
Abstract
Focal segmental glomerulosclerosis (FSGS) is a major cause of nephrotic syndrome and end-stage kidney disease (ESKD). Many cases are attributable to pathogenic variants in podocyte-related genes, such as inverted formin 2 (INF2). However, no specific treatment exists for hereditary FSGS, and experimental platforms that faithfully model podocyte injury remain limited. Therefore, in this study, we developed an injury model by generating induced pluripotent stem cells (iPSCs) from a patient with INF2-associated FSGS and differentiating them into kidney organoids. Podocytes were validated by immunofluorescence staining for podocyte-specific markers. We induced podocyte injury in this model using puromycin aminonucleoside (PAN) in a dose-dependent manner. Injury severity was quantified by measuring podocalyxin fluorescence intensity. Cyclosporine A (CsA) or voclosporin (VCS) was administered as a 1-h pretreatment before PAN exposure to evaluate their podocyte-protective effects. The kidney organoids exhibited well-defined podocyte marker expression, confirming successful differentiation. PAN exposure caused a significant and concentration-dependent reduction in podocalyxin fluorescence, indicating robust induction of podocyte injury in organoids harboring the INF2 variant. Pretreatment with CsA or VCS significantly attenuated PAN-induced podocyte injury and preserved podocyte marker expression. CsA and VCS reduced podocyte injury to similar extents. In conclusion, we established a patient-specific kidney organoid model of INF2-associated FSGS that reliably recapitulated podocyte vulnerability to toxic injury. This platform demonstrated that calcineurin inhibitors, including the novel agent VCS, exert direct podocyte-protective effects in a genetic FSGS background and provide a practical system for mechanistic studies and therapeutic screening.
Introduction
Focal segmental glomerulosclerosis (FSGS) is a major cause of nephrotic syndrome (NS) and end-stage kidney disease (ESKD), accounting for ∼10% of patients with NS undergoing kidney biopsy in Japan. 1 Although FSGS and minimal change disease often present with similar clinical features and are initially treated with corticosteroids, FSGS is distinguished by a markedly lower remission rate and a high frequency of steroid resistance. Moreover, FSGS accounts for ∼10% of patients who progress to ESKD and require renal replacement therapy.
FSGS occurs in patients of all ages, races, and ethnic backgrounds, with a similar prognosis regardless of these factors.2,3 FSGS is a major cause of steroid-resistant NS and can be triggered by a wide variety of etiological factors. Accordingly, FSGS is now recognized not as a single disease entity but rather as a podocyte lesion resulting from diverse pathogenic mechanisms. While traditionally classified based on histological findings, recent approaches have integrated clinical and laboratory data to categorize FSGS into four subtypes: (i) primary, presumably immune-mediated; (ii) genetic; (iii) secondary; and (iv) undetermined. 4
Among these, genetic forms are a major cause of FSGS and often develop at a relatively young age, thus constituting a clinically significant disease. Currently, no curative treatment exists for genetic FSGS, and management is largely restricted to nephroprotective strategies to slow disease progression against the backdrop of gradually declining kidney function. In this study, we established an in vitro model of podocyte injury using induced pluripotent stem cells (iPSCs) derived from a patient with FSGS and NS caused by an inverted formin 2 (INF2) variant. 5 NS caused by FSGS resulting from an INF2 gene variant follows an autosomal dominant pattern of inheritance and has been reported to develop between the ages of 10 and 50 years. 6 The INF2 gene is involved in the actin cytoskeleton and plays a role in reinforcing the podocyte cytoskeleton. According to reports from Europe and the United States, this condition accounts for approximately 10%–15% of all autosomal dominant NS cases, making it the most common form. FSGS develops following podocyte dysfunction. Furthermore, immunosuppressive therapy is ineffective for hereditary FSGS, and no curative treatment is currently available. Therefore, it is necessary to consider strategies to protect podocytes and mitigate podocyte dysfunction. The use of puromycin aminonucleoside (PAN) as a model for podocyte dysfunction has been reported. 7 Calcineurin inhibitors (CNIs) have been reported to stabilize the podocyte cytoskeleton and improve clinical outcomes in FSGS,8,9 including hereditary forms of FSGS. Hence, we used the model established in this study to evaluate the potential podocyte-protective effects of CNIs.
Materials and Methods
Ethics
This study was approved by the Institutional Review Board of Nagoya University Hospital (Approval Number: 2019-0414-7). Written informed consent was obtained from all participants.
iPSC culture and differentiation
We generated a human iPSC (hiPSC) line (NUMNi002-A) derived from a patient with NS harboring an INF2 variant. The line was generated using peripheral blood mononuclear cells and episomal vectors (pCE-hOCT3/4 [#41813], pCE-hSK [#41814], pCE-hUL [#41855], pCE-mp53DD [#41856], and pCXB-EBNA-1 [#41857]; Addgene, USA). 5 As a control, we used the hiPSC line (NUMNi001-A). 10 The hiPSCs were cultured in human iPS medium (StemFit AK02N; Ajinomoto Healthy Supply Co., Inc.) in a 5% CO2/20% O2 atmosphere. The medium was changed three times weekly. iPSCs were passaged every 5–7 days in culture dishes coated with iMatrix (Matrixome). Kidney organoids were differentiated using the STEMdiff Kidney Organoid Kit (STEMCELL Technologies) according to the manufacturer’s protocol. Briefly, during passaging iPSCs, they were dissociated into single cells using 0.125% trypsin and seeded at 7.5 × 104 to 1.0 × 105 cells/well in a 24-well plate coated with Matrigel (Corning, 1:100). The next day, the iPSCs were overlaid with Matrigel, and the medium was replaced with StemFit AK02N. The following day, kidney organoid differentiation was induced by incubating the cells for ∼40 h in Kidney Basal Medium supplemented with Kidney Supplement SG. Subsequently, differentiation was continued for ∼18 days using Kidney Basal Medium supplemented with Kidney Supplement DM.
Immunohistochemistry of differentiated kidney organoids
Differentiated kidney organoids were fixed in 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) containing 0.1% Triton X-100 for 10 min. After fixation, the samples were blocked with 10% Blocking One (Nacalai Tesque) in PBS for 20 min. The organoids were then incubated with primary antibodies diluted in PBS and 0.1% Triton X-100 for 16–18 h at 4°C.
Following primary antibody incubation, the samples were washed twice with PBS and incubated with appropriate secondary antibodies diluted in PBS containing 0.1% Triton X-100 for 30 min at room temperature. Nuclear counterstaining was performed using 4′,6-diamidino-2-phenylindole (1:5000; Sigma). Immunofluorescence images were acquired using a BZ-800X microscope (Keyence).
The primary antibodies and concentrations were as follows:
Human podocalyxin antibody (R&D Systems), 1:100 Purified mouse anti-E-cadherin (BD Transduction Laboratories), 1:100 Lotus tetragonolobus lectin, biotinylated (Vector Laboratories), 1:100 Anti-Wilms tumor protein antibody (abcam), 1:500 Human nephrin antibody (R&D Systems), 1:50 Anti-alpha Actinin 4 antibody (abcam), 1:100
The secondary antibodies and concentrations were as follows:
Streptavidin, Alexa Fluor™ 488 conjugate (Invitrogen), 1:500 Donkey anti-goat IgG (H+L) cross-adsorbed secondary antibody, Alexa Fluor™ 555 (Invitrogen), 1:1000 Donkey anti-mouse IgG (H+L) highly cross-adsorbed secondary antibody, Alexa Fluor™ Plus 647 (Invitrogen), 1:1000 Goat anti-rabbit IgG (H+L) cross-adsorbed secondary antibody, Alexa Fluor 555 (Invitrogen), 1:500 Donkey anti-sheep IgG (H+L) cross-adsorbed secondary antibody, Alexa Fluor 488 (Invitrogen), 1:1000
Electron microscopy
Electron microscopy observation was outsourced to Tokai Electron Microscopy, Inc. Fixation and observation were performed according to their protocol. Briefly, samples were fixed with 2% PFA/PBS and 2% glutaraldehyde (GA) in 0.1 M phosphate buffer (Pb), pH 7.4, at 37°C and then placed in a refrigerator at 4°C. Thereafter, they were fixed with 2% GA in 0.1 M Pb at 4°C overnight. After fixation, the samples were rinsed three times with 0.1 M Pb for 30 min each, followed by post fixation with 2% osmium tetroxide in 0.1 M Pb at 4°C for 1 h. The samples were dehydrated through a graded ethanol series (50%, 70%, 90%, and 100%). The schedule was as follows: 50% and 70% for 15 min each at 4°C, 90% for 15 min at room temperature, and three changes of 100% for 30 min each at room temperature. The samples were transferred to resin (Quetol-812; Nisshin EM Co.) and polymerized at 60°C for 48 h. The blocks were ultra-thin sectioned at 70 nm using a diamond knife with an ultramicrotome (Ultracut UCT; Leica), and the sections were placed on Cu grids. They were stained with 2% uranyl acetate at room temperature for 15 min and then rinsed with distilled water, followed by secondary staining with Lead stain solution (Sigma-Aldrich) at room temperature for 3 min. The grids were observed using a transmission electron microscope (JEM-1400PlusEX; JEOL Ltd.) at an acceleration voltage of 100 kV. Digital images (3296 × 2472 pixels) were acquired using a CCD camera (EM-14830RUBY2; JEOL Ltd.).
Western blotting
Total protein from kidney organoids with or without PAN exposure was extracted using Blue Loading Buffer Pack (Cell Signaling Technology). Protein concentrations were determined using the Pierce BCA Protein Assay Kit (Thermo Fisher). Approximately 10 μg of protein lysates were separated by a 4%–12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and the proteins were transferred to a nitrocellulose membrane by wet blotting. Membranes were then incubated with primary antibodies, followed by compatible secondary antibodies. β-actin was used to normalize protein expression. Protein bands were visualized using ECL Western Blotting Detection Reagents (Thermo Fisher) and detected in a UV chamber. Band intensity was quantified Image Lab software (Ver.6.1, BioRad) and was normalized to β-actin levels.
The primary antibodies and concentrations were as follows:
beta-Actin (8H10D10) Mouse Monoclonal Antibody (Cell Signaling Technology), 1:1000 p53 (1C12) Mouse Monoclonal Antibody (Cell Signaling Technology), 1:1000 Cleaved Caspase-3 (Asp175) Antibody (Cell Signaling Technology), 1:1000 Phospho-Histone H2A.X (Ser139) (20E3) Rabbit Monoclonal Antibody (Cell Signaling Technology), 1:1000
The secondary antibodies and concentrations were as follows:
Anti-rabbit IgG, HRP-linked Antibody #7074 (Cell Signaling Technology), 1:5000 Anti-mouse IgG, HRP-linked Antibody #7076 (Cell Signaling Technology), 1:5000
Inducing podocyte injury
Kidney organoids were prepared, and PAN (Gibco) was added to the culture medium. Evaluation was performed after 48 h. The final PAN concentration in the medium was adjusted to a range of up to 30 µg/mL for evaluation.
Assessing the degree of impairment
Podocalyxin-positive areas were identified as podocytes and evaluated based on the podocalyxin fluorescence intensity. The mean fluorescence intensity of podocytes under non-impaired conditions was set to 100%, and the percentage reduction was assessed. Measurements were obtained from three locations per photograph using a minimum of five photographs. Statistical analysis was performed using the Kruskal–Wallis test, with significance set at P <0.05. Comparisons within each group were adjusted using the Bonferroni method. The Mann–Whitney U test was used to compare the strain derived from the healthy donor at each PAN concentration.
Cyclosporine and voclosporin treatment of kidney organoids
Cyclosporine A (CsA) (Sigma-Aldrich) and voclosporin (VCS) (Sigma-Aldrich) were dissolved in dimethyl sulfoxide (DMSO) (Nacalai Tesque) at a concentration of 8.3 mM. These compounds were added to the culture medium at a 1:1000 dilution, resulting in a final concentration of 8.3 μM. The control comprised DMSO diluted 1:1000 in the medium. CsA and VCS were added to the medium as a pretreatment 1 h before exposure in the kidney organoids. After pretreatment, the procedure was the same as that used for PAN-induced podocyte injury.
Results
Creation of kidney organoids from iPSCs
We previously established iPSCs from a patient with FSGS and harboring an INF2 variant. 5 Using this cell line, we generated kidney organoids. The organoids were generated over ∼3 weeks according to the manufacturer’s protocol (Fig. 1A). Immunofluorescent antibody staining revealed areas positive for podocalyxin, a podocyte marker (Fig. 1B). Additionally, we confirmed the expression of podocyte-specific proteins, such as WT1, nephrin, and α-actinin 4, by immunofluorescent antibody staining (Fig. 1B). Furthermore, we assessed the microstructure of foot processes by electron microscopy (Fig. 1C). These findings confirmed that the differentiated cells contained podocytes.
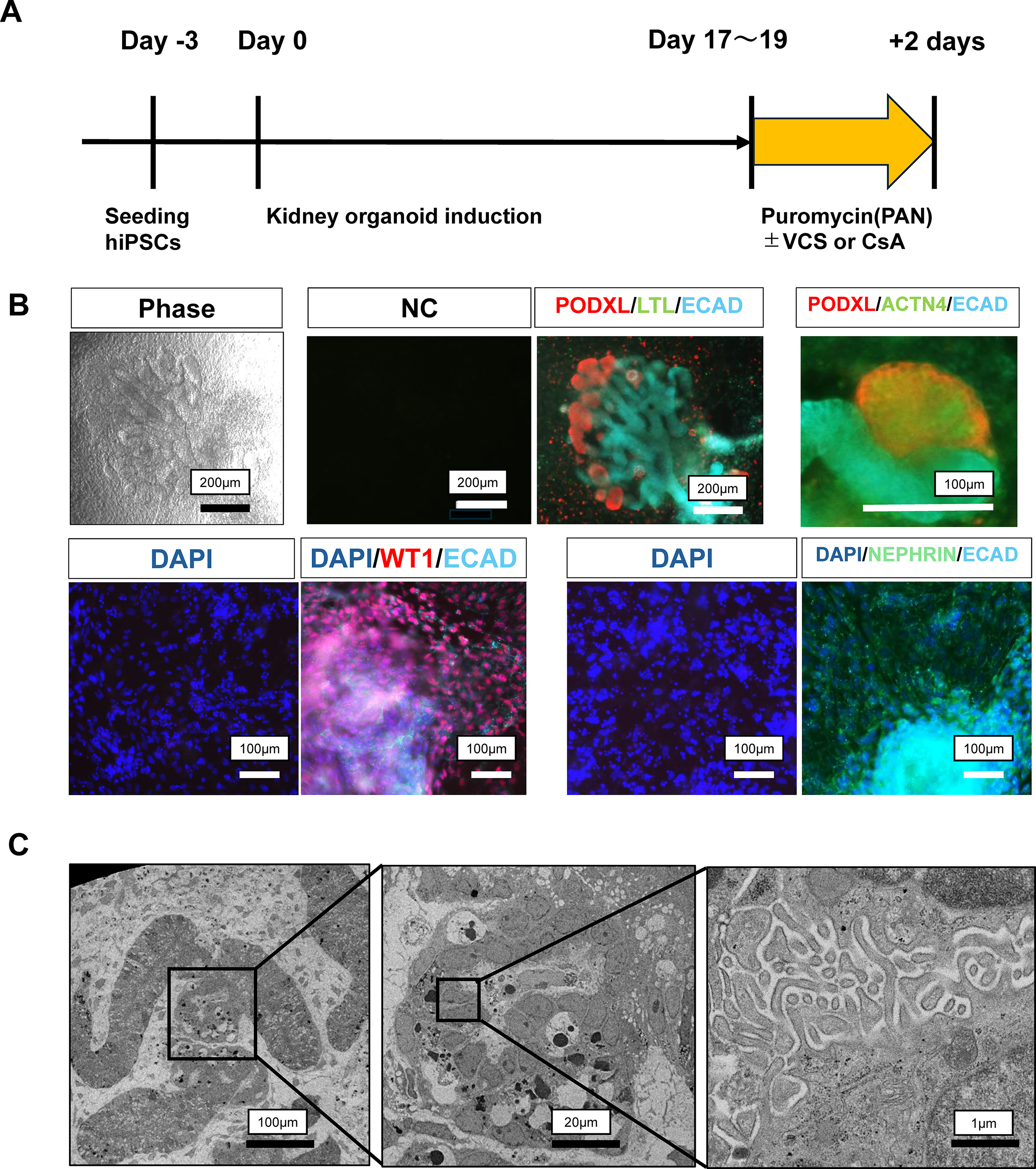
Induction of podocytes from patient-derived iPSCs.
Establishment of a model for PAN-induced podocyte injury
Podocyte injury was induced by adding PAN to the culture medium for 2 days (Fig. 1A). The mean fluorescence intensity of podocyte-positive areas decreased in a PAN concentration-dependent manner (Fig. 2A, B). At all concentrations from 3 to 30 μg/mL, the mean fluorescence intensity of podocalyxin was significantly reduced compared with that in the uninjured state (PAN 0 μg/mL). In addition, organoids derived from hiPSCs from healthy donors were used as controls (Supplementary Fig. S1). The organoids derived from patient-derived hiPSCs harboring the INF2 variant used in this study were more vulnerable to PAN-induced podocyte injury than those derived from healthy donors.
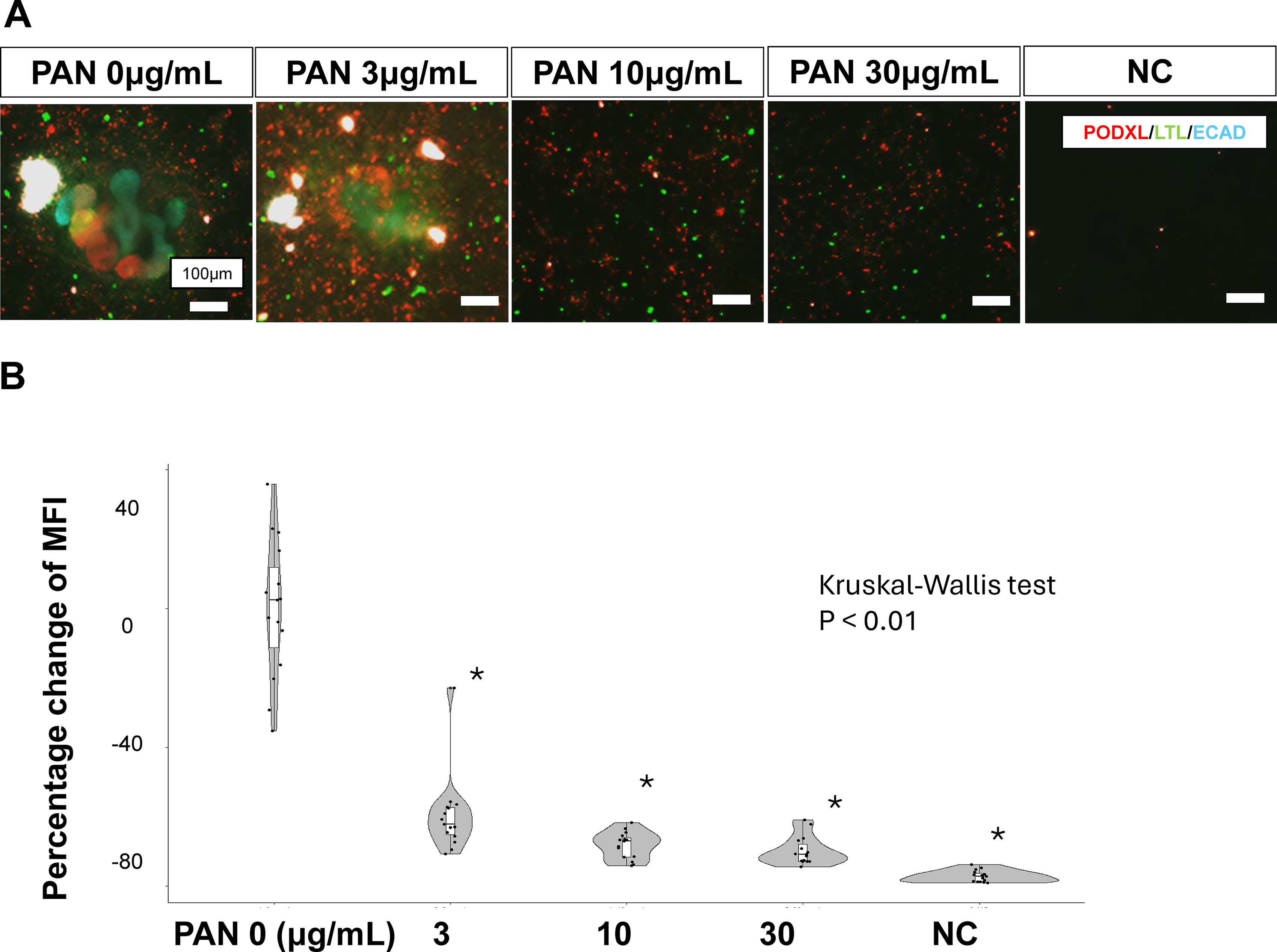
Concentration-dependent podocyte dysfunction in PAN.
Rescue using CNIs for podocyte impairment induced by PAN
After confirming PAN concentration-dependent podocyte injury, we evaluated whether CNIs (CsA and VCS) exerted podocyte-protective effects. Pretreatment with CsA or VCS before PAN (3 μg/mL) exposure reduced the decrease in mean fluorescence intensity (Fig. 3A, B), suggesting reduced cellular damage. Furthermore, based on a previous report, 7 we evaluated DNA damage, apoptosis, and p53 levels in organoids following PAN treatment (Fig. 3C). The results showed that, similar to the findings in Figure 3A and B, PAN-induced injury tended to be mitigated by CsA and VCS.
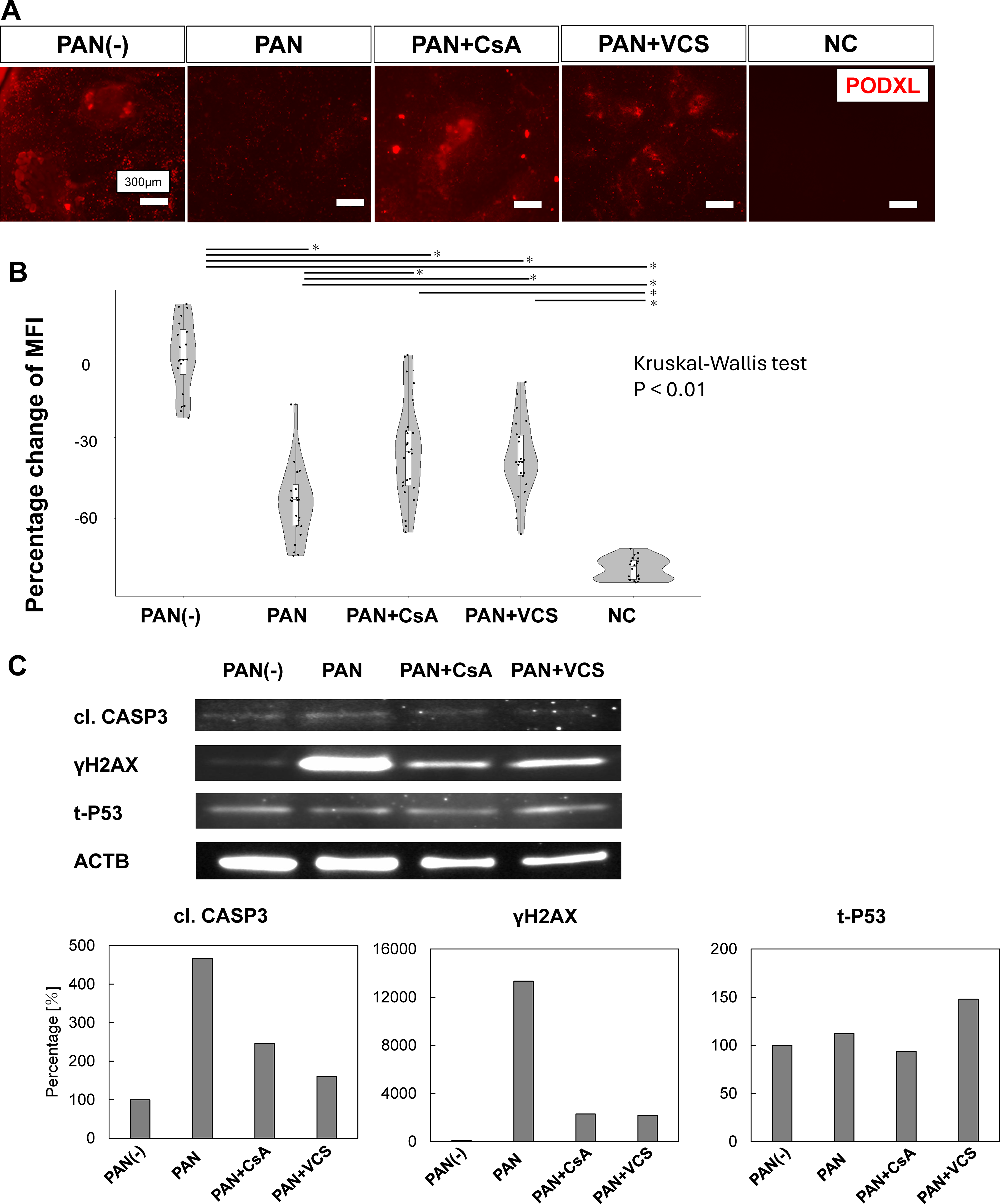
Reduced PAN-induced podocyte injury following CsA or VCS administration.
Discussion
In the present study, we established a model of podocyte injury using hiPS cells derived from a patient with FSGS harboring an INF2 variant. PAN administration in this model demonstrated concentration-dependent podocyte injury. Furthermore, using this system, we verified the protective effects of CNIs, which are sometimes used clinically, on podocytes. Additionally, our results suggest a podocyte-protective effect of VCS, a novel CNI.
Previous reports have described PAN-induced podocyte injury in kidney organoids and alleviation of this damage using CsA. 11 Other reports have demonstrated PAN-induced podocyte injury in animal models and human immortalized podocyte cell lines, as well as the effects of CsA in mitigating this damage.12,13 However, whether PAN could similarly induce podocyte injury, or whether rescue experiments were feasible in patients with INF2 variants, given their fragile cytoskeleton, remained unclear. This gap was addressed in the present study. Additionally, previous studies used slightly higher concentrations of PAN (e.g., 30 μg/mL or higher). Because the organoids used in those studies were three-dimensional, higher-order structures, drug penetration into the interior of the cell mass may have been limited, or cell–cell interactions may have influenced the response. In the present study, we observed cytotoxicity at a lower concentration than previously reported, possibly owing to easier drug penetration into the interior and weaker cell–cell interactions induced by two-dimensional organoid culture. Although three-dimensional organoids are advantageous for mimicking the kidney, the method proposed in the present study may be simpler and more useful for drug screening.
Furthermore, this study is also the first to examine the effects of novel CNIs such as VCS. VCS is currently covered by insurance for lupus nephritis in Japan, and its clinical efficacy has been reported.14,15 Moreover, a randomized controlled trial demonstrated that CsA significantly reduced proteinuria in patients with steroid-resistant FSGS, with 70% achieving partial or complete remission compared with only 4% in the placebo group, 9 presumably including patients with hereditary FSGS. The podocyte-protective effects of CsA have been attributed to improved cytoskeletal stability independent of immunological mechanisms. Clinically, its use has been reported in FSGS involving INF2 variants.16,17 Therefore, VCS was expected to provide similar podocyte-protective effects. Some reports have indicated that VCS possesses higher activity than CsA 18 ; however, clinically, it can be used without measuring its blood concentration. In the present study, the VCS concentration was determined based on reports in other diseases 19 ; however, the efficacy did not differ significantly between VCS and CsA. Although VCS exerted podocyte-protective effects, it is not a fundamental treatment targeting the cytoskeleton, which may explain why the results were similar to those observed with CsA.
Conclusions
We generated kidney organoids using iPSCs established from a patient with FSGS harboring an INF2 variant and confirmed the usefulness of the PAN-induced podocyte injury model in this setting. The degree of impairment increased with increasing PAN concentrations and was mitigated by CNIs, including VCS.
Footnotes
Acknowledgments
The authors thank the donor for participating in this study. They also thank the staff of the Department of Nephrology at Nagoya University for their assistance.
Ethical Considerations
This study was approved by the Institutional Review Board of Nagoya University Hospital (Approval Number: 2019-0414-7).
Consent to Participate
Written informed consent was obtained from all participants.
Data Availability
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. However, the authors are planning future clinical trials with Otsuka Pharmaceutical Co., Ltd., the distributor of VCS in Japan, and may receive financial support related to those studies.
Funding Information
This study was supported by a research grant from the Ministry of Education, Culture, Sports, Science, and Technology of Japan KAKENHI (grant no. 24K11429).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.