
Abstract
Most cellular and physiological activities are temporally organized, with many following robust circadian oscillations over the 24-h day. Mineralized dental tissues formation during odontogenesis provide an especially tractable model to study such temporal control, as they display clear daily patterns of cell behavior and matrix accretion. In this pilot study, the temporal dimension of odontogenesis was examined using two complementary circadian knockout (KO) mouse models that exhibit opposing chronotypes; Cryptochrome1 (Cry1) KO, characterized by a 1-h shortened circadian period, and Cryptochrome2 (Cry2) KO, characterized by a 1-h lengthened period. Our data revealed distinct chrono/genotype–phenotype coupling across the dental tissues examined. Cry2 targeting resulted in more pronounced changes with significantly augmented dentin/cementum deposition accompanied by constricted pulp chambers. In enamel, targeting the Cry genes resulted in less noticeable effects with an accelerated early enamel matrix deposition rate observed in Cry2 mutants, while enamel mineralization was slightly slowed down, whereas targeting Cry1 resulted in an opposite pattern, slowed down secretion, and precocious enamel mineralization. Collectively, our results strongly suggest that there is a correlation between circadian period length and dental matrix proteins deposition, whereby longer periods favor elevated secretion rates while shorter periods result in a slightly reduced output. On the other hand, no similar effects were noted in mineral content, as mature enamel, bone, and dentin mineral densities did not differ significantly across the two KO models. Overall, this work identifies the cryptochrome 2 gene as a potent regulator of dentinogenesis and mandibular bone remodeling with a limited impact on amelogenesis. It also highlights intrinsic biological time as a fundamental axis along which the architecture of mineralized craniofacial tissues is patterned.
Keywords
Introduction
Dental mineralized tissues are profoundly time-structured systems in which growth and differentiation proceed in a rhythmic rather than continuous manner. 1 Indeed, incremental markings in enamel (i.e., daily cross-striations) and in dentin (i.e., von Ebner’s lines) record successive growth fronts and demonstrate that matrix apposition is organized in discrete, circadian-scale increments rather than as a smooth gradient.1,2 These features have been used to reconstruct tooth formation rates, eruption timing, and life-history traits in both extant species and fossil vertebrates, underlining how odontogenesis is coupled to the 24-h cycle. 3 At the cellular and molecular levels, previous works by our group and others in rodent models and ameloblast-lineage cells have shown that multiple core clock components, including and brain and muscle ARNT-Like 1 (Bmal1), Clock, Period (Per)1 & 2, oscillate in ameloblasts at regular 24-h intervals at both RNA and protein levels.4–7 Together, these anatomical and molecular observations support the view that odontogenesis is an intrinsically rhythmic process governed by the circadian clock.
Within the molecular clockwork, Cryptochrome 1 (CRY1) and Cryptochrome 2 (CRY2) act as essential repressors in the negative limb, partnering with PER proteins to inhibit BMAL1-CLOCK activity and thereby close the core circadian transcription translation feedback loop.8–10 Genetic studies in mice and cellular clock reporters have established that Cry1 and Cry2 are not functionally redundant; deletion of Cry1 typically shortens the circadian period by roughly 1 h, whereas deletion of Cry2 lengthens it by a similar amount, creating “short-period” and “long-period” chronotypes, respectively.11,12 These chronotypes are reflected in locomotor behavior and in cell-autonomous rhythms in peripheral tissues.11–13 Beyond timekeeping, cryptochromes have been implicated in bone biology and metabolism.14,15 However, the Crys significance in other mineralized tissues and/or in the craniofacial bone complex has never been investigated.
Despite the strong evidence that odontogenic cells possess functional clocks, the gene-specific contributions of clock genes to odontogenesis remain essentially unexplored. Indeed, it remains largely unknown whether and how circadian disruption would translate into changes in dental formation, morphology, and architecture. Addressing this gap is crucial not only for basic understanding of dental development but also for emerging chronobiology-informed strategies in oral medicine and tissue engineering. In this context, the present study uses well-characterized Cry1 and Cry2 knockout (KO) mice as genetic models of short- and long-period chronotypes to interrogate the temporal control of odontogenesis. 11 Our central hypothesis is that misalignment of the intrinsic circadian period length will differentially impact odontogenesis dynamics across the enamel, dentin, and the periodontium, yielding gene- and chronotype-specific phenotypes.
Materials and Methods
Circadian knockout experimental models
Cry1 and Cry2 KO mice were purchased from Jackson laboratories (JAX, MA, United States; stock # 016186 and 016185). 12 All animal procedures were approved by the University of Saskatchewan (protocol# 20170014) and Universite Laval (protocol# 2023-1451) animal research ethics boards Successful targeting of the Cry genes was validated by PCR genotyping as we previously described. 10
X-Ray and micro-computed tomography analyses
Micro-Computed Tomography (micro-CT) parameters, region of interest (ROI) and analyses are detailed in Supplementary Data.
Scanning electron microscopy (SEM)
Cry2 KO and their wildtype counterparts were comparatively analyzed by SEM. Detailed in Supplementary Data.
Histomorphometry
We quantified the thickness/area of root dentin, acellular cementum (AC) and cellular cementum (CC) in the distobuccal roots of the first maxillary molar using Hematoxylin & Eosin (H&E) stained parallel sections. Detailed in Supplementary Data.
RNA extraction and quantitative PCR & immunohistochemistry (IHC)
The gene expression levels of amelogenin (Amelx), kallikrein 4 (Klk4), dentin silaophosprotein (Dspp), bone sialoprotein (Bsp), and alkaline phosphatase (Alpl) were assessed in enamel organ (EO) and dental mesenchymal cells. AMELX protein was assessed in incisor ameloblasts by IHC. Detailed in Supplementary Data.
Statistical evaluation
All data are presented as means ± standard error of mean (SEM). Differences were considered statistically significant if P < 0.05. Detailed in Supplementary Data.
Results
Circadian period misalignment effects on incisor growth and the different stages of amelogenesis
Upon gross inspection, Cry2 KO mice showed noticeably shorter mandibular incisors compared to their WT littermates, while Cry1 KO mice showed no apparent differences (Fig. 1A). To fully assess and quantify the lengths of the curved mandibular incisor’s different segments, we measured three linear distances on similarly oriented X-ray images in 7-week-old mice (n = 3/genotype): (1) from the EO follicle margin to the incisor tip (total incisor length), (2) from the EO margin to the alveolar bone edge (unerupted segment), and (3) from the alveolar bone edge to the incisor tip (erupted segment) (Fig. 1B,C). All three lengths were reduced in Cry2 KO mandibular incisors versus their WT and Cry1 KO counterparts (Fig. 1D). To evaluate the role of Cry genes in the temporal regulation of amelogenesis, we analyzed continuously growing mandibular incisors to capture all developmental stages within a single specimen as previously described16,17 (Fig. 1E). Enamel thickness and mineral density were measured to assess enamel deposition and biomineralization, respectively. The first 2 mm from the onset of mineralization (secretory stage) was analyzed at 0.2 mm intervals, followed by measurements at 3 and 5 mm (maturation stage) and 7 and 9 mm plus the incisor tip (fully mature enamel) 16 (Fig. 1F). Cry1 and Cry2 deletion produced opposing effects. Cry2 KO mice exhibited increased enamel matrix secretion but delayed initial mineralization during the secretory stage, whereas Cry1 KO showed reduced matrix deposition and increased mineralization, particularly within the first 1 mm (Fig. 1G,H). These differences diminished during later stages, although Cry2 KO enamel remained slightly thicker than WT and Cry1 KO (Fig. 1I,J).
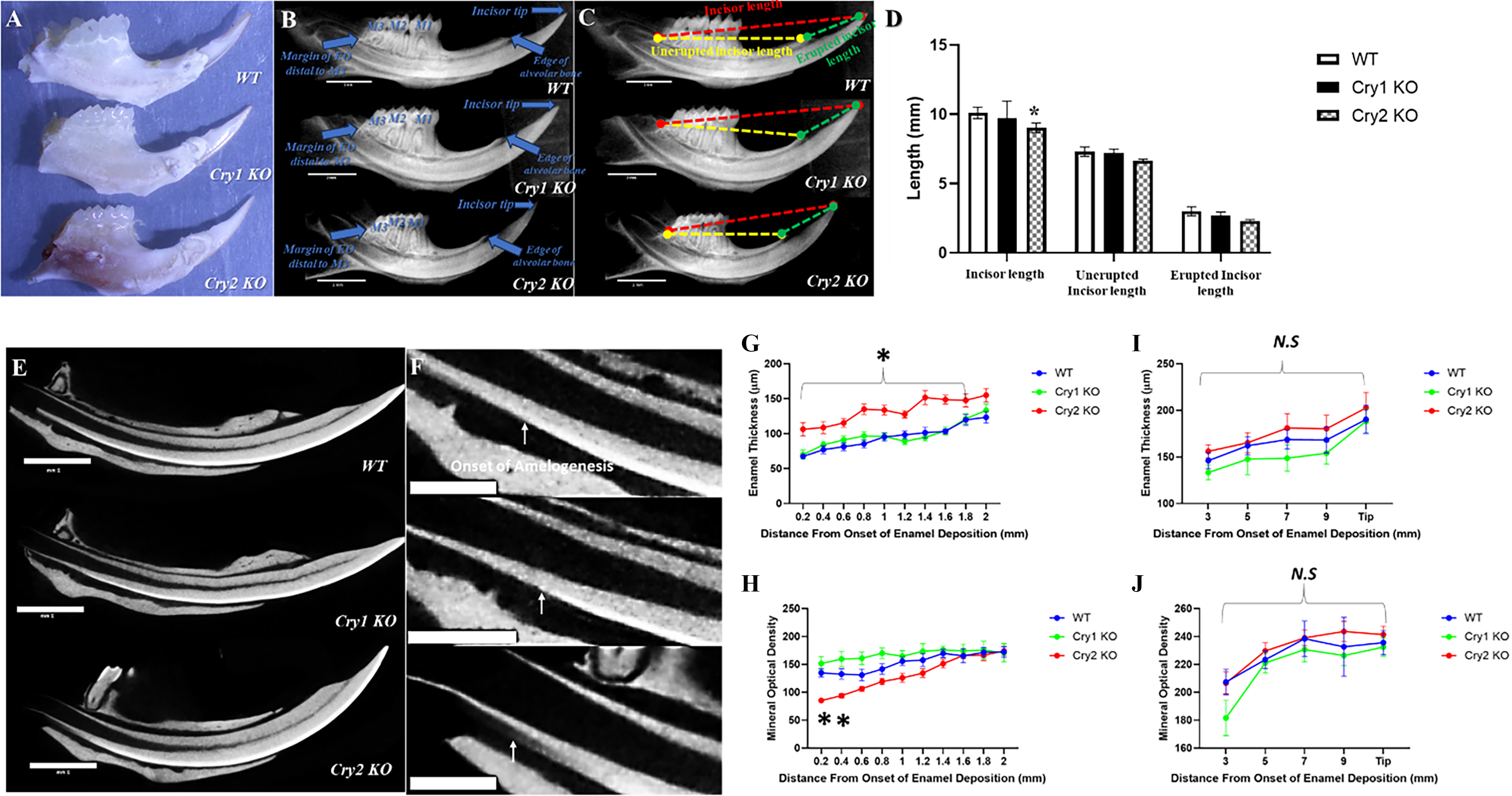
The differential effects of targeting the cryptochrome genes on incisors and the different stages of amelogenesis.
Circadian period misalignment effects on mature enamel
SEM imaging showed that targeting Cry 2 didn’t result in evident changes in the general shape or size of the molars with no clear surface defects observed when compared to WT (Fig. 2A). Cry2 KO molars showed similar enamel thickness as their WT littermates (Fig. 2B). Interestingly, however, a thicker layer of the highly mineralized and less porous aprismatic surface enamel was consistently observed in the Cry2 KO molars (Fig. 2B). At the ultrastructural level, higher magnification of the sectioned bulk enamel revealed a more organized prismatic arrangement with typical keyhole patterns in WT molars compared to the thinner and more compact prisms observed in Cry2 KO (Fig. 2B). In incisors, imaging of transverse ground sections of mandibular incisors cut at the level of the alveolar crest showed increased dentin and enamel thickness in Cry2 KO mice compared to WT, consistent with our micro-CT analyses (Fig. 2C). BSE imaging of the transversely cut enamel showed that WT mice enamel showed slightly lower mineral density with higher porosity than Cry2 KO mice, although this difference proved insignificant (Fig. 2D). Higher magnification of the surface of Cry2 KO knockout incisor enamel didn’t reveal any evident defects or significant differences compared to WT (Fig. 2E).
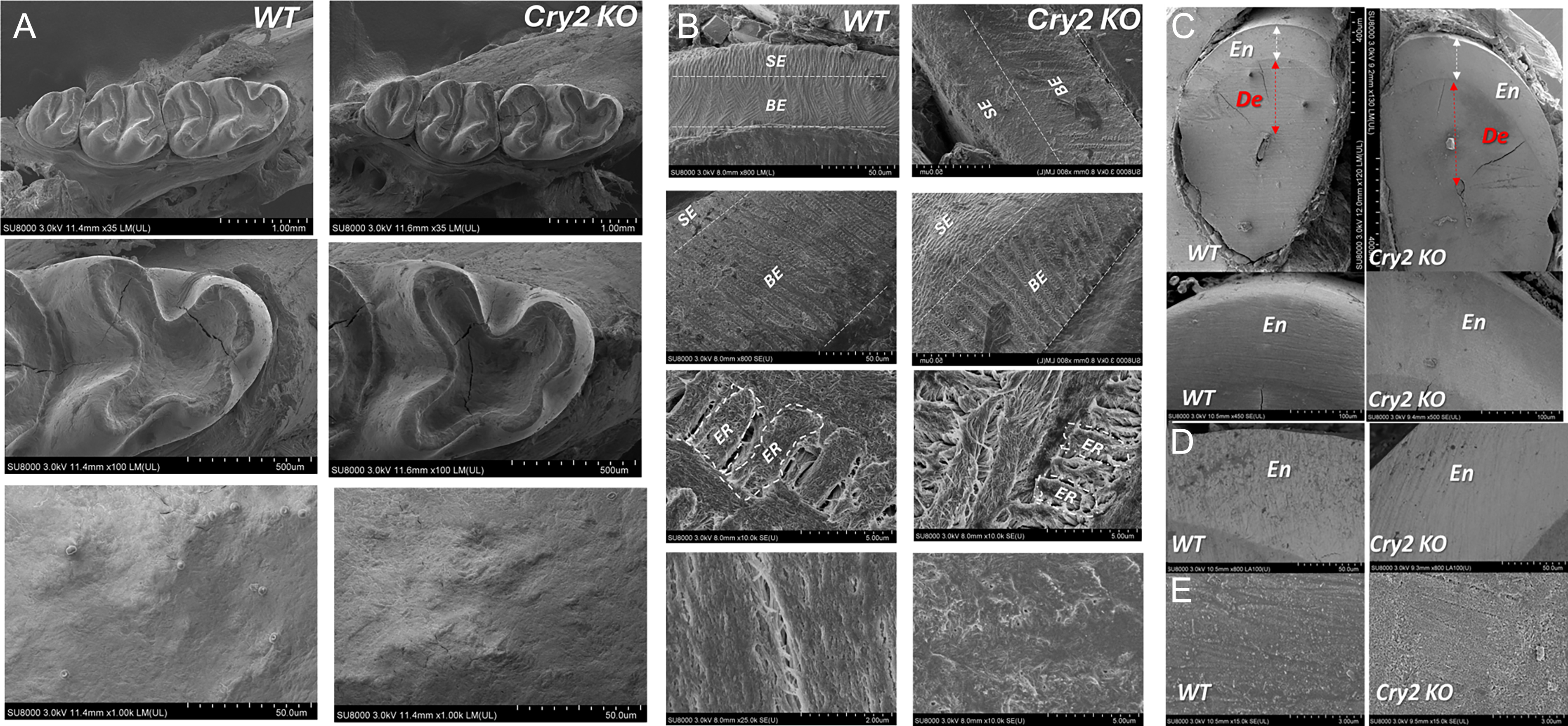
Molar enamel phenotype in Cry2 KO mice.
Circadian period misalignment effects on dentin, cementum, and mandibular bone
Our micro-CT analysis of the mandibular first molar ROI (n = 3/genotype, 7 weeks; Fig. 3A) revealed that cryptochrome gene deletion differentially affects dentin, pulp, and mandibular bone volumes. Cry2 KO mice showed more pronounced changes, including shorter roots and increased dentin volume, particularly in the incisor region (Fig. 3B). When normalized to total tooth volume (TV), Cry2 KO teeth exhibited significantly increased dentin (DV/TV) and reduced pulp space (PV/TV), whereas Cry1 KO showed the opposite pattern (Fig. 3C). With regard to bone, then targeting either of the Cry genes resulted in similar trends with reduced mandibular bone volume, shorter interradicular ridges, and decreased relative trabecular bone volume (BV/TV), with stronger effects in Cry2 KO (Fig. 3B,C). Histological analysis of parallel sections of the maxillary M1 further confirmed the micro-CT findings (i.e., sectioned at the level where the distobuccal root showed the largest portion of its apical cellular cementum) (Fig. 4A). Cry2 KO showed significantly thicker root dentin and larger amounts of apical cellular cementum when compared to both WT and Cry1 KO counterparts (Fig. 4B). Interestingly, the coronal pulp chambers and root canals of the Cry2 KO molars showed diffuse pulp calcification, which may further explain the constriction of pulp spaces noted in the micro-CTs, but this phenomenon was not consistent in all their molars (Fig. 4A). Finally, no significant changes were observed in dentin, alveolar, or basal bone mean mineral optical density across the three groups (Supplementary Fig. S2).
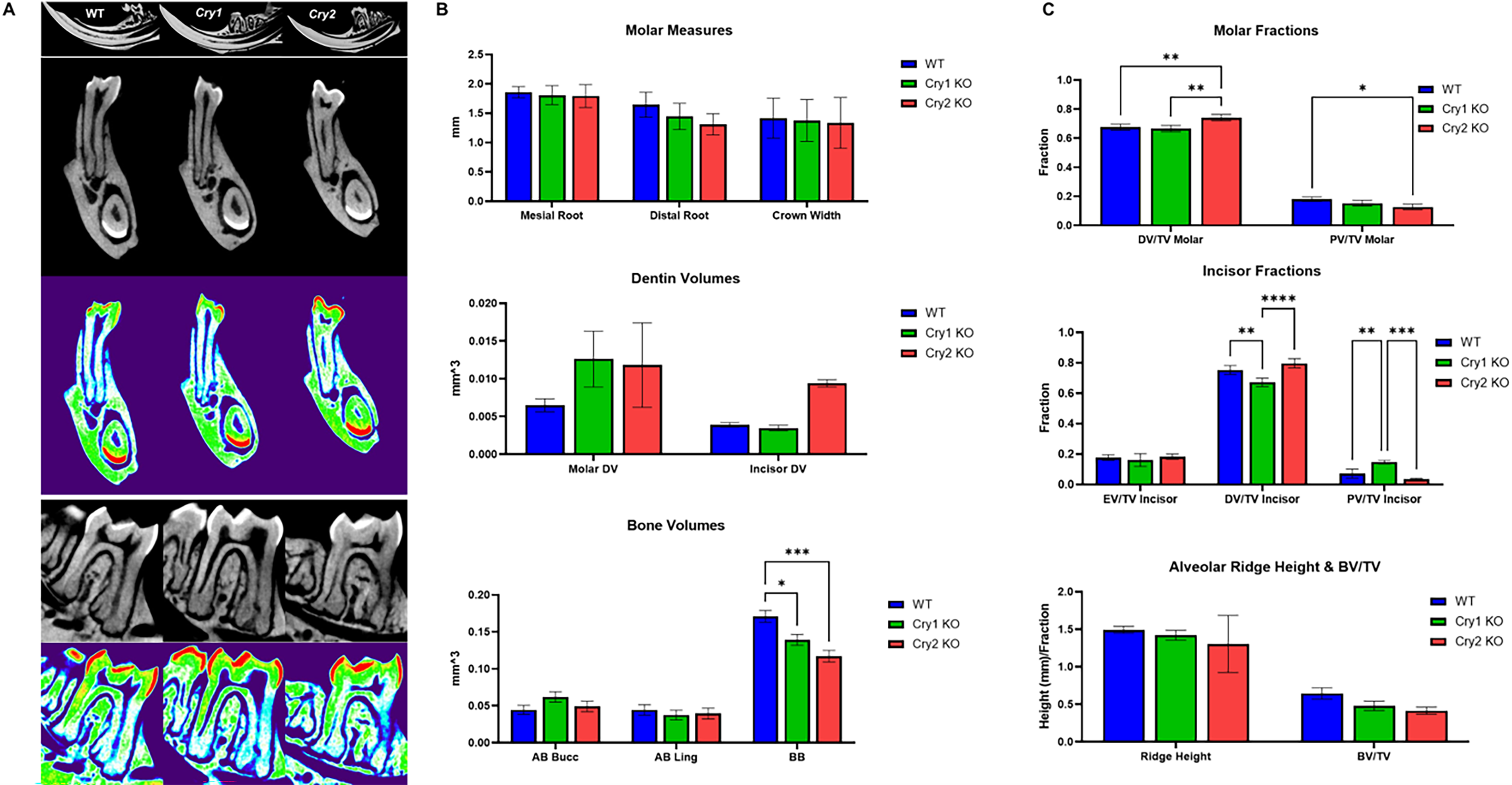
Effects of targeting the cryptochrome genes on dentoalveolar complex.
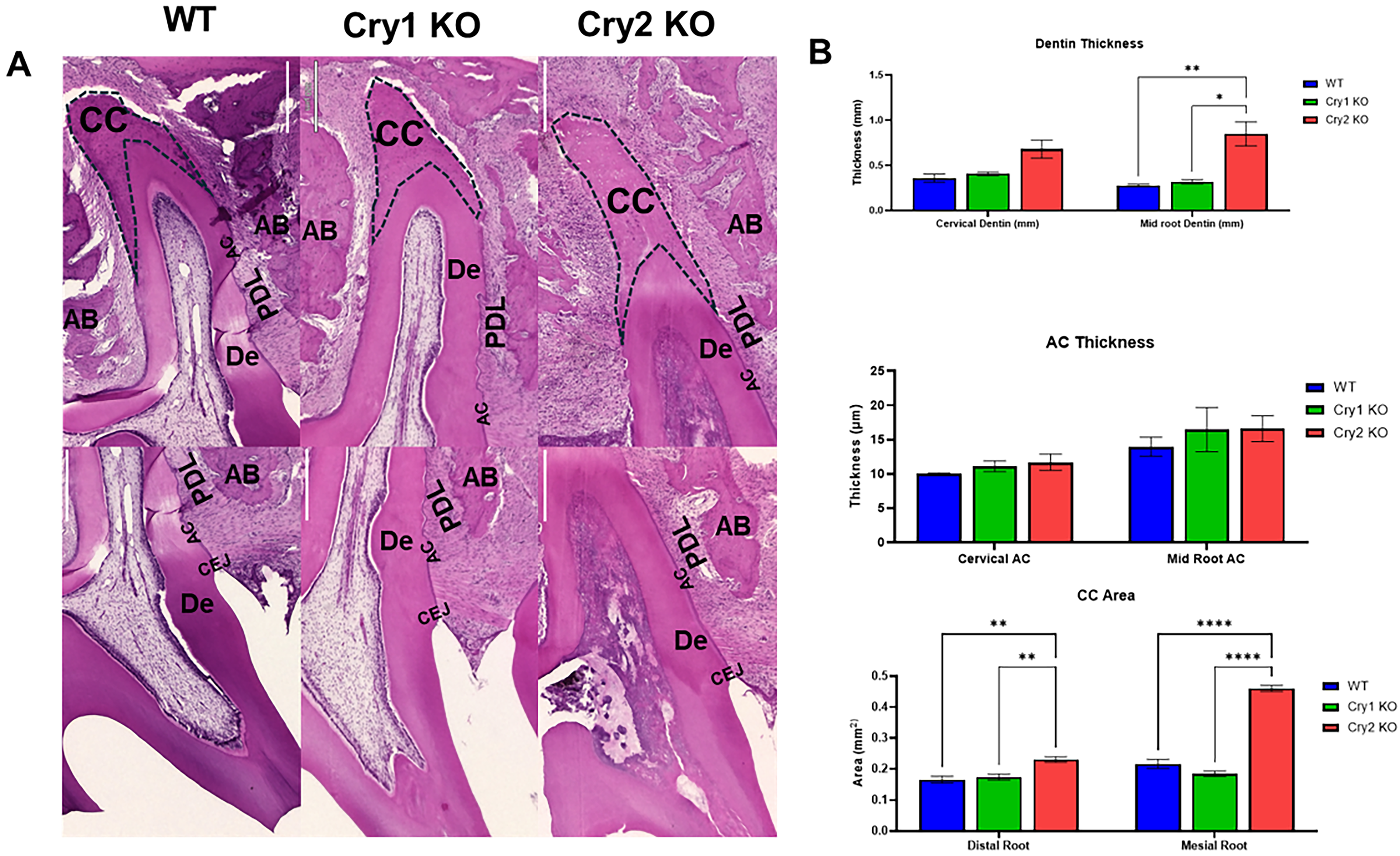
Chronotype-dependent alterations in root dentin, cementum, and inter-radicular alveolar bone.
Circadian period misalignment effects on ameloblasts and odontoblast functional markers
To determine Cry1 & 2 potential downstream molecular targets, we measured the gene expression levels of the main enamel matrix protein Amelx and the enamel proteinase Klk4 by RT-qPCR in enamel organ cells. RNA levels were quantified at both activity (i.e., 1:00 AM) and rest (i.e., 1:00 PM) circadian periods to assess any circadian variation (n = 2–3 per genotype, 12-week-old). Targeting cryptochrome genes did not result in any significant differences in the expression levels of any marker (Fig. 5A). IHC staining also did not show any significant changes in the protein signals of AMELX after targeting Cry2 (Fig. 5B,C). For odontoblasts, the dentin matrix protein Dspp, the odontoblast differentiation marker Bsp, and the dentin mineralization marker Alpl were examined in dental mesenchymal cells isolated from the maxillary molars dental pulp at the same activity and rest time points (n = 2–4 per time point, 12-week-old). Generally, Cry2 deficient cells showed higher levels of three markers that may explain the observed increased dentin deposition (Fig. 5D).
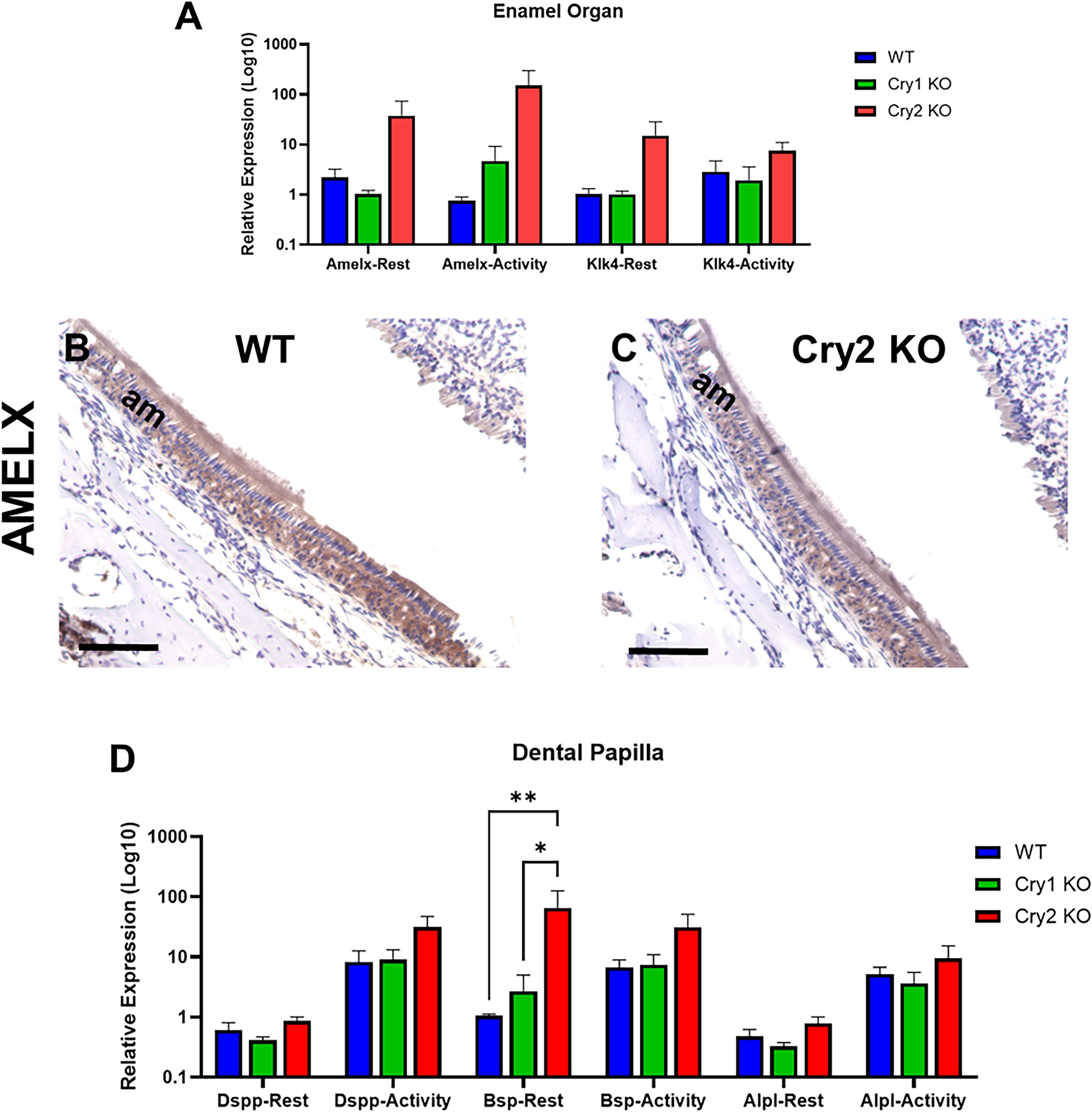
The effects of targeting Cry1 & Cry2 on ameloblast and odotoblasts functional markers.
Discussion
The present study provides the first in vivo evidence that targeting the core clock genes differentially influenced the architecture of the dentoalveolar complex, revealing a previously unappreciated temporal dimension in craniofacial mineralized tissue development. We demonstrate a subtle but clear chrono/genotype–phenotype coupling across enamel, dentin, cementum, and alveolar bone. The most prominent phenotype was observed in Cry2-deficient mice, which exhibited reduced mandibular bone volume, shortened roots with constricted pulp chambers, and enhanced dentin and cementum deposition, whereas enamel alterations were comparatively modest and largely restricted to early stages of amelogenesis. Importantly, despite the differences in tissue architecture, mineral density remained largely unchanged across genotypes, suggesting that circadian disruption primarily affects matrix deposition dynamics rather than final mineral content. These findings extend prior in vitro and molecular observations of circadian oscillations in ameloblasts and odontoblasts and position intrinsic biological times as an important but understudied regulator of craniofacial tissue patterning.
With respect to enamel, our data indicate that cryptochrome genes only exert subtle effects on amelogenesis. Cry2 deletion was associated with accelerated early enamel matrix secretion and delayed initial mineralization, whereas Cry1 deletion produced the opposite trend. These early differences largely dissipated in mature enamel, with SEM observations showing largely preserved enamel microarchitecture and noticeable changes noted in the expression patterns of ameloblasts markers. Collectively, our findings suggest that the cryptochrome genes do not appear to be a major ameloblast regulator, in contrast to what we previously showed with regard to the other clock genes; Bmal1 & Per2. Indeed, we previously showed that the diurnal variations of both AMELX and KLK4 protein expression levels closely mirror PER2 and BMAL1 in vitro.4,7 More specifically, production of both AMELX and KLK4 showed an inverse relationship with Per2 and a positive relationship with Bmal1 in the HAT-7 ameloblast cell line.2,4,7 Moreover, we recently showed that the ameloblasts in the arhythmic Per2 KO mice showed significantly upregulated AMELX and KLK4 expression associated with increased enamel deposition and hyper-mineralization (article under review). The relative lack of clear enamel phenotype changes in the Cry1 & 2 KOs compared to the robust alterations we observed in Per2 KO suggests that enamel formation may be buffered against circadian perturbations limited to period length but is more susceptible to arhythmic chronotypes. It must be noted, however, that the increased ratio of the aprismatic surface enamel noted in Cry2 KO mice supports a potential delayed transition from secretion to maturation, 16 which may indicate the that longer circadian periods resulted in elongated secretory phase.
Unlike enamel, dentin appeared to exhibit a robust and sustained response to circadian period disruption, particularly in Cry2-deficient mice. The observed increase in dentin thickness, coupled with constricted pulp chambers, suggests enhanced odontoblast activity and/or prolonged dentin apposition. The concomitant increase in CC further suggests coordinated regulation of root-associated mineralized tissues, potentially via shared mesenchymal progenitor populations or common signaling pathways influenced by circadian timing (e.g., RUNX2). The stronger impact of targeting Cry2 on dentin and cementum compared to enamel could be attributed to the fact that dentinogenesis and cementum apposition proceed throughout life, so changes in circadian period or amplitude can integrate over long time windows into measurable increases effects. Whereas ameloblasts are less able to “accumulate” a similar chronic circadian-period effect after eruption. Odontoblasts, like ameloblasts, form dentin incrementally, with short- and long-period growth lines visible in the dentin matrix.18,19 The amount of collagen deposited by odontoblasts is approximately twofold higher during the light phase than during the dark phase, indicating strong daily rhythmicity in matrix production. 20 The impact of circadian disruption on dentin structure remains unclear, however, as lesion studies of the suprachiasmatic nucleus (SCN) have yielded conflicting results regarding the persistence of daily dentin growth lines.21,22 At the molecular level, very little is known about the molecular circadian mechanisms that regulate dentin formation. We previously showed that the core clock genes are expressed in odontoblasts during tooth development in parallel with ameloblasts, and BMAL1 has been shown to promote dentinogenic differentiation of dental pulp stem cells under mild inflammation but inhibit it under severe inflammatory conditions.6,23 Clock gene expression has also been demonstrated in cementoblasts and cementocytes.2,24 A recent report by Liu et al., 2023 showed that pharmacological circadian modulation regulated cementoblast proliferation and mineralization in vitro, supporting a direct role for local peripheral clocks in cementum. 24 Collectively, our results here and the abovementioned reports strongly suggest that dentin and cementum formation are under robust circadian control, and this temporal regulation may be essential for maintaining tooth integrity throughout the lifespan.
The mandibular bone phenotype observed in both Cry1 & 2 KO mice, characterized by reduced absolute and fractional bone volumes, highlights an important and common role for both cryptochromes in craniofacial bone remodeling. These findings align with previous studies demonstrating a similar impact of both cryptochrome genes on bone remodeling. The study by Fu et al. 2005 was the first to show altered bone mass and osteoblastic activity in Cry1/2-deficient mice vertebrae. 25 A subsequent study by Maronde et al., 2010 showed that Cry2 deletion altered vertebral and tibial bone mass with lower levels of osteoclastic serum markers. 14 Generally, the circadian regulation of osteogenesis is well documented, with osteoblasts rhythmically expressing all the canonical clock genes and ∼26% of the osteoblast functional genes display a diurnal expression pattern.14,26 Samsa et al., 2016 reported that loss of Bmal1 leads to reduced osteoblast activity and decreased bone mass. 15 Xu et al., 2016 found that mice lacking Bmal1 in osteoclasts instead display increased bone mass due to impaired osteoclast differentiation. 27 Other studies also showed that the pineal circadian hormone melatonin can modulate osteoblast and osteoclast proliferation.28,29 Our findings here further extend these observations to the dentoalveolar context and reinforce the significance of clock genes in regulating bone turnover. The reduction in bone volume despite increased dentin and cementum deposition suggests a decoupling of osteogenesis and odontogenesis under circadian disruption. This may reflect differential sensitivity of osteoblasts and odontoblasts to clock gene perturbation, or distinct regulatory inputs from systemic versus local clocks.
A central question arising from this work is whether the observed phenotypes are driven primarily by the altered chronotype, by gene-specific functions of CRY1 and CRY2, or by circadian-dependent systemic metabolic changes. While the opposing trends of Cry1 and Cry2 KO in enamel & dentin strongly support a chronotype-dependent mechanism, where longer periods favor increased matrix deposition and shorter periods constrain it, this interpretation is likely incomplete. Cryptochromes are known to have pleiotropic roles beyond timekeeping, including modulation of NF-κB signaling. 30 It is well established that NF-κB signaling contributes to tooth and craniofacial development, with Ectodysplasin/NF-κB required for odontogenesis and cusp formation, and NF-κB activity influencing osteogenic cells involved in craniofacial bone repair. 31 Moreover, both CRYs have been shown to interact with glucocorticoid signaling and metabolic pathways, raising the possibility that systemic hormonal or metabolic alterations contribute to the observed phenotypes. 32 The lack of significant changes in gene expression of enamel and dentin markers further suggests that the systemic mechanisms may play a more dominant role, while local transcriptional regulation may be preserved or not largely affected. Finally, the more pronounced effects in Cry2 KO compared to Cry1 KO may reflect that Cry2 plays stronger and more critical roles in odontogenic lineages. Another study by Lowe et al., 2018 showed similar patterns in mesenchymal myogenic cells where targeting Cry2 has led to impaired regeneration and weakened rhythmic expression of cell-cycle and differentiation genes, whereas Cry1 deficient skeletal muscle cells showed relatively preserved rhythms. 33 Collectively, it’s safe to deduce that the phenotype likely arises from a complex interplay between intrinsic period length, gene-specific regulatory functions, and systemic circadian misalignment. Future studies should utilize tissue-specific KOs to disentangle systemic versus local clock effects.
In conclusion, this study establishes clock gene-dependent circadian timing as a regulator of dentoalveolar tissue development. Our findings suggest that dentinogenesis in particular appears to be highly sensitive to alterations in intrinsic period length time. These insights have important implications for chronobiology-informed approaches in dentistry and craniofacial regeneration, including the potential optimization of treatment timing and the design of clock-targeted therapeutics. However, several limitations should be acknowledged, including the small sample size, the use of only male mice, and the limited temporal resolution of gene expression analyses. Future studies should incorporate longitudinal designs, higher-frequency sampling, and dental tissue-specific KOs.
Statements and Declarations
All authors have approved the submitted version of the article. All authors have read and approved the final version to be published and take responsibility for the affirmations regarding article submission, and the integrity of the data presented. The authors minimally used an AI-assisted language model (Perplexity, powered by GPT-5.1) to support language editing and refinement of the article draft.
Ethical Considerations
All animal procedures were approved by the Animal Care and Use Committees/Ethics Boards at the University of Saskatchewan (USASK) (protocol# 20170014) and Université Laval (ULaval) (protocol# 2023-1451) and were conducted in accordance with national and institutional guidelines for the care and use of laboratory animals.
Data Availability
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request. The full reconstructed micro-CT image stacks from the 7-week-old WT Cry1 & Cry2 KO mice (4 hemimandibles per genotype) have been deposited in Zenodo (DOI: https://doi.org/10.5281/zenodo.20336721). Processed quantitative imaging data can be regenerated from these primary files using the workflow described in the Methods. The qPCR datasheets containing the derived relative quantities (RQ) for all analyzed genes of interest, as well as a summary spreadsheet with micro-CT and histology processed quantitative imaging data are provided as research data files in the journal submission system and will be made available with the published article as Supplementary Data.
Supplemental Material
sj-docx-1-stc-10.1177_15473287261466918 — Supplemental material for The Effects of Circadian Period Misalignment on Odontogenesis
Supplemental material, sj-docx-1-stc-10.1177_15473287261466918 for The Effects of Circadian Period Misalignment on Odontogenesis by Raed Said, Helya Mortazavi, Ibrahim Hoja, David Cooper, Silvana Papagerakis, and Petros Papagerakis
Footnotes
Author Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
This research was funded by the University of Saskatchewan’s VPFR-Centennial Enhancement Chair in One Health grant (PP), a Discovery Grant from Natural Sciences and Engineering Research Council of Canada (NSERC); PP, Grant ID: RGPIN-2022–04617 and a Team Research Project from Fonds de recherche du Québec – Nature et Technologies (FRQNT) (PP & SP).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.