
Abstract
Neural crest (NC) cells are a transient population of migratory multipotent cells that give rise to a wide variety of derivatives, including neurons, glial cells, Schwann cells, melanocytes, endocrine cells, smooth muscle cells, and the skeletal and connective tissue components of the craniofacial complex. Although the multipotency of NC cells is generally considered to be transient during the early stages of NC formation, accumulating evidence indicates that these cells retain their multipotent characteristics during embryonic migration. Moreover, multipotent NC stem-like cells (NCSCs) persist even within target tissues in the fetal and adult stages. Recent advances in high-throughput and integrative transcriptomic analyses have provided a comprehensive understanding of the genetic and molecular profiles of NC cells. These studies have revealed that NC cells exhibit remarkable transcriptional diversity and simultaneously express genes associated with pluripotency, lineage specification, and differentiation, underscoring their intrinsic plasticity. The multipotency and plasticity of NC cells and NCSCs thus represent a compelling field of study with significant implications for developmental biology and regenerative medicine. In this review, we summarize advances in research on NC cells and multipotent NCSCs as well as the transcription factors that maintain the multipotency of NC cells.
Introduction
Neural crest (NC) cells are migratory multipotent cells that give rise to diverse derivatives. 1 Based on these multipotent characteristics, NC cells play critical roles in mammalian embryogenesis. During their migration, NC cells respond to a repertoire of factors, such as bone morphogenetic proteins, epidermal growth factor (EGF), Wnt proteins, stem cell factor, and endothelin 3, 2 and give rise to a large array of cell types, including neurons and glial cells of the peripheral sensory and autonomic ganglia, enteric neurons, Schwann cells, chromaffin cells of the adrenal medulla, melanocytes, smooth muscle cells, and skeletal and connective tissue cells of the craniofacial complex. 1 Due to their high migratory ability and multipotency, NC cell abnormalities have been implicated in various diseases, including Waardenburg’s syndrome, Hirschsprung’s disease, CHARGE syndrome, and DiGeorge’s syndrome. 3
NC cells are generated at the border regions of neural folds during neurulation. 1 They then delaminate from the neuroepithelium and migrate in various streams along with the axis from anterior to posterior within the embryo. NC cells start to migrate in the cephalic region in the embryo; the cephalic NC cells differentiate into cartilage, bone, muscle, connective tissues for the skull and facial structures, and facial innervations. 1 Following the stream, NC cell migration is initiated vagal and trunk regions; the vagal NC cells give rise to the aortic outflow tract of the heart and to the enteric nervous system with connective tissue and neuronal and glial cells, whereas trunk NC cells generate the most posterior region of the enteric nervous system, dorsal root ganglia (DRG), and the adrenal gland and melanocytes. Due to their multipotency, NC cells are often referred to as the fourth germ layer. 3
The multipotency of NC cells was previously considered to be lost after their delamination from the neural tube and their migration in the embryo; however, cells derived from NC cell, such as melanoblasts and Schwann cells precursors, also exhibit a multipotent cell fate.4–7 Furthermore, target tissues into which NC cells migrate and differentiate, such as the peripheral nerve, gut, skin, and carotid bodies (CBs), have been shown to maintain multipotent NC stem-like cells (NCSCs).8–11 Recent analyses for large-scale information of NC cells provided novel insights into NC cell differentiation; NC cells are not fate-restricted, as previously considered, but rather maintain high plasticity. We herein review and briefly discuss the findings on the plasticity of NC cells, multipotent NCSCs in tissue, and transcription factors relevant to NC cell multipotency.
NC Cell Plasticity: Recent Findings on NC Cell Generation and Differentiation with Single-Cell Transcriptomics
Research on the differentiation potential of NC cells began with studies using chicken–quail chimeric embryos. In these early in vivo analyses, NC cells were thought to lose their multipotency before or soon after delaminating from the neural tube. Most of the initial NC cell population appeared to be lineage-restricted, producing clones of only a single-cell type 12 or, at most, bipotent precursors. 13 In contrast, in vitro culture experiments of migrating NC cells revealed the presence of multipotent progenitor cells in the trunk NC of mice and rats.14–17 Furthermore, NC cells have been shown to retain pluripotent characteristics even after migration to target tissues in embryos.8,18 It remains unclear whether these apparently contradictory results arise from the NC population being highly heterogeneous or from NC cells themselves possessing intrinsic plasticity. However, recent advances in comprehensive analyses and technologies for combining large-scale information have clarified the genetic profiles of NC cells, providing new insights into this issue.
Lignell et al. combined multiplex single-molecule fluorescence in situ hybridization with cell segmentation based on machine learning algorithms to analyze the NC cell at the single-cell resolution in in vivo neural tube in Hamburger and Hamilton stage 9 chick embryos. 19 They demonstrated that premigratory NC cells localized at the midline of the dorsal neural tube coexpressed marker of NC identity, pluripotency, and differentiation, whereas migratory NC cells expressed subsets of NC markers along with the differentiation marker, such as epidermal marker Krt19 (Fig. 1). Soldatov et al. performed single-cell RNA sequencing analysis to elucidate the process of NC cell-fate determination. 20 By isolating NC cells from Wnt1-Cre/R26R-Tomato and Sox10-CreERT/R26R-Tomato mice across embryonic stages from E9.5 to fetal stages, they showed that NC cells competitively activated gene modules related to fate determination prior to branching into specific lineages. For example, in the delaminated NC cell population, epithelial to mesenchymal transition-related genes and NC cell-specific genes were upregulated, whereas some neuroectoderm boundary-specific genes remained expressed (Fig. 1). Qiu et al. profiled the transcriptional state of single cells from mouse embryos from E3.5 to E13.5, annotated them by cell type, and systematically integrated these data to construct a roadmap of the trajectory taken by each cell in the embryo. 21 This analysis reconstructed mouse embryogenesis based on transcriptional similarities and key transcription factors upregulated at lineage bifurcations. In this analysis, NC-fated cells that diverged from the neuroectoderm highly expressed Sox9, Msx1, and Id2, genes associated with NC cell multipotency22–24 (Fig. 1). Zalc et al. revealed that mouse cranial NC cells are transcriptionally diversity population during delamination and ultimately reactivate epiblast pluripotent factor Oct4 in combination with single-cell RNA-seq, lineage tracing, loss-of-function screening, and epigenomic profiling 25 (Fig. 1). Similarly, Pajanoja et al. reported that both chick and mouse cranial NC cells express pluripotency factors. By tracing the transcriptional changes from gastrulation to neurulation using single-cell multiplex spatial transcriptomics and bulk RNA sequencing, they showed that pluripotency factors maintained in the pan-ectoderm during late neurulation persist within NC cells 26 (Fig. 1). Although transcriptional profiles of NC cells vary depending on isolation methods and spatiotemporal context, these findings collectively indicate that many NC cells simultaneously express genes related to pluripotency, generation, and differentiation, suggesting that NC cells themselves demonstrate transcriptional diversity (Fig. 1). These transcriptionally diversity were shown in other vertebrate NC cells by a single-cell RNA sequencing analysis of zebrafish premigratory NC cells.27,28 This transcriptional diversity may lead NC cells itself to a highly plastic state. Thus, due to the plasticity of NC cells, chick trunk NC cells forcibly expressing Twist1, a cranial NC gene, were shown to differentiate into the cranial NC cell type lineage, mesenchymal lineage. Conversely, Twist1 knockdown in cranial NC cells induced differentiation into the trunk NC cell type derivatives such as glial and neuronal cells. 20 These comprehensive analyses underscore the broad plasticity inherent in NC cells themselves.
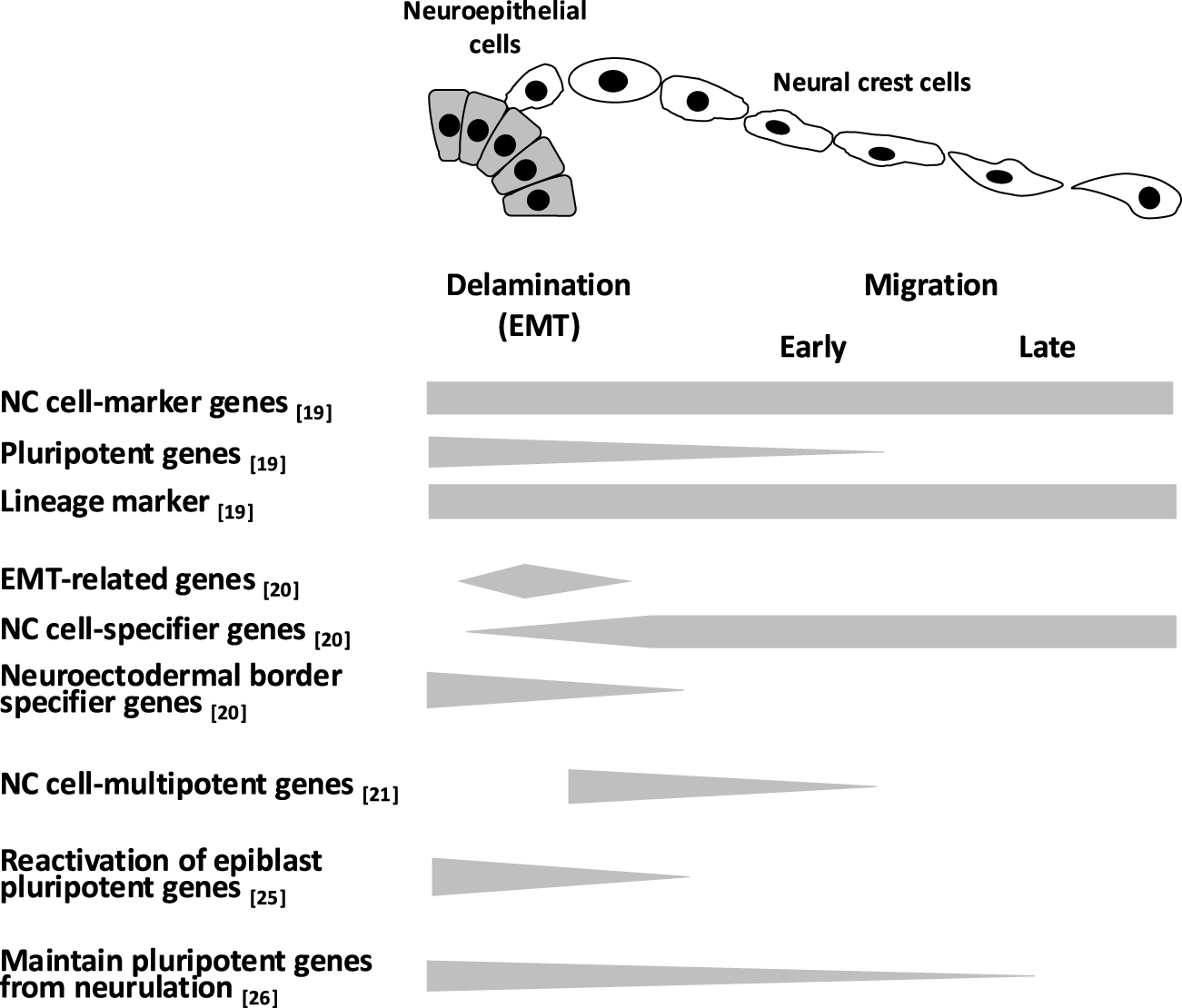
Gene expression during the generation and migration of NC cells. Although various transcriptional profiles of NC cells have been reported depending on the isolation method, spatiotemporal differences, and analytical approaches, NC cells are thought to simultaneously express genes involved in pluripotency, specification, and differentiation. This transcriptional diversity may contribute to the high plasticity of NC cells. Gray bars indicate gene expression levels. NC, neural crest.
Multipotent NCSCs in Tissues
Recent advances in gene technologies have revealed the presence of postmigratory multipotent NCSCs in tissue. NCSCs were detected in the tissues of NC-labeled transgenic mice, such as P0-promoter Cre/Floxed-EGFP and Wnt1-promoter Cre/Floxed-EGFP or in self-renewing sphere-forming cultures.
It currently remains unclear whether these NCSCs in tissues are derived from embryonic NCs or are generated by the dedifferentiation of NC cell derivatives (Fig. 2). However, Schwann cell precursors (SCPs), which have multiple fates, were recently suggested to represent a source of adult NCSCs residing in numerous tissues (Fig. 2). SCPs correspond to intermediate cells in the early development stage between NC cells and immature Schwann cells, migrate to distal regions with developing nerves in growing embryos, and maintain multipotency before the onset of myelination.5,29,30
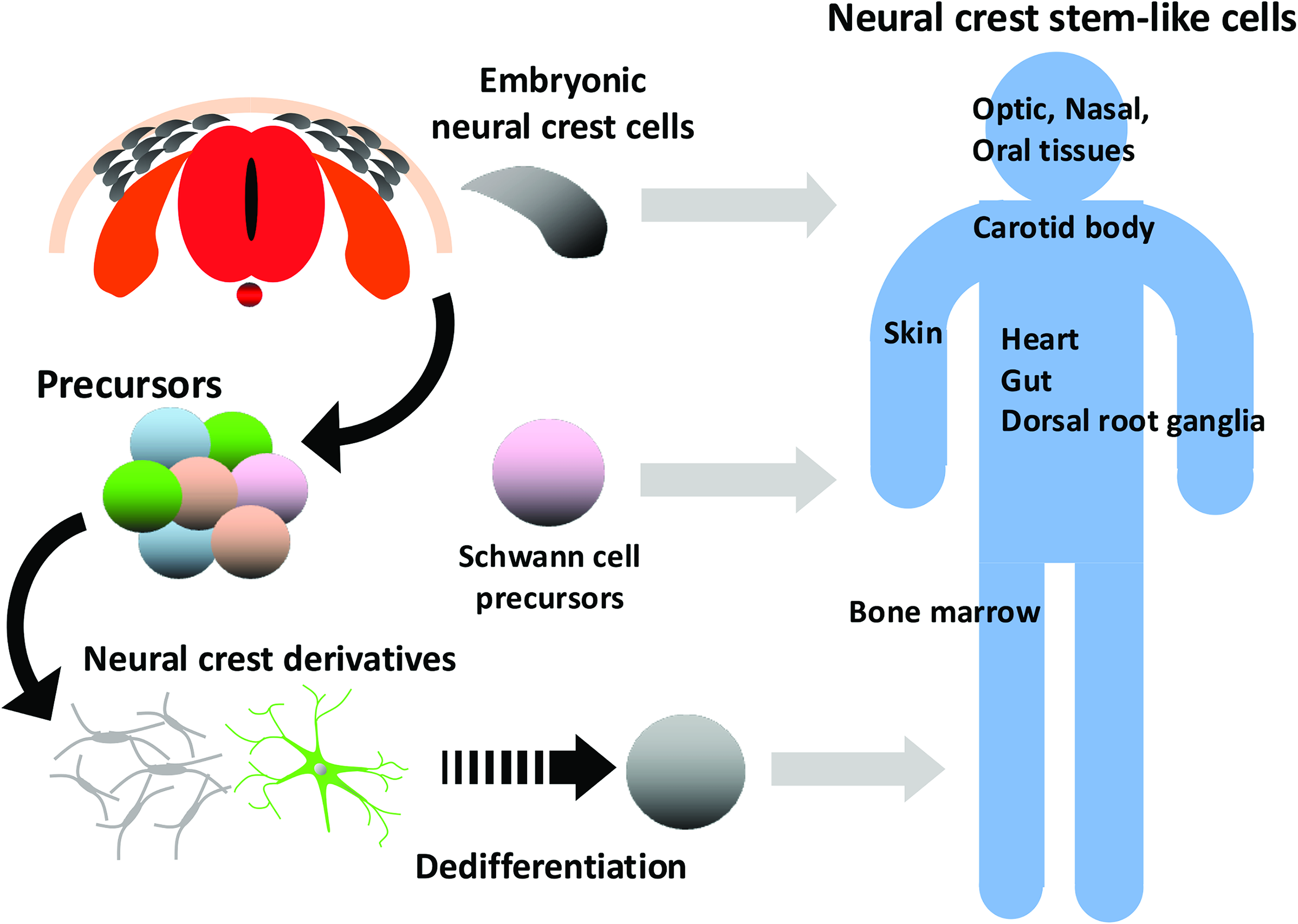
Origin of NCSCs in tissues. Although multipotent NC cells are a transient population that give rise to various derivatives in embryogenesis, multipotent NCSCs are present in some tissues to which NC cells migrate. Three putative origins of NCSCs in tissues have been proposed, maintained multipotent embryonic NC cells, Schwann cell precursors that attach to nerves and have multiple fates, and dedifferentiation from NC cell derivatives. NCSC, neural crest stem-like cell.
SCPs differentiate from migrating NC cells associated with growing nerves in E11.5 embryos. SCPs migrate with nerves, reach distal parts in the embryo, and those that detach from the nerve differentiate into melanocytes, parasympathetic nerves, sympathetic nerves, and enteric nerves, 31 chromaffin cells, and mesenchymal cells, which differentiate further into chondrocytes and mature osteocytes, whereas those that remain attached to the nerve differentiate into Schwann cells.5,32 A RNA-Seq analysis of SCPs recently showed that they have a similar transcription factor profile to that of migratory NC cells, for example, the expression of Sox9, Sox10, FoxD3, Tfap2a/b, and Ets1, which are linked to stem cell properties. 33 In addition to the multipotency of SCPs in early development, some SCPs remain and maintain multipotency in the postnatal stages. For example, in adult mouse incisors, which continuously grow, SCPs associated with nerves contribute to the dental mesenchymal stem cell pool, from which dental pulp and odontoblasts are generated. 34 These findings suggest that SCPs utilize peripheral nerve networks for distribution in the tissues of embryos and as niches that support the maintenance of their pluripotency. The maintenance of multipotency by SCPs in adulthood may represent a source of the adult NCSCs residing in numerous tissues. 33
In contrast, the reemergence of NCSC-like states in vivo suggests that tissue NCSCs arise from the dedifferentiation of NC derivatives. NC-derived peripheral glial cells dedifferentiate by downregulating glial lineage markers after injury and acquire an NCSC-like state with increased proliferation and the upregulated expression of p75. 35 A subset of melanoma cells reexpress CD271 (p75NTR) with a gene expression profile that is reminiscent of NCSCs, such as Sox10.36,37 Schwann cells were previously shown to dedifferentiate into their immature states in response to injury and contribute to wound healing.38–40 These findings suggest that NC-derived cells, the lineage of which is fully restricted, unexpectedly dedifferentiate into multipotent NCSCs in response to changes in their microenvironment.
In each section of this chapter, we summarize the current status of tissue NCSCs, as shown in Table 1.
Isolation and Differentiation of Tissue-Derived Neural Crest Stem-Like Cells
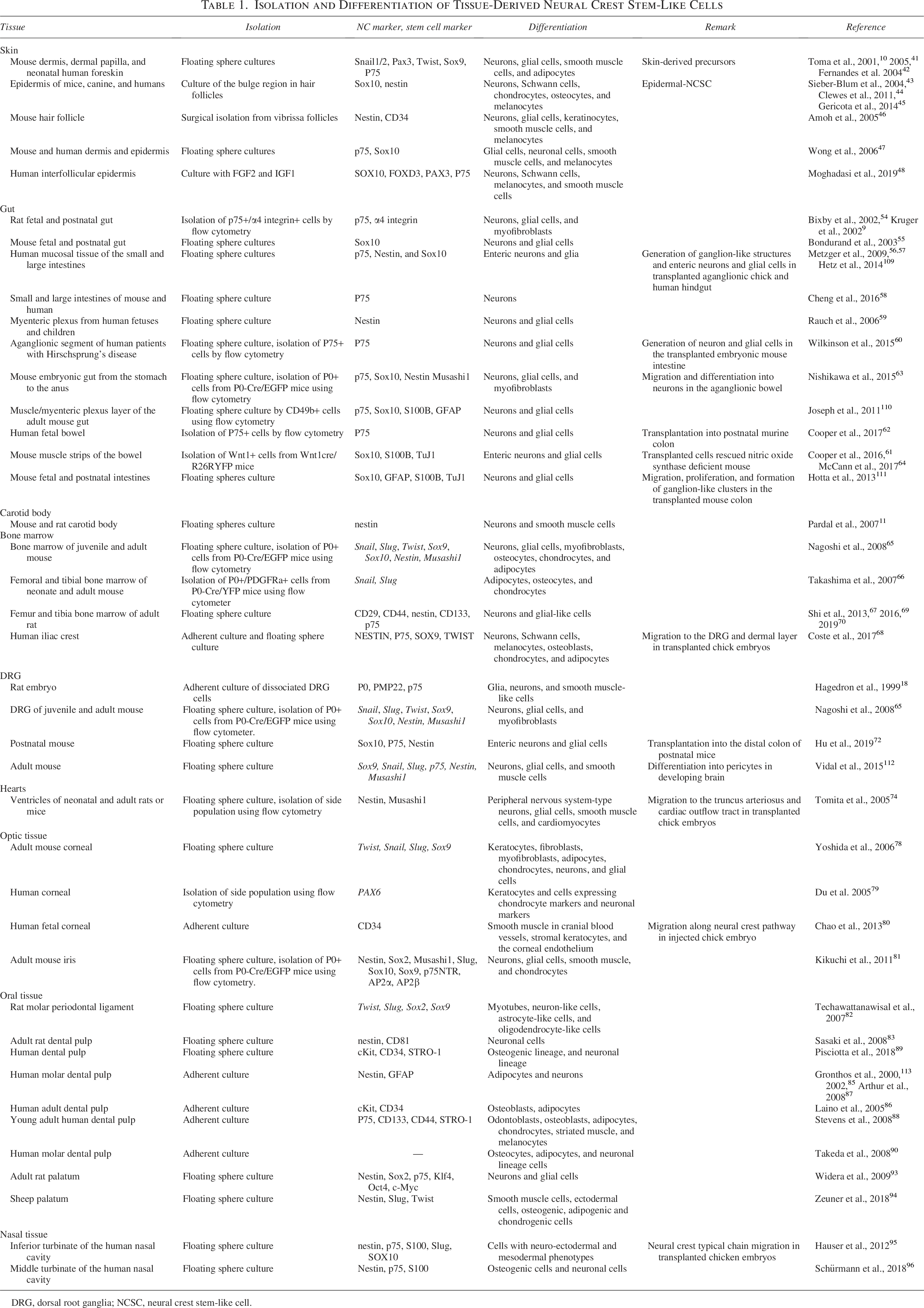
DRG, dorsal root ganglia; NCSC, neural crest stem-like cell.
NCSCs in skin
NCSCs were described in the mouse dermis, the dermal papilla of whisker hair follicles in facial skin, and neonatal human foreskin as skin-derived precursors.10,41,42 Previous studies also reported the existence of multipotent NCSCs in skin; epidermal NCSCs (EPI-NCSCs) in the epidermis of mice, canines, and humans; NCSCs in human interfollicular keratinocytes; nestin and stem cell marker CD34-expressing cells in the bulge region of hair follicles; and sphere-forming cells in adult mice and human skin.43–48
The therapeutic potential of skin NCSCs is enhanced by easy accessibility to skin for their isolation. EPI-NCSCs transplanted into injured rat spinal cords were shown to differentiate into glial cells and promote the release of the neurotrophic factors, brain derived growth factor (BDNF) and neutrophin-3, which ameliorated the injury. 49 Furthermore, grafted EPI-NCSCs with olfactory ensheathing cells in injured peripheral nerves upregulated the expression of BDNF and nerve growth factor, which attenuated the injury. 50 In ischemic stroke rats, treatments with skin-NCSCs and its conditioned medium reduced infarct sizes and mitigated neurological impairments.51,52 Moreover, in facial nerve defects, exosomes from hair follicle EPI-NCSCs promoted cellular nerve allografts. 53 Collectively, these findings show that skin NCSCs contribute to the repair damaged tissues, not only through their differentiation but also via factors derived from the NCSCs.
Enteric NCSCs
Enteric NCSCs were initially identified in the nervous system in the wall of the gut during fetal development 54 and were shown to persist throughout adulthood.9,55 Since then, enteric enteric NCSCs have been isolated from the tissue or mucosal tissue of the small and large intestines56,57,58,59 and even from the aganglionic segments from human patients with Hirschsprung’s disease. 60
Enteric NCSCs have been shown to form spheres with self-renewal potency that proliferate, migrate, and differentiate into neurons and glial cells in the transplanted embryonic mouse intestine and functionally integrate with the host neuromusculature.61,62 Transplanted Enteric NCSCs were found to migrate and differentiate into neurons in an aganglionic bowel 63 and also restored gut motility in a mouse model of human enteric neuropathy. 64 Although many transplantation studies have demonstrated that enteric NCSCs are effective for treating enteric nervous disorder models, further studies are needed before their application to the treatment of enteric neuropathy.
NCSCs in CBs
NCSCs have been detected in CB, which is an oxygen-sensing organ at the bifurcation of the carotid artery. CB contains clusters of O2-sensitive, neuron-like glomus cells enveloped by glial cells originating from NC cells. The CB-NCSCs form spheres with self-renewal and differentiate into dopaminergic neurons and smooth muscle cells. 11 However, the use of CB-NCSCs in transplantation is limited due to the small amount of available tissue.
NCSCs in bone marrow (BM)
NCSCs have been identified in BM, where NC-derived cells contribute to the formation and maintenance of the hematopoietic stem cell niche.65–68 BM-NCSCs represent an attractive source for autologous cell therapies, as BM can be readily accessed for harvesting from patients. Several studies have provided compelling evidence that BM-NCSCs may have regenerative potential, such as in the treatment of peripheral nerve injuries.69,70 However, expanded mouse BM-NCSCs have been reported to form tumors following in vivo transplantation. 71 Therefore, before clinical application, it is essential to rigorously assess whether the biological characteristics of NCSCs are suitable and safe for therapeutic use. 71
NCSCs in DRG
NCSCs in DRG were initially characterized as sphere-forming cells expressing two peripheral myelin proteins: P0 and PMP22. 18 A previous study demonstrated that NCSCs from postnatal DRG survived and generated neuron–muscle connections when transplanted into the distal colon of postnatal mice. 72 Similar to CB-NCSCs, the use of DRG-NCSCs in transplantation is limited due to the small amount of available tissue.
Cardiac NCSCs
Cardiac NCSCs have also been detected in the side population (SP) cells of the heart. SP cells are isolated by their ability to actively efflux the fluorescent DNA binding dye Hoechst 33342 using a flow cytometric analysis and are generally regarded as tissue-specific progenitor cells. 73 Cardiac NCSCs generate spheres and differentiate into NC-derived cell types, including neurons and glial cells in the peripheral nervous system and smooth muscle cells in addition to cardiomyocytes. 74 Sphere-derived cells have also been shown to migrate and integrate into NC-targeted tissues, DRG, spinal nerves, and aortic smooth muscle cells in transplanted chick embryos. 714 In a myocardial infarction model using P0-promoter Cre/Floxed-EGFP mice, EGFP+ NCSCs accumulated at the ischemic border zone area, suggesting that persistent NCSCs in the heart contributed to the regeneration of cardiomyocytes. 75
NCSCs in optic tissues (corneal and iris)
Corneal keratocytes, which form the stroma of the cornea, continually heal the cornea throughout the lifespan and their developmental origin was identified as cranial NC cells.76,77 A previous study detected multipotent keratocyte precursor cells in the adult mouse cornea (cornea-derived precursors, COPs). 78 In humans, multipotent keratocyte progenitor cells were isolated as SP cells by flow cytometric cell sorting. 79 Although limited information is currently available, SP cells are known to differentiate into cells expressing chondrocyte markers and neuronal markers at the genetic level. 79 Another study demonstrated that human fetal keratocytes injected into avian embryos migrated along the NC pathway, differentiated into NC derivatives, including smooth muscle in cranial blood vessels, stromal keratocytes, and the corneal endothelium. 80 These findings suggest that human keratocytes also contain multipotent NCSCs that differentiate into NC-derived cells.
The iris also possesses multipotent NCSCs. Iris cells isolated from P0-Cre/Floxed-EGFP transgenic mice formed sphere structures that differentiated into smooth muscle and cartilage. 81 These multipotent keratocytes and iris cells have great potential in regenerative therapy for eye diseases.
NCSCs in oral tissues (periodontal ligament, dental pulp, and palate)
NCSCs were identified as the serially sphere-forming cells with bFGF in the periodontal ligament, a soft connective tissue embedded between the tooth root and the alveolar bone socket, in the dental pulp, the connective tissue at the center of teeth encased in mineralized dentin, and in the palate.82–84
Human dental pulp stem cells differentiate into NC cell derivatives: chondrocytes, osteocytes, adipocytes, odontoblasts, myocytes, and neural-like cells in vitro85–90 and, furthermore, into other types of cells in vitro, such as islet cell aggregates 91 and to insulin producing cells, 92 demonstrating their potential for clinical use in regenerative therapies.
NCSCs have also been identified in the palate of adult rats using sphere-forming cultures and differentiated into neurons and glial cells. 93 The sheep palate was also shown to contain NCSCs. 94 However, it remains unknown whether they can differentiate into other lineages other than neurons and glial cells, or if they have self-renewal potency.
Nasal NCSCs
The nasal inferior turbinate contains NCSCs. These NCSCs formed the sphere structures in the presence with bFGF and EGF, differentiated into cell with the neuro-ectodermal phenotype, and migrated along the NC cell-specific pathway in avian embryos. 95 NCSCs were also identified in the middle turbinate of the human nasal cavity. They differentiated into osteogenic and neuronal cell types similar to the cells of the inferior turbinate. 96
Transcription Factors that Control the Multipotency of NC Cells
Numerous studies have revealed the individual transcription factors involved in induction, specification, migration, and differentiation of NC cell and have demonstrated the involvement of various transcription factors in the multipotency of NC cells. Among these transcription factors, FoxD3 and Sox10 are known to be involved in the multipotency of NC cell. FoxD3, a winged-helix, forkhead transcription factor, is one of the early NC cell lineage molecular markers. 97 The ectopic expression of Foxd3 by migrating NC cells retains an undifferentiated state, and the NC cell-specific deletion of Foxd3 resulted in the aberrant differentiation of NC cells, such as the loss of neural derivatives in the cranial, vagal, and cardiac NC cells, suggesting the contribution of FoxD3 to NC self-renewal, multipotency, and the control of differentiation.98,99 Sox10 is an NC cell specifier-like FoxD3, is expressed in premigrating NC cells, maintained in migratory NC cells, and has been implicated in the glial and melanocytic differentiation of NC cells. 100 The constitutive expression of Sox10 inhibits the differentiation of NC cells into neurons, 101 and multipotent postmigratory NC cells were shown to undergo cell death in the Sox10–/– embryos before maturation, 102 indicating that Sox10 is essential for the maintenance and survival of multipotent NC cells. Therefore, conventional studies revealed that several transcription factors, including FoxD3 and Sox10, are involved in the multipotency of NC cells. However, since the multipotency of NC cells is considered to be controlled not by a single or a few transcription factors but rather by multiple transcription factors working in coordination with each other (Fig. 3), the molecular mechanisms that maintain and control the multipotency of NC cells remain unclear. Moreover, multipotency may be a transient state exhibiting transcriptional heterogeneity that is related to cell fate decisions during the developmental process 20 ; therefore, difficulties are associated with identifying molecule clusters that are critical for the control of NC cell multipotency using a conventional “point analysis.” Recent technologies for combining information obtained from conventional analyses, such as gene arrays, lineage tracing, and single-cell RNA-seq, provide “continuous information” and have achieved breakthroughs.
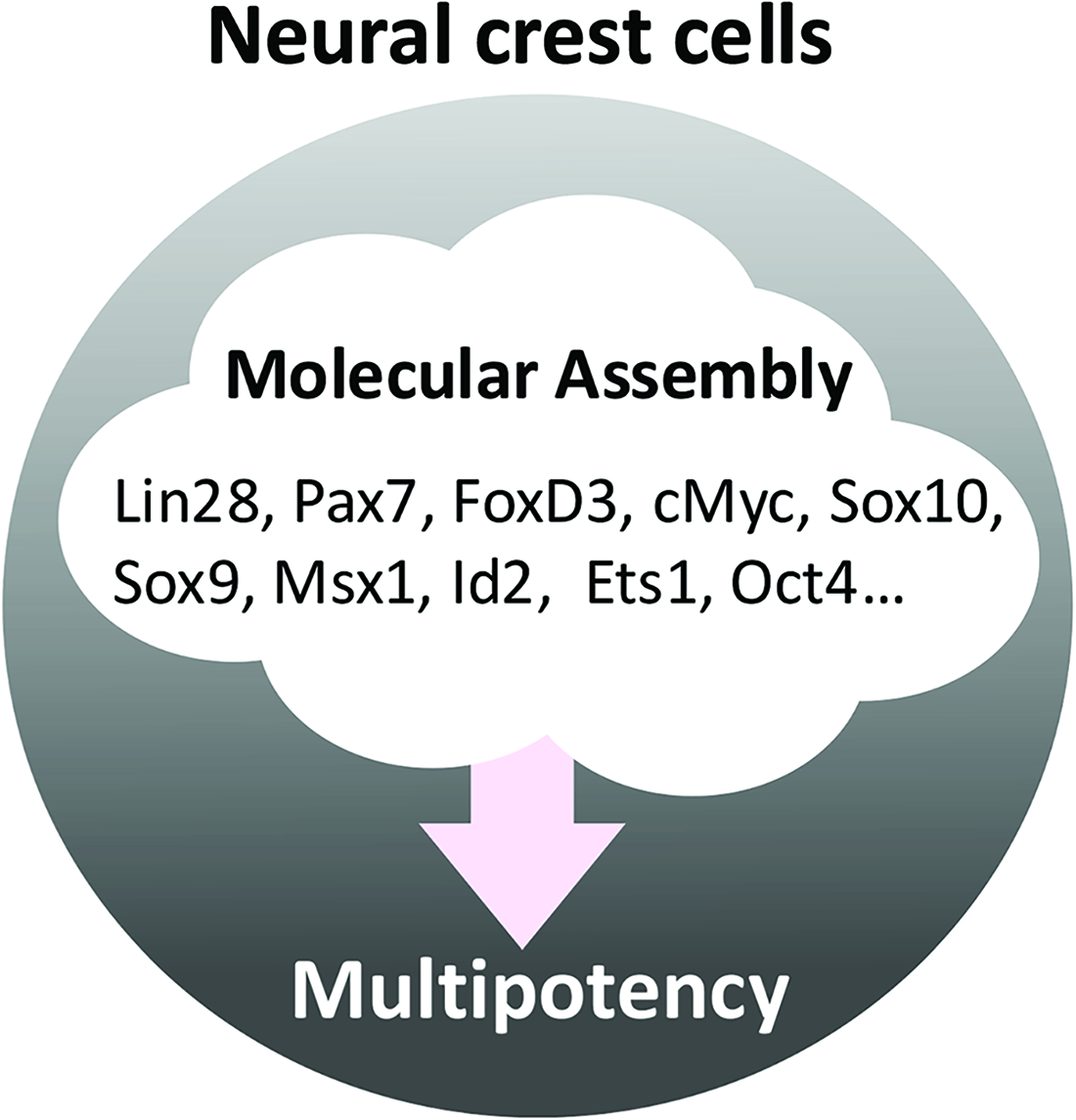
Multipotency of NC cells. Since the multipotency of NC cells is considered to be controlled by multiple transcription factors, difficulties are associated with clarifying the roles of transcription factors.
Simões-Costa et al. attempted to elucidate the genetic regulatory network (GRN) that governs NC development and differentiation with BioTapestry, a pathway modeling tool that integrates data from different sources, including microarrays, ChIP-seq, and RNA-seq experiments, and facilitates the visualization of large numbers of linked genes. 100 By using a GRN analysis, Simões-Costa et al. revealed the involvement of the RNA-binding protein Lin28a in multipotency in the cranial NC. 103 In early chick NC cells, Lin28a transcription was activated by Wnt signaling and maintained NC stemness by inhibiting let-7 miRNA, which silenced multiple components consisted of Pax7, FoxD3, cMyc, and Sox10. 103
With a roadmap of the cellular trajectories of mouse embryogenesis described in Chapter 1, Qiu et al. identified key transcription factors that were upregulated at the divergence of the NC cell lineage from the neuroectoderm. 21 The key transcription factors involved were Sox9, Msx1, and Id2, each of which were relevant to NC cell multipotency.22–24 Moreover, the forced expression of Sox9 and Msx1 directly converted somatic cells into NC-like cells, 104 and the ectopic expression of Id2 was shown to convert ectodermal cells to a NC fate, 24 indicating their potential to confer NC multipotent characteristics.
In Xenopus, NC cells share pluripotency regulatory factors, such as FoxD3, Ets1 and Snail1, in blastula animal pole cells, 105 suggesting that NC cells maintain pluripotency by continuing to express or reactivating these transcription factors. This was confirmed in combination with experimental information using mouse and chick NC cells by Lignell et al., Zalc et al., and Pajanoja et al. as shown in Chapter 119,25,26 (Fig. 1). Furthermore, Lignell et al. found that NC cells simultaneously expressed pluripotency and lineage markers, 19 suggesting the plasticity and multiple fates of NC cells, which may be the “pluripotency” of NC cells. Therefore, by using a comprehensive analysis of multiple information sources for NC cell, a more detailed understanding of the transcription factors involved in their pluripotency is being obtained.
The forced expression of transcription factors involved in NC development has potential in the direct conversion of some somatic cells into NC cells. Since the transcription factors used for direct conversion confer NC cell pluripotency to somatic cells, they may be relevant to the pluripotency of NC cells. FoxD3, Sox10, and Msx1 strongly induce the direct conversion of somatic cells into NC-like cells through their forced expression.104,106–108 These NC-like cells were previously shown to undergo chondrogenic, adipogenic, osteogenic, neurogenic, and melanogenic differentiation, 106 differentiated into neuronal cell, glial cells, chondrocytes, adipocytes, osteocytes, and smooth muscle cells104,107 and showed potency for neuronal differentiation, adipogenesis, and smooth muscle cell differentiation, 108 respectively. These finding revealed the involvement of FoxD3, Sox10, and Msx1 in conferring the potency for differentiation into these cells.
Concluding Remarks
We herein focused on the plasticity of NC cells, multipotent NCSCs in tissues, and transcription factors relevant to their multipotency. Recent advances in comprehensive single-cell-level analyses revealed various properties of NC cells, including their differentiation and multipotency. However, there is still much to be revealed about NC cells.
The origin of tissue NCSCs remains obscure. In this review, we proposed SCPs and dedifferentiation from NC cell derivatives as the possible origin of tissues NCSCs (Fig. 2). Furthermore, it is possible that multipotent embryonic NC cells are also maintained in adult tissue (Fig. 2). One possibility is that a specialized cell population is present and maintained in vivo as NCSCs from the early induction stage of NC cells, whereas another possibility is that migratory NC cells inadvertently reach and reside in stem cell niches in each tissue.
In addition, the leading machinery for maintaining NC cell multipotency remains unclear. Since the multiple transcription factors may be working in combination with each other to maintain the multipotency of NC cells (Fig. 3), a conventional single-molecule level analysis is not possible. Therefore, further advances in comprehensive analyses or technologies to combine large-scale information are expected and will provide insights on this issue.
NCSCs represent attractive candidates for cell therapy. However, current NCSC isolation, expansion, and differentiation protocols need to be improved in order to facilitate the therapeutic applications of NCSCs to clinical settings. Direct reprogramming offers another promising alternative to obtain NCSCs; however, converted NCSCs may display limited multipotency, low conversion efficiency, and the risk of tumorigenesis due to the forced expression of transcription factors.104,106,107 A detailed examination of the generation, regulation, and maintenance of stem cell potential in NC cells is important for understanding not only embryonic development but also stem cell biology and their therapeutic application to regenerative medicine. The latest comprehensive omics analysis is expected to enable detailed studies on NC cell stemness, which will lead to further advances in NC cell research.
Footnotes
Acknowledgments
The authors thank the other members of their laboratory for their discussions and critical reading of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Infomation
This study was supported by a Grant from the program Grants-in-Aid for Scientific Research (C) by the Japan Society for the Promotion for Science (JP22K06808, JP25K10143).