
Abstract
Elevated levels of Bicaudal-D Family Like Cargo Adaptor 1 (BICDL1) are associated with poor prognosis in various cancers. However, the role of BICDL1 in breast cancer (BC) has not been reported. We analyzed BICDL1 expression in BC using bioinformatics and molecular experiments. Gene Set Enrichment Analysis was conducted to identify significantly enriched signaling pathways related to BICDL1. The effects of BICDL1 on BC cell proliferation, migration, invasion, and expression of epithelial-mesenchymal transition (EMT)-related proteins were assessed using colony formation assays, Transwell assays, and Western blot. The influence of BICDL1 on pyroptosis in BC cells was determined by measuring the expression of key pyroptosis proteins (NLRP3, GSDMD-N, procaspase-1, cleaved caspase-1, ASC), lactate dehydrogenase release, and cytokines (IL-1β and IL-18). We also investigated whether BICDL1 promotes BC proliferation and metastasis by regulating pyroptosis using Nigericin, an agent that induces NLRP3 inflammasome activation. The results showed that BICDL1 was significantly overexpressed in BC tissues and cells. Knockdown of BICDL1 inhibited BC cell proliferation, migration, invasion, and the EMT process, while overexpression of BICDL1 had the opposite effects. BICDL1 was enriched in the pyroptosis pathway; overexpression of BICDL1 suppressed pyroptosis, thus promoting BC cell proliferation, inhibition, and migration. Notably, the addition of Nigericin reversed the effects of BICDL1 overexpression on BC cells. These results suggested that BICDL1 promoted BC cell proliferation, inhibition, and migration by hindering pyroptosis. These findings indicate that BICDL1 is a potential biomarker for BC treatment.
Introduction
Breast cancer (BC) ranks as the most prevalent cancer among women worldwide, posing a significant threat to their health. 1 Although survival rates for BC patients have improved due to notable advancements in research and treatment, metastasis and recurrence remain substantial barriers to patient survival.2,3 Investigating important treatment targets and molecular processes underlying BC metastasis and recurrence is crucial for improving patient outcomes.
Aging, infected, or damaged cells undergo programmed cell death at designated times, a process essential for maintaining homeostasis. However, in varying pathological conditions, including cancer, this cell death becomes dysregulated, exacerbating disease progression. 4 Resistance to cell death is recognized as a hallmark of cancer, allowing malignant cells to proliferate and metastasize. 5 Therefore, inducing cell death is considered a promising strategy in cancer therapy. 6 Pyroptosis, a newly recognized form of inflammatory programmed cell death initiated by caspase-1/4/5/11, has garnered attention in cancer research. 7 As research advances, it has become increasingly clear that pyroptosis is intricately linked to cancer initiation and progression. For example, inducing pyroptosis hampers growth and metastasis in colorectal cancer, 8 nonsmall cell lung cancer, 9 and hepatocellular carcinoma. 10 Likewise, activating pyroptosis in BC also suppresses tumor progression. 11 Overall, the potential of pyroptosis in oncology has garnered extensive interest, yet its regulatory mechanisms in BC remain incompletely understood and warrant further investigation.
Bicaudal-D Family Like Cargo Adaptor 1 (BICDL1) is a member of the BICD family, which is primarily responsible for facilitating intracellular vesicle and organelle transport, playing a vital role in secretion and cell division. 12 Members of the BICD family are implicated in cancer progression; for instance, BICD1 is a prognostic biomarker in glioma, with its downregulation associated with improved patient outcomes. 13 However, research on BICDL1 in cancer remains limited, with only a study by Luo et al. 14 linking BICDL1 to poor prognosis and immune infiltration in colorectal cancer patients. Therefore, it is essential to explore the role of BICDL1 in BC.
This study identified BICDL1 as a gene significantly upregulated in BC and a key factor promoting BC proliferation and metastasis. This effect was mediated through the inhibition of pyroptosis. These findings demonstrated the functional and molecular regulatory mechanisms of BICDL1 in BC, offering new insights into the understanding of tumor proliferation and metastasis.
Materials and Methods
Bioinformatics analysis
mRNA expression data for adjacent (n = 113) and BC tumor tissues (n = 1117) were sourced from TCGA (https://portal.gdc.cancer.gov/). Differential analysis was performed using the edgeR package (|logFC| > 2, FDR <0.05) to identify differentially expressed genes, particularly BICDL1. Gene Set Enrichment Analysis (GSEA) was conducted to investigate signaling pathways associated with BICDL1. Pearson correlation analysis evaluated relationships between BICDL1 and pyroptosis markers NLRP3 and caspase-1 (CASP1).
Cell culture
Human normal breast epithelial cells (MCF 10A) and human BC cell lines (MCF-7, MDA-MB-453, and MDA-MB-468) were obtained from SUNNCELL (China). MCF 10A cells were cultured in a specific medium designed for them (SUNNCELL, China), while MCF-7 cells were maintained in their respective medium (SUNNCELL, China). MDA-MB-453 and MDA-MB-468 cells were grown in Dulbecco’s modified Eagle medium (Gibco, USA) with 10% fetal bovine serum (FBS) (Gibco, USA) and 1% penicillin-streptomycin (Gibco, USA). All cell lines were incubated at 37°C in a 5% CO2 atmosphere.
Cell transfection and treatment
si-BICDL1, oe-BICDL1, and their negative controls were designed and synthesized by GenePharma (China). Transfections were completed with Lipofectamine 3000 reagent (Thermo Fisher, USA) when cell density reached approximately 70%–80%. Cells were incubated for 24 hours before proceeding with further experiments. Nigericin was acquired from MCE (USA) and applied at 2 μg/mL for 24 hours in subsequent experiments. 15
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was extracted from cells with Trizol reagent (Beyotime, China), and cDNA was synthesized with the GeniuScript III Select RT Kit for qPCR (UGBIO, China). qPCR reactions were conducted using uGreener Flex qPCR 2X Mix (UGBIO, China) on an ABI7500 fluorescent qPCR instrument (Thermo Fisher, USA). After the reaction was completed, the expression levels of the target genes were normalized using GAPDH as the internal reference. The specific steps were as follows: First, the Ct values of the target genes and GAPDH were measured by qPCR. Then, the ΔCt values of the target genes and GAPDH were calculated (ΔCt = Ct target gene—Ct GAPDH). Next, the ΔΔCt values were calculated (ΔΔCt = ΔCt experimental group—ΔCt control group). Finally, the relative expression levels of the target genes in the experimental group compared to the control group were calculated using the 2−ΔΔCt formula. Primer (Tsingke, China) sequences are listed in Table 1. To ensure the reproducibility of the experimental data, this experiment was conducted with three independent replicates.
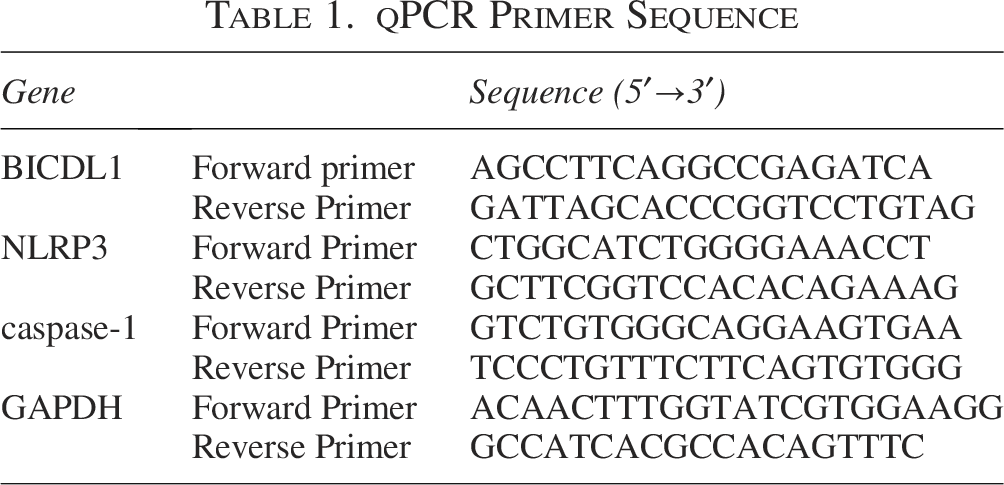
qPCR Primer Sequence
Western blot (WB)
Cells from each treatment group were collected, and total protein was extracted using Western and IP cell lysis buffer (Beyotime, China). Protein concentration was assessed with a bicinchoninic acid protein assay kit (Beyotime, China). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels (Epizyme, China) were employed to separate proteins, which were transferred to PVDF membranes (Millipore, USA). The membranes were blocked with 5% nonfat milk for 1 hour, then incubated with primary antibodies at 4°C overnight. Following this, membranes were treated with secondary IgG antibodies at room temperature for 1 hour. Bands were visualized with an enhanced chemiLuminescence chemiluminescent reagent kit (Beyotime, China). Detailed information on the antibodies used is provided in Table 2. To ensure the reproducibility of the experimental data, this experiment was conducted with three independent replicates.
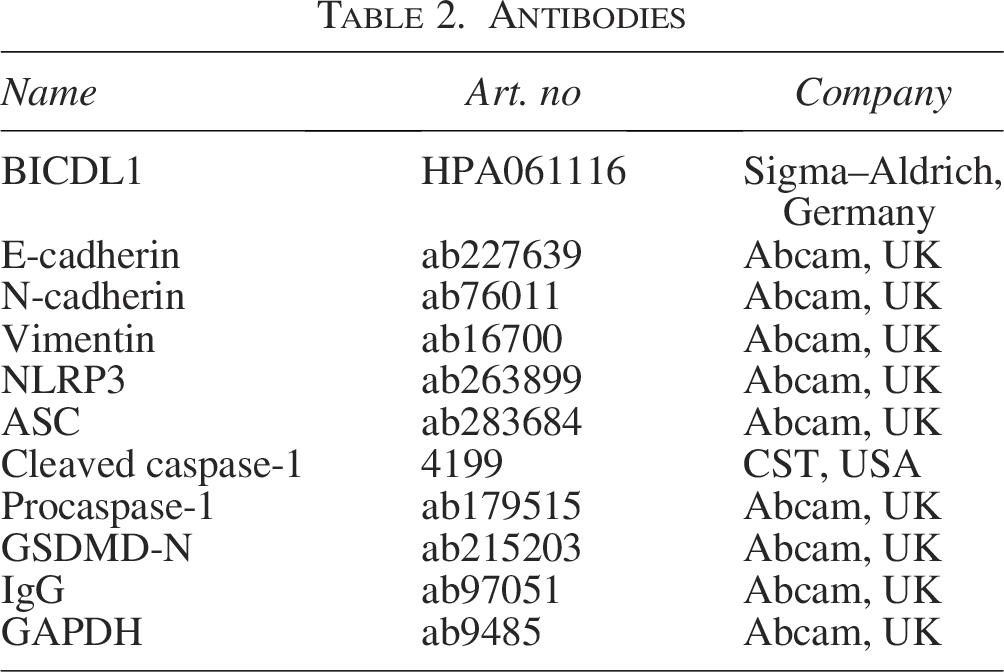
Antibodies
Colony formation assay
Cells were seeded into 12-well plates with 2 mL of complete medium (200 cells per well) and cultured for 7 to 14 days. After fixation with 75% ethanol for 30 minutes, cells were stained with crystal violet solution (Beyotime, China) for 30 minutes. The staining solution was carefully removed, and wells were gently rinsed with distilled water. After drying, colonies were photographed using a Nikon camera (Japan) and counted. To ensure the reproducibility of the experimental data, this experiment was conducted with three independent replicates.
Transwell assay for cell migration and invasion
Transwell chambers with an 8 μm pore size (Corning, USA) were utilized for migration and invasion assays. The lower chamber was precoated with Matrigel (Corning, USA) for invasion assays. 1 × 104 cells were suspended in 200 μL of serum-free medium and placed in the upper chamber, while 600 μL of medium containing 10% FBS was added to the lower chamber. Nonmigrated and noninvaded cells in the top chamber were carefully removed using a cotton swab after a 24-h incubation period at 37°C. Cells adhering to the lower surface of the membrane were fixed with 75% ethanol and stained with crystal violet solution for 30 minutes. The staining solution was carefully removed, and wells were rinsed with distilled water. Subsequently, they were photographed and counted under a microscope. To ensure the reproducibility of the experimental data, this experiment was conducted with three independent replicates.
Lactate dehydrogenase (LDH) cytotoxicity assay
LDH cytotoxicity assay used an LDH cytotoxicity detection kit (Beyotime, China). Absorbance was tested at 490 nm with a microplate reader (Thermo Fisher, USA), and cytotoxicity levels of LDH in each group were calculated based on the provided formula per instructions. To ensure the reproducibility of the experimental data, this experiment was conducted with three independent replicates.
Detection of IL-1β and IL-18 levels
Human IL-1β and IL-18 levels were quantified using enzyme-linked immunosorbent assay kits (Elabscience, China). Absorbance was tested at 450 nm with a microplate reader, and concentrations of IL-1β and IL-18 in the supernatants of each group were determined by constructing standard curves. To ensure the reproducibility of the experimental data, this experiment was conducted with three independent replicates.
Data analysis
All experiments were conducted in triplicate; data are presented as mean ± standard deviation. Data were analyzed on GraphPad Prism 8.0, employing t-tests or one-way ANOVA to compare differences between experimental groups. A p value <0.05 was deemed statistically significant.
Results
BICDL1 is overexpressed in BC
To assess BICDL1 expression in BC, a bioinformatics approach was applied to mRNA expression data from TCGA, comparing tumor tissues with adjacent normal tissues. As illustrated in Figure 1A BICDL1 expression was significantly elevated in BC tissues. Further analysis using qRT-PCR and WB revealed a significant increase in BICDL1 expression in BC cells (MCF-7, MDA-MB-453, and MDA-MB-468) relative to normal breast epithelial cells (MCF 10A) (Fig. 1B–C). Collectively, these findings confirm that BICDL1 is highly expressed in BC.
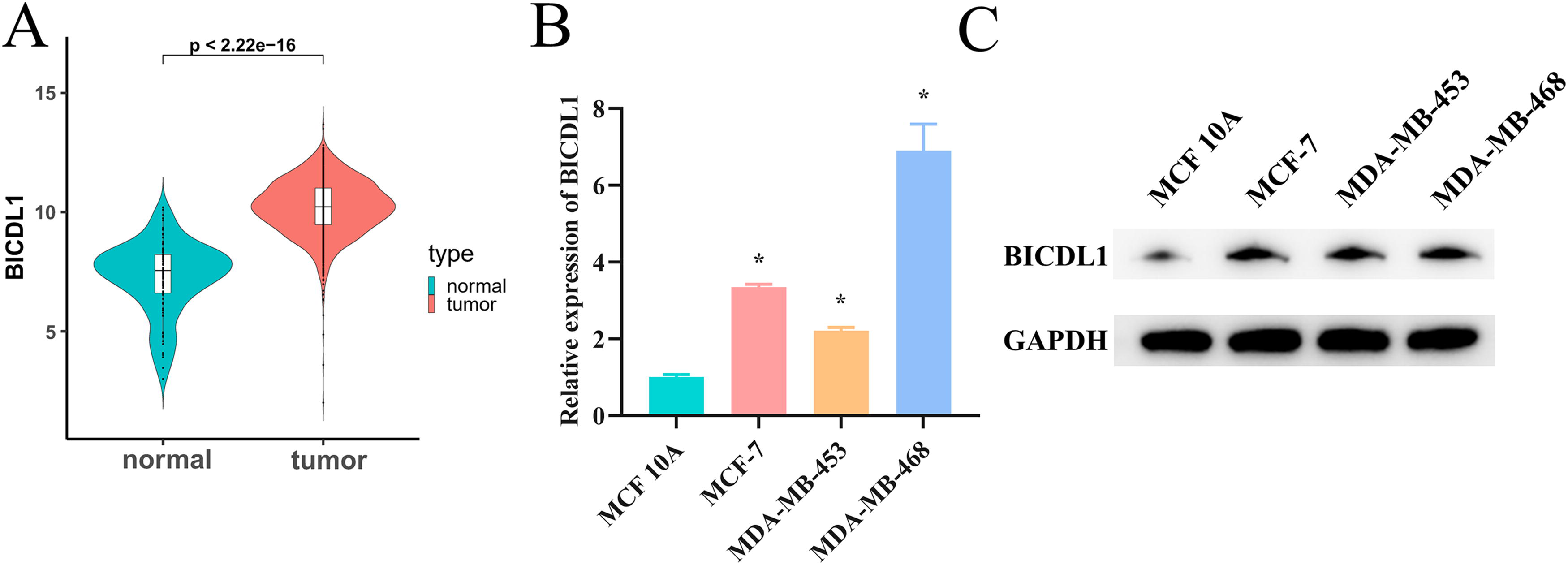
High expression of BICDL1 in BC.
BICDL1 promotes BC cell proliferation, inhibition, and migration
To elucidate the functional role of BICDL1 in BC, we focused on MDA-MB-468 cells for BICDL1 knockdown and MDA-MB-453 cells for BICDL1 overexpression, guided by preliminary findings. We confirmed transfection efficiency through qRT-PCR and WB analyses, ensuring the results were adequate for further experiments (Fig. 2A–B). Colony formation assays revealed that knocking down BICDL1 in MDA-MB-468 cells led to a significant suppression in both colony number and proliferative capacity. Conversely, overexpressing BICDL1 in MDA-MB-453 cells enhanced colony formation (Fig. 2C). Transwell assays unveiled that BICDL1 knockdown significantly suppressed the migration and invasion capabilities of MDA-MB-468 cells. In contrast, overexpression of BICDL1 in MDA-MB-453 cells significantly increased these abilities (Fig. 2D–E). Examining expressions of epithelial-mesenchymal transition (EMT) markers unveiled that knockdown of BICDL1 elevated E-cadherin while decreasing N-cadherin and Vimentin expressions in MDA-MB-468 cells. The opposite pattern was seen with BICDL1 overexpression in MDA-MB-453 cells (Fig. 2F). These findings collectively indicate that BICDL1 is crucial in promoting BC cell malignant progression.
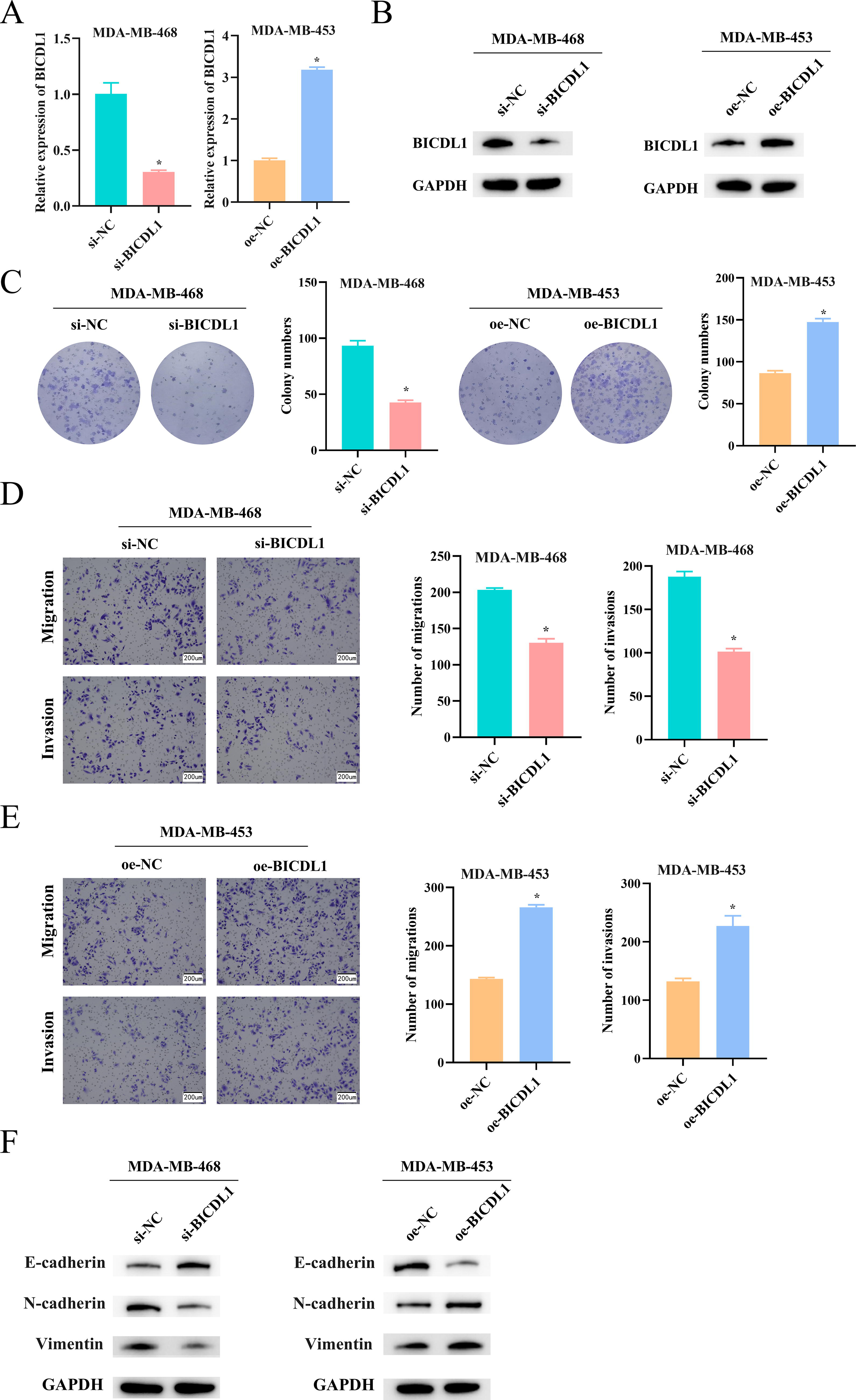
BICDL1 promotes BC proliferation, inhibition, and migration.
BICDL1 hinders the pyroptosis pathway in BC cells
To explore signaling pathways associated with BICDL1, we performed GSEA. BICDL1 was enriched in the pyroptosis pathway and was significantly negatively correlated with NLRP3 and CASP1, two pyroptosis markers (Fig. 3A–B). The qRT-PCR results showed that the expression of NLRP3 and CASP1 was upregulated upon BICDL1 knockdown (Fig. 3C), whereas BICDL1 overexpression exhibited the opposite trend (Fig. 3D). WB analysis of key pyroptosis proteins showed that BICDL1 knockdown increased the expression of NLRP3, procaspase-1, cleaved CASP1, GSDMD-N, and apoptosis-associated speck-like protein containing a CARD (ASC), indicating activation of the pyroptosis signaling pathway. Conversely, BICDL1 overexpression exhibited a reduction in these markers (Fig. 3E). Furthermore, we observed significant increases in LDH release and proinflammatory cytokines IL-1β and IL-18 levels following BICDL1 knockdown compared to the control group, while BICDL1 overexpression produced an inhibitory effect (Fig. 3F–I). These results suggest that BICDL1 effectively inhibits pyroptosis in BC cells.
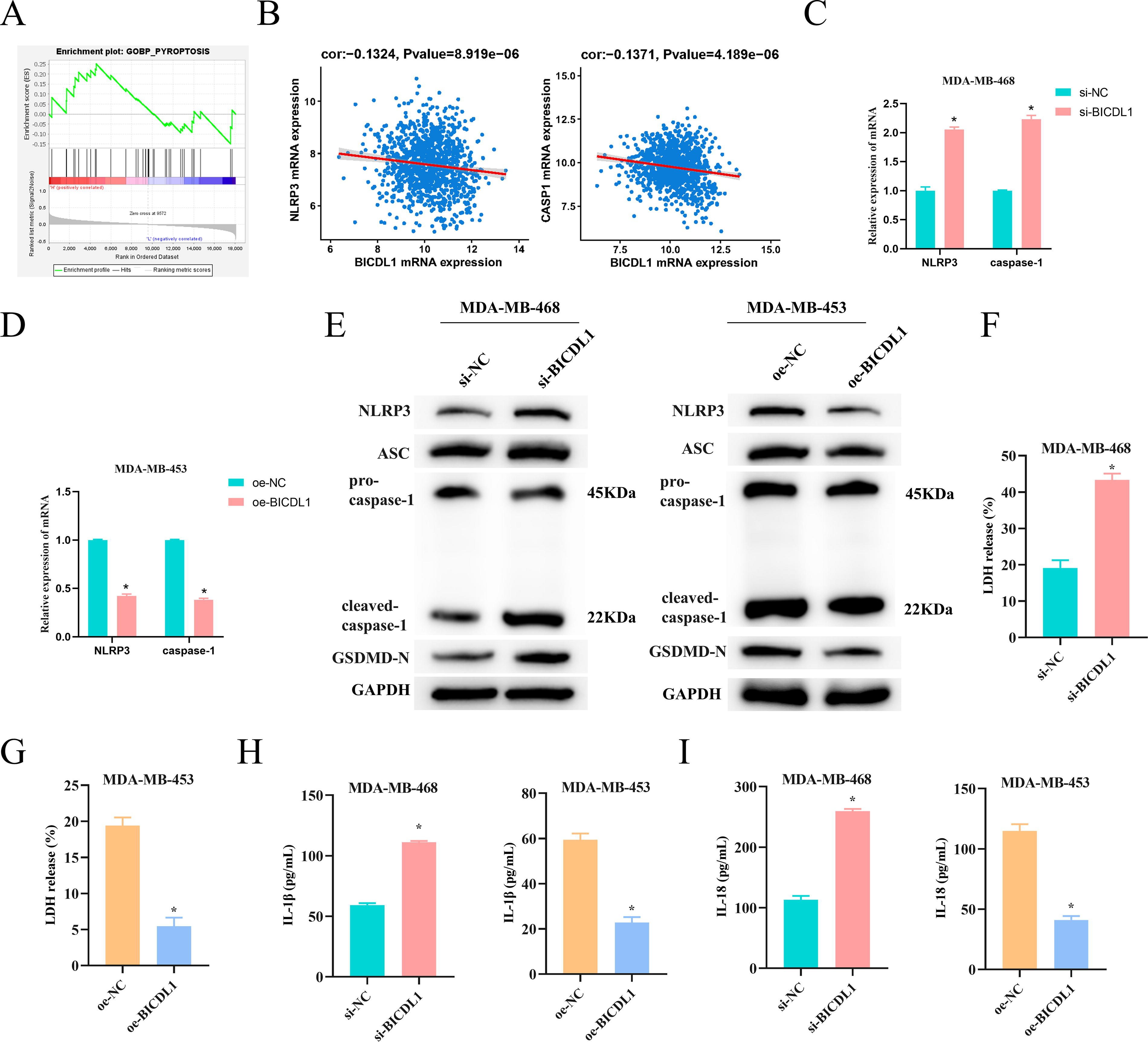
BICDL1 mediates the pyroptosis pathway in BC Cells.
BICDL1 hinders pyroptosis to promote BC cell proliferation, inhibition, and migration
Building on our previous findings, we hypothesized that BICDL1 enhanced BC cell proliferation, inhibition, and migration through the NLRP3-mediated pyroptosis signaling pathway. To test this, we utilized Nigericin, an ionophore known to induce pyroptosis by activating the NLRP3 inflammasome. 16 In our experiments with MDA-MB-453 cells, we established four groups: oe-NC, oe-BICDL1, oe-NC + Nigericin, and oe-BICDL1 + Nigericin. Colony formation assays demonstrated that while BICDL1 overexpression promoted cell proliferation, the introduction of Nigericin effectively reversed this effect (Fig. 4A). Moreover, Nigericin also negated the increased migration and invasion stimulated by BICDL1 overexpression (Fig. 4B). Similar trends were evident in expressions of EMT-related proteins in MDA-MB-453 cells (Fig. 4C). WB analysis of pyroptosis-related proteins revealed that BICDL1 overexpression inhibited expression of ASC, GSDMD-N, NLRP3, procaspase-1, and cleaved CASP1, but addition of Nigericin restored levels of these proteins (Fig. 4D). Additionally, the presence of Nigericin alleviated the suppressive effects of BICDL1 overexpression on LDH release and IL-1β and IL-18 levels (Fig. 4E–G). Together, these data support our hypothesis that BICDL1 enhances BC cell proliferation, inhibition, and migration via the NLRP3-mediated pyroptosis signaling pathway.
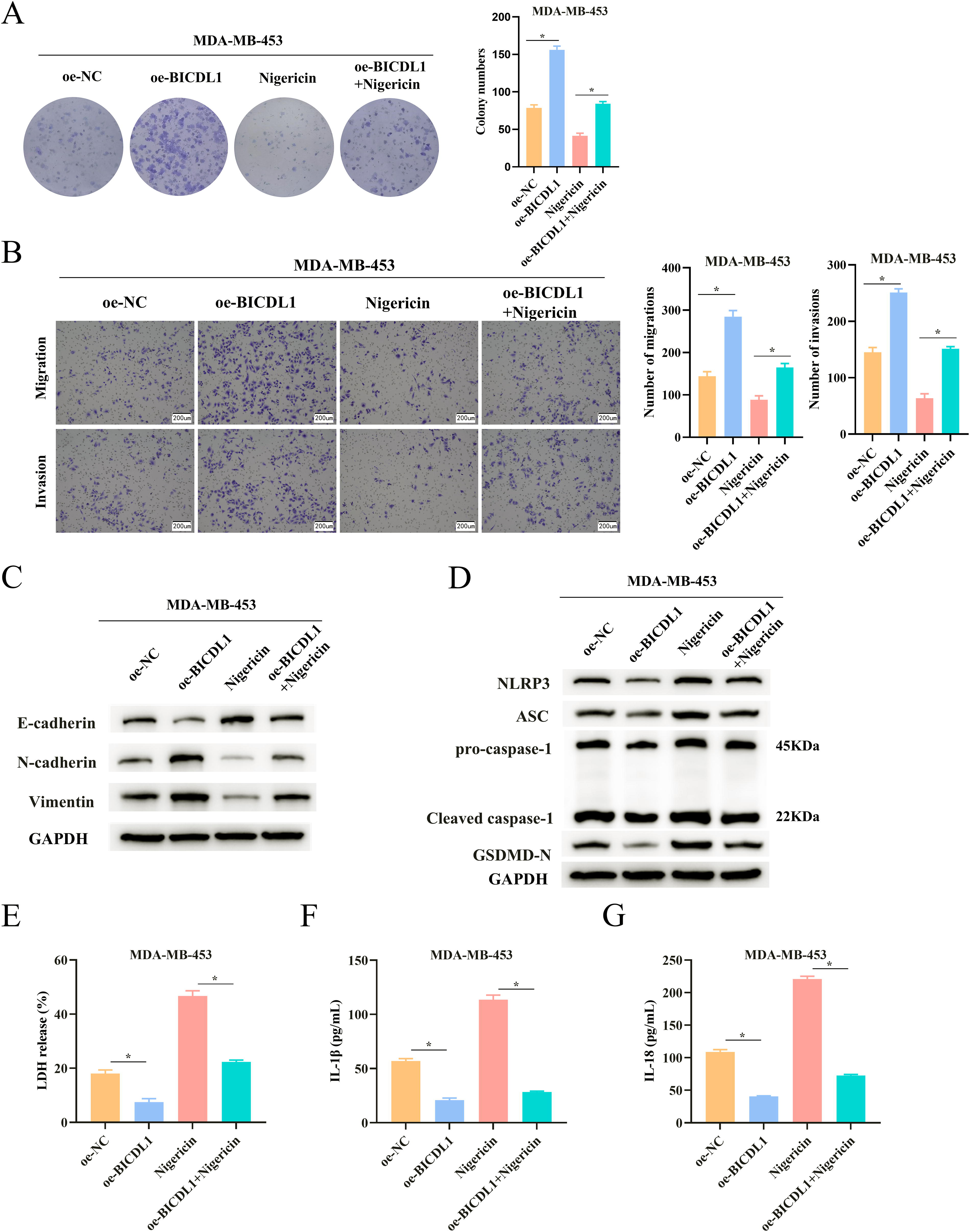
BICDL1 hinders pyroptosis to promote BC cell proliferation, inhibition, and migration.
Discussion
BC is a leading cancer threatening women’s health, underscoring the urgent need to identify effective molecular markers and therapeutic targets. 17 We identified BICDL1 overexpressed in BC, and our functional assessments revealed its role in fostering proliferation, inhibition, and migration of BC cells. Moreover, we elucidated the regulatory mechanism of BICDL1 in BC progression, demonstrating that it promoted tumor growth and metastasis by inhibiting pyroptosis.
BICDL1 has been identified as a crucial component in regulating vesicular transport within developing neurons. 18 BICDL1 is also implicated in cancer, with a study showing its significant overexpression in colorectal cancer tissues and a correlation with poorer overall survival in patients exhibiting high levels of BICDL1. 14 We initially analyzed BICDL1 expression in BC through bioinformatics methods, revealing a notable upregulation in BC tissues. This trend was confirmed in BC cell lines, suggesting that BICDL1 may be vital in BC progression. Subsequent cell-based experiments demonstrated that knocking down BICDL1 repressed BC cell malignant properties, while its overexpression exerted the opposite effects. BICD1, another member of the BICD family, facilitates malignant progression in hepatocellular carcinoma, with its knockdown causing reduced tumor cell properties. 19 This supports our findings regarding BICDL1. However, research on BICDL1 in cancer remains limited, and our study addresses this gap by confirming its oncogenic role in BC progression.
To delve deeper into the mechanisms by which BICDL1 influences BC proliferation and metastasis, we performed gene enrichment analysis, which highlighted a significant association of BICDL1 with the pyroptosis pathway. Further investigations established that BICDL1 suppressed expressions of pivotal pyroptotic proteins, such as NLRP3, GSDMD-N, procaspase-1, cleaved CASP1, and ASC, as well as the release of LDH and proinflammatory cytokines in BC cells. Previous research has underscored the central role of NLRP3 in initiating pyroptosis; CASP1 activation follows upon NLRP3 inflammasome formation, triggering pyroptosis and releasing inflammatory cytokines IL-1β and IL-18.20,21 Pyroptosis leads to cell swelling and lysis, causing a significant release of intracellular LDH. 22 Our results indicated that BICDL1 effectively repressed pyroptosis in BC cells. To investigate whether BICDL1 could affect BC progression through modulation of pyroptosis, we utilized Nigericin in our experiments. We found that overexpression of BICDL1 facilitated BC malignant progression by repressing pyroptosis, while the addition of Nigericin counteracted these effects. Inhibition of pyroptosis often accelerates BC progression, whereas its activation hampers tumor growth. 11 For instance, in triple-negative BC, activating pyroptosis reduces tumor cell proliferation and metastasis. 23 Lei et al. 24 also demonstrated that inducing pyroptosis effectively curbs abnormal BC proliferation. Hence, targeting BICDL1 to modulate pyroptosis may offer a promising therapeutic strategy for BC.
In summary, our study reveals a novel mechanism through which BICDL1 enhances BC proliferation, inhibition, and migration by hindering pyroptosis. These data suggest that BICDL1 has potential as a diagnostic marker for BC, contributing to improved diagnostic and therapeutic strategies. Nevertheless, our study has limitations; we focused exclusively on the cellular aspects of BICDL1 functions, necessitating further validation through animal models to bolster the reliability of our conclusions. Furthermore, given the critical role of BICDL1 in intracellular transport, it is essential to examine how its regulation of transport mechanisms may influence cancer progression.
Authors’ Contributions
Conceptualization: X.C. Data curation: L.X. and Y.Z. Formal Analysis: Y.Z. and X.L. Investigation: X.C. Methodology: L.X. and X.L. Project administration: X.C. Resources: X.C. and Y.Z. Supervision: X.L. Validation: L.X. Visualization: X.L. Writing—original draft: X.C. and Y.Z. Writing—review and editing: L.X. and X.L.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Data Availability Statement
The data in this study are available from the corresponding author on reasonable request.