
Abstract
Taurine and tauroursodeoxycholic acid are used for treating cerebral ischemia. In this study, we used the combination of taurine and tauroursodeoxycholic acid to establish a model of ischemia-reperfusion injury and evaluate the effect and target of the drug combination using the neurovascular unit (NVU) model of hypoxia and reoxygenation in vitro. Results showed that the combined application of these two drugs improved the survival of neurons, astrocytes, and endothelial cells; reduced inflammatory damage, levels of tumor necrosis factor-alpha, interleukin (IL)-6, and IL-1β, oxidative stress response, and the release of malondialdehyde and nitric oxide; and enhanced the activity of superoxide dismutase. Simultaneously, they acted on the blood–brain barrier (BBB) and improved the transendothelial electrical resistance value, reduced lactate dehydrogenase levels, improved the activity of γ-glutamyl transferase, and protected the integrity of the BBB against damage caused by oxygen-glucose deprivation/reoxygenation. At the same time, they prevented neuronal apoptosis, reduced the expression of Bax and caspase-3, and increased the expression of Bcl-2. These two drugs regulated the expression of connexin 43 (CX43) and aquaporin 4 (AQP4) in astrocytes, reducing the level of AQP4, and improving the activity of CX43. In addition, the drug combination increased the expression of tight junction proteins in endothelial cells, such as zona occludens-1, occluding, and claudin-5, and decreased the expression of matrix metalloproteinase (MMP)2 and MMP9. Furthermore, they acted together on the p38 mitogen-activated protein kinase (MAPK) signaling pathway, and the addition of the p38 inhibitor SB203580 partially inhibited the expression of p38MAPK. Thus, the combined action of these drugs protected the NVU.
Introduction
Although ischemic stroke is the leading cause of adult death and severe neurological dysfunction worldwide, effective treatments are scarce. 1 Tissue plasminogen activator (tPA) is the only Food and Drug Administration-approved drug for treating ischemic stroke, and currently, drugs for the treatment of hemorrhagic stroke are not available. Over the past few decades, many neuroprotective agents have been studied, which were either unsuccessful or are still in preclinical transition to clinical use. 2
Taurine (2-aminoethylsulfonic acid) is a sulfur-containing free amino acid abundant in mammals and is known to play a key nutritional role in the growth, differentiation, and development of brain cells. 3 Taurine differs from other amino acids in the presence of a sulfur group instead of a carboxyl group, which prevents its incorporation into proteins. It is found mainly in excitable tissues such as the brain, retina, heart muscle, and skeletal muscle. 4 In addition, taurine can be obtained from meat, dairy products, poultry, fish, and shellfish. Taurine, combined with tPA, can reduce the protein levels of matrix metalloproteinase (MMP)2 and MMP9, cerebral edema, and acute stroke injury. 5 Taurine exerts a protective effect on cortical neurons by inhibiting the hypoxia/reoxygenation or glutamate-induced endoplasmic reticulum stress. Under chronic hypoxia/reoxygenation or glutamate exposure, taurine significantly inhibits activation of the ATF6 and IRE1 pathways but not the pancreatic endoplasmic reticulum kinase (PERK) pathway.6,7 In contrast, taurine delays the activation of the PERK pathway during transient glutamate exposure.8,9 The neuroprotective effect of taurine on cerebral infarction was demonstrated by inhibiting the activities of 12/15-LOX, p38 mitogen-activated protein kinase (MAPK), and cPLA2. 10
Tauroursodeoxycholic acid (TUDCA) is an endogenous cholic acid that has been reported to regulate endoplasmic reticulum stress, mitochondrial function, reactive oxygen production, cytochrome C release, and neuroinflammation. 11 TUDCA exerts neuroprotective effects in different animal models of stroke and nervous system diseases. Furthermore, TUDCA has been shown to exert anti-inflammatory effects on glial cell culture and mouse models of acute neuroinflammation.12,13 TUDCA can reduce apoptosis by inhibiting the expression of caspase-12 and, thus, plays a protective role against secondary nerve injury.14,15 In addition, TUDCA protects retinal structures in mice by inhibiting microglia activation and inflammation. 16
Although neuroprotection has been demonstrated in experimental animal stroke models, many neuroprotective agents have not provided protection against ischemic stroke in clinical trials. These results showed the limitations of treatments focusing only on neurons in case of stroke. 17 The neurovascular unit (NVU), which is composed of neurons, astrocytes, and vascular endothelial cells, was conceptualized in 2001. 18 The NVU encompasses all neuronal components, interactions between cells, and signal transduction between cells and the extracellular matrix, which is critical for understanding the mechanism of ischemic stroke. Microglia, pericytes, and oligodendrocytes also play an important role in the maintenance of NVU function and can be used as new targets for stroke treatment.
This conceptual framework of NVUs links microvascular and neuronal function and their response to injury, as well as microvascular components to neurons via astrocytes. Therefore, in this study, the influence of these drugs on neurons and the blood–brain barrier (BBB) was investigated by establishing brain NVUs from cortical cells of rats subjected to oxygen-glucose deprivation/reoxygenation (OGD/R).
Materials and Methods
Animals
Sprague Dawley (SD) rats were purchased from Beijing Weitong Lihua Technology Co., Ltd. (license no. SCXK 2020-0012). All studies were conducted in accordance with the institutional guidelines of the Beijing University of Chinese Medicine.
Preparation of TUDCA and taurine
TUDCA (Mw: 499.7, purity ≥ 98%) was purchased from the National Institute for Food and Drug Control (Beijing, China). Taurine (Mw: 125.15, purity ≥ 98%, ab141063) was purchased from Abcam.
Cell culture
Primary culture of rat cortical neurons
Primary cortical neurons were obtained from SD rats within 24 hours of birth. The meninges and blood vessels were removed, and only the cerebral cortex was retained. The cerebral cortex was cut into pieces using ophthalmic shears, following which 0.125% ethylenediaminetetraacetic acid trypsin (Gibco 25200-056, USA) was added and incubated at 37°C for 30 minutes. Then, Dulbecco’s modified Eagle’s medium (DMEM)-F12 (HyClone SH30023.01, USA) containing 10% fetal bovine serum (FBS; Gibco 10099141C, USA) was added, and the suspension was beaten with a 1 mL tip to form a cell suspension; the cells were strained with a 70 μm strainer and centrifuged at 1000 rpm for 5 minutes. The supernatant was discarded, and the cells were resuspended by adding medium and then plated in cell culture plates precoated with poly-L-lysine (0.01%, P8920, Sigma) at the density of 1 × 106 cells/mL. The culture plates were incubated in a 5% CO2 humidified incubator at 37°C for 4−8 hours, after which the medium was replaced with neurobasal-A medium (Gibco 10888022, USA) supplemented with 2% B27 supplement and 0.25% GlutaMAX (Gibco 35050061, USA). Cortical neurons were cultured after 2−3 days.
Primary culture of rat cortical astrocytes
Primary cortical astrocytes were obtained from SD rats 3 days after birth. The meninges and blood vessels were removed, and the cerebral cortex was dissected using ophthalmic scissors, following which it was treated with 0.125% trypsin and incubated for 20 minutes. After that, DMEM-F12 containing 10% FBS was added, and the suspension was beaten with a 1 mL tip and then filtered using a 70 μm filter. Then, the suspension was centrifuged at 1000 rpm for 5 minutes, the supernatant was discarded, and the cells were plated in a culture plate at the density of 1 × 105 cells/mL. Primary astrocytes were cultured for 6−7 days, after which they were purified and subcultured for subsequent experiments.
Primary culture of rat cortical capillary endothelial cells
Primary microvascular endothelial cells were obtained from SD rats at 7−10 days of age. The meninges and blood vessels were removed under aseptic conditions, and only the cerebral cortex was retained. The cerebral cortex was cut to shreds using ophthalmic shears, following which 25% bovine serum albumin (BSA) solution was added and centrifuged at 3000 rpm for 5 minutes. The tissues were resuspended and centrifuged thrice, following which the supernatant was discarded, and the microvascular endothelial cells were retained. Then, 1% collagenase/dispase was added and incubated for 10 minutes, following which the cells were resuspended in the medium, centrifuged at 1000 rpm for 5 minutes; the supernatant was discarded, and the cells were plated at the density of 1 × 105 cells/mL. After 5−7 days of in vitro culture, 80% of the purified endothelial cells were used for subsequent experiments.
Establishment of OGD/R-injured NVU model in vitro
The neurons were derived from the 24-hour-old fetal rats born from SD rats, the astrocytes were derived from the 3-day-old fetal rats born from SD rats, and the endothelial cells were derived from the 7–10-day-old fetal rats born from SD rats.
An in vitro model of NVU was established as described previously. In this study, cortical capillary endothelial cells, astrocytes, and neurons of SD rats were cocultured according to their relative positions in vivo. First, the neurons (1 × 106 cells/mL) were harvested and plated in a transwell culture plate coated with Poly-L-Lysine (PLL) for 2−3 days. Then, the astrocytes (2 × 105 cells/mL) were plated on the back of transwell inserts (Millipore, 1.0 μm, USA) and incubated in a humidified atmosphere of 5% CO2 at 37°C for 3 hours. After 4 hours, the neurons were cocultured with DMEM-F12 culture medium containing 10% FBS. Then, microvascular endothelial cells (3 × 105 cells/mL) were implanted inside the transwell to complete the three-cell coculture.
Construction of the OGD/R model in vitro
Neurons, astrocytes, and endothelial cells were cultured for 3−5 days and then used for the OGD/R model culture. Earle's balanced salt solution was used as the medium for OGD for 1 hour, and 20% FBS DMEM-F12 medium was used as the medium for reoxygenation for 24 hours.19,20
Experimental groups and treatment
The in vitro NVU models were randomly divided into five groups: control group, OGD/R group, taurine + TUDCA (T + T) group, SB203580 group, and T + T + SB203580 group. With the exception of the control group, each group was exposed to OGD for 1 hour and R for 24 hours. The drug treatment groups were treated with TUDCA and taurine (1 μM and 1 mM, respectively) for 24 hours before OGD and during OGD (TUDCA, taurine, and SB203580 were added into the culture media of the insert).
Detection of cell viability
The cell counting kit 8 (CCK8) assay was used to determine cell viability, which was expressed as the percentage of living cells. Primary astrocytes cultured to the second generation were seeded in 96-well plates at a density of 1 × 105 cells/mL. Primary neurons were plated on PLL-coated 96-well plates at a density of 1 × 106 cells/mL. The microvascular endothelial cells were plated on gelatin-coated 96-well plates at a density of 1 × 105 cells/mL. OGD/R was performed after the 96-well plate was incubated for 24 hours. One hundred microliters of the CCK8 reagent diluted 10-fold with 10% FBS-supplemented DMEM-F12 was added to each well and incubated for 1 hour, following which the absorbance was detected at 450 nm using a microplate analyzer.
Neuronal apoptosis assay
Apoptosis was assessed using flow cytometry with a fluorescein isothiocyanate (FITC) annexin V apoptosis detection kit (Sigma, San Francisco, CA, USA) following the manufacturer’s instructions. Briefly, the cells were digested with trypsin and centrifuged, the supernatant was discarded, and the cells were resuspended in 100 μL binding buffer. Five microliters of annexin V and 5 μL propidium iodide (PI) were added to each sample and incubated at room temperature for 10 minutes in the dark. Next, 200 μL binding buffer was added, and the apoptosis rate was detected using flow cytometry.
Immunofluorescent staining
The cells were washed thrice with phosphate-buffered saline (PBS), fixed with paraformaldehyde for 10 minutes, washed thrice with PBS, and sealed using 5% BSA for 1 hour. Neurons were stained using antimicrotubule-associated protein 2 rabbit monoclonal antibody (17490-1-AP, 1:400, Proteintech), astrocytes with antiglial fibrillary acidic protein (GFAP) mouse polyclonal antibody (3670S, 1:400, Cell Signaling Technology), and endothelial cells with antivon Willebrand factor (vWF) rabbit polyclonal antibody (11778-1-AP, Proteintech). Next, the rabbit secondary antibody (ab6717, Abcam, 1:300) and mouse secondary antibody (SA00013-4, Proteintech, 1:300) were added to the cells. The cells were incubated with 4′,6-diamidino-2-phenylindole staining solution for 5 minutes, after which images were captured using a microscope.
Transendothelial electrical resistance (TEER) and serum-free (SF)
The TEER value of NVU was measured using an epithelial-volt-ohmic resistance meter (ERS-2, Millipore, USA). The cell-free group was used as the blank. The TEER value was calculated using the following equation: TEER = (sample TEER value-background TEER value) × the area of membrane (1.12 cm2). The values were expressed as Ω·cm2.
Detection of lactate dehydrogenase (LDH)
The release of LDH in the supernatant was determined as an indicator of cell membrane integrity. After OGD/R, LDH levels in rats were detected according to the manufacturer’s instructions (Jiancheng, Nanjing, China).
Detection of inflammatory markers
The expression of proinflammatory factors, tumor necrosis factor (TNF)-α, interleukin (IL)-6, and IL-1β, was determined using a rat TNF-α enzyme-linked immunosorbent assay (ELISA) kit (EK0526), rat IL-6 ELISA kit (EK0412), and rat IL-1β ELISA kit (EK0393), respectively (Boster Biological Technology Co., Ltd.).
Assay for oxidative markers and γ-glutamyltransferase
Nitric oxide (NO, A01212, Jiancheng, Nanjing, China), malondialdehyde (MDA; A00312, Jiancheng, Nanjing, China), superoxide dismutase (SOD; A00131, Jiancheng, Nanjing, China), and γ-glutamyl transpeptidase (γ-GT; c01711, Jiancheng, Nanjing, China) levels were measured according to the manufacturer’s instructions.
Detection of neuronal apoptosis
The annexin V/PI apoptosis kit (Sigma, San Francisco, CA, USA) was used to detect neuronal apoptosis. The cells were coincubated with annexin V-FITC and PI at room temperature for 10 minutes, followed by the addition of 200 μL buffer. In total, 1 × 104 cell/mL was used, and the apoptosis of neurons was detected using flow cytometry.
Western blotting
The effect of taurine and TUDCA on the expression of NVU proteins was detected using western blotting. In brief, after preparing brain cell protein samples using radioimmunoprecipitation assay lysis buffer, protein concentration was measured using a kit, and equal amounts of protein were electrophoresed. Next, the cells were treated with 5% BSA for 1 hour, and primary antibodies, namely, rabbit anti-Bax (ab32503), rabbit anti-Bcl-2 (ab59348), anticaspase-3 (66470-2-1g), rabbit anti-zona occludens-1 (ZO-1; ab216880), antioccludin (ab216327), mouse anticlaudin-5 (Ta347919), rabbit anti-MMP2 (10373-2-AP), rabbit anti-MMP9 (ab76003), rabbit anti-β-actin (SA00001-2), rabbit anti-connexin 43 (CX43; 26980-1-AP), rabbit anti-p38MAPK (14064-1-AP), and rabbit anti-aquaporin 4 (AQP4; 16473-1-AP), were added and incubated overnight at 4°C. After washing the membrane thrice with Tris-buffered saline, the corresponding secondary antibodies (1:2000) were added. The protein bands were observed using the Clarity Western ECL substrate kit. The ImageJ software was used to quantify the band densities (National Institutes of Health, USA).
Statistical analysis
Data were expressed as the mean ± standard deviation of values from at least three independent experiments and analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test of the GraphPad Prism 8.0 software (GraphPad, La Jolla, CA, USA). The statistical differences among the groups were analyzed through one-way ANOVA, and subsequent multiple comparisons were conducted using the post hoc Tukey’s test. p < 0.05 was considered statistically significant.
Results
Characterization of cells in the NVU model
The morphological characteristics of cells in the NVU model were assessed using immunofluorescence staining (Fig. 1) after 1–2 days of coculture. Neurons, stained using anti-MAP2 antibodies, were characterized by stereoscopic nuclear structure and long synapses and were interwoven into a network. Astrocytes, stained using anti-GFAP antibodies, were irregular in shape and had long protrusions. Endothelial cells, characterized by vWF immunostaining, occurred as a nonoverlapping continuous monolayer of spindle-shaped cells. 21
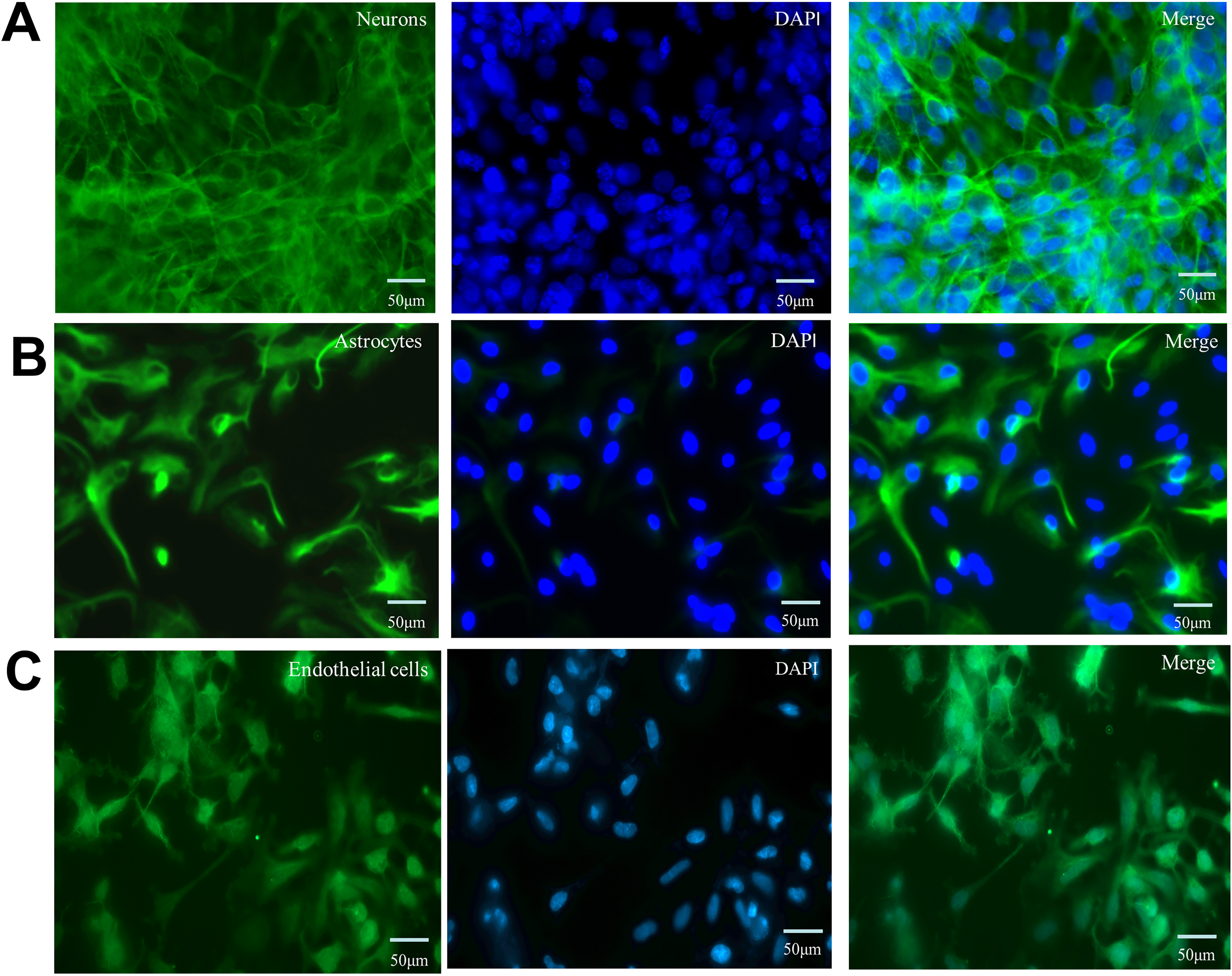
Immunofluorescence staining for identification of neurons, astrocytes, and endothelial cells in the NVU model.
Survival and viability of neurons, astrocytes, and endothelial cells
To investigate the synergistic protective effect of taurine and TUDCA on OGD/R-induced cell death, the CCK-8 assay was used to detect the viability of neurons, astrocytes, and endothelial cells (Fig. 2). We observed that 1 mM taurine exerted protective effect on neurons (p < 0.01). Neuronal activity was the strongest in the presence of 1 mM taurine and 1 μM TUDCA (p < 0.01). Furthermore, the combination of 1 mM taurine and 1 μM TUDCA showed the strongest protective effect on astrocytes (p < 0.01) and endothelial cells (p < 0.01); in addition, 1 mM taurine alone also exerted a protective effect on endothelial cells (p < 0.01). Therefore, 1 mM taurine and 1 μM TUDCA were selected as the optimal dose.
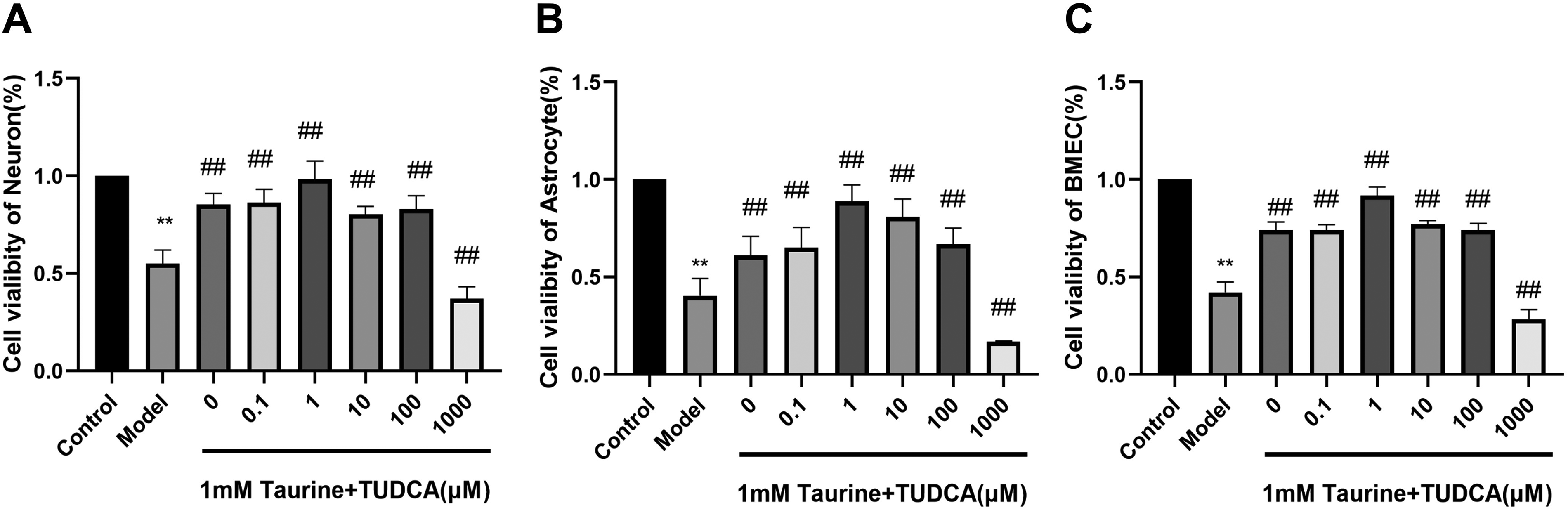
Effects of taurine and tauroursodeoxycholic acid on cell viability. Data are presented as mean ± SD. n = 4. Compared with the control group, **p < 0.01. Compared with the model group. ##p < 0.01. SD, standard deviation.
Effects of taurine and TUDCA combination on BBB permeability and LDH after OGD/R
TEER reflects the function of BBB, LDH reflects the degree of cell damage, and γ-GT is a BBB-specific enzyme. The activity of γ-GT indicates the integrity of BBB (Fig. 3). LDH levels in the model group were higher than those in the normal group (p < 0.01), whereas TEER value and γ-GT were lower (p < 0.01). LDH levels in the T + T group were lower than those in the model group (p < 0.01), whereas TEER value and γ-GT were higher (p < 0.01), which indicated that the taurine and TUDCA application reduced cell damage and protected the BBB.
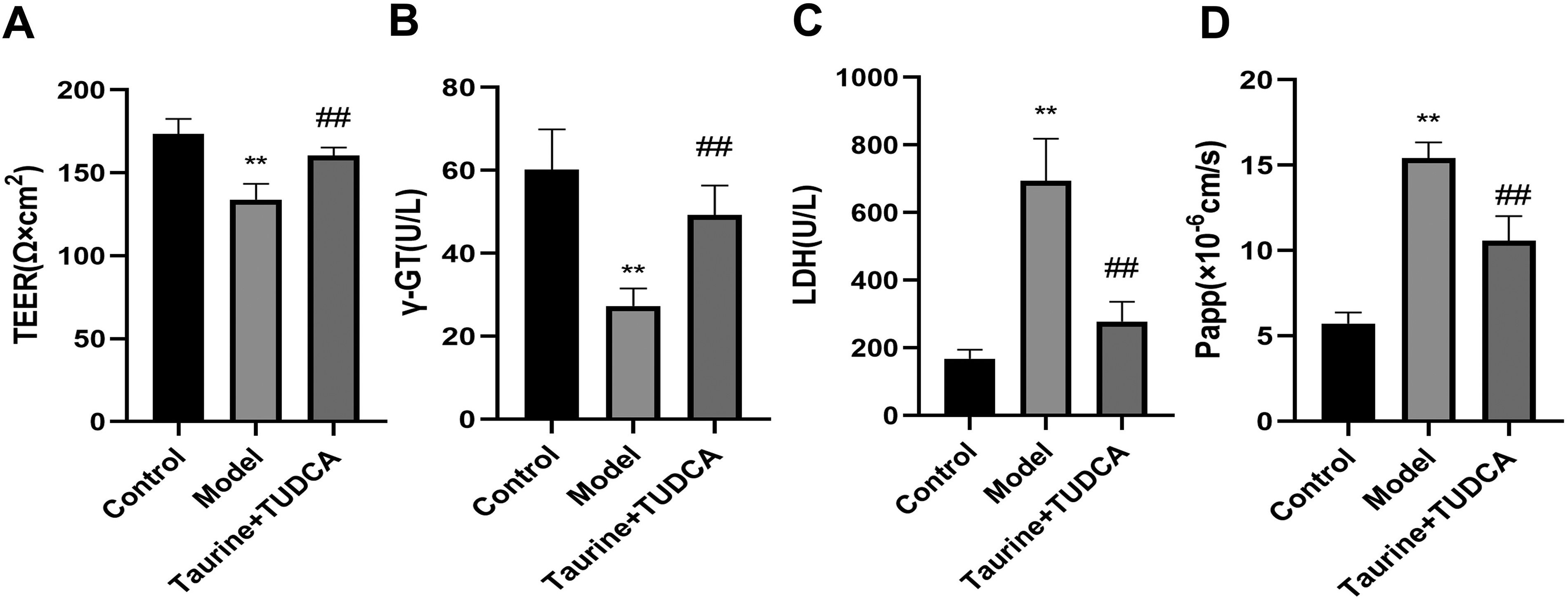
Effects of taurine and tauroursodeoxycholic acid (TUDCA) on blood–brain barrier function in vitro. Effects of taurine + TUDCA on LDH, TEER, γ-GT, and Papp on NVU after OGD/R. Data are presented as mean ± SD. N = 4, compared with the control group. **p < 0.01 compared with the model group, ##p < 0.01. LDH, lactate dehydrogenase; OGD/R, oxygen glucose deprivation/reoxygenation; TEER, transendothelial electrical resistance; TUDCA, tauroursodeoxycholic acid; γ-GT, γ-glutamyl transpeptidase.
Effects of taurine and TUDCA combination on oxidative stress injury after OGD/R
To investigate the effects of taurine and TUDCA on oxidative stress induced by OGD/R injury, NO, MDA, and SOD levels were detected in the cell supernatant (Fig. 4). Results showed that NO and MDA levels in the model group were higher than those in the control group, whereas SOD activity was lower (p < 0.01). NO and MDA levels in the T + T group were lower than those in the model group (p < 0.01), whereas SOD activity was higher (p < 0.01), indicating that oxidative stress was inhibited after the administration of the combination of taurine and TUDCA.
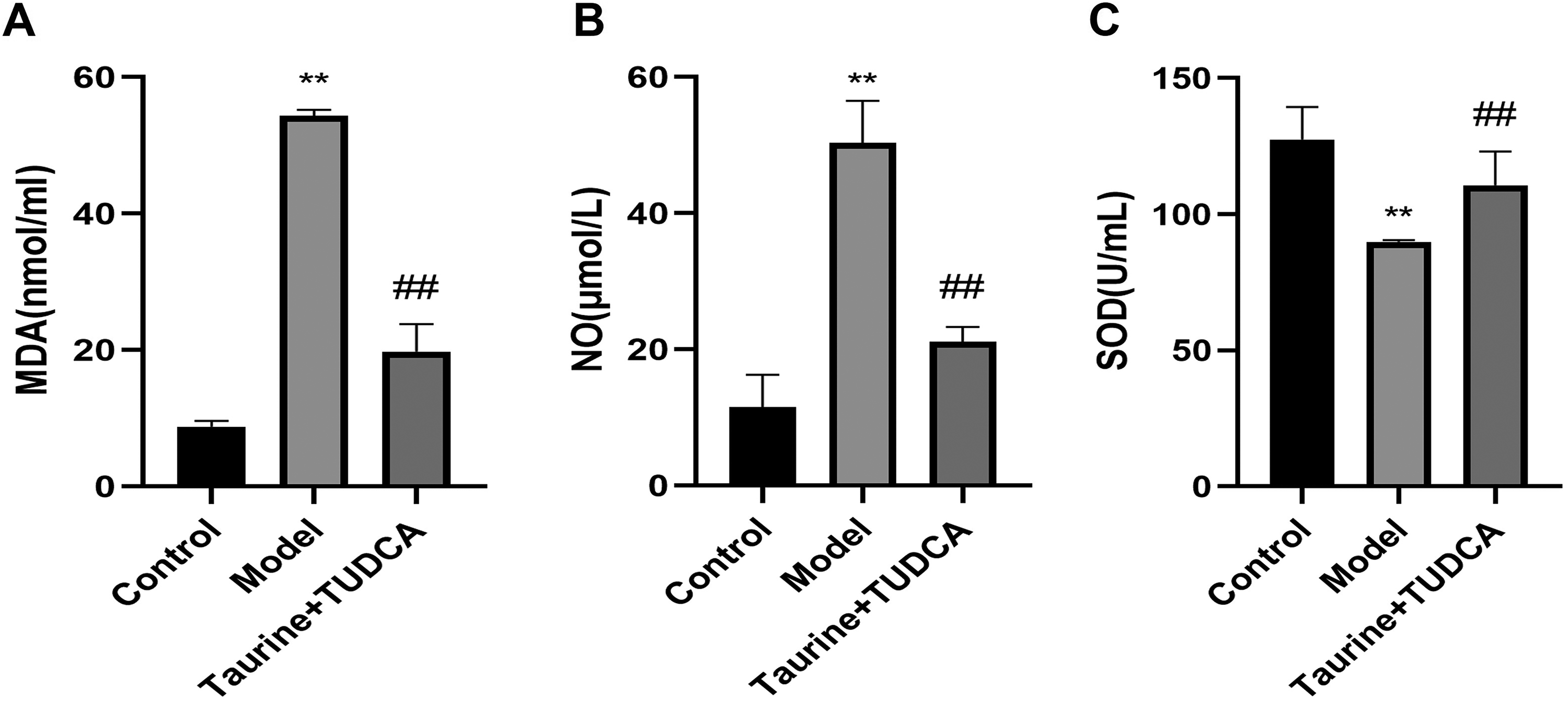
Taurine and tauroursodeoxycholic acid attenuated oxidative stress in NVU induced by OGD/R injury. Data are presented as mean ± SD. n = 4. Compared with the control group, **p < 0.01; compared with the model group, ##p < 0.01.
Effects of taurine and TUDCA combination on inflammatory factor after OGD/R
To determine whether ischemic reperfusion injury causes inflammation, we measured IL-1β, IL-6, and TNF-α levels after OGD/R (Fig. 5). Results showed that TNF-α, IL-1β, and IL-6 levels in the model group were higher after OGD/R than those in the control group (p < 0.01), whereas TNF-α and IL-6 levels decreased after treatment with taurine and TUDCA (p < 0.05). IL-1β level decreased after treatment with taurine and TUDCA (p < 0.01) indicating that the combination of these two drugs can reduce the inflammatory response caused by OGD/R.
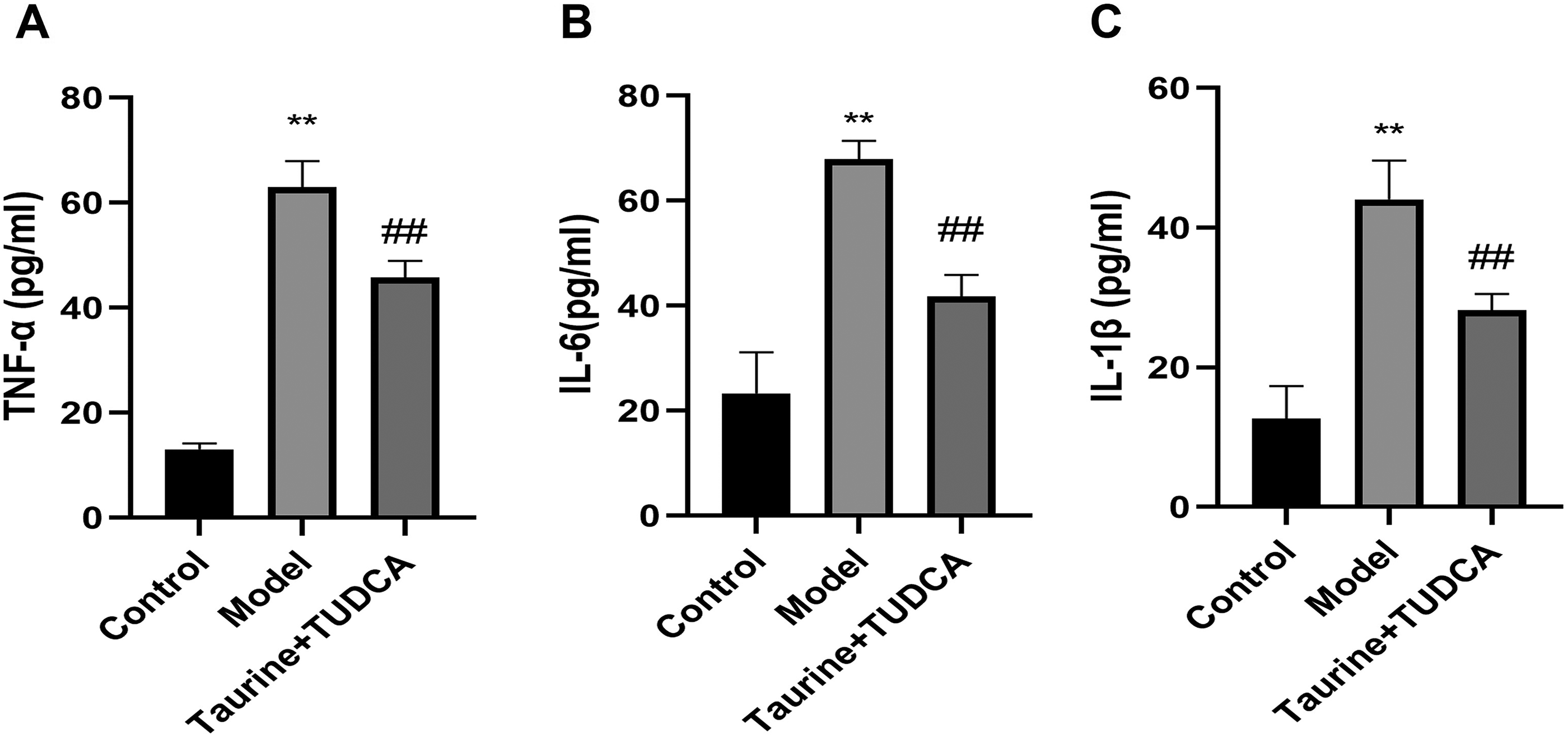
Combined administration of taurine and TUDCA attenuated the effect of OGD/R on inflammatory factors. Data are presented as mean ± SD. n = 4. Compared with the control group, **p < 0.01; compared with the model group, ##p < 0.01.
Effects of combined taurine and TUDCA on apoptosis and expression of apoptotic proteins, Bax, Bcl-2, and caspase-3, after OGD/R
Apoptosis was detected using flow cytometry (Fig. 6). Apoptotic proteins such as Bax, Bcl-2, and caspase-3 were detected using western blotting (Fig. 6). The rate of neuronal apoptosis was higher in the OGD/R group than in the control group (p < 0.01), whereas it was lower in the T + T group (p < 0.01). The rate of neuronal apoptosis was higher in the T + T + SB203580 group than in the T + T group (p < 0.05). Compared with that in the control group, OGD/R increased Bax and caspase-3 levels and decreased Bcl-2 levels (p < 0.01). Bax levels were lower and Bcl-2 levels were higher in the T + T group than in the model group (p < 0.01). Caspase-3 levels were lower in the T + T group (p < 0.05), whereas Bax levels were lower and Bcl-2 levels were higher in the SB203580 group than in the model group (p < 0.05). Bcl-2 levels were lower and caspase-3 levels were higher in the T + T + SB203580 group than in the T + T group (p < 0.05).
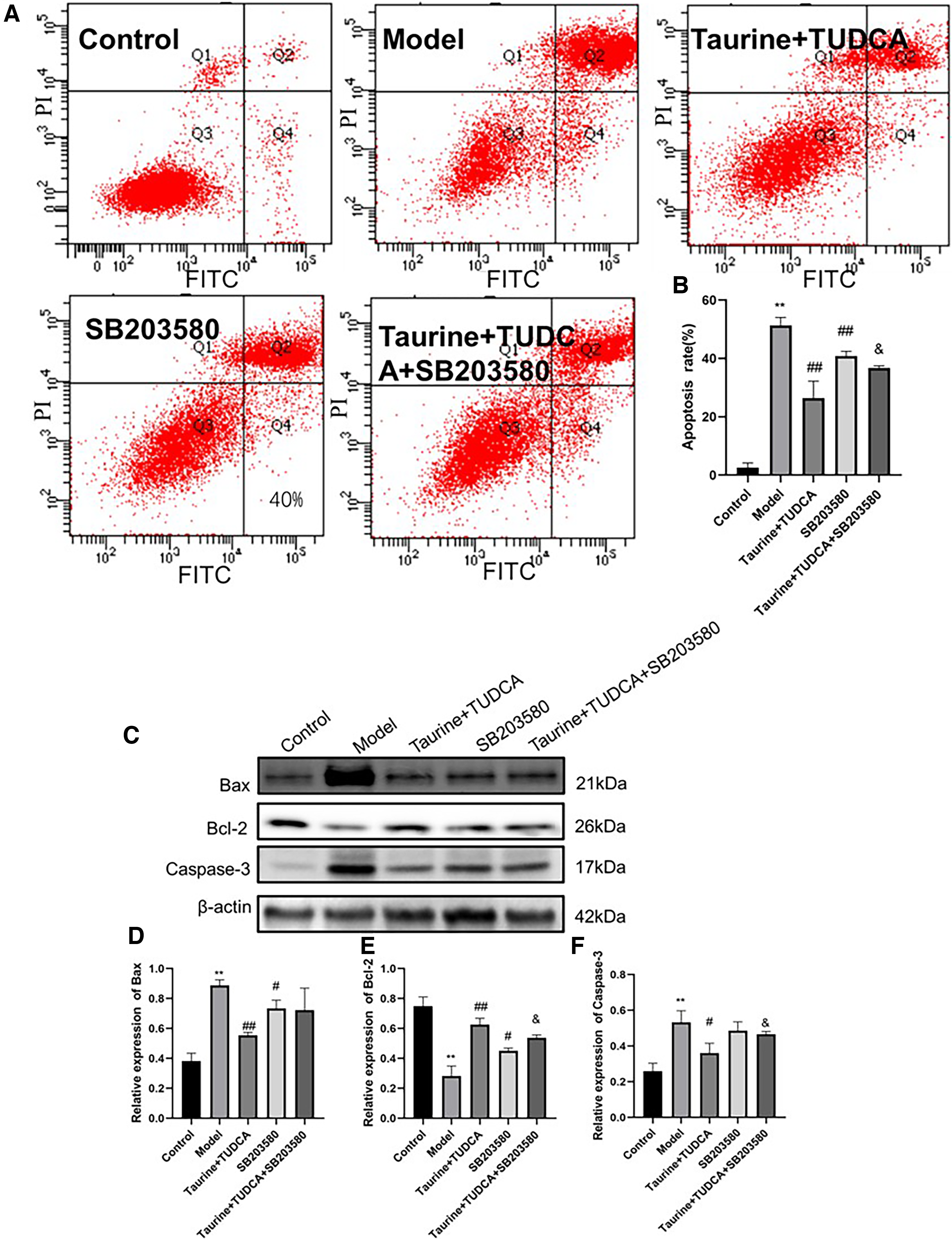
Effects of taurine and TUDCA on neuronal apoptosis after OGD/R.
Effects of taurine and TUDCA combination on the expression of tight junction (TJ) proteins, ZO-1, occludin, and claudin-5
To investigate the effect of T + T on the BBB after OGD/R preconditioning, we detected the expression of TJ proteins, occludin, claudin-5, and ZO-1 in endothelial cells using western blotting (Fig. 7). Compared with that in the control group, the expression of ZO-1, occludin, and claudin-5 in the model group (p < 0.01) decreased after OGD/R, whereas that in the T + T group was higher than that in the model group (p < 0.01). The expression of ZO-1 and claudin-5 in the T + T + SB203580 group was lower than that in the T + T group (p < 0.05). Occludin expression decreased in the T + T + SB203580 group (p < 0.01).
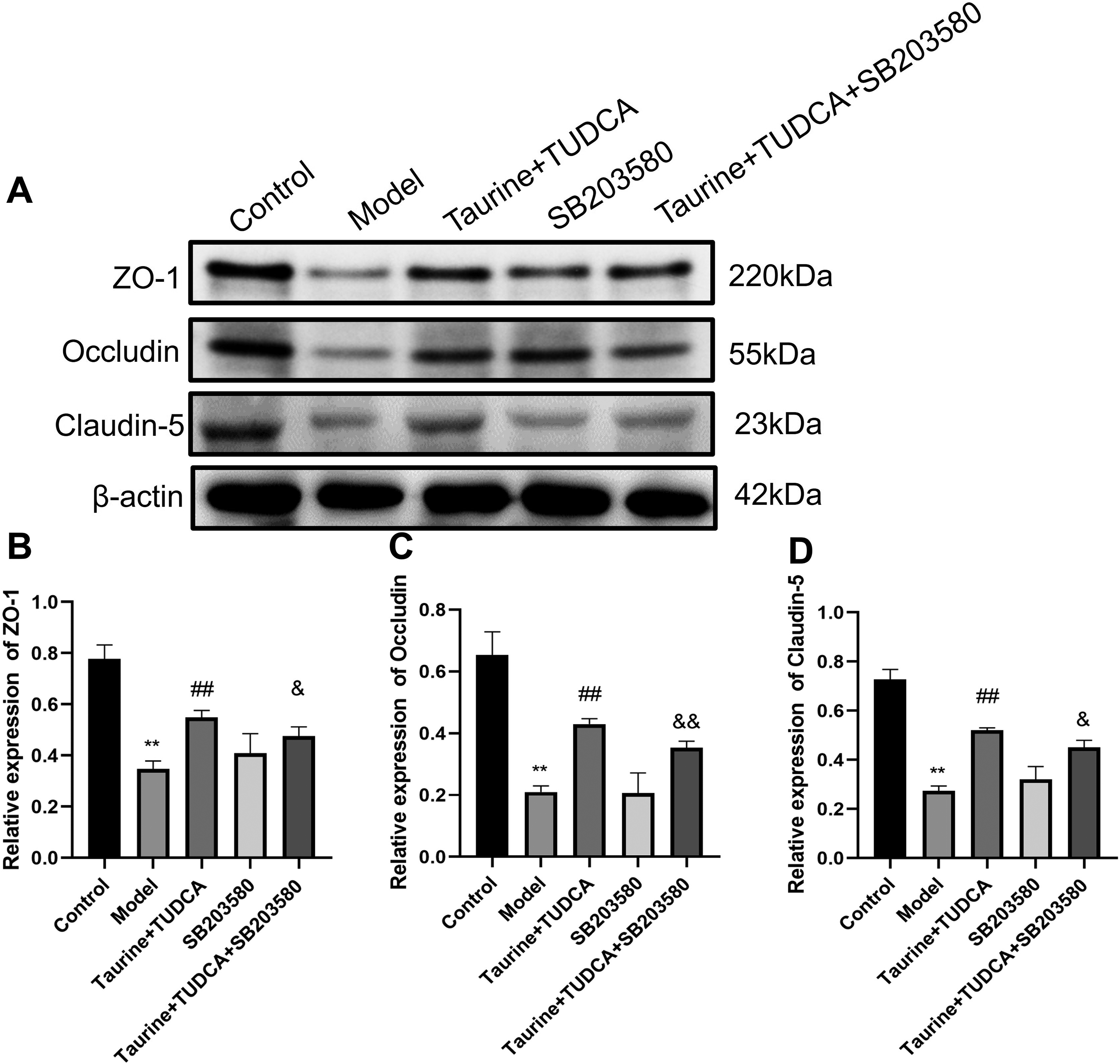
Effects of taurine and TUDCA on BBB tight junction protein expression after OGD/R.
Effects of taurine and TUDCA combination on the expression of BBB metalloproteinases, MMP2 and MMP9, after OGD/R
Many types of proteases are activated during ischemic stroke, and MMPs, particularly MMP-2 and MMP-9, are critical for TJ complex degradation and destruction of the BBB (Fig. 7). MMP-2 is produced by neurons, glial cells, and endothelial cells under ischemia, and its level is significantly increased and activated in the first 24 hours after reperfusion, contributing to early and reversible BBB leakage. Peripheral leukocytes cross the BBB and enhance glial cell activation, inducing MMP-9 activation. MMP-9 is important for maintaining long-term leakage of the BBB. Compared with that in the control group, the expression of MMP2 and MMP9 in the model group increased after OGD/R (Fig. 8, p < 0.01), whereas that in the T + T group was lower than that in the model group (p < 0.05). MMP2 level in the T + T + SB203580 group was lower than that in the T + T group (p < 0.05).
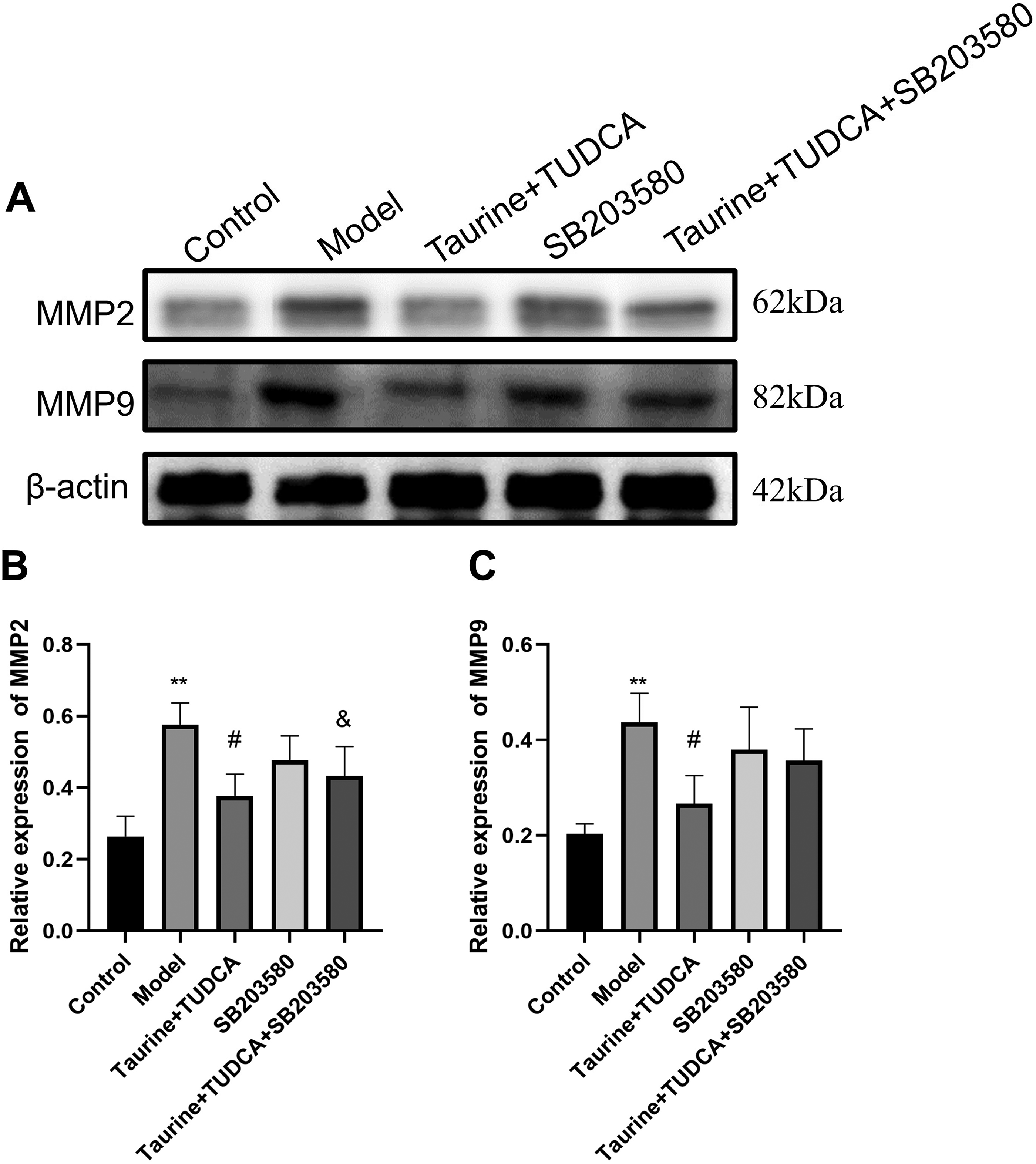
Effect of taurine and tauroursodeoxycholic acid (TUDCA) on expression of NVU metalloproteinases, MMP2 and MMP9, after OGD/R.
Effects of taurine and TUDCA combination on CX43 and AQP4 protein levels after OGD/R
We detected CX43 and AQP4 protein levels, markers of brain edema, in astrocytes (Fig. 9). CX43 levels were lower and those of AQP4 were higher in the model group than in the control group (p < 0.01). CX43 levels were higher (p < 0.05) and those of AQP4 were lower (p < 0.01) in the T + T group than in the model group, whereas the levels of AQP4 decreased in the SB203580 group (p < 0.05). These results showed that the combined application of taurine and TUDCA inhibited the occurrence of brain edema and promoted nerve repair.
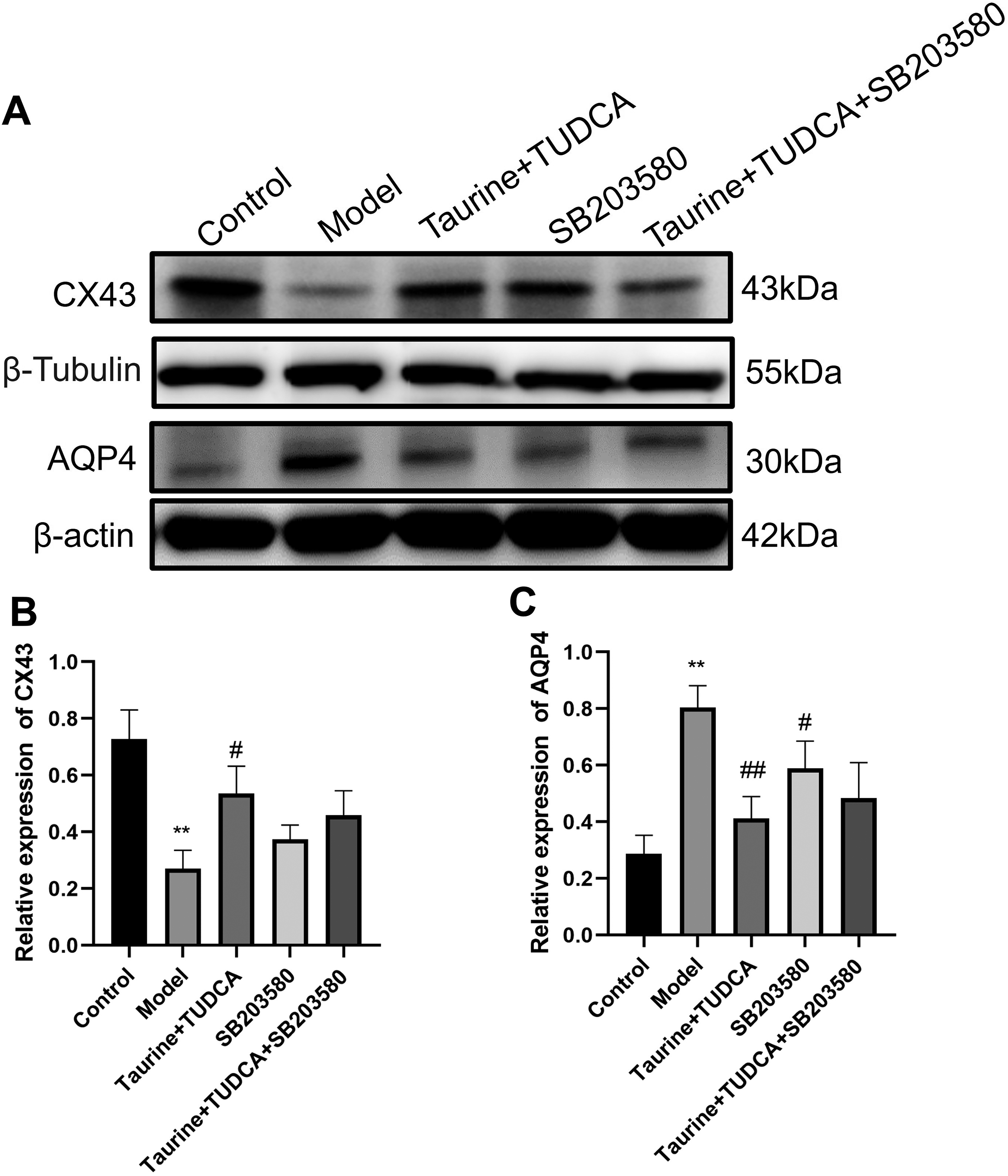
Effects of taurine and tauroursodeoxycholic acid on CX43 and AQP4 protein expression in astrocytes after OGD/R.
Taurine and TUDCA inhibited the activation of the p38MAPK pathway and protected the NVU against OGD/R damage
The p-p38/p38 MAPK expression in BBB after OGD/R was studied using western blotting (Fig. 10). p-p38/p38 MAPK levels in the model group were higher than those in the normal group (p < 0.01); compared with that in the model group, the protein level of p-p38/P38 MAPK in the T + T group and SB203580 group decreased (p < 0.01; p < 0.05); protein levels in the T + T + SB203580 group were lower than those in the T + T group (p < 0.05), which indicated that after treatment, the p38MAPK pathway was inhibited, and that SB203580 partially blocked p38MAPK.
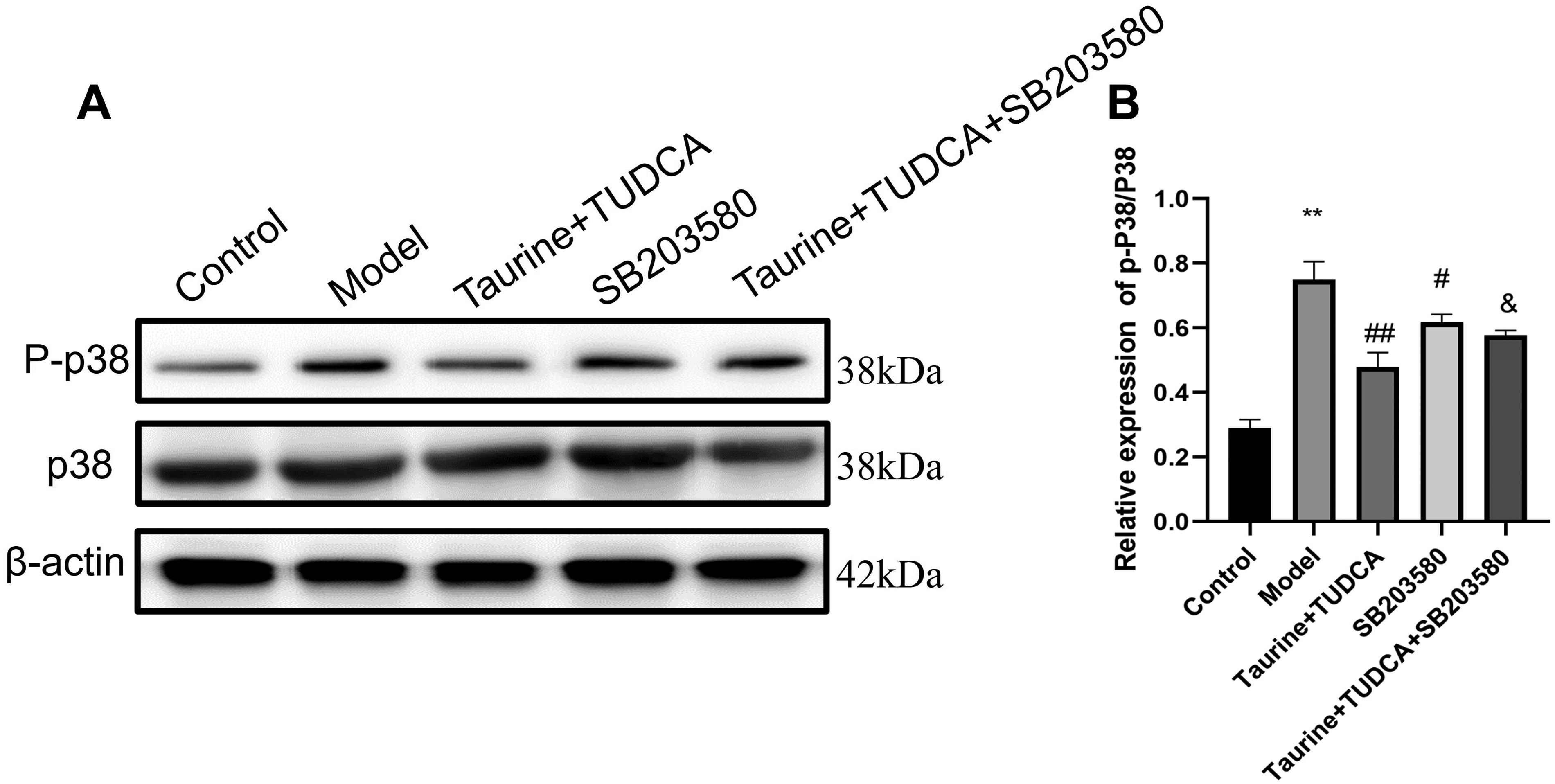
P-p38 and p38 protein expression in the BBB detected using western blotting. *p < 0.05, model group vs. control group, &p < 0.05, T + T+SB203580 group vs. model group.
Discussion
This study fills the gap in the research on the protective effects of taurine and TUDCA on the BBB. It has protective effects on the related proteins of the BBB, such as ZO-1, Occludin, and Claudin-5, and can also protect neurons and inhibit apoptosis. The combined use of the two drugs yields better therapeutic effects than when used alone.
As a naturally occurring amino acid, taurine is unlikely to cause excessive side effects in the human body. According to toxicity studies, it has not shown any effects of causing genetic mutations, carcinogenesis or teratogenesis. 22 Taurine is one of the most abundant metabolites in the central nervous system. Taurine has a neurotrophic effect and plays a significant role in the early stages of brain development, including the growth of nerve projections, the formation of synapses, and synaptic transmission. 23
A risk assessment study indicates that the maximum daily intake of taurine is 3 g. Based on the toxicological analysis of all relevant human clinical trials, this dose has not shown any adverse effects. However, the minimum intake level causing adverse reactions has not yet been determined. 24
The safety of the experimental compounds is of crucial importance for human clinical trials. The safety of hydrophilic bile acids has gained extensive experience in liver and gallbladder indications. TUDCA has been used for the treatment and prevention of cholesterol gallstones, as well as various diseases such as primary biliary cirrhosis, liver cirrhosis, primary sclerosing cholangitis, polycystic liver disease, pregnancy-induced intrahepatic cholestasis, and chronic liver disease fatigue. All these studies have generally confirmed good safety, with mostly mild gastrointestinal adverse events reported, such as diarrhea, abdominal pain, nausea, and vomiting, whereas rashes and itching are less common.25–27
The use of drug combinations is applicable to diseases where the BBB is compromised, such as stroke. However, as this study only involved cell experiments, the time range of drug administration still needs to be clarified through animal experiments. Taurine has the function of a neurotransmitter in the central nervous system. Taurine is an inhibitory neuroregulator. 28
As a potential drug component, its effects in combating oxidative stress and inflammation have been explored in various studies. It can prevent various diseases and disorders in different organ systems (such as the skin, cardiovascular, respiratory, muscular, skeletal, circulatory, and endocrine systems). 29
Taurine can reduce neural inflammation by lowering inflammatory factors IL-6 and TNF-α, improve oxidative stress, and treat cerebral ischemia through the P38 MAPK pathway and by inhibiting apoptosis. 22 Taurine may have the ability to reduce ischemia by increasing the level of calpain and reducing the expression of calpain and caspase-3. 5 Taurine can also achieve the goal of improving neurological function and treating ischemic stroke by inhibiting endoplasmic reticulum stress. 6
The anti-inflammatory properties of TUDCA have also been verified under various pathological conditions. The molecular basis for TUDCA’s anti-inflammatory effect is to reduce nitrite production by transcriptionally inhibiting inducible NO synthase and to inhibit the proinflammatory stimulus-induced activation of Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells (NF-κB) in glial cells. 30 TUDCA has proposed an additional anti-inflammatory mechanism by activating the transforming growth factor β pathway in the mouse brain. 31 TUDCA not only alters the phenotype of microglia in vitro but also in vivo toward an anti-inflammatory state. By binding to the bile salt receptor GPBAR1/TGR5, TUDCA increases the intracellular cAMP level in microglia, inducing anti-inflammatory markers while reducing proinflammatory markers. 13 TUDCA treatment effectively alleviated behavioral disorders, reversed the downregulation of TGR5, weakened the activation of microglia, and prevented neuroinflammation by inhibiting NF-κB signaling pathway, thereby reducing proinflammatory cytokines (IL-1β, TNFα, and IL-6) in the hippocampus and increasing anti-inflammatory cytokine (IL-10). 32
The pathological process of cerebral ischemia is complex and mainly includes inflammatory reaction, energy metabolism disorder, oxidative stress, and destruction of the BBB. The BBB, an important component of the NVU, contains TJ proteins and various ion and nutrient transporters that maintain normal brain physiology. Disruption of the BBB is the main pathological feature of ischemic stroke, which is the result of a combination of various factors, including MMPs, inflammatory regulators, blister transport, and oxidative pathways. 33
Compared with cells cultured alone, the coculture of neurons, astrocytes, and endothelial cells simulates the environment of ischemia reperfusion injury and closely resembles the in vivo physiological and pathological characteristics in terms of morphology and function. 34 The cerebral cortex cells of SD rats were selected for coculture of neurons, astrocytes, and endothelial cells, which are important components of the NVU, the basic structure of the central nervous system. 35 Previous studies have examined the effects of drugs on individual cells such as neurons, while ignoring the interactions among various cell types. Here, we established an in vitro NVU model and studied the therapeutic effect of drugs on ischemia-reperfusion. Neuronal injury is an important aspect of ischemia-reperfusion, and cerebral capillary endothelial cells are damaged after OGD/R. TJs are the most significant features of cerebral capillary endothelial cells and are key for ensuring the integrity of the BBB. 36 TJ proteins include ZO-1, occludin, and claudin-5. The entry of inflammatory cells into brain tissue leads to poststroke inflammation, which is another important mechanism of ischemia-reperfusion injury. 37 IL-1β is one of the most widely studied cytokines in stroke and neuroinflammation. Studies have shown that IL-1β-induced brain injury is associated with an increase in BBB permeability. 38 IL-6, a proinflammatory cytokine, has been shown to destroy the integrity of the BBB in rat brain endothelial cells by reducing TEER values. 39
In this study, the neuronal apoptosis rate increased, BBB TEER value decreased, LDH level increased, the γ-GT activity decreased, and the expression levels of ZO-1, occludin, and claudin-5 decreased significantly after OGD/R. The levels of inflammatory factors increased, oxidative stress response was enhanced, and edema of degree of astrocytes increased. These findings suggested that OGD/R damaged neurons, astrocytes, and endothelial cells.
Previous studies have suggested that neuronal cell death may occur via various mechanisms. 40 After stroke, the expression of neuronal apoptosis-related proteins, Bcl-2 and Bax, is perturbed. 41 Bcl-2 is an inhibitor of apoptosis that inhibits the release of apoptotic factors such as cytochrome C by inhibiting mitochondrial membrane permeability. 42 After OGD/R, the Bcl-2 level in neurons decreased, whereas that of Bax increased. Taurine and TUDCA pretreatment significantly downregulated Bax and Caspase-3 and upregulated Bcl-2. Therefore, these results suggested that the combined administration of the two drugs with antiapoptotic effects can prevent neuronal death caused by OGD/R injury.
Previous studies have shown that AQP4, the most abundant aquaporin in the brain, is mainly present in astrocyte foot and is associated with various inflammatory diseases such as brain edema.42,43 Hypoxia can increase AQP4 expression in astrocytes; furthermore, AQP4 expression increased in brain edema. 44 We detected AQP4 in BBB and showed that AQP4 level increased after OGD/R and decreased after preadministration of taurine and TUDCA, which reduced brain edema. AQP4 expression selectively affects the expression of the CX43 subtype of gap junction proteins. 45 Furthermore, AQP4 and CX43 were located around the blood vessels in the astrocyte terminal foot, 46 where CX43 acts as an immunomodulatory factor of glial blood vessels, namely the BBB, 47 and contributes to the release of glial neurotransmitters. 48 CX43 is highly expressed in astrocytes, especially at the interface of BBB, 46 where it forms gap junctions and hemichannels to maintain the normal morphology and function of astrocytes. 49 CX43 and AQP4 are known to be involved in the formation, development, and clearance of brain edema after brain injury. 50 On the surface of astrocytes, CX43 and AQP4 coordinate the transfer of ions and water between cells to maintain intracellular homeostasis.
The p38 MAPK pathway regulates cell death and survival. 51 The activation of p38 MAPK has been well demonstrated to exert beneficial or protective effects in ischemic models of cerebral ischemia or other organs, especially in ischemic preconditioning models. 52 In addition, many other studies have reported that p38 MAPK activation can protect neurons in ischemic preconditioning organs. 53
In addition, p38MAPK is associated with AQP4-mediated regulation of cortical astrocytes after ischemic injury. 54 SB203580 inhibited p38 MAPK significantly, which reduced AQP4 expression, indicating that p38 MAPK was related to AQP4 expression. 55 Recent studies have shown that the p38/MAPK pathway is activated during ischemia-reperfusion injury in astrocytes and neurons. 56 P38MAPK, as an upstream signaling molecule, under oxidative stress and inflammatory conditions, activates the apoptotic protein (caspase BAX) and inhibits the antiapoptotic protein (Bcl-2). 57 The proapoptotic proteins released during apoptosis can further activate P38MAPK, amplifying the apoptotic signal. Inhibiting the apoptotic proteins may inhibit the activation of P38MAPK, thereby limiting the extent of apoptosis. 58
Conclusions
Our study provides a new idea for using a combination of drugs for treating ischemic stroke and shows that the effect of the drug combination is better than that of a single drug. Both drugs can be used to treat ischemic stroke, and the mechanism of action of the two drugs is similar. In the NVU model, the two drugs simultaneously acted on different proteins of neurons, the BBB, astrocytes, and endothelial cells, leading to target diversification. Thus, this approach will be conducive for the development of future therapies for ischemic stroke. This article only focuses on the protective effects of taurine and TUDCA on the in vitro brain nerve vascular unit. It cannot represent the actual brain ischemic environment in vivo, and further animal experiments are needed to verify this. After drugs enter the body, they need to go through processes such as absorption, distribution, metabolism, and excretion. These processes are closely related to the functions of various organs, and cell experiments cannot fully simulate them. The drug needs to travel through the bloodstream to reach the target organ, and its distribution is affected by factors such as vascular permeability and plasma protein binding rate. Animal experiments can directly reflect the efficiency of drug distribution in the body and the drug concentration in the target organs, whereas cell experiments cannot demonstrate this process. The drug concentration in the in vitro cell experiment is the local concentration directly acting on the cells. However, for drugs in the body, the absorption efficiency differences caused by the administration route, as well as the clearance ability of the body towards the drugs, need not be taken into account. Animal experiments can accurately establish the dose–effect relationship that conforms to the in vivo reagents.
Authors’ Contributions
X.D. conceived and designed the project. S.-Y.L. and G.-L.W. implemented the methods and analyzed the data. C.-Y.S., H.-Y.C., and H.L. drafted the article. Y.-F.C. and Q.G.W. revised the article. All the authors read and approved the final article.
Footnotes
Acknowledgment
The authors thank the support of the Classical Prescription Basic Research Team of the Beijing University of Chinese Medicine.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This work was supported by the Natural Science Foundation of Xiamen (No. 3502Z20227108), Natural Science Foundation of Fujian Province (No. 2024J011370), 2025 Xiamen Municipal Key Clinical Specialty Construction Project (Xiawei Medical Administration 2023 No. 485), Panyu Science and Technology Plan Project (No. 2022-Z04-022), Guangdong Basic and Applied Basic Research Foundation (No. 2023B1515120075), and Guangdong Provincial Key Laboratory of Research on Emergency in TCM (No. 2023B1212060062).
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.