
Abstract
Mesenchymal stromal cell-derived extracellular vesicles (MSC-EVs) are emerging as potent cell-free mediators of tissue repair, whose composition and function can be tuned by the cellular microenvironment. Although inflammatory cues modulate mesenchymal stromal cell (MSC) behavior, how defined preconditioning strategies program extracellular vesicle (EV) functional outputs remains incompletely understood. Here, we systematically evaluated how priming bone marrow-derived MSCs with interferon-gamma/tumor necrosis factor-alpha (I/T) or lipopolysaccharide (LPS) generates EV populations with distinct immunomodulatory and regenerative properties. Using a murine full-thickness wound model, we performed an integrative analysis of biodistribution, immune response, and extracellular matrix (ECM) remodeling, complemented by single-cell, transcriptomic, and proteomic profiling. All EV populations were retained at the wound site following subcutaneous delivery and supported wound contraction; however, they drove distinct, treatment-specific repair trajectories. I/T-EVs promoted a coordinated regenerative response characterized by balanced macrophage (MΦ) activation, controlled immune modulation, and efficient resolution of inflammation, resulting in organized ECM remodeling. In contrast, LPS-EVs induced a more pro-inflammatory response, accelerating wound contraction and promoting compensatory matrix stiffening with reduced structural coordination. Control EVs primarily facilitated early immune resolution with limited induction of regenerative remodeling pathways. Proteomic profiling of EVs identified enrichment of proteins associated with insulin-like growth factor signaling, MΦ recruitment, and ECM remodeling, consistent with in vivo protein expression patterns and linking EV cargo to functional outcomes. These findings demonstrate that preconditioning does not uniformly enhance EV efficacy but instead selectively programs distinct MSC-EV functional states, establishing EV preconditioning as a tunable strategy for engineering cell-free therapeutics with predictable and context-specific therapeutic outcomes.
Keywords
Introduction
Mesenchymal stromal cells (MSCs) are characterized by their capacity for self-renewal and multipotency. Clinically, they have been explored for the treatment of injuries across various organs, including bone, lungs, and liver, as well as for restoring homeostasis in tissues with limited regenerative potential. 1 MSCs have also been employed as adjuvant cells to promote angiogenesis, mitigate inflammation, and enhance engraftment in cell and organ transplantation.2,3 Owing to their ability to modulate both resident and circulating immune cells, 4 MSCs are often administered during the inflammatory phase of injury. Following administration, MSCs dynamically reprogram their cellular activities in response to biochemical cues, thereby enhancing their tissue-targeting capacity and therapeutic potential. 5 Preconditioning through environmental stressors further alters gene expression and molecular profiles, 6 improving immunomodulation, extracellular matrix (ECM) remodeling, and vascularization.7,8
MSCs influence tissue repair through their extracellular vesicles (EVs), which act as key mediators of intercellular communication. While MSCs respond dynamically to the in vivo microenvironment, EVs represent a stable snapshot of the cellular state at release, enabling controlled delivery of bioactive signals independent of subsequent environmental reprogramming. 9 Unlike their parent MSCs, EVs are minimally immunogenic and remain functionally consistent after isolation, allowing them to deliver therapeutic cargo effectively, and circumventing limitations of cell-based therapies such as immune rejection and variable responsiveness. 10 Their nanoscale size and intrinsic targeting capabilities allow them to cross biological barriers while preserving therapeutic efficacy. 11 These properties position EVs as scalable, stable, and reconfigurable cell-free therapeutics. 12
The bioactive cargo of MSC-derived EVs (MSC-EVs), including coding and noncoding RNAs, proteins, and lipids, is influenced by environmental conditions such as hypoxia,13,14 glucose deprivation, 15 and exposure to pro-inflammatory cytokines [ie, interleukin-1β (IL-1β), tumor necrosis factor-alpha (TNF-α), interferon-gamma (IFN-γ), and bacterial lipopolysaccharide (LPS)]. 16 LPS-conditioned MSC-EVs promote immunotolerance and reduce inflammation,17,18 conferring protection against septic liver injury 19 and myocardial infarction. 20 Similarly, IL-1β-licensed MSC-EVs exert protective effects against sepsis and acute lung injury.21,22 Preconditioning with TNF-α and IFN-γ enhances the anti-fibrotic,23–26 anti-inflammatory,4,27 angiogenic, 28 and cell proliferative 29 properties of MSC-EVs. Across these conditions, macrophages (MΦs) consistently emerge as primary targets, with EVs promoting the transition from pro-inflammatory (M1) to anti-inflammatory (M2) phenotypes. 30
Despite these advances, a comprehensive understanding of how distinct inflammatory licensing cues program EV functional states and downstream regenerative trajectories remains limited. In particular, the effects of preconditioning on EV biodistribution, tissue retention, and the coordination of molecular and cellular responses in vivo are not fully defined.
In this study, we systematically evaluated how bone marrow-derived MSC (BM-MSC) preconditioning under physiological and inflammatory conditions modulates EV function in vivo. BM-MSCs are among the most extensively characterized MSC populations 31 and are widely used in EV studies. 32 We employed IFN-γ/TNF-α (I/T) to model injury-induced inflammation 33 and LPS to simulate bacterial infection, 34 generating EVs that were directly compared to unprimed control (CTRL) group. EVs were characterized for physicochemical and molecular properties, and biodistribution was assessed using an in vivo imaging system (IVIS) at 1 and 3 days post-subcutaneous (SubQ) administration in healthy mice. Regenerative outcomes were evaluated in a full-thickness wound model, with EV delivery at wound margins and healing monitored over 14 days. The inflammatory microenvironment was profiled at molecular and single-cell resolution to capture immune dynamics and ECM remodeling. By integrating biodistribution, EV cargo proteomics, and in vivo functional analyses, this study defines how MSC preconditioning programs EV-mediated tissue repair, providing a mechanistic framework for the development of targeted, cell-free regenerative therapies.
Methods
Cell culture
Human adult BM-MSCs from a single healthy donor were purchased from American Type Culture Collection (ATCC; PCS-500-012) and cultured at 37°C with 5% CO2. Cells were maintained in high-glucose Dulbecco’s modified Eagle medium supplemented with 10% exosome-depleted fetal bovine serum, 1 mM
EV enrichment and characterization
Cells used for EV production were maintained within the same passage range (P3–P5) and at ∼80% confluence to minimize variability. Conditioned media were centrifuged at 2,300g for 30 min, filtered through 0.22-μm membranes, and concentrated using Amicon® Ultra-15 centrifugal filters (Millipore). EVs were enriched using a polyethylene-glycol-based precipitation method (Total Exosome Isolation Reagent, Invitrogen), followed by centrifugation at 10,000g for 1 h at 4°C. Pellets were resuspended in filtered phosphate-buffered saline (PBS) and stored at −80°C. All EV preparations were processed in parallel using identical workflows to ensure consistency across conditions. EV characterization was performed in accordance with MISEV2023 guidelines. 35 Nanoparticle Tracking Analysis (NTA, NS3000 Nanosight System, Malvern) was used to determine EV size and concentration. Samples were diluted 1:100 in PBS, and five 60-s videos were acquired per sample. Western blotting confirmed EV markers (TSG101, CD63, CD9, and Alix; Proteintech). EV morphology was assessed by cryo-transmission electron microscopy (Cryo-TEM). Quantifoil R2/1, 200-mesh Cu holey carbon grids were glow-discharged for 20 s using a Pelco easiGlow (Ted Pella Inc). Vitrification was performed with a Vitrobot Mark IV (FEI) at 20°C and 100% humidity. Each grid was treated with 3.5 μL of sample solution, blotted for 2 s, and immediately plunged into liquid ethane for preservation. EV images were collected at 100,000×, and scale bars were set to 50 nm.
EV proteomic analysis
Proteins were isolated from EV-enriched preparations and analyzed by nanoLC-MS/MS (System Biosciences, USA). Samples were processed using SDS denaturation, SDS-PAGE separation, and in-gel trypsin digestion, followed by analysis on a Q Exactive mass spectrometer (Thermo Fisher Scientific). Spectra were searched against the UniProt Homo sapiens database using MASCOT (Matrix Science), with standard parameters (≤2 missed cleavages, carbamidomethylation as fixed modification, oxidation, N-terminal acetylation, pyroglutamate formation, and deamidation). Protein identifications and validation were performed using Scaffold software, requiring ≥2 unique peptides per protein. A total of 920 proteins were identified across samples. The resulting EV proteomic dataset was used as the reference cargo pool for subsequent integrative analysis with tissue-level protein expression.
Animal model
All animal experiments were conducted under an approved Institutional Animal Care and Use Committee (IACUC) protocol (IS00006596) at the Houston Methodist Research Institute and followed ARRIVE 2.0 guidelines. Male C57BL/6J mice (8 weeks old) were housed under standard conditions. Power analysis (GraphPad StatMate) indicated—four to five animals per time point (α = 0.05). Humane endpoints and euthanasia criteria followed institutional guidelines.
Biodistribution studies
EVs were labeled with Vybrant™ DiD (Invitrogen) following the manufacturer’s instructions. Mice (n = 4) were anesthetized with 1%–3% isoflurane and shaved at the injection site. DiD-labeled CTRL, I/T, or LPS EVs (1 × 109 particles in 200 μL PBS) were administered via SubQ dorsal injection. The Sham group received PBS. To minimize confounders, mice from each group were injected prior to handling subsequent groups. EV biodistribution was assessed in vivo and ex vivo at days 1 and 3 post-injection using the IVIS® SpectrumCT imaging system (PerkinElmer). Signals were quantified as radiance photons using Living Image® 4.0 software, with regions of interest (ROIs) defined at the injection site and explanted organs. Fluorescence signals were normalized to the Sham group by subtracting total flux [p/s].
Surgery
Mice (n = 5) were anesthetized with 1%–3% isoflurane, and oral carprofen (5 mg/kg) was administered as analgesia. A 6 mm full-thickness dorsal skin defect was created using a biopsy punch, followed by SubQ injections of CTRL, I/T, or LPS EVs (1 × 109 particles in 200 μL PBS) around wound margins. The Sham group received PBS. Wounds were photographed and measured at days 0, 1, 3, 7, and 14. Wound area was calculated using A = πab, where a and b represent semi-major and semi-minor axes, and normalized to day 0 values. Mice were euthanized by CO2 inhalation (30%–70% displacement, >5 min) followed by cervical dislocation. Full-thickness skin explants were collected for downstream analyses.
RT-PCR immune gene expression array
Total RNA was extracted from day 1 skin explants using TRIzol™ reagent (Invitrogen). Complementary DNA synthesis was performed using the High-Capacity RNA-to-cDNA™ Kit, and qPCR was conducted using TaqMan™ Fast Advanced Master Mix (Applied Biosystems™) with a Mouse Immune Fast 96-well immune gene expression array plate (Applied Biosystems). Reactions were run on a StepOnePlus™ Real-Time PCR System. Relative expression was calculated using the 2−ΔΔCt method, with samples analyzed in biological triplicate. Differentially expressed genes (DEGs) relative to the Sham group were subjected to gene ontology (GO)/Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis using SRplot.
Proteome profiler mouse XL cytokine array
Proteins were extracted from day 3 skin explants using M-PER™ Mammalian Protein Extraction Reagent supplemented with 1× Halt™ Protease and Phosphatase Inhibitor Cocktail (Thermo Scientific). Protein concentration was determined by the Pierce™ BCA Protein Assay Kit (Thermo Scientific). A membrane-based immunoassay (Proteome Profiler Mouse XL Cytokine Array; R&D Systems) was used to assess 111 cytokines and related proteins. Signals were acquired using a ChemiDoc™ MP Imaging System and quantified with Fiji (Protein Array Analyzer plugin).
Integrative analysis of EV cargo and tissue protein expression
To identify EV-associated components potentially contributing to the observed tissue-level effects, an integrative comparative analysis was performed between EV proteomic cargo and protein expression changes in treated skin explants (day 3). Differential protein expression in skin explants was assessed using the Proteome Profiler Mouse XL Cytokine Array, comparing EV-treated groups to the Sham group. Only proteins exhibiting statistically significant changes relative to the Sham group were selected for downstream analysis. These differentially expressed proteins (DEPs), as well as DEGs from the transcriptomic analysis, were then cross-referenced with proteins identified in EV preparations from different treatments by LC–MS/MS. Overlapping proteins, as well as those functionally related through shared biological pathways, were identified, including inflammation, immunomodulation, chemotaxis, angiogenesis, growth factors, and ECM remodeling (including collagen synthesis). This targeted integration enabled the prioritization of a subset of EV-associated proteins most likely to contribute to the observed biological effects, rather than considering the full EV proteomic datasets. Importantly, these associations are correlative and do not establish direct mechanistic causality. Instead, they highlight biologically plausible links between EV cargo composition and protein expression changes within the wound microenvironment.
Western blot for collagen detection
Protein extracts (20 µg) were separated by SDS-PAGE and transferred to Immobilon®-P PVDF membranes (Millipore). Membranes were blocked in 5% bovine serum albumin and incubated overnight with primary antibodies against Collagen I, Collagen III, Collagen IV, and β-tubulin (Abcam, 1:1000). Following incubation with horseradish peroxidase-conjugated goat anti-rabbit (ab205718) or anti-mouse (ab205719) secondary antibodies, proteins were visualized using enhanced chemiluminescence with Immobilon Forte Western horseradish peroxidase (HRP) substrate (Millipore) and imaged using a ChemiDoc MP Imaging System.
Quantification of cell and ECM density in the skin
Skin explants (days 1, 3, 7, and 14) were fixed in 4% paraformaldehyde, paraffin embedded, and sectioned (5 μm). Sections were stained with hematoxylin and eosin (H&E) and Masson’s trichrome. Cell and ECM density were quantified using Fiji (Color Deconvolution plugin). Images were acquired using a Keyence BZ-X800 microscope (20× for H&E, 10× for trichrome). Cell nuclei and collagen were quantified after defining a threshold for blue signal (H&E) and green signal (trichrome), respectively. Three fields per sample and five ROIs per field were analyzed (n = 3; N = 45 ROIs/group), and results were expressed as average integrated density.
Imaging mass cytometry
Imaging mass cytometry (IMC, Hyperion™) was performed to assess 15 markers across 12 ROIs in skin explants (n = 3 mice/group). Cells were segmented using Ilastik and CellProfiler, based on DNA staining (Ir191) and other cell surface markers. Data were processed with the Histology Topography Cytometry Analysis Toolbox (HistoCAT) and R. Marker intensities were clipped at the 99.5th percentile and normalized (0-1) prior to clustering using Seurat (Louvain algorithm). Cell clusters were annotated based on mean marker expression and consolidated into 15 cell types. Cell densities were normalized to ROI areas. Neighborhood analysis identified spatial relationships between cell types, visualized as heatmaps (−1 to 1 scale).
Exploratory phase-integrated biological relevance scoring of EV cargo proteins
To further stratify EV-associated proteins potentially contributing to the distinct wound healing responses observed across treatment groups, an exploratory phase-integrated biological relevance scoring system was developed. This framework integrated EV cargo proteomic datasets with wound-site molecular, histological, and immune profiling analyses generated across the inflammatory, proliferative, and remodeling phases of wound healing. The objective of this analysis was not to infer direct mechanistic causality, but rather to prioritize EV-associated proteins showing the strongest temporal and biological concordance with tissue-level responses following EV treatment. A composite score was calculated based on molecular alignment with wound-site alterations (A), literature-supported functional relevance to wound healing biology (B), concordance with inflammatory-phase responses (C1), concordance with proliferative/remodeling transition responses (C2), concordance with late-stage remodeling outcomes (C3), and system-level biological relevance within wound healing-associated signaling networks (D).
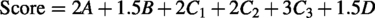
Greater weighting was assigned to remodeling-phase concordance to emphasize proteins associated with regenerative tissue resolution rather than persistent fibrotic remodeling. The resulting prioritization was intended as a hypothesis-generating framework to identify candidate EV cargo proteins for future mechanistic investigation. Detailed scoring criteria are provided in Supplementary Data.
Statistical analyses
Data are presented as mean ± standard deviation. Statistical comparisons were performed using one-way or two-way analysis of variance with Tukey’s post hoc test (GraphPad Prism 8.0). Results were normalized to Sham controls and expressed as fold-change or percentage change. Statistical significance was set at P < 0.05.
Results
MSC preconditioning affects EV production
EV-enriched extracellular particle preparations derived from BM-MSCs under inflammatory (I/T and LPS) and physiological (CTRL) conditions were characterized for concentration, size, and structural integrity. Figure 1a shows representative EV size distribution profiles, highlighting the intrinsic heterogeneity of EV populations across all conditions. Quantitative analysis revealed that LPS-MSCs produced EVs at concentrations comparable to CTRL-MSCs (4.8 × 1010 ± 6.3 × 109 particles/mL vs. 4.3 × 1010 ± 9.6 × 109 particles/mL), whereas I/T-MSCs exhibited a significant reduction in EV yield (2.0 × 1010 ± 2.3 × 109 particles/mL) compared to both CTRL (P < 0.05) and LPS (P < 0.01) (Fig. 1b). EV size distribution remained consistent across all groups, with mean diameters of ∼100 nm (CTRL: 102 ± 10.8 nm, I/T: 97.0 ± 13.1 nm, LPS: 100.5 ± 8.3 nm), confirming enrichment within the small EV range (<200 nm; Fig. 1c). Ultrastructural analysis confirmed preserved EV morphology across conditions, including a defined lipid bilayer and electron-lucent core (Fig. 1d). Consistently, Western blot analysis validated the presence of established EV markers (CD63, CD9, Alix, and TSG101) in all preparations (Fig. 1e).
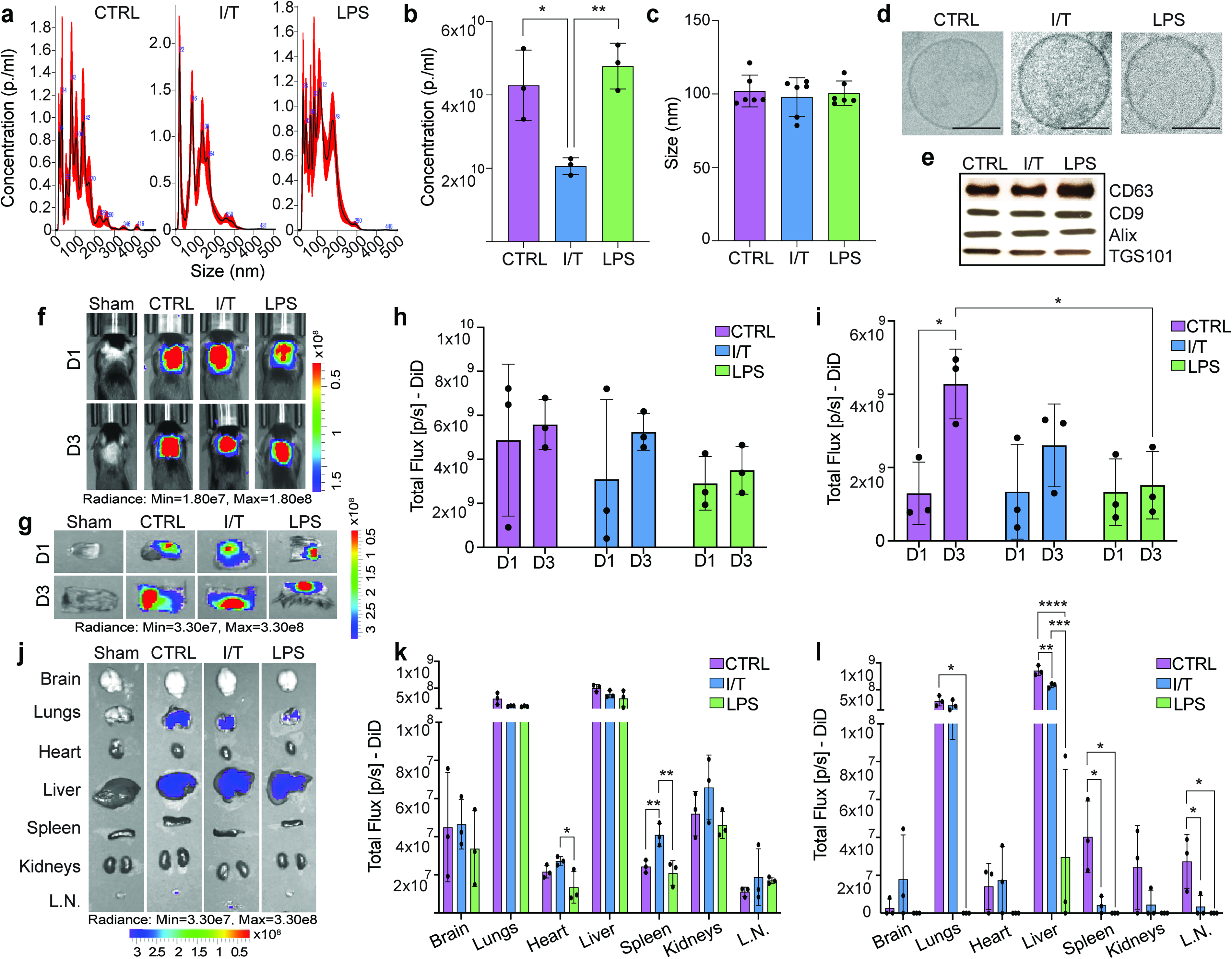
Characterization and biodistribution of extracellular vesicles (EVs) derived from conditioned mesenchymal stromal cells (MSCs). EV particle distribution
Subcutaneously injected EVs mainly accumulate at the site of injection
Fluorescently labeled EVs were SubQ administered dorsally, and spatial distribution was monitored at days 1 and 3 post injection (Fig. 1f). At day 1, EV accumulation at the injection site was comparable across all groups, and this pattern was maintained at day 1 and 3 (Fig. 1h, g) A time-dependent increase in CTRL-EV signal was observed at day 3 compared to day 1 (P < 0.05), as well as higher retention relative to LPS-EVs (P < 0.05) (Fig. 1i, j). Systemic biodistribution analysis at day 1 demonstrated that, despite local delivery, EVs were detectable across multiple organs, with a preferential accumulation in the liver and lungs across all groups (Fig. 1k). Condition-specific differences were limited but notable, including increased accumulation of I/T-EVs in the heart compared to LPS-EVs (P < 0.05), and elevated splenic localization of I/T-EVs relative to both CTRL-EVs (P < 0.05) and LPS (P < 0.01) groups. By day 3, overall systemic EV signal decreased across organs for all groups (Fig. 1l). Significant differences emerged in these organs, with reduced liver accumulation observed for both I/T-EVs and LPS compared to CTRL-EVs (P < 0.01 and P < 0.0001, respectively) and for LPS-EVs compared to I/T-EVs (P < 0.001). Lung distribution was also lower in LPS-EVs compared to CTRL-EVs (P < 0.05). In secondary lymphoid organs, both I/T-EVs and LPS-EVs showed reduced accumulation in the spleen and lymph nodes compared to CTRL (P < 0.05).
EV treatment differentially modulates wound contraction dynamics and cellular responses
Since DiD-labeled EVs remained detectable at the wound site for at least 3 days following SubQ administration, the selected EV particle dose was considered sufficient to support sustained local exposure throughout the early phases of wound healing. A full-thickness punch biopsy wound healing model was used to assess SubQ administration of I/T- and LPS-EVs relative to CTRL-EVs over 14 days. Comparable overall closure was observed across all conditions (Fig. 2a). Figure 2b shows wound contraction, with an initial increase at day 1 across all groups (CTRL: 119%, I/T: 133%, LPS: 121%; baseline = 100%). By day 3, wound areas in CTRL and LPS decreased to 45% and 58%, respectively, whereas I/T remained unchanged (99%), indicating delayed contraction and a significant difference from Sham (P < 0.05). Contraction from day 1 to 3 was significant in CTRL and LPS (P < 0.05). By day 7, CTRL and LPS measured 49% and 33%, respectively, while I/T decreased to 37% (P < 0.05). From days 7 to 14, all EV-treated groups showed significant contraction (CTRL and I/T: P < 0.01; LPS: P < 0.0001), with near-complete closure by day 14 (CTRL: 3%, I/T: 3%, LPS: 1%). Dermal cellularity (Fig. 2c) showed that at day 1, I/T increased compared to Sham (P < 0.0001), whereas CTRL and LPS showed nonsignificant trends. By day 3, cellularity increased in all groups, with significant changes in I/T (P < 0.05) and LPS (P < 0.0001). At day 7, cellular density plateaued in I/T and LPS, remaining higher than Sham (P < 0.01), while CTRL increased from day 3 (P < 0.0001) without differences from Sham. By day 14, cellular density was reduced in all EV-treated groups compared to Sham (CTRL: P < 0.01; I/T and LPS: P < 0.0001), with I/T showing the lowest values (vs. CTRL: P < 0.0001 and LPS: P < 0.001). Compared to day 7, cellular density increased at day 14 in all groups (P < 0.0001) while remaining below Sham levels.
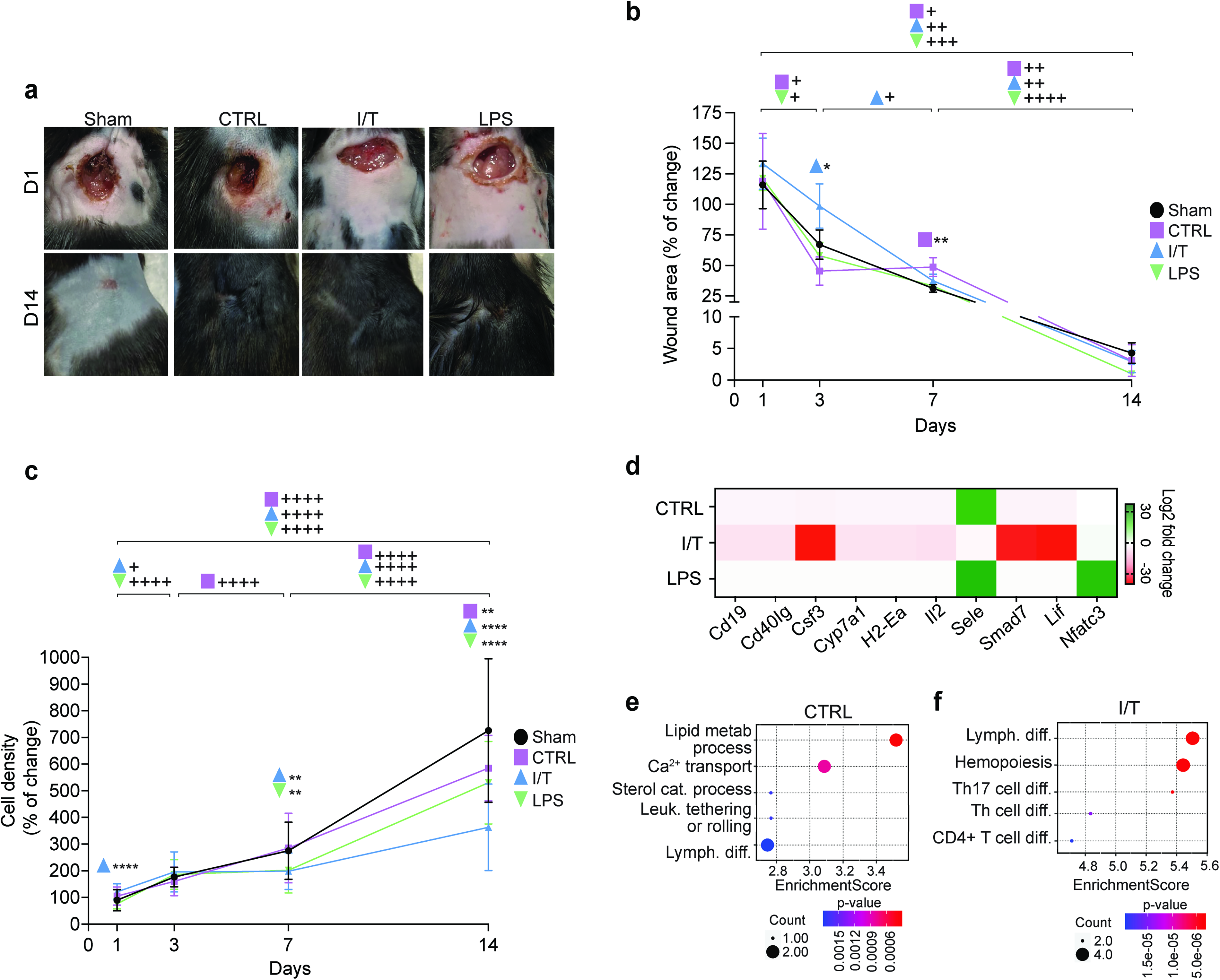
Wound contraction, cellular density, and gene expression analysis following EV treatment. Representative images illustrate wound contraction following SubQ injection of PBS (Sham), CTRL-EVs, I/T-EVs, or LPS-EVs
Differentially conditioned MSC-EVs drive distinct molecular signatures
A qPCR-based cytokine array was performed on skin explants to profile immune-related gene expression following treatment (Fig. 2d). Distinct transcriptional signatures were identified, with 10 DEGs reaching statistical significance. As summarized in Table 1, CTRL-EVs induced a moderate response, with Nfatc3 upregulated and Cd19, Cd40lg, Cyp7a1, H2-Ea, and Sele downregulated. In contrast, I/T-EVs elicited a broader transcriptional shift, with Csf3, Smad7, Lif, and Nfatc3 upregulated and Cd19, Cd40lg, Cyp7a1, H2-Ea, and Il2 downregulated, with stronger suppression compared to CTRL-EVs for shared targets. LPS-EVs did not induce significant changes. GO enrichment analysis further highlighted functional divergence between EV conditions. CTRL signatures were enriched in lipid metabolism, sterol catabolism, Ca2+ transport, leukocyte tethering and rolling, and lymphocyte differentiation (Fig. 2e), whereas I/T-EV signatures were associated with hematopoiesis and adaptive immune regulation, including differentiation of T helper cells (Th), CD4+ T cells, and Th17 cells (Fig. 2f).
Differentially Expressed Genes in Skin Explants After Mesenchymal Stromal Cell-Derived Extracellular Vesicle Treatment in vivo
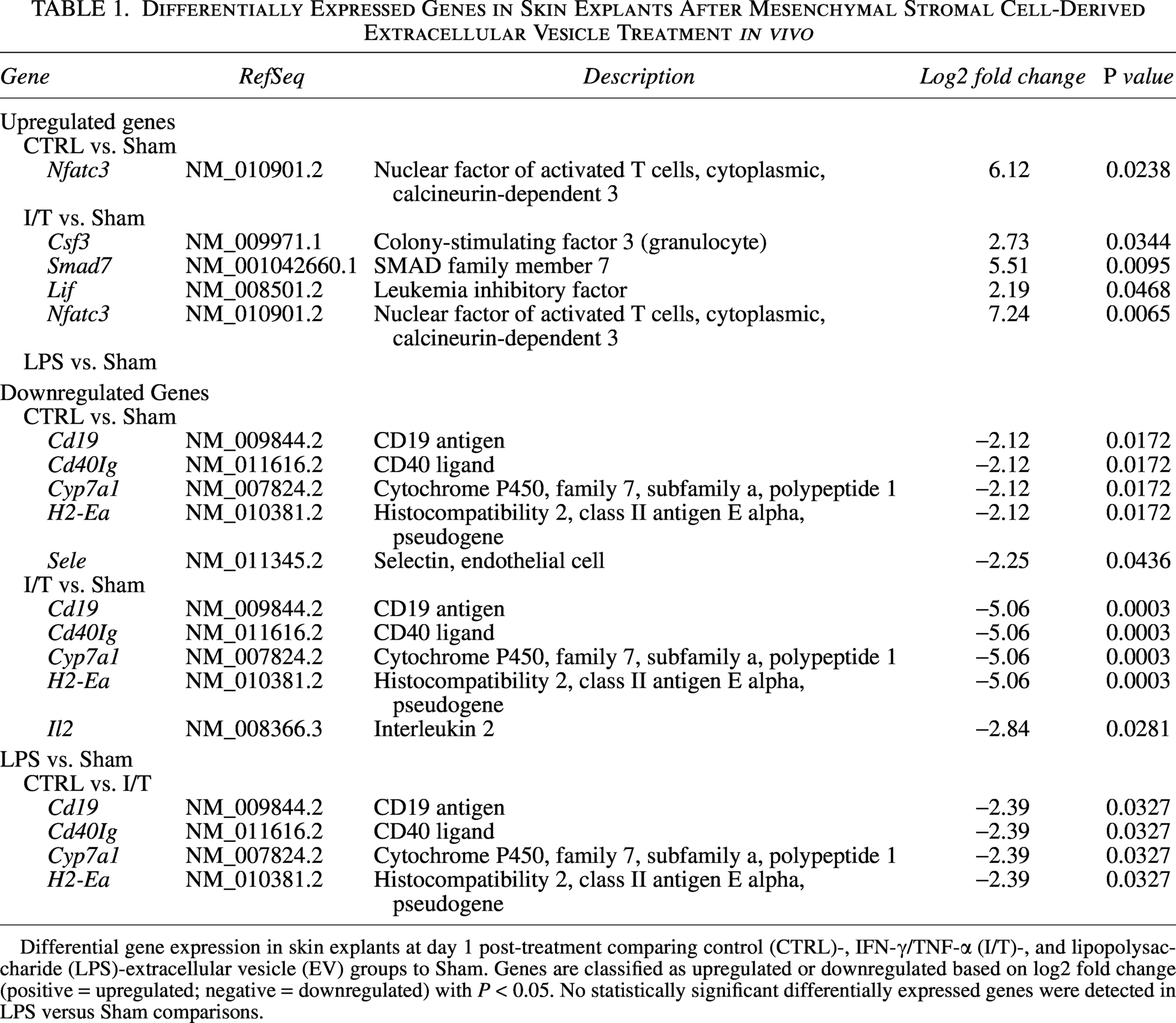
Differential gene expression in skin explants at day 1 post-treatment comparing control (CTRL)-, IFN-γ/TNF-α (I/T)-, and lipopolysaccharide (LPS)-extracellular vesicle (EV) groups to Sham. Genes are classified as upregulated or downregulated based on log2 fold change (positive = upregulated; negative = downregulated) with P < 0.05. No statistically significant differentially expressed genes were detected in LPS versus Sham comparisons.
MSC-EV conditioning reshapes early immune cell landscapes at the injury site
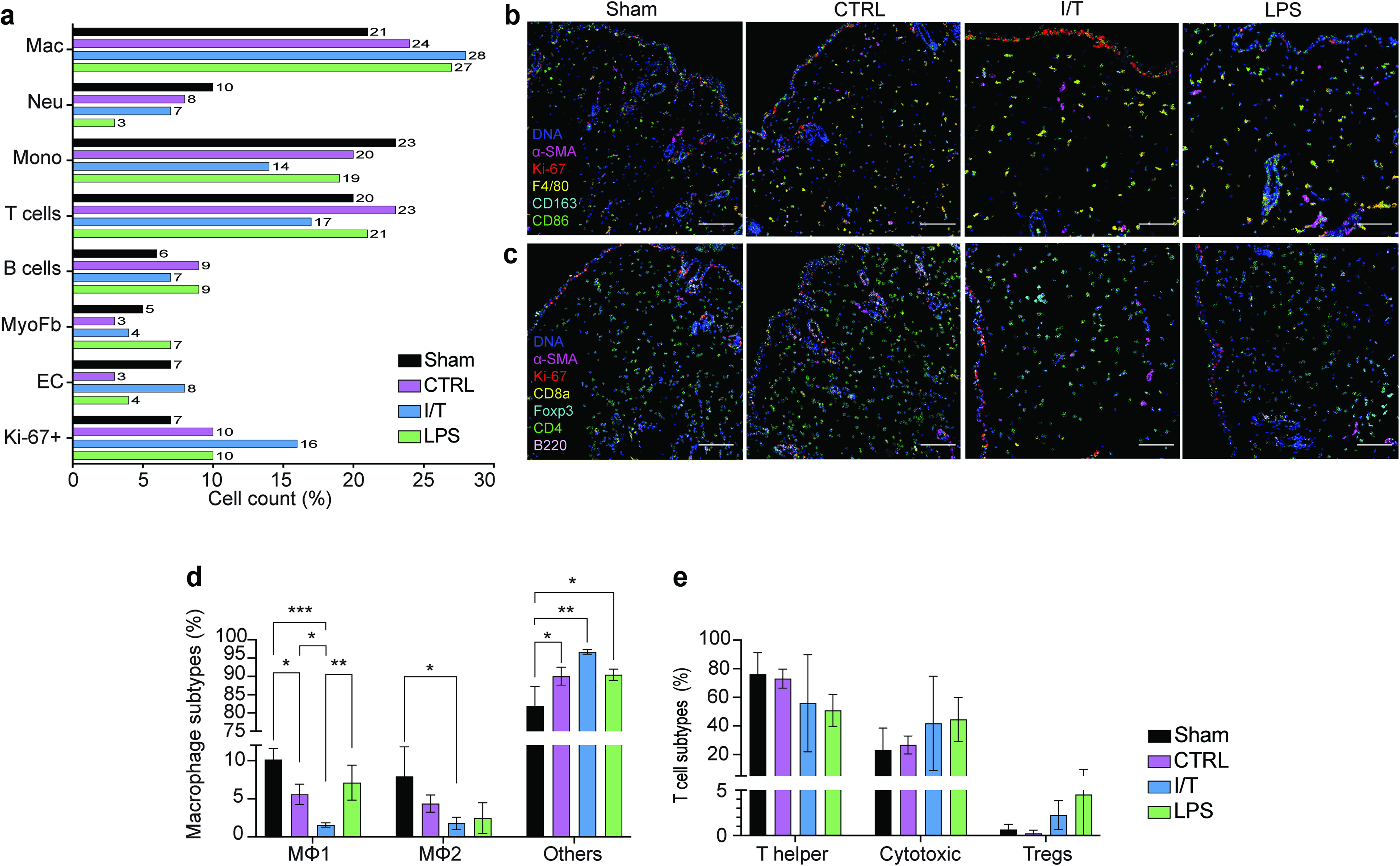
Skin explants collected at day 1 were analyzed using IMC for single-cell profiling. Analysis revealed distinct tissue architectures and cell distribution across treatments (Supplementary Fig. S1a), while neighborhood analysis indicated treatment-specific cellular interactions (Supplementary Fig. S1b). All EV treatments shifted the inflammatory landscape relative to Sham, with increased MΦs and B cells, and reduced neutrophils and monocytes (Fig. 3a). T cell responses varied, with increased total T cells in CTRL and LPS, and reduced levels in I/T.
Immune and stromal cell population dynamics at the wound site 1-day post treatment. t-SNE analysis of immune and stromal cell populations obtained from Hyperion imaging mass cytometry across CTRL, I/T, and LPS treatment groups, showing distinct clustering of cell populations at the wound site
All EV groups showed increased Ki67+ proliferative cells, most pronounced in I/T, predominantly in the epidermal compartment. MΦ subpopulation analysis revealed expansion of undifferentiated MΦs accounting for 97% in I/T (P < 0.01) and 90% in CTRL and LPS (P < 0.05) (Fig. 3b, d), with concurrent reduction in MΦ1 and MΦ2 subsets. MΦ1 decreased in I/T (P < 0.001) and CTRL (P < 0.05), while MΦ2 reduction was significant only in I/T (P < 0.05).
T cell subset analysis showed decreased CD4+ T cells and increased CD8+ T cells across all EV groups (Fig. 3c, e). Regulatory T cells (Tregs) decreased in CTRL but increased in I/T and LPS.
Nonimmune compartments were also affected: endothelial cells decreased in CTRL (3%) and LPS (4%) but increased in I/T (8%), while myofibroblasts decreased in CTRL (3%) and I/T (4%) but increased in LPS (7%). Histology and phenotype maps confirmed the spatial organization of these populations (Supplementary Fig. S1c).
EVs suppress inflammation and orchestrate ECM remodeling through distinct protein signatures
Day 3 skin explants were analyzed using a cytokine array. Normalization to Sham revealed downregulation of immune-related proteins across EV-treated groups (Supplementary Fig. S2a). Of 111 proteins analyzed, 36 DEPs were identified, of which 30 were downregulated and 6 upregulated (Table 2). CTRL-EVs induced moderate suppression, whereas I/T-EVs exhibited a broader effect. LPS-EVs exhibited a more limited, overlapping profile without uniquely downregulated proteins. Despite global suppression, proteins associated with tissue repair and ECM remodeling remained elevated, including mediators of growth factor signaling and matrix turnover. Key mediators such as C-C motif chemokine ligand 6 (CCL6), epidermal growth factor (EGF), insulin-like growth factor-binding protein 3 (IGFBP-3), matrix metallopeptidase 3 (MMP3), matrix metallopeptidase 9 (MMP9), and osteopontin (OPN) were upregulated (Fig. 4a), whereas TNF-α, IFN-γ, IL-1α, and MMP2 were reduced. GO enrichment revealed condition-specific programs (Fig. 4b): CTRL-EVs were associated with cytokine signaling and lymphocyte recruitment, I/T-EVs with leukocyte migration and IL-1 responses, and LPS-EVs with epithelial morphogenesis and lymphocyte co-stimulation.
Differentially Expressed Proteins in Skin Explants after Mesenchymal Stromal Cell-Derived Extracellular Vesicle Treatment in vivo
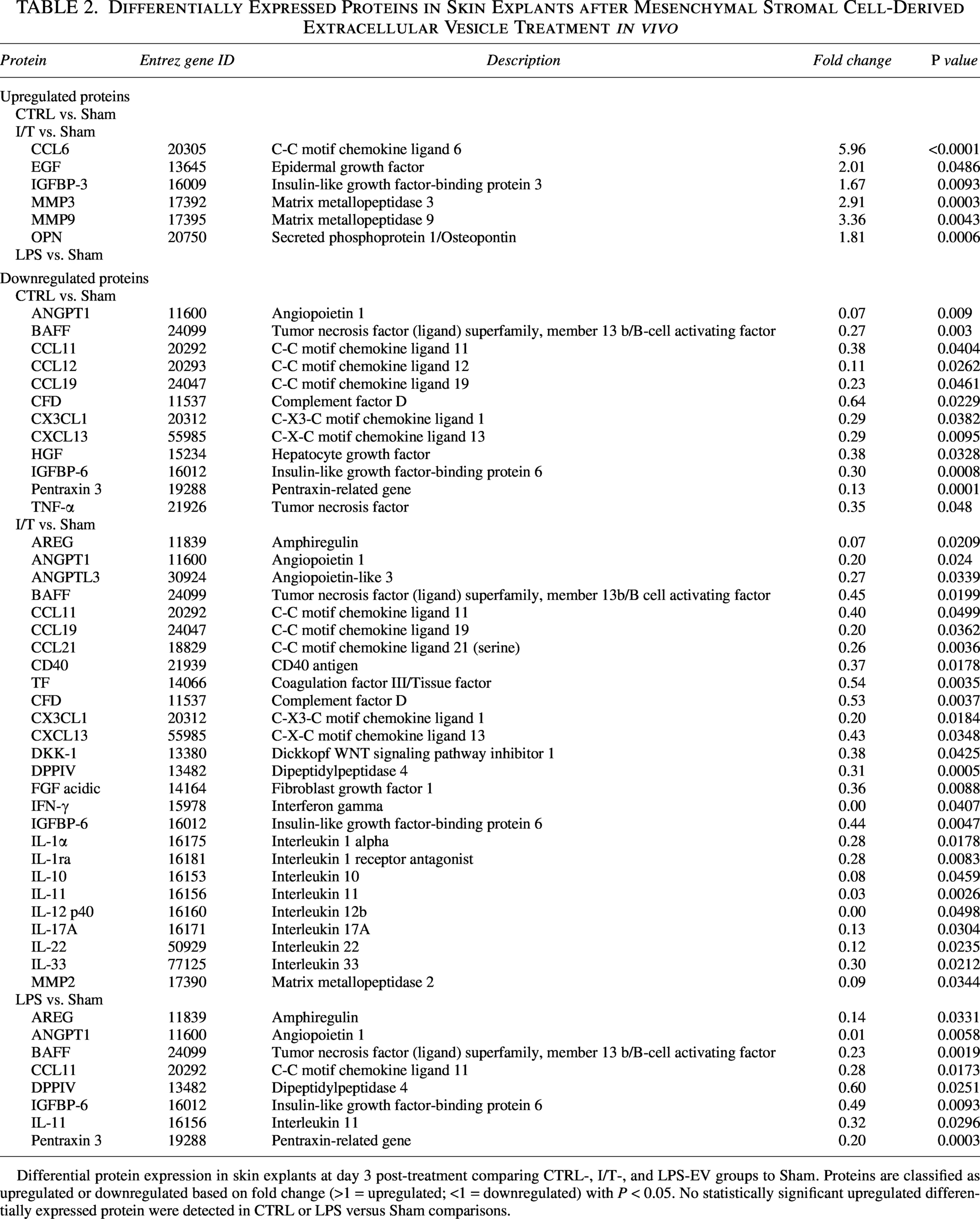
Differential protein expression in skin explants at day 3 post-treatment comparing CTRL-, I/T-, and LPS-EV groups to Sham. Proteins are classified as upregulated or downregulated based on fold change (>1 = upregulated; <1 = downregulated) with P < 0.05. No statistically significant upregulated differentially expressed protein were detected in CTRL or LPS versus Sham comparisons.
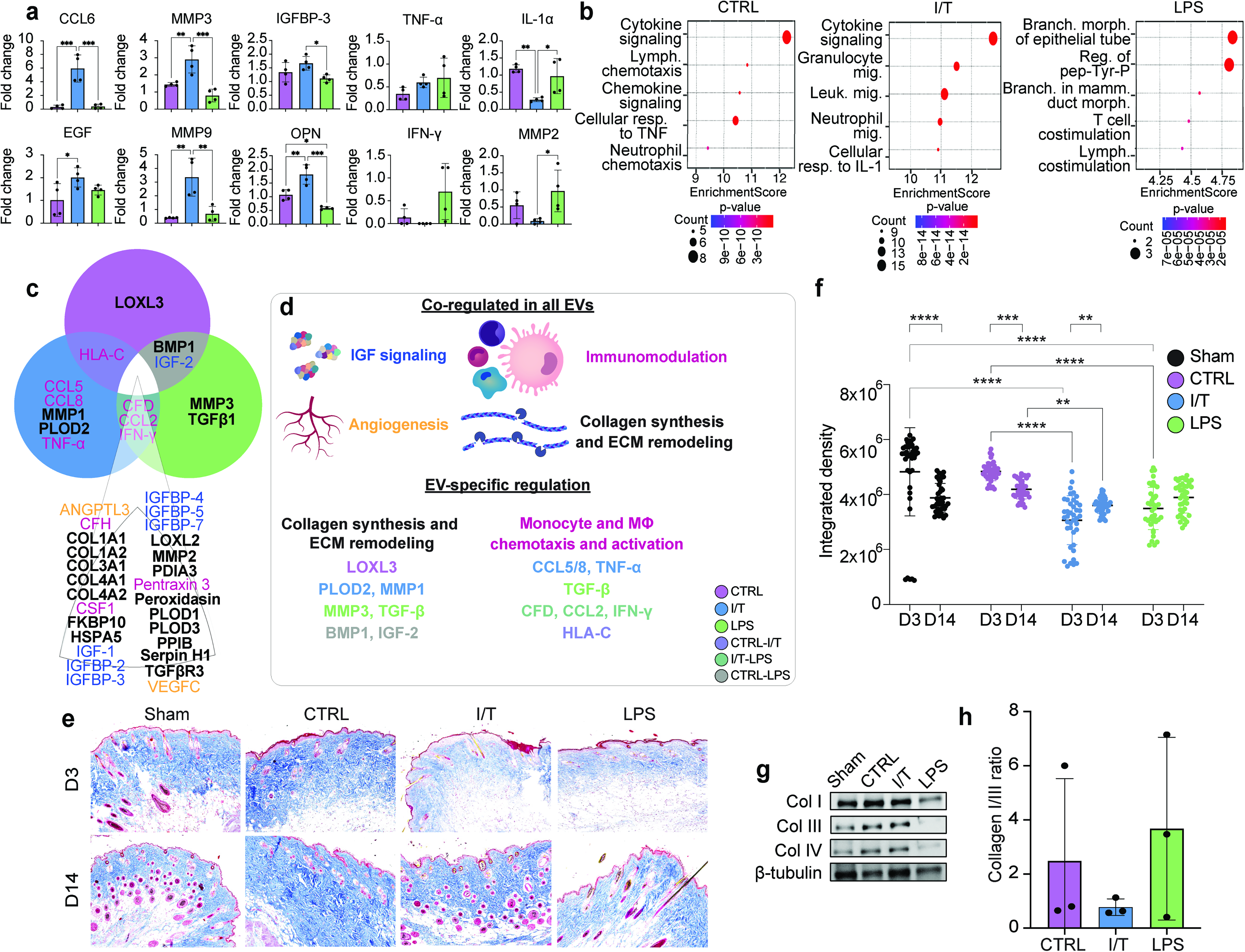
Protein expression and extracellular matrix (ECM) density at the wound site. Protein expression in skin explants at day 3 was analyzed using an immunoarray-targeting immune response-related proteins. A selection of some of the key proteins upregulated and downregulated by EV treatments that are directly associated with immunomodulation and tissue repair
Integration with EV proteomic cargo demonstrated correspondence between EV composition and in vivo protein modulation (Fig. 4c). EVs across all groups contained proteins related to immunomodulation, ECM remodeling, collagen synthesis, and angiogenesis (Fig. 4d), with conditioning-specific differences and partial overlap across groups. CTRL-EVs contained unique structural remodeling components, I/T-EVs were enriched in immune and matrix remodeling mediators, and LPS-EVs carried factors linked to matrix turnover and signaling pathways.
Differentially conditioned MSC-EVs modulate ECM density and collagen organization
ECM remodeling was assessed by Masson’s Trichrome staining (Fig. 4e). At day 3, ECM density was significantly reduced in I/T and LPS compared to Sham and CTRL (P < 0.0001), while CTRL remained comparable to Sham (Fig. 4f). By day 14, ECM density increased in CTRL (P < 0.05), remained unchanged in LPS, and was persistently reduced in I/T compared to all groups (P < 0.05–0.0001).
Temporal analysis (Supplementary Fig. S2b, c) showed early ECM reduction in all EV groups (most pronounced in I/T and LPS), followed by recovery in CTRL and LPS, while I/T remained lower overall. By day 14, LPS most closely matched Sham ECM levels.
Western blot analysis of collagen I, III, and IV at day 3 showed qualitative differences without significant changes (Fig. 4g). However, the collagen I/III ratio was higher in CTRL and LPS (2.5 ± 3.0- and 3.7 ± 3.4-fold change, respectively) and lower in I/T (0.8 ± 0.3-fold change) (Fig. 4h).
Exploratory prioritization of EV cargo proteins identifies treatment-associated wound healing signatures
To further stratify EV-associated proteins potentially contributing to the distinct wound healing responses observed across treatment groups, an exploratory phase-integrated biological relevance scoring system was applied, integrating EV proteomics with wound-site transcriptomic, proteomic, histological, and immune profiling datasets across healing stages. Prioritized proteins differed across treatment groups and aligned with distinct tissue remodeling trajectories. Following this approach, the top 5 highest-ranked EV cargo proteins by treatment group were identified and presented in Supplementary Table S1. CTRL-EVs preferentially enriched proteins associated with collagen organization and matrix stabilization, including MMP2, LOXL2, PLOD1, COL1A1, and COL1A2, consistent with the persistent ECM density and delayed wound contraction observed during later healing stages. I/T-EVs showed enrichment of proteins associated with inflammatory modulation and regenerative remodeling, including IGFBP-3, MMP2, TGFβR3, CSF1, and HSPA5. These proteins demonstrated concordance with reduced inflammatory signatures, altered MΦ-associated responses, decreased ECM burden, and improved remodeling outcomes observed in vivo. LPS-EVs preferentially enriched proteins associated with active matrix remodeling, including MMP2, LOXL2, TGFβ-1, MMP3, and PLOD3, consistent with the early remodeling dynamics and transitional ECM responses observed following treatment. Importantly, incorporation of late remodeling outcomes into the prioritization framework enabled discrimination between proteins associated with regenerative tissue resolution and those associated with persistent ECM accumulation. Together, these analyses supported the presence of distinct EV-associated biological programs across treatment conditions.
Discussion
Preconditioning MSCs with inflammatory stimuli has been shown to enhance the therapeutic efficacy of their EVs, particularly in models of skin and soft tissue repair.36,37 In this study, we investigated how preconditioning BM-MSCs with either I/T or LPS, mimicking sterile inflammation and bacterial cues, modulates distinct regenerative and immunomodulatory programs mediated by their EVs in wound healing.
Consistent with previous reports that inflammatory cues influence MSC-EV biogenesis, 38 I/T preconditioning significantly reduced EV yield compared to LPS treatment and unconditioned controls. Despite these differences, all EV populations exhibited comparable size distributions and canonical marker expression, supporting direct functional comparisons independent of major physicochemical differences. This contrasts with findings from Zheng et al., 39 who reported increased EV production in adipose-derived MSCs following LPS stimulation, indicating that EV output is context- and source-dependent rather than uniformly enhanced by inflammatory priming. 40
Fluorescent tracking revealed preferential EV retention at the wound site, supporting a predominantly local mode of action linked to injury tropism, consistent with prior studies.41,42 Although limited systemic dissemination was observed, mainly to the liver and lungs, this pattern mirrors prior reports of EV biodistribution across various administration routes. 43
Importantly, EVs persisted at the injection site over time, echoing findings by Hsu et al. 44 At day 1, all EV-treated groups demonstrated similar retention, with differences emerging only in the heart and spleen. By day 3, EV signal decreased across organs and skin explants, indicating progressive clearance and redistribution while maintaining local activity, consistent with a balance between systemic uptake and site-specific persistence.
Our EV proteomic analysis demonstrates that EV cargo is not uniformly altered by preconditioning but selectively reprogrammed, with all groups sharing a core set of pro-repair proteins and factors involved in MΦ activation, ECM synthesis and remodeling, and angiogenic signaling,45–47 whereas condition-specific differences align with the observed in vivo responses. Accordingly, EV populations can be interpreted as functionally distinct: CTRL-EVs bias early immune resolution, I/T-EVs coordinate immune modulation with active matrix remodeling, and LPS-EVs promote early contraction and epithelial remodeling with intermediate control of inflammation (Fig. 5).
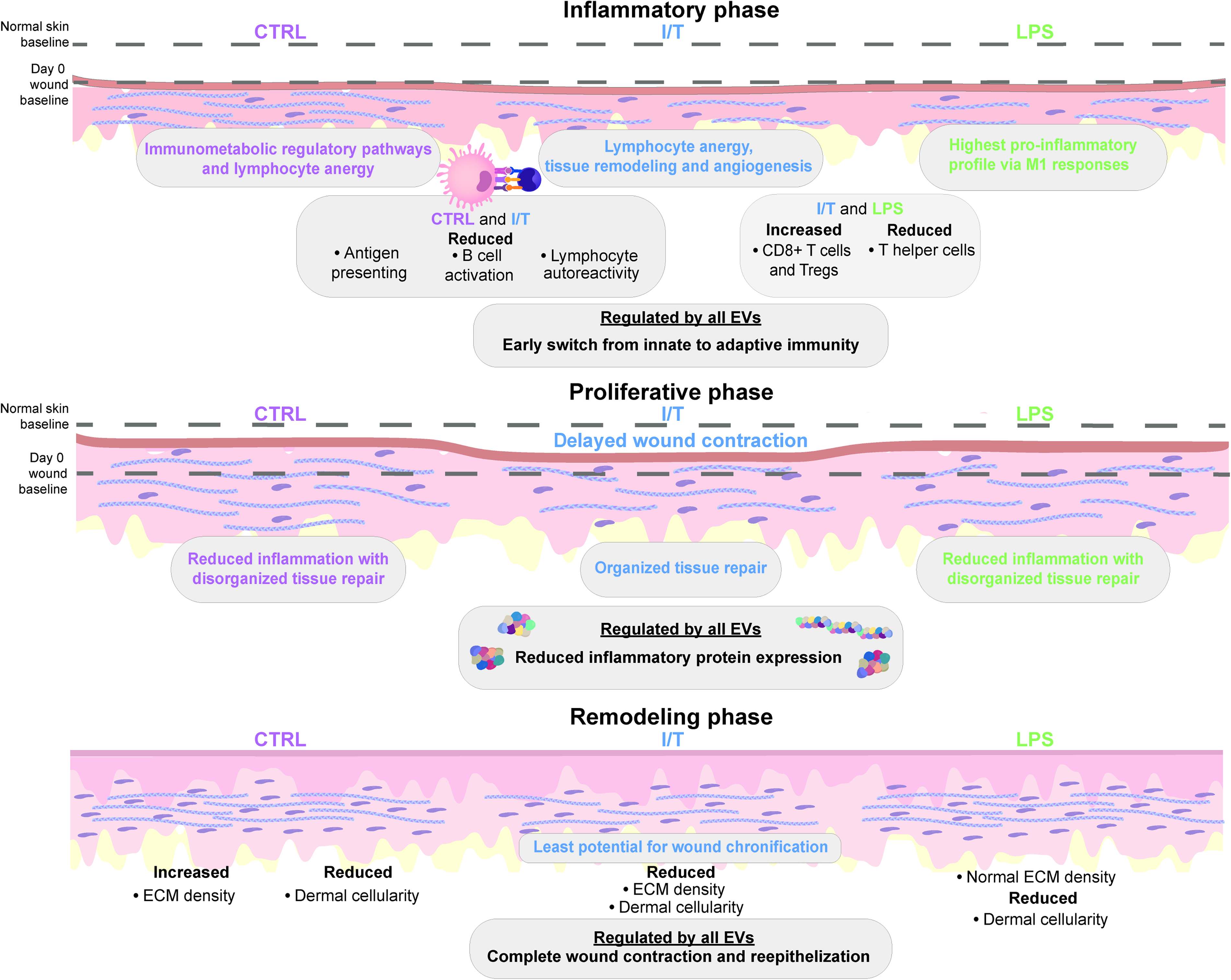
Schematic representation of MSC-EV preconditioning-dependent effects on wound healing. Schematic illustrating the functional outcomes of preconditioned MSC-EVs in an in vivo full-thickness wound healing model. The figure depicts three panels showing the differential results on early immune modulation, highlighting the effects on immune cell populations and inflammatory signaling across conditions during the inflammatory, proliferative, and remodeling phases of wound healing. Downstream effects on ECM remodeling are also depicted, showing condition-specific differences in matrix deposition, organization, and collagen remodeling. The schematic emphasizes how preconditioning defines EV functional states, linking molecular signatures to distinct wound healing trajectories.
Among these, CTRL-EVs generated an immune landscape most closely resembling Sham. Despite this similarity, CTRL-EVs accelerated early contraction, promoting a shift from innate to adaptive immunity by day 1, with decreased neutrophils and monocytes and increased MΦ and B cell infiltration. This contrasts with the delayed adaptive engagement observed in Sham. 48
Transcriptomic analysis revealed enrichment in lipid metabolism, calcium transport, leukocyte tethering/rolling, and lymphocyte differentiation. Downregulation of Cyp7a1 may affect cholesterol homeostasis and bile acid-mediated immune regulation,49,50 while reduced Sele may impair leukocyte recruitment. 51 Suppression of Cd19, Cd40lg, and H2-Ea suggests reduced B cell activation and antigen presentation. 52 Conversely, Nfatc3 upregulation indicates modulation rather than suppression of immune signaling. 53 By day 3, CTRL-EVs further suppressed inflammation through inhibition of TNF-α-associated pathways and complement activation, with reduced Pentraxin 3 and CFD, supporting resolution. 54 Decreased chemokines (CCL12, CCL19, CCL11, CXCL1, and CXCL13) and BAFF indicate reduced immune recruitment over time. In parallel, pro-regenerative factors (HGF, IGFBP-6, and ANGPT1) were modulated without inducing strong matrix-remodeling programs. 55
CTRL-EV cargo, including LOXL3 and shared factors (BMP1 and IGF-2), also present in LPS-EVs, further supports collagen maturation and ECM deposition,56–58 while HLA-C (shared with I/T-EVs) suggests engagement of adaptive immunity, including CD8+ T and NK cells. 59 Structurally, ECM density paralleled Sham at day 3 but had a higher (∼2-fold) collagen I/III ratio, indicating accelerated collagen remodeling and increased matrix stiffness without proportional structural organization. 60 Overall, CTRL-EVs reduce inflammation but promote a less organized repair due to limited activation of coordinated pro-regenerative and remodeling programs.
In contrast, I/T-EVs delayed early contraction, accompanied by a transient spike in dermal cellularity, consistent with enhanced early immune recruitment. 61 The immune profile resembled that of CTRL-EVs but with a lower M1/M2 MΦ ratio and increased undifferentiated MΦs, indicating a more plastic MΦ compartment. This aligns with IFN-γ and TNF-α roles: IFN-γ favors M2 polarization and Treg expansion, 62 while TNF-α supports regeneration and limits fibrosis. 63 Tregs increased modestly, and endothelial cells were elevated, supporting angiogenesis.
Transcriptomics revealed a distinct signature, with Il-2 downregulation promoting immune tolerance 64 and upregulation of Csf3, Smad7, and Lif, implicating coordinated activation of regenerative, hematopoietic, and immune-regulatory pathways.65–68 By day 3, I/T-EVs increased CCL6, OPN, MMP3, MMP9, EGF, and IGFBP-3, enhancing MΦ recruitment, ECM remodeling, and epithelial and fibroblast proliferation, thereby supporting the transition into the proliferative phase. 69 Pro-inflammatory mediators were suppressed without IL-10 compensation, indicating self-resolving and controlled immune modulation. 70 Downregulation of DKK-1 may alleviate Wnt signaling constraints and support matrix turnover, 71 while reduced TF, ANGPTL3, FGF acidic, AREG, DPPIV, and IL-11 may temper angiogenesis and fibrosis. 72 EV proteomic analysis showed strong cargo enrichment of chemotactic and remodeling factors (CCL2/5/8, CFD, MMP1, and PLOD2),73–76 indicating coordinated ECM degradation and synthesis rather than unidirectional remodeling.
The presence of IFN-γ and TNF-α within EV cargo, coupled with a net anti-inflammatory outcome, suggests context-dependent tolerization enabling controlled inflammation resolution with sustained regenerative signaling.77,78 This aligns with prior reports showing enhanced M1-to-M2 switching under combined I/T stimulation. 79 ECM remodeling was balanced, with a collagen I/III ratio near 1 normalized to Sham, indicative of an active remodeling process and ECM breakdown that favors granulation tissue formation. 80 By day 14, I/T-treated wounds exhibited the lowest ECM and cell density, reflecting efficient resolution and advanced maturation. These features indicate a more synchronized transition from inflammation to tissue repair compared with other conditions, with reduced risk of wound chronification.
In contrast, LPS-EVs accelerated early contraction and induced a distinct immune profile, with a higher M1/M2 ratio and increased CD8+ T cells. This diverges from prior studies showing LPS-driven M2 polarization,18,19,39 likely reflecting dose- or context-dependent effects.81,82 Early transcriptomic changes were minimal, suggesting post-transcriptional regulation. However, by day 3, inflammation resolved with reduced Pentraxin 3, BAFF, and CCL11, alongside decreased AREG, DPPIV, and IL-11, pointing to effective inflammation control without fibrotic overactivation. GO analysis indicated enrichment in epithelial morphogenesis and receptor signaling pathways (EGFR and VEGFR), consistent with epithelial repair. 83
LPS-EV cargo included MMP3 and chemotactic factors (CFD, CCL2), supporting matrix reorganization and immune recruitment.76,84,85 Delivery of TGF-β and IFN-γ further supports epithelial remodeling and matrix regulation. 86 Structurally, reduced ECM density with elevated collagen I/III ratios suggests compensatory stiffening supporting contraction. 87 Overall, LPS-EVs favor rapid structural closure with less integrated regenerative remodeling compared with I/T-EVs. By day 14, the ECM density normalized, indicating effective remodeling without fibrosis.
Integration of multidimensional wound healing datasets suggested that MSC-EV treatments engage coordinated and temporally distinct regenerative programs rather than isolated molecular pathways. Although all EV conditions influenced ECM remodeling and immune regulation, the analysis highlighted divergent healing trajectories across treatment groups. I/T-EVs preferentially aligned with inflammatory resolution, controlled matrix turnover, and reduced late-stage ECM accumulation, whereas LPS-EVs showed features of active remodeling associated with both transient profibrotic and compensatory remodeling responses. In contrast, CTRL-EVs preferentially enriched proteins associated with matrix stabilization and collagen retention. Importantly, incorporation of late remodeling outcomes enabled prioritization of candidate EV cargo proteins showing concordance with regenerative remodeling rather than persistent fibrotic matrix accumulation. Together, these findings support the concept that MSC preconditioning reshapes EV cargo composition and downstream tissue remodeling behavior during wound repair.
Our study establishes a mechanistic framework for precision EV-based therapies and highlights key challenges in EV research, including variability in yield, isolation and purification methods, cargo heterogeneity, and the ongoing difficulty in defining biologically relevant EV potency and dosing strategies. 88 Standardization of preconditioning, characterization, and administration parameters will be essential for clinical translation. Limitations of this study include the use of healthy murine models and inherent variability in wound healing responses, which may differ in pathological conditions such as diabetes or cardiovascular disease. Given the context-dependent effects observed, these differences may be amplified in disease settings, warranting future studies in comorbidity models to better define translational potential.
Conclusions
MSC preconditioning with distinct inflammatory cues generates EVs with unique molecular signatures and functionally distinct regenerative profiles. Our findings reinforce the value of preconditioned EVs as tunable, cell-free platforms for targeted tissue repair. Unlike MSCs, which exhibit significant phenotypic plasticity and source- and context-specific variability in vivo, EVs provide a more stable and controllable representation of the cellular state at the time of release, enabling more predictable and tunable therapeutic responses. Importantly, the differential cargo and functional outputs observed across preconditioning conditions suggest that EV-mediated effects can be deconvoluted into discrete molecular drivers. This creates an opportunity to identify minimal sets of proteins, RNAs, and lipids responsible for specific regenerative and immunomodulatory functions, enabling the design of EV-inspired or mimetic platforms.89,90 Such approaches could support the development of simplified, scalable, and precisely defined therapeutic strategies aligned with precision medicine frameworks.
Ethical Considerations and Consent to Participate
The original source of BM-MSCs (ATCC) has confirmed initial ethical approval for the collection of human cells, and the donor has signed an informed consent form.
Animal Ethics
All animal experiments were conducted in compliance with an approved protocol from the IACUC at the Houston Methodist Research Institute (approval title: Therapeutic studies in gynecological cancer models; approval number: IS00006596; approval date: December 06, 2022). These studies adhered to the guidelines of the Animal Welfare Act and the Guide for the Care and Use of Laboratory Animals.
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Footnotes
Acknowledgments
Histological examination and single-cell analyses were performed through the Houston Methodist Research Institute Research Pathology Core and ImmunoMonitoring Core, respectively (fee-for-service). Cryo-TEM analyses were performed at the CryoEM Core at Baylor College of Medicine. The authors declare that no artificial intelligence-generated content was used in this article.
Author Disclosure Statement
A.G. is a cofounder and chief scientific advisor of Continuity Bioscience.
Funding Information
B.C. received salary support from the
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.