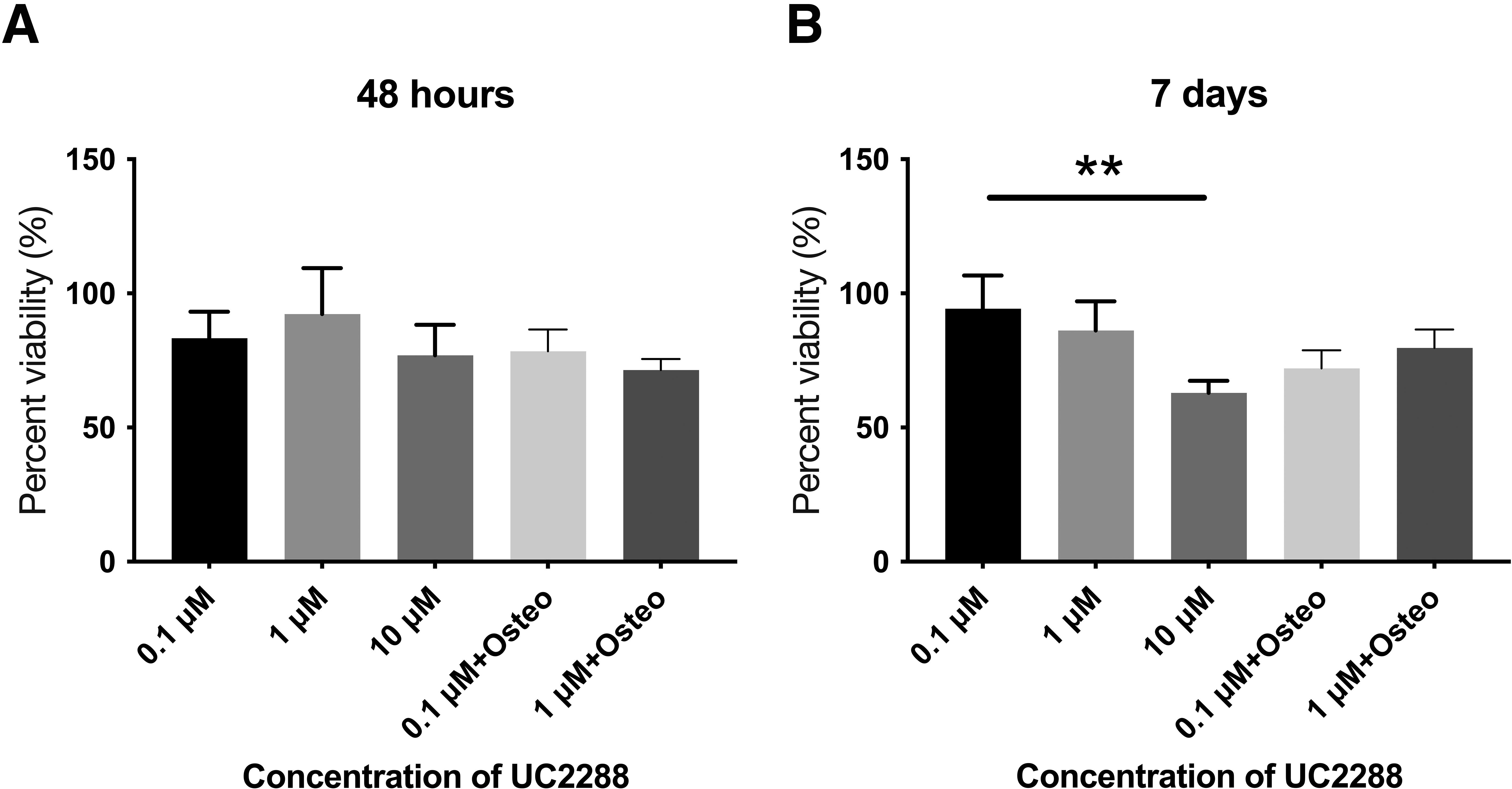
Percent viability of BM-MSCs when incubated with UC2288.
UC2288 is a derivative of sorafenib that is used clinically to treat cancers 15 . To evaluate if other cells respond differentially to UC2288, specifically cancer cells versus stem cells, we tested cell viability and morphology of both cancer and stem cells when treated with different concentrations of UC2288. We tested MDA-MB-231 (breast cancer) cells (Fig. 2A) and BM-MSCs (Fig. 2B) over 7, 14, and 21 days using Live/Dead staining. Green fluorescence represents live cells, while red indicates dead cells. Our results, like previous studies 15,17,21 , show that UC2288 is cytotoxic to cancer cells, while BM-MSCs exhibit greater resistance, particularly at lower concentrations. With MDA-MB-231 cells, all conditions involving UC2288 demonstrated diminishing confluency, with cell death increasing with time and concentration. At 2.5 µM, a slight decrease in cell density was observed, while 5 µM led to increased cell death, especially by day 14. By day 21, surviving cancer cells appeared slightly elongated, likely as a stress response rather than epithelial–mesenchymal transition, as this morphological change coincided with increased cell death, suggesting cytoskeletal disruption and apoptosis. 15 At 10 µM, cancer cells exhibited substantial cell death as early as day 7. In BM-MSCs, the no-drug control showed a characteristic spindle-shaped morphology with minimal red fluorescence over all time points. Cells treated with 2.5 µM UC2288 maintained morphology and confluency similar to no-drug controls. At 5 µM UC2288, a slight reduction in cell density was observed, but widespread cell death did not occur. At 10 µM, cells became more elongated, and by day 21, a noticeable reduction in cell density was evident, demonstrating concentration and time-dependent cytotoxicity. However, a substantial number of live cells remained compared with the cancer cells. Overall, MDA-MB-231 cancer cells were significantly more sensitive to UC2288, with widespread cell death at concentrations of 5 µM and higher, leading to almost complete cell death at 10 µM. In contrast, BM-MSCs exhibited greater resistance, with only mild reductions in viability at higher concentrations.
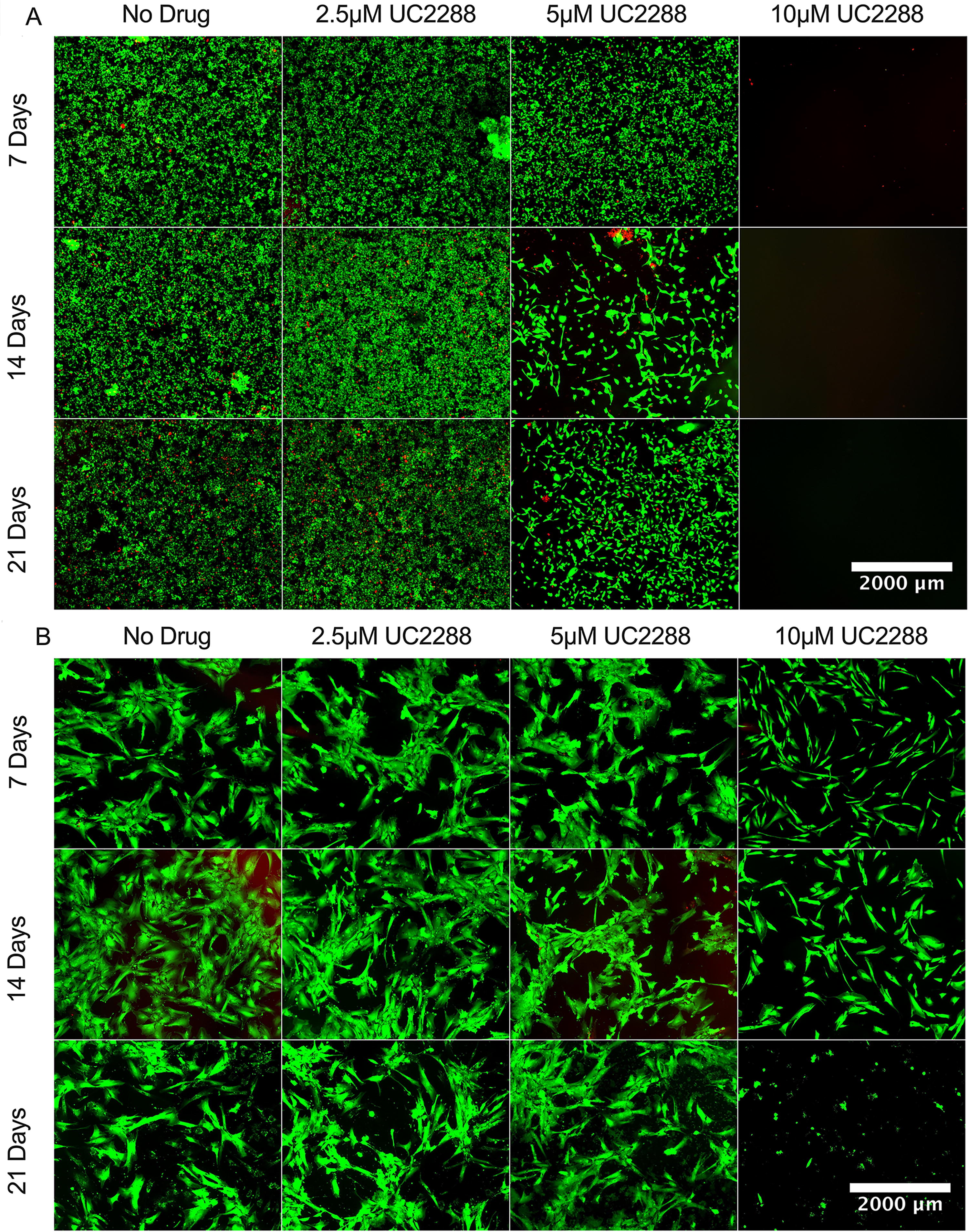
Cytotoxic effects of UC2288 on MDA-MB-231 cancer cells and BM-MSCs.
RT-qPCR analysis was performed to evaluate the relative gene expression of p21 and osteogenic markers in BM-MSCs cultured in DMEM/F12 (Fig. 3A) and alpha-MEM (Fig. 3B) after 7 days of treatment with UC2288. In DMEM/F12, p21 expression was significantly upregulated across all UC2288 concentrations. The expression of key osteogenic markers, Runx2 and Col1a1, also increased, particularly in the 1 µM condition, indicating that UC2288 may enhance osteogenic differentiation. In alpha-MEM, a contrasting expression pattern emerged, with only Sp7 showing the most significant upregulation, particularly in the OSTEO + 2.5 µM UC2288 condition. Runx2 displayed an upward trend but no statically significant increase in the treatment conditions compared with no-drug control. In contrast, p21 expression remained relatively unchanged, exhibiting a downward trend with different concentrations of UC2288 compared with no-drug control. This suggests that UC2288’s influence on osteogenic differentiation may be medium-dependent, with alpha-MEM specifically favoring Sp7 activation under osteogenic conditions.
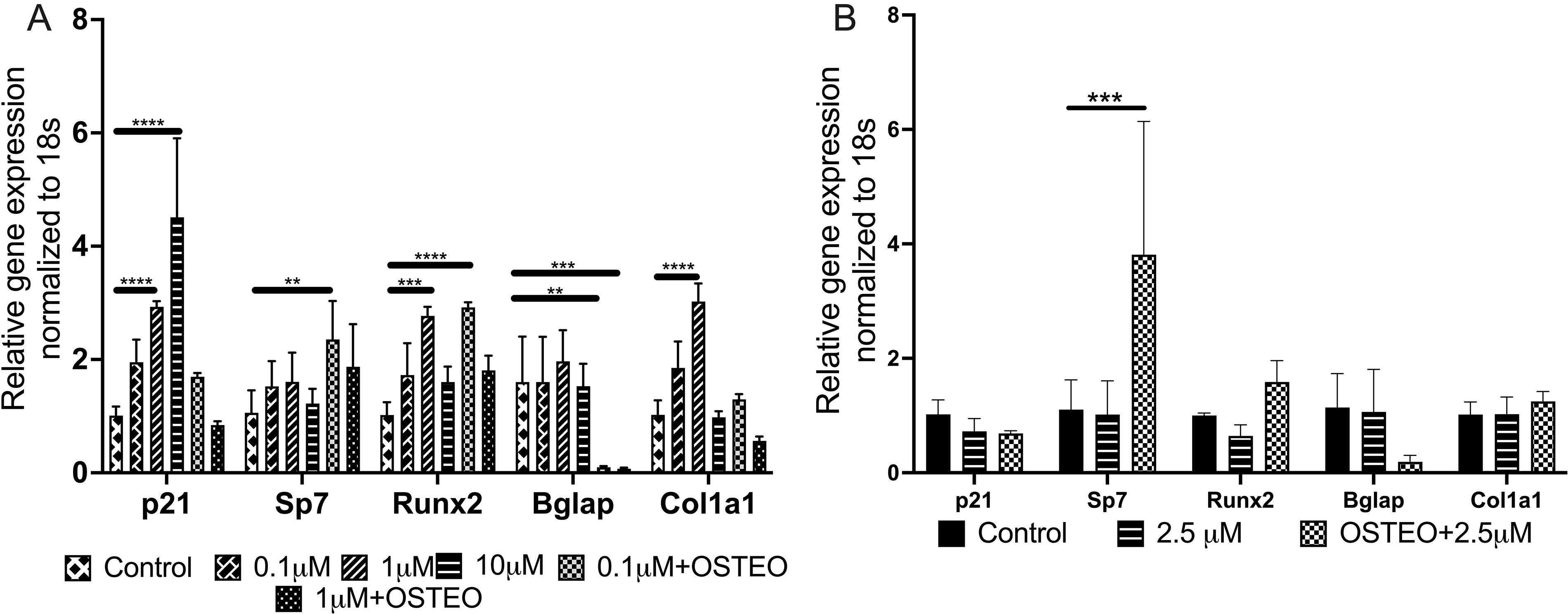
Relative expression of osteogenic genes and p21 after BM-MSCs were treated with varying concentrations of UC2288 for 7 days.
Alizarin Red staining assay was performed to assess functional outcomes such calcium deposition and osteogenic differentiation in BM-MSCs cultured under different conditions (Fig. 4). In the no-drug alpha-MEM group (Fig. 4A), minimal staining was observed, indicating negligible mineralization and a lack of osteogenic differentiation in the absence of osteogenic supplements or UC2288. In the alpha-MEM + OSTEO condition (Fig. 4B), increased red staining was evident, signifying successful induction of osteogenesis. The addition of 2.5 µM UC2288 to the osteogenic medium (alpha-MEM + OSTEO + 2.5 µM UC2288) (Fig. 4C) resulted in the most pronounced Alizarin Red staining, suggesting a significant enhancement in mineralization compared with the osteogenic media alone. These results indicate that UC2288 may promote osteogenic differentiation and mineralization in BM-MSCs when combined with osteogenic factors.
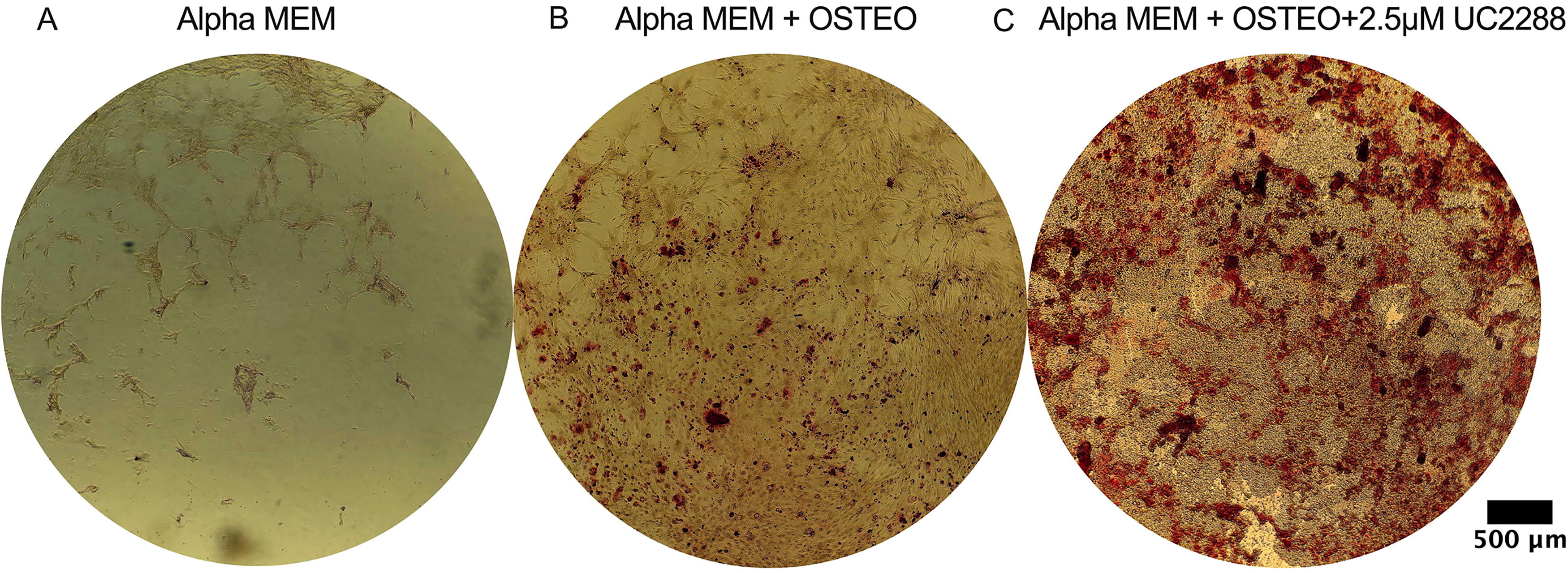
Alizarin Red staining of BM-MSCs incubated with different concentrations of UC2288 and osteogenic media for 21 days.
Gene expression analysis showed that BM-MSCs cultured in DMEM/F12 exhibited p21 upregulation, whereas those cultured in alpha-MEM showed no significant changes. We next examined whether these differences were also reflected at the protein level. Western blot analysis was performed to assess protein expression of p21, p-p21, and osteocalcin (a late osteogenic marker) in response to two different concentrations (2.5 µM of UC2288 in alpha-MEM and 1 µM of UC2288 in DMEM/F12) of the treatment, compared with control groups in one biological replicate of each condition. The experiments were conducted in two sets: one with BM-MSCs cultured in DMEM/F12 as a control and the addition of 1 µM UC2288 (shown in image 5A), and the other with BM-MSCs cultured in alpha-MEM basal medium as a control and the addition of 2.5 µM UC2288 (shown in image 5B). The presence of p-p21 and osteocalcin was not detected in any of the conditions. Densitometric quantification of p21, normalized to GAPDH protein levels, shows a slight increase with 1 µM UC2288 in DMEM/F12 (Fig. 5C) and a decrease in p21 expression with 2.5 µM UC2288 in alpha-MEM (Fig. 5D) and.
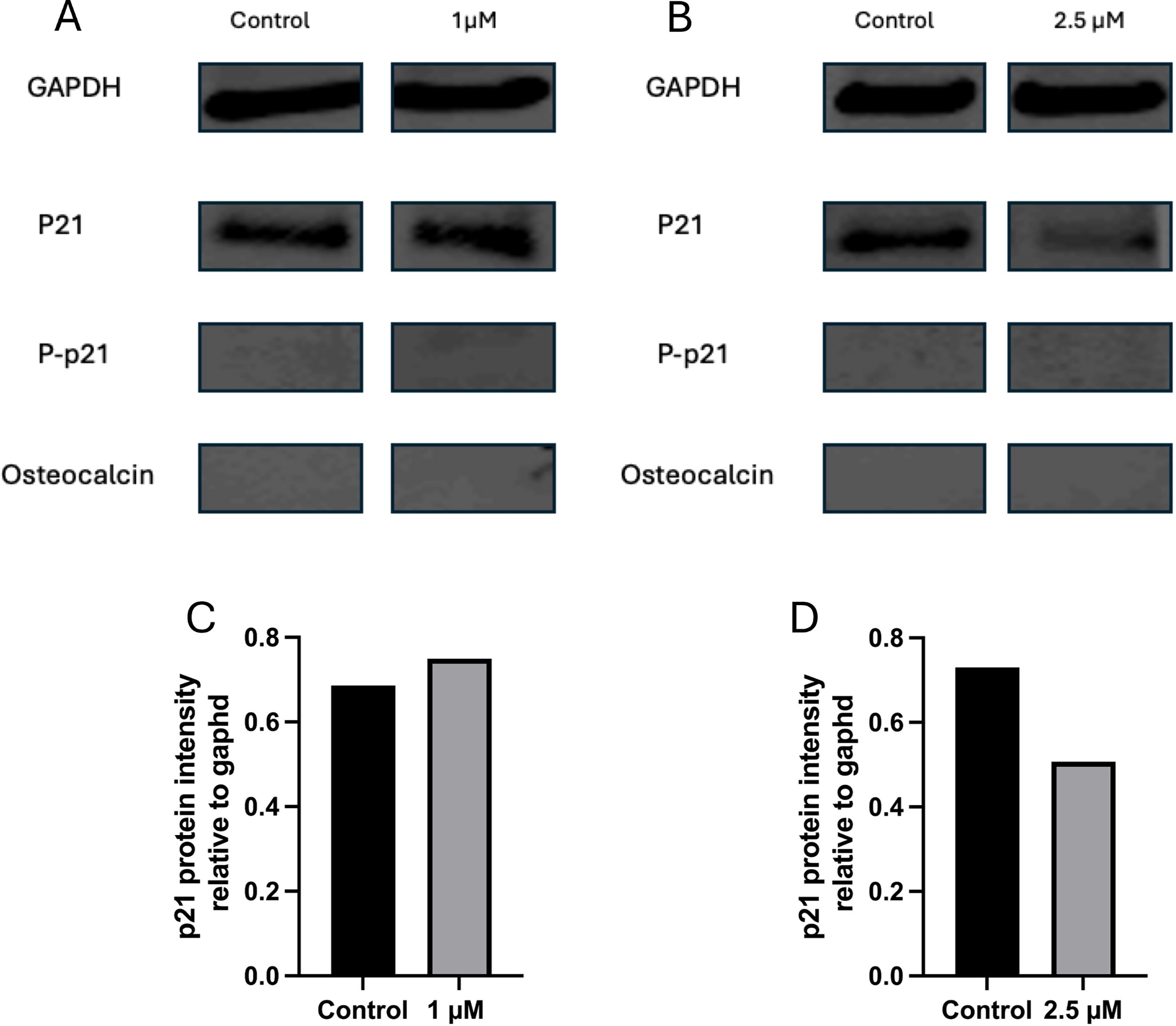
Western blot analysis of BM-MSCs treated with UC2288 for 7 days.
Discussion
UC2288 was assessed for its ability to enhance the osteogenic capacity of BM-MSCs by modulating p21 expression. 12,22 The cytotoxicity assay results align with previous findings, 23 demonstrating that 10 μM of UC2288 induced significant cell death in both cancer cells and MSCs. However, while MDA-MB-231 cancer cells exhibited sensitivity to UC2288 at lower concentrations, BM-MSCs showed notable resistance, with significant cell death observed at only 10 μM, as confirmed by both MTT and Live/Dead assays. These findings suggest that UC2288 may have a concentration-dependent dual effect, where lower concentrations (2.5 µM and below) promote osteogenic differentiation without inducing cytotoxicity in BM-MSCs. The differential sensitivity between BM-MSCs and MDA-MB-231 cells highlights UC2288’s potential as a selective therapeutic agent, enhancing osteogenesis while maintaining viability in stem cell populations.
One striking result observed in gene and protein expression analysis of BM-MSCs with DMEM/F12 revealed that UC2288 interaction at all concentrations increased and did not decrease p21 expression. This is contrary to previously published studies where p21 mRNA levels decreased after interaction with UC2288. 21 When cells are cultured with UC2288 for longer periods of time, a compensatory protective mechanism could be activated where the endoplasmic reticulum increases its function activating adaptive responses 24 causing increase in p21. Similar results were obtained by Sorteberg et al. where they treated neuroblastoma cell lines with 10 µM of UC2288 and determined levels of p21 and p-p21 (Thr145) (p21 that has been localized to the cytoplasm and phosphorylated). They observed an increase in p21 expression but an associated decrease in p-p21, similar to our results. 17
Interestingly, when BM-MSCs were cultured in alpha-MEM, the gene expression pattern following UC2288 treatment differed significantly from that observed in DMEM/F12. Unlike in DMEM/F12, where multiple osteogenic markers (Runx2, Bglap, Col1a1) exhibited increased gene expression, alpha-MEM cultures primarily showed a significant upregulation of Sp7 (Osterix). This suggests that the combination of alpha-MEM and UC2288 enhances osteogenesis through a distinct regulatory pathway compared with UC2288 with DMEM/F12. One possible explanation is that alpha-MEM’s unique composition—higher amino acid, vitamin, and nucleoside content, 25 along with lower glucose levels—creates an environment that promotes early osteogenic commitment via Sp7 activation before substantial upregulation of Runx2 and downstream markers. Additionally, the metabolic differences between these media may influence the effects of UC2288 differently, potentially altering p21’s role in osteogenic differentiation depending on the cellular energy state and stress-related interaction. 26 The absence of substantial changes in other osteogenic genes in alpha-MEM suggests that UC2288’s regulatory effects on p21 and osteogenesis are not solely dependent on media composition but rather on the specific interplay between UC2288 and the metabolic and signaling environment provided by each medium.
This is the first study to assess the potential of UC2288 to increase osteogenic activity in MSCs. Conventionally, inhibition of p21 through UC2288 has been associated with initiating apoptosis in cancer cells. 16,17,27 Yet, in MSCs, UC2288 at lower concentrations did not cause apoptosis but instead increased osteogenic capacity. This suggests that the effects of UC2288, whether promoting differentiation or inducing apoptosis, are both concentration-dependent and cell-type specific. The varying responses across different cell types indicate that the threshold of p21 expression required for these outcomes may differ between cells. We hypothesized that the effects of UC2288 could be mediated through phosphorylation, which we evaluated by western blot. However, the absence of p21 phosphorylation suggests that alternative pathways may be involved. Therefore, future studies will include transcriptomic analyses to further elucidate UC2288’s interactions with various signaling pathways.
Conclusions
UC2288 is a p21 attenuator with selective cytotoxicity against cancer cells that also holds potential for enhancing osteogenic differentiation in BM-MSCs. Our study demonstrates that UC2288 significantly increases the expression of osteogenic genes, particularly Sp7, Runx2, Bglap, and Col1a1, in a medium-dependent manner. In alpha-MEM, Sp7 showed the most pronounced upregulation, while DMEM/F12 led to broader osteogenic gene activation alongside increased p21 expression, suggesting a complex regulatory mechanism. This study represents a critical first step in evaluating the role of UC2288 as a potential osteogenic small molecule that could enhance bone regeneration. While UC2288 alone may initiate osteogenic signaling, its full potential may be realized in combination with an osteogenic environment. Future research will focus on in vivo studies to assess its effects on bone formation, and explore its potential for clinical translation in regenerative medicine and skeletal repair applications.
Footnotes
Acknowledgments
The authors acknowledge the College of Engineering and Applied Sciences (Dr. Andrew Graettinger) for providing equipment used in this study. The authors also thank Angela Schmoldt for aiding in PCR.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the University of Wisconsin, Milwaukee’s Research Foundation (UWMRF) (MIL118990) and NSF I-Corps (2331093). The funding agency was not involved in any aspect of the study. J.S. was supported by SURF Awards through University of Wisconsin, Milwaukee.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.