
Abstract
Mild cognitive impairment (MCI) lacks approved disease-modifying therapies. Classical multicomponent prescriptions may act on convergent neurobiological nodes. We combined network pharmacology with in vivo testing to evaluate Yizhi Dihuang Decoction (YZDHD). Constituents were curated from traditional chinese medicine systems pharmacology database and analysis platform (TCMSP) and high-throughput experiment- and reference-guided database of Traditional Chinese Medicine (HERB under blood–brain barrier-aware SwissADME criteria. Targets were inferred, intersected with MCI genes, organized into STRING and MCODE networks, and examined by Gene Ontology and Kyoto Encyclopedia of Genes and Genomes enrichment. Structure-based docking evaluated ligand–protein interactions across network-identified hub targets and ranked complexes by predicted binding energy. Predictions were tested in a D-galactose mouse model using the Morris water maze and novel object recognition, hippocampal histology with hematoxylin and eosin and Nissl staining, transmission electron microscopy, and molecular readouts by Western blot and quantitative reverse transcription polymerase chain reaction (qRT-PCR), including PI3K, p-AKT/AKT, p-mTOR/mTOR, LC3-II/LC3-I, and p62/SQSTM1. We identified 152 bioavailable compounds and 381 overlapping targets that converged on hub kinases including AKT1, PIK3CA, PIK3CD, and mTOR; docking supported feasible engagement. In vivo, YZDHD improved spatial learning and recognition memory, preserved hippocampal cytoarchitecture and mitochondrial integrity, increased LC3-II/LC3-I, decreased p62/SQSTM1, and reduced activation indices of AKT and mTOR. YZDHD ameliorates MCI-like deficits by rebalancing PI3K–AKT–mTOR signaling and restoring autophagy-related activity. Signals for mitogen-activated protein kinase (MAPK), hypoxia-inducible factor 1 (HIF-1), epidermal growth factor receptor (EGFR), and toll-like receptor 4 (TLR4) broaden the mechanistic hypothesis space and warrant targeted follow-up.
Keywords
Introduction
Mild cognitive impairment (MCI) occupies the boundary between healthy aging and dementia and has emerged as a common and growing public health concern in later life. 1 Diagnosis requires objective impairment in one or more cognitive domains with largely preserved everyday function; however, heterogeneity across settings, comorbidities, and cognitive profiles complicates prognosis and the design of clinical trials. 2 Although neuropsychological algorithms differ across clinics and cohorts, their shared aim is early risk detection with structured longitudinal follow-up. Demographic aging is projected to markedly increase the number of individuals living with cognitive impairment, while scalable interventions that reliably alter trajectories at the population level remain limited. 3 Against this backdrop, clarifying operational phenotypes and follow-up frameworks, improving the precision of early risk stratification and outcome prediction, and embedding generalizable assessment and management pathways within routine care represent the central needs for contemporary MCI research and practice.
There is currently no medicine specifically licensed for MCI. Management focuses on risk reduction, comorbidity control, and nonpharmacological support. Exercise programs and combined physical–cognitive training produce modest but clinically useful gains, although effects vary with dose, modality, and baseline status. 4 Computerized cognitive training can improve memory outcomes, and multisession protocols show benefits over months to years. 5 Yet, durable effects remain uneven, and translation at scale is challenging. Biology helps explain why MCI reflects network disruption rather than a single lesion: proteostasis stress, tau-related injury, neuroinflammation, cholinergic deficits, mitochondrial strain, and early synaptic failure interact and compensate. 6 Approaches that modulate several pathways in concert are therefore attractive. Among upstream nodes, the PI3K–AKT–mTOR axis regulates autophagic flux, a core mechanism for neuronal homeostasis. When the axis is overactive, autophagosome initiation is suppressed and degradative capacity falls; when rebalanced, clearance improves. 7 In tissue studies, LC3 lipidation (LC3-II/LC3-I ratio), together with reduced p62/SQSTM1, is used—following current guidelines—as an operational signature of enhanced autophagy. These concepts have practical value in hippocampal vulnerability and aging models, and they inform target selection for early-stage interventions.
Traditional Chinese medicine takes a systems perspective. Classical formulae combine multiple herbs to act across linked axes of pathology. Yizhi Dihuang Decoction (YZDHD) is one such prescription. It is an innovation that draws on two established formulae—Dihuang Yin Zi and Kongsheng Zhenzhong Dan—and adapts them for practice. Dihuang Yin Zi nourishes yin and replenishes essence, while Kongsheng Zhenzhong Dan calms the spirit and supports sleep. In YZDHD, prepared Rehmannia glutinosa (Gaertn.) DC. replenishes kidney essence; Cornus officinalis Siebold et Zucc., Morinda officinalis F.C.How, Cistanche deserticola Y.C.Ma, Alpinia oxyphylla Miq., and Cinnamomum cassia (L.) J.Presl support yang; Dendrobium nobile Lindl., Ophiopogon japonicus (Thunb.) Ker Gawl., Schisandra chinensis (Turcz.) Baill., Chinemys reevesii (Gray), and Fossilia ossis mastoid support yin; Acorus tatarinowii Schott and Polygala tenuifolia Willd. open the orifices and resolve phlegm; Prunus persica (L.) Batsch, Carthamus tinctorius L., and Salvia miltiorrhiza Bunge promote blood flow. Modern studies suggest anti-inflammatory, antioxidant, microcirculatory, and neuroprotective actions for constituent herbs, 8 but claims require transparent pipelines and prospective validation.
Network pharmacology—linking chemistry to targets, pathways, and interaction networks—provides a tractable route to generate testable, mechanism-level hypotheses for complex formulae when applied with careful curation and bias control. Network pharmacology can connect multicomponent therapy to coherent biology. It helps prioritize targets that sit at pathway junctions and predicts plausible ligand–protein interactions before bench work. In this study, we used network pharmacology to interrogate YZDHD for MCI: we identified bioavailable constituents, inferred putative protein targets, intersected these with MCI-related genes, mapped protein–protein interaction (PPI) hubs, and performed functional enrichment and structure-based docking to nominate core pathways. We then validated the working model in vivo. A D-galactose mouse model provided an aging-like context to assess behavior, hippocampal histology and ultrastructure, and molecular indices of autophagic flux—LC3-II/LC3-I and p62/SQSTM1—together with activity markers along the PI3K–AKT–mTOR axis. Our goal is to offer a clear, testable rationale for how a classical, multicomponent formula might engage autophagy-permissive signaling relevant to MCI, while treating other network-suggested routes as exploratory leads pending targeted tests.
Materials and Methods
Screening of YZDHD constituents and target prediction
Constituents of YZDHD were retrieved from the TCMSP and HERB databases.9,10 Compounds were initially filtered by oral bioavailability ≥ 30% and drug-likeness ≥ 0.18. A secondary screen incorporated Lipinski’s Rule of Five 11 and pharmacokinetic criteria with blood–brain barrier (BBB) permeability: on SwissADME, 12 we required molecular weight <500 Da, rotatable bonds ≤ 10, hydrogen-bond acceptors ≤10, hydrogen-bond donors ≤5, gastrointestinal absorption classified as “High,” and BBB permeability predicted as “Yes.” SMILES strings of retained compounds were standardized via PubChem 13 and submitted to SwissTargetPrediction 14 for target inference, retaining putative targets with probability > 0. Protein targets were then normalized using the UniProt database. 15
Collection of MCI-associated targets
Genes associated with MCI were retrieved from GeneCards 16 using the keyword “mild cognitive impairment.” Candidate entries were filtered by the platform’s disease-to-gene association score, and those with a score >10 were retained for subsequent analyses to ensure high confidence relevance to MCI pathobiology.
Identification of therapeutic targets of YZDHD for MCI and network visualization
Predicted targets of YZDHD constituents were intersected with the MCI gene set. Overlaps were assessed in FunRich 3.1.4 17 and visualized by Venn diagrams. Shared targets were imported into STRING 18 to build a PPI network with a confidence threshold ≥0.900; disconnected nodes were removed to optimize topology. A multilayer “YZDHD–compound–target” network was constructed in Cytoscape 3.10.3, 19 and topological features were computed via NetworkAnalyzer. Compounds were ranked by degree, and a top-30 degree threshold was applied to define core constituents of YZDHD against MCI, yielding 35 core compounds due to ties at the cutoff.
Prioritization of core targets
Therapeutic targets were analyzed in Cytoscape 3.10.3 using the MCODE plugin to identify densely connected functional modules. After parameter optimization, settings were degree cutoff = 2, k-core = 2, max depth = 100, and node score cutoff = 0.2. Nodes within the highest-scoring modules were designated as core targets implicated in YZDHD’s effects on MCI.
GO and KEGG enrichment analyses
Core targets underwent Gene Ontology (GO) enrichment using the DAVID database 20 with organism set to Homo sapiens and significance at p < 0.05. Terms were summarized for biological process (BP), molecular function (MF), and cellular component (CC) and visualized as bubble plots. Gene identifiers were then converted to Ensembl formats via BioMart, and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment was performed on the OmicShare platform. 21 Pathways with p < 0.001 were considered significant; the top 20 by enrichment were visualized using bubble and circular hierarchy plots to outline putative mechanisms of YZDHD in MCI.
Molecular docking
The three-dimensional (3D) structures of the core compounds were downloaded from PubChem. Protein crystal structures with ligand-binding domains and a resolution better than 2.5 Å were obtained from the Protein Data Bank (PDB). 22 After receptor preparation by removing water molecules and optimizing polar hydrogens, docking was performed on the DockThor platform. 23 The search grid was set to 40 Å × 40 Å × 40 Å, centered on the binding pocket, and 500 Monte Carlo sampling runs were executed. Complexes were ranked by binding free energy, and the top five were visualized in PyMOL 2.4.0. 24 Hydrophobic contacts and key hydrogen bond interactions were examined to evaluate conformational stability.
Reagents and resources
For immunoblotting, proteins were extracted in SDS lysis buffer (Beyotime, P0013G) and quantified with a BCA kit (Beyotime, P0010S). Equal amounts were resolved on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels (BolaiBio, BP112/BP108) and transferred to PVDF membranes (Immobilon-P, Merck, IPVH00010). Membranes were blocked in skim milk (Biosharp, BS102-100 g) and probed with primary antibodies against LC3 (Proteintech, 14600-1-AP), p-AKT (Proteintech, 28731-1-AP), AKT (Proteintech, 10176-2-AP), PI3K p85 (Proteintech, 11748-1-AP), p62/SQSTM1 (Proteintech, 10222-1-AP), p-mTOR (CST, 2971S), mTOR (CST, 2972S), and β-actin (Beyotime, AF5003), followed by HRP-conjugated goat antirabbit IgG (Abbkine, AS014). Signals were developed with enhanced chemiluminescence (Meilunbio, MA0186-1) and molecular weight referenced with prestained markers (Thermo Fisher, 26616; 26625). TBST (Biosharp, BL315B) and antibody diluent (Boster, AR1017) were used as indicated.
For RNA analyses, total RNA was isolated with RNAeasy™ Animal RNA kit (Beyotime, R0027), reverse-transcribed using BeyoRT™ III First-Strand cDNA Mix (Beyotime, D7182M), and quantified by qPCR with ChamQ Universal SYBR qPCR Master Mix (Vazyme, Q711).
Equipment and software
Behavioral testing used the Morris water maze (MWM), novel object recognition (NOR) arenas, and tracking software (Xinruan, Shanghai, China). Microscopy included a rotary microtome (Leica, Germany) and an upright light microscope (Olympus, Japan). Ultrastructural imaging employed a transmission electron microscope (TEM; Hitachi, Japan) with carbon-support copper grids (Zhongjing Keyi, China). Electrophoresis and transfer were performed on mini vertical PAGE and blotting systems (LiuYi, China) with dedicated power supplies, and chemiluminescence was captured on a Tanon 5200 imaging system (Tanon, China). Additional equipment included a precision electronic balance (Ohaus, USA), paraffin and cryo embedding systems (Kedi, China), a plate reader (Laibao, China), a tissue homogenizer and dry block heater (Allsheng, China), a refrigerated high-speed centrifuge (Beckman, USA), a horizontal shaker (LiuYi, China), and a real-time PCR instrument (LongGene, China).
Preparation of YZDHD
Prepared according to the 2020 Pharmacopoeia of the People’s Republic of China, YZDHD was supplied by the hospital pharmacy. In brief, the herbs were soaked in eight volumes of water for 30 minutes, boiled for 40 minutes, filtered, and the residue was decocted six additional times for 30 minutes each. The combined filtrates were concentrated to a stock decoction equivalent to 6.37 g crude herb per mL. Before administration, this stock was diluted with saline to prepare working solutions corresponding to the low-, medium-, and high-dose regimens used in the animal experiment, while maintaining a constant gavage volume of 0.1 mL/10 g body weight. The decoction was stored at 4°C for short-term use and at −20°C in aliquots for long-term storage and was mixed thoroughly before gavage.
Animals and experimental design
Male C57BL/6J mice, 2 months old, weighing 20 ± 2 g, SPF grade, were obtained from Liaoning Changsheng (license SCXK-2020-0001) and housed at 22 ± 2°C, 40%–60% humidity, 12-hour light/dark cycle with ad libitum food and water. Animals acclimated for 7 days. All procedures were approved by the Institutional Animal Care and Use Committee (no. 2024091201).
MCI was induced by intraperitoneal injection of D-galactose in normal saline at a volume of 0.1 mL/10 g body weight once daily for 4 consecutive weeks. After this period, eligible mice were randomized into a control group (C), a model group (M), a positive drug group (Y), and YZDHD low-/medium-/high-dose groups (YZDHD-d/m/h; n = 8). YZDHD doses were 15.925, 31.85, and 63.7 g/kg/day; the donepezil dose was 0.65 mg/kg/day, calculated according to the human-to-mouse conversion factor of 9.1; gavage volume was 0.1 mL/10 g. During the subsequent 4-week treatment phase, D-galactose administration was maintained in all groups except C and was given in parallel with vehicle, donepezil, or YZDHD. Dosing and behavioral testing were conducted at consistent times to minimize circadian effects. Investigators were blinded to group allocation during testing and analysis.
Behavioral testing
Behavioral testing was performed during the light phase at fixed times by investigators blinded to group allocation. Task order was counterbalanced across cages, and the apparatus was cleaned with 70% ethanol between trials to minimize olfactory cues. Tracking was conducted with video analysis software. Spatial navigation was assessed with MWM in a circular pool of 100 cm diameter containing a hidden platform of 10 cm diameter; water temperature was maintained at 22 ± 2°C. Animals underwent five consecutive days of acquisition with start positions varied across trials, and escape latency, path length, and swim speed were recorded. At 24 hours after the last acquisition session, a 60-second probe trial was conducted with the platform removed. Primary outcomes were time spent in the target quadrant and the number of crossings over the former platform location. Recognition memory was assessed with NOR in a 40 × 40 × 40 cm arena. After habituation to the empty arena, animals were exposed to two identical objects placed equidistant from the walls. In the test phase, one object was replaced with a novel object; object identity and position were counterbalanced across animals. Exploration was defined as the nose oriented toward an object at close range without climbing. The discrimination index was calculated as the time spent exploring the novel object divided by the total exploration time for the novel and familiar objects.
Histopathology and ultrastructure
At the endpoint, mice were euthanized by intraperitoneal injection of sodium pentobarbital at 150 mg/kg body weight. Brains were collected immediately. A portion of the hippocampus was fixed in 4% paraformaldehyde, embedded in paraffin, and used for hematoxylin and eosin (H&E) and Nissl staining. Another portion was fixed in 2.5% electron microscopy fixative at 4°C under light protection for TEM. The remaining tissue was snap-frozen at −80°C for molecular assays. Paraffin-embedded blocks were cut into sections with a thickness of 5–7 µm according to standard protocols. Imaging and quantification were performed in the CA1 subfield, CA3 subfield, and dentate gyrus. For TEM, samples were fixed in glutaraldehyde, postfixed in osmium tetroxide, and dehydrated. They were embedded in Epon 812 resin and cut into ultrathin sections with a thickness of approximately 90 nm. Sections were stained with uranyl acetate and lead citrate and examined for mitochondrial and autophagosome ultrastructure.
Western blotting
Hippocampal lysates were prepared in SDS buffer and quantified by BCA. Equal protein was separated by SDS-PAGE, transferred to PVDF membranes, blocked with 5% milk, and incubated with primary and HRP-conjugated secondary antibodies per manufacturer’s instructions; signals were developed by ECL. Targets included PI3K, p-AKT/AKT, p-mTOR/mTOR, p62/SQSTM1, and LC3-II/LC3-I, with β-actin as the loading control. Band intensities were quantified in ImageJ and normalized; control and treated samples were run on the same blot.
qRT-PCR
Total hippocampal RNA was purified using a column method, treated with DNase, and assessed for quality. The absorbance ratio A260/280 ranged from 1.8 to 2.1. cDNA was synthesized with the BeyoRT kit. qPCR was carried out with SYBR Green chemistry in 20 μL reaction volumes, using primers at a final concentration of 0.2 μM. The cycling program was as follows: initial denaturation at 95°C for 30 seconds, followed by 40 cycles of 95°C for 10 seconds and 60°C for 30 seconds. A melt-curve confirmed the presence of single specific products. Relative gene expression was determined by the 2−ΔΔ Ct method with normalization to Actb. Amplification efficiency ranged from 90% to 110% with R2 ≥ 0.99, as verified by standard curves. Target genes included Sqstm1 and Mtor (Table 1).
Primer Sequences Used for qRT-PCR

Statistical analysis
Data are presented as mean ± standard error of the mean. MWM acquisition was analyzed by two-way repeated-measures ANOVA with Greenhouse–Geisser correction when appropriate; probe trials, novel NOR, Western blot, qPCR, and histological quantifications were analyzed by one-way ANOVA with Tukey post hoc. If assumptions were violated, Kruskal–Wallis with Dunn post hoc or Welch ANOVA was used. For multiple markers/genes, Benjamini–Hochberg FDR correction was applied. All tests were two-tailed with p < 0.05 considered significant. Analyses and plots were performed in GraphPad Prism 10.1.2; normality/variance tests were run in SPSS 26.0. Full statistical details and any predefined exclusion criteria are provided in the Supplementary Data.
Results
Acquisition of active components and targets of YZDHD
We interrogated TCMSP and HERB to delineate the chemical space of the 16-herb formula YZDHD. Ten herbs were indexed in both repositories (Morinda officinalis F.C.How, Salvia miltiorrhiza Bunge, Carthamus tinctorius L., Cistanche deserticola Y.C.Ma, Cinnamomum cassia (L.) J.Presl, Cornus officinalis Siebold et Zucc., Acorus tatarinowii Schott, Rehmannia glutinosa (Gaertn.) DC., Prunus persica (L.) Batsch, and Schisandra chinensis (Turcz.) Baill.), whereas six (Chinemys reevesii (Gray), Fossilia ossis mastoid, Ophiopogon japonicus (Thunb.) Ker Gawl., Dendrobium nobile Lindl., Alpinia oxyphylla Miq., and Polygala tenuifolia Willd.) were retrieved exclusively from HERB. After taxonomy harmonization and prespecified ADME/BBB filters, we retained 152 nonredundant active compounds across the 16 herbs. Representative sources included Acorus tatarinowii Schott with 29 active compounds, Salvia miltiorrhiza Bunge with 21, Carthamus tinctorius L. with 20, Cinnamomum cassia (L.) J.Presl with 20, Alpinia oxyphylla with 18, and Polygala tenuifolia with 15. Morinda officinalis F.C.How and Chinemys reevesii (Gray) each contributed one active, and Fossilia ossis mastoid contributed none after filtering (Supplementary Table S1).
Putative protein targets were inferred for each compound using SwissTargetPrediction. Low-confidence calls were discarded and target lists were deduplicated across herbs, yielding 1027 unique human targets from an initial compound–target mapping of 8040 associations. These 1027 targets underpinned all subsequent network construction, enrichment analyses, and docking.
Screening of MCI-related targets
Based on the GeneCards database, 11,907 candidate genes associated with MCI were initially obtained. By applying a high-relevance threshold, 3224 genes were retained. These standardized target names were used for subsequent analyses.
Screening of therapeutic targets and construction of the PPI network
By integrating the targets of YZDHD active components with MCI-related genes, 381 common targets were identified using FunRich 3.1.4 (Fig. 1A). A PPI network was then constructed using the STRING platform, containing 1058 interaction relationships. After applying a confidence threshold and removing isolated nodes, a core PPI network consisting of 307 targets was obtained (Fig. 1B).
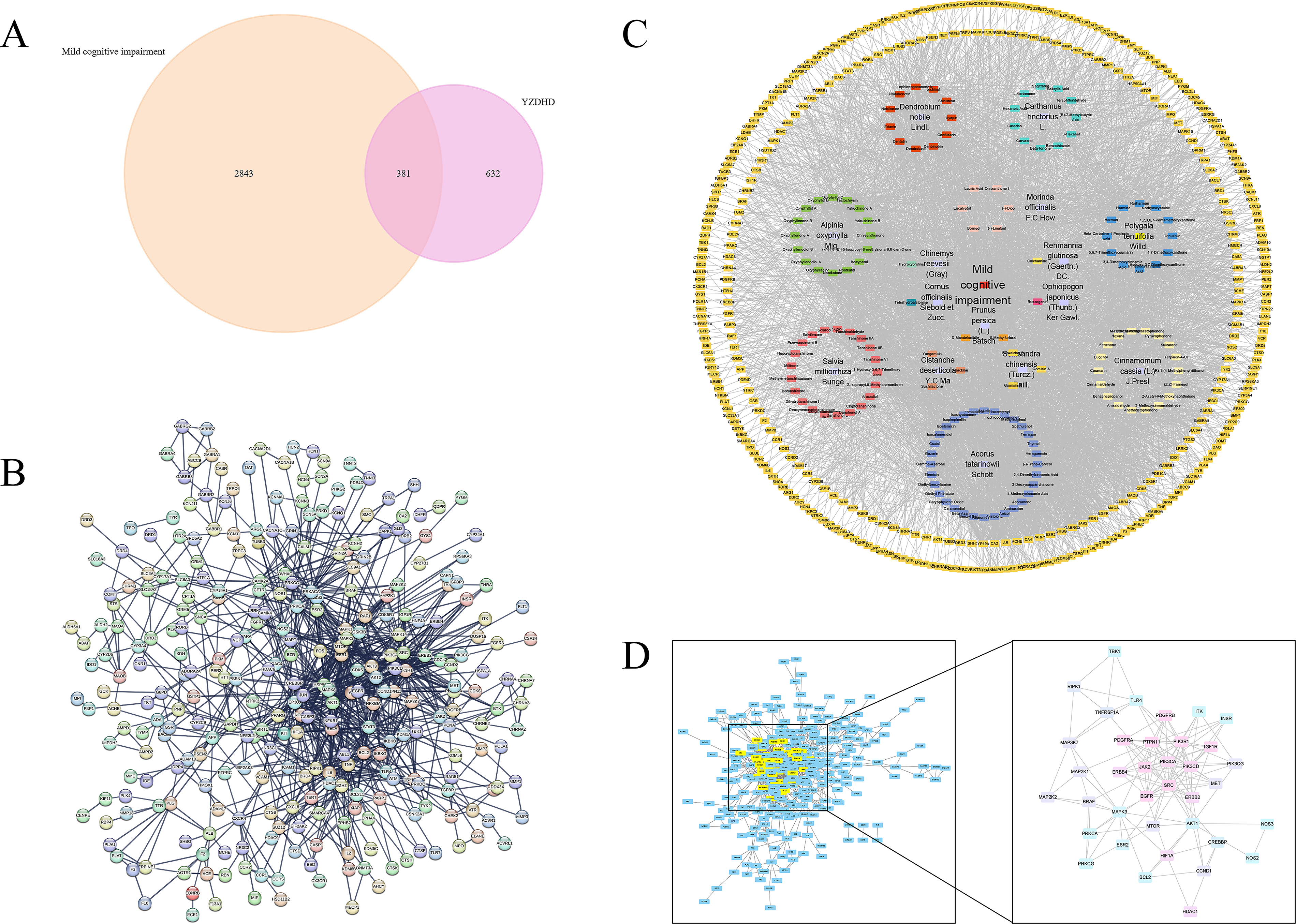
Network pharmacology-based identification of YZDHD targets for MCI.
Screening of core compounds and core therapeutic targets
A multidimensional “YZDHD–compound–therapeutic target” network model was constructed using Cytoscape 3.10.3 (Fig. 1C). The network consisted of 536 nodes and 3862 edges, with topological parameters indicating an average node degree of 14.41, a network diameter of 6, a characteristic path length of 2.424, and a network centralization of 0.688, which reflected high connectivity and regulatory significance of core nodes. Based on the top-30 degree-ranking threshold, 35 core compounds were identified, including Orchinol, Erianin, D-Mandelonitrile, Yakuchinone A, Gomisin A, and Nodakenetin. Due to ties at the 30th rank, six compounds—Tanshinone IIb, Isoelemicin, Colchamine, Oxyphyllol B, 1-Hydroxy-3,7-Dimethoxyxanthone, and 3,4,5-Trimethoxycinnamic Acid—shared the same degree value and were therefore also retained within this 35-compound core set (Table 2). Using the MCODE plugin in Cytoscape, modular analysis of the PPI network containing 307 therapeutic targets was performed, which revealed 17 functional modules in total (Fig. 1D). The highest-scoring module, with a score of 7.833, contained 37 core targets, including AKT1, MAPK3, EGFR, BCL2, HIF1A, PIK3CA, MTOR, SRC, and TLR4 (Supplementary Table S2).
Core Active Compounds of YZDHD Against MCI, Their Network Degree Values, and Source Herbs

MCI, mild cognitive impairment; YZDHD, Yizhi Dihuang Decoction.
Enrichment analysis of core therapeutic targets
Functional annotation of core therapeutic targets was performed using the DAVID database, with p < 0.05 as the significance threshold. A total of 444 GO enrichment results were obtained, including 303 BPs, mainly enriched in insulin-like growth factor receptor signaling pathway, epidermal growth factor receptor signaling pathway, positive regulation of ERK1/2 cascade, protein autophosphorylation, and positive regulation of phosphatidylinositol 3-kinase/protein kinase B (PI3K–AKT) signal transduction. There were 106 MFs, mainly enriched in protein tyrosine kinase activity, ATP binding, transmembrane receptor protein tyrosine kinase activity, and histone H3Y41 kinase activity. There were 35 CCs, mainly enriched in receptor complex, plasma membrane, cytosol, and phosphatidylinositol 3-kinase complex. The GO enrichment analysis results were visualized as bar charts, and the top 20 results for BPs, MFs, and CCs were visualized as bubble plots (Fig. 2A–C).
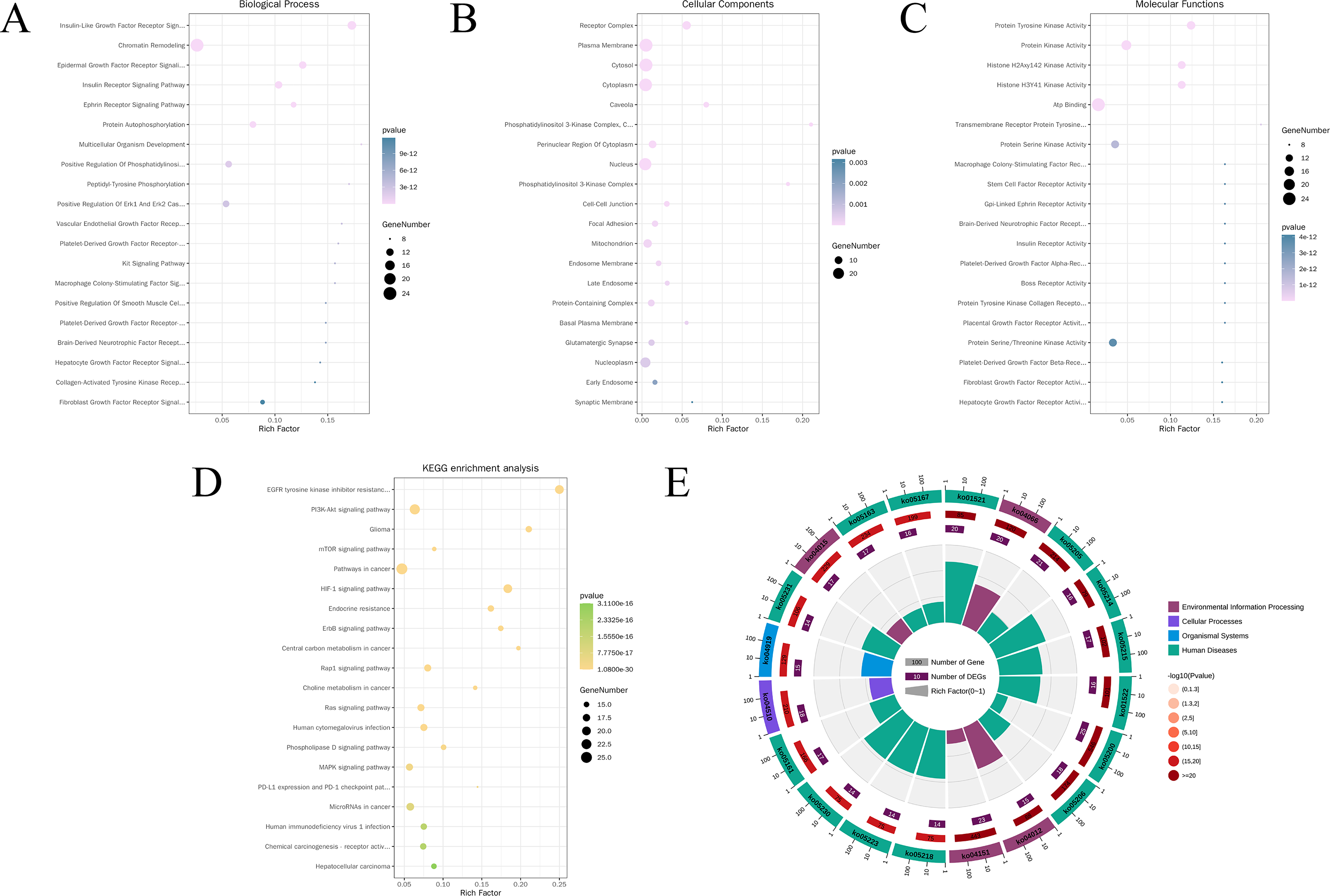
GO and KEGG enrichment analysis of core therapeutic targets.
KEGG pathway enrichment analysis identified 150 significantly associated pathways, including PI3K–AKT signaling pathway, MAPK signaling pathway, EGFR tyrosine kinase inhibitor resistance, mTOR signaling pathway, and HIF-1 signaling pathway. In addition, entries such as pathways in cancer, glioma, and choline metabolism in cancer suggested that the core targets may participate in MCI pathology by modulating neuroinflammation, energy metabolism imbalance, and apoptosis across multiple diseases. The top 20 pathways with the smallest p-values were visualized using bubble plots and circular hierarchy diagrams (Fig. 2D, E and Table 3).
KEGG Pathway Enrichment of Core Therapeutic Targets

KEGG, Kyoto Encyclopedia of Genes and Genomes.
Molecular docking results of core therapeutic targets and core compounds
The 3D SDF structures of 35 core compounds were downloaded from PubChem, and the structures of 37 core target proteins were obtained from the PDB database. These were imported into the DockThor platform for molecular docking analysis. According to docking score criteria, scores < 0 kcal/mol indicate spontaneous binding between compounds and proteins; scores <−4.25 kcal/mol suggest moderate binding activity; and scores <−7.0 kcal/mol indicate strong binding activity. The docking results showed that the core compounds of YZDHD had good binding affinity with the core therapeutic targets, with all docking scores <−4.25 kcal/mol, and an average docking score of −7.29 kcal/mol. Among them, 58.1% had scores ≤−7.0 kcal/mol, indicating significant binding activity. Notably, AKT1 showed the lowest average docking score with compounds (−8.61 kcal/mol), suggesting strong binding potential and its likely key role in YZDHD treatment of MCI.
Analysis of the docking heatmap (Fig. 3A) revealed the top five compound–target combinations: Suchilactone–PI3K p110δ (PIK3CD) with a docking score of −11.48 kcal/mol (Fig. 3B); suchilactone–AKT1 with a docking score of −11.12 kcal/mol (Fig. 3C); acoramone–mTOR with a docking score of −10.91 kcal/mol (Fig. 3D); 3,4,5-trimethoxycinnamic acid–PI3K p110δ (PIK3CD) with a docking score of −10.89 kcal/mol (Fig. 3E); and elemicin–PI3K p110α (PIK3CA) with a docking score of −10.87 kcal/mol (Fig. 3F). These results were visualized using PyMOL 2.4.0, and the stability of molecular interactions was analyzed by examining hydrogen bond numbers and bond lengths.
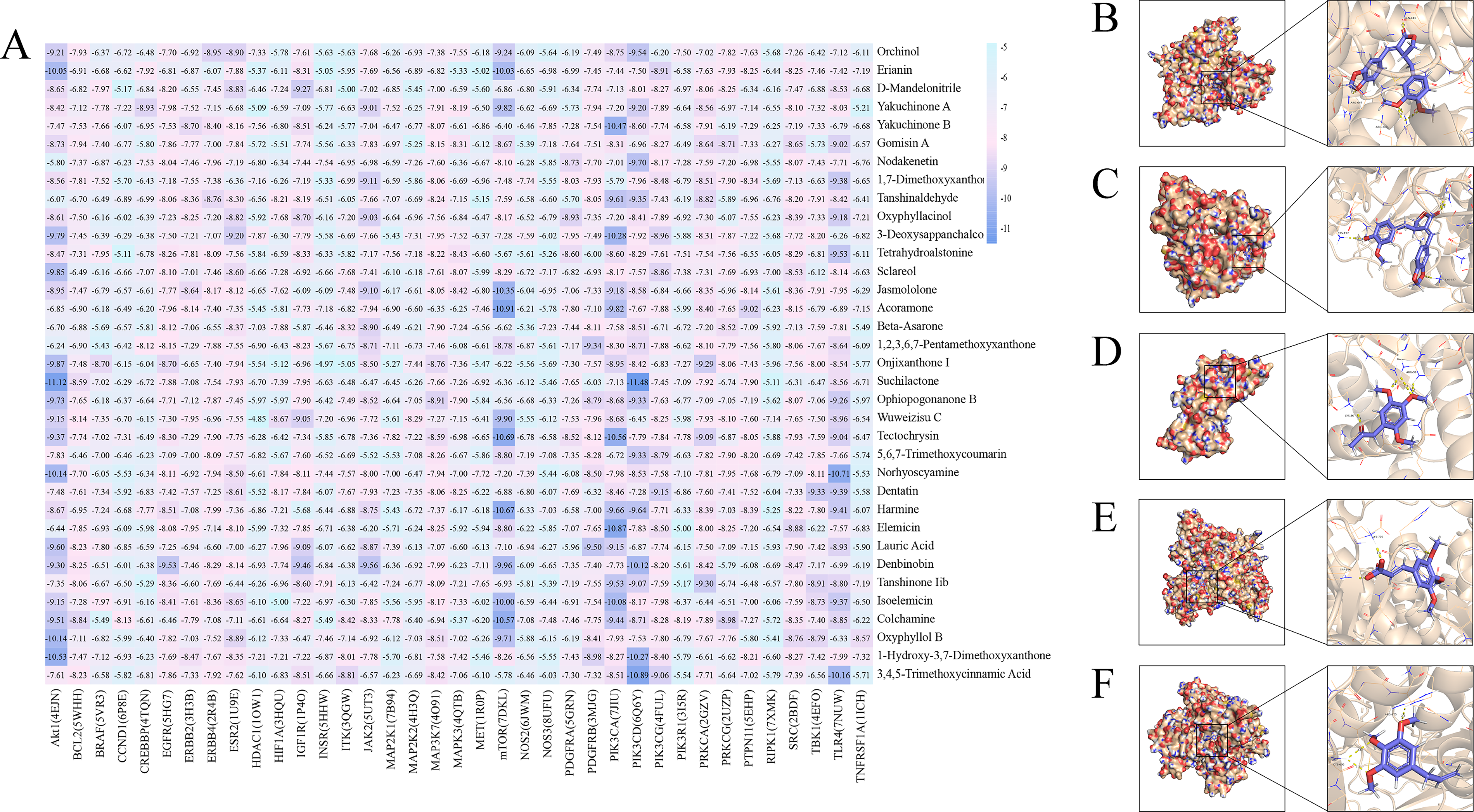
Molecular docking analysis of core YZDHD compounds with hub therapeutic targets.
Effects of YZDHD on spatial memory ability in MCI mice
In the navigation trial (Fig. 4A–D), compared with M, the C, Y, and YZDHD-d/m/h groups exhibited significantly shorter escape latency, reduced total swimming distance, and increased swimming speed (p < 0.05 or p < 0.01). In the probe trial (Fig. 4E–F), the number of platform crossings was significantly increased in C, YZDHD-m, and YZDHD-h (p < 0.01), while moderate increases were observed in Y and YZDHD-d (p < 0.05). Together, these results indicate that YZDHD treatment effectively improved spatial learning and memory in MCI mice.
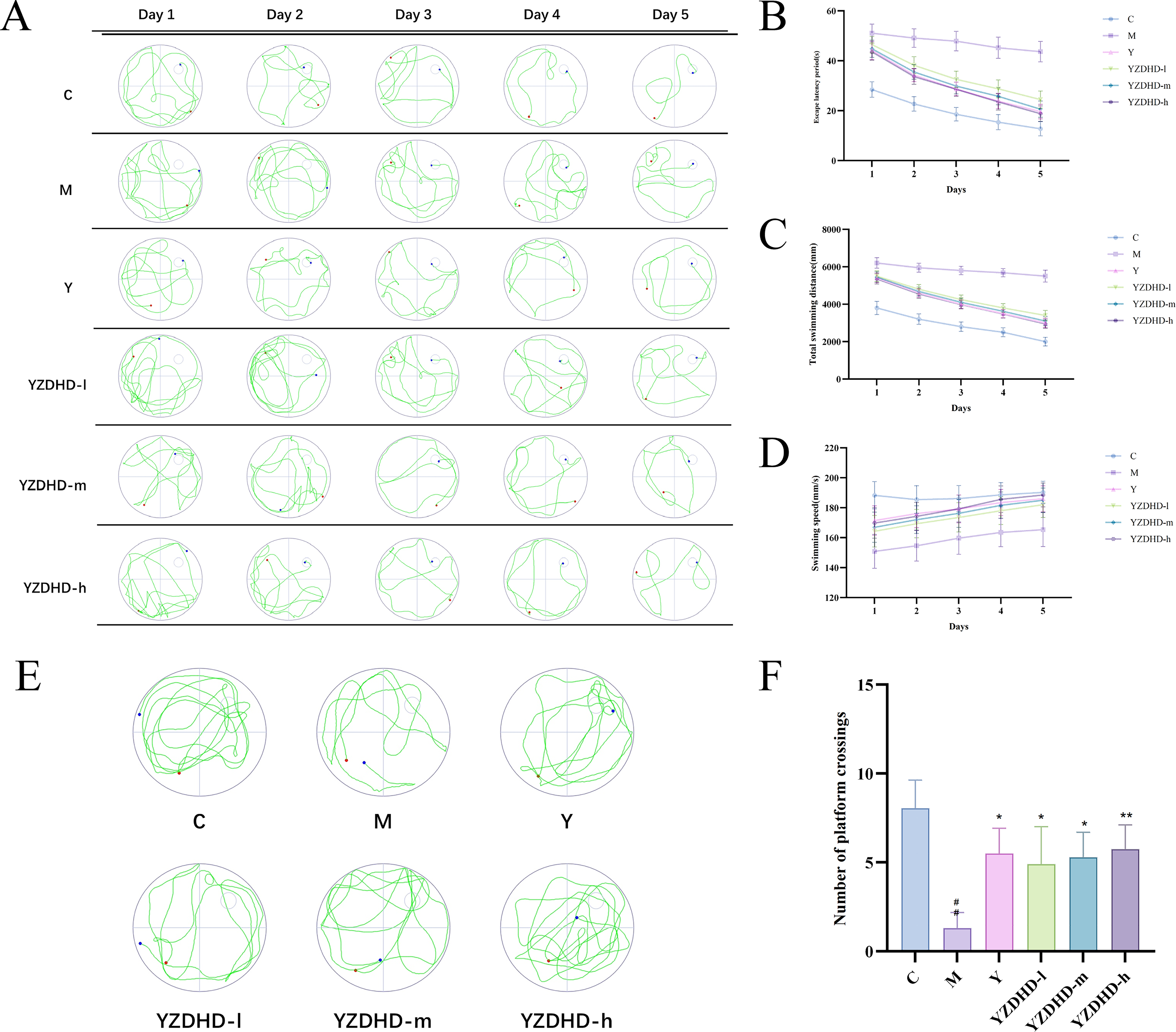
Effects of YZDHD on spatial learning and recognition memory in D-galactose-induced MCI mice.
During the familiarization phase, no significant differences were observed among groups in the exploration time of objects A1 and A2 (p > 0.05). In the test phase, compared with M, C and all YZDHD-d/m/h groups showed a significant increase in the discrimination index for the novel object (p < 0.01), indicating a marked improvement in recognition memory (Fig. 4G–H).
Effects of YZDHD on hippocampal neurons and autophagosomes in MCI mice
H&E staining
Hippocampal neurons in C were densely arrayed with clear lamination, large round nuclei, and uniformly distributed chromatin. M exhibited loose and disordered layers with blurred cytoarchitecture, nuclear pyknosis, enhanced cytoplasmic eosinophilia, and scattered vacuoles. YZDHD attenuated neuronal loss and improved cellular morphology, with YZDHD-m, YZDHD-h, and Y showing more orderly neuronal arrangement, fewer abnormal cells, and reduced pyknosis. YZDHD-d displayed only mild improvement without a significant difference from M (Fig. 5A).
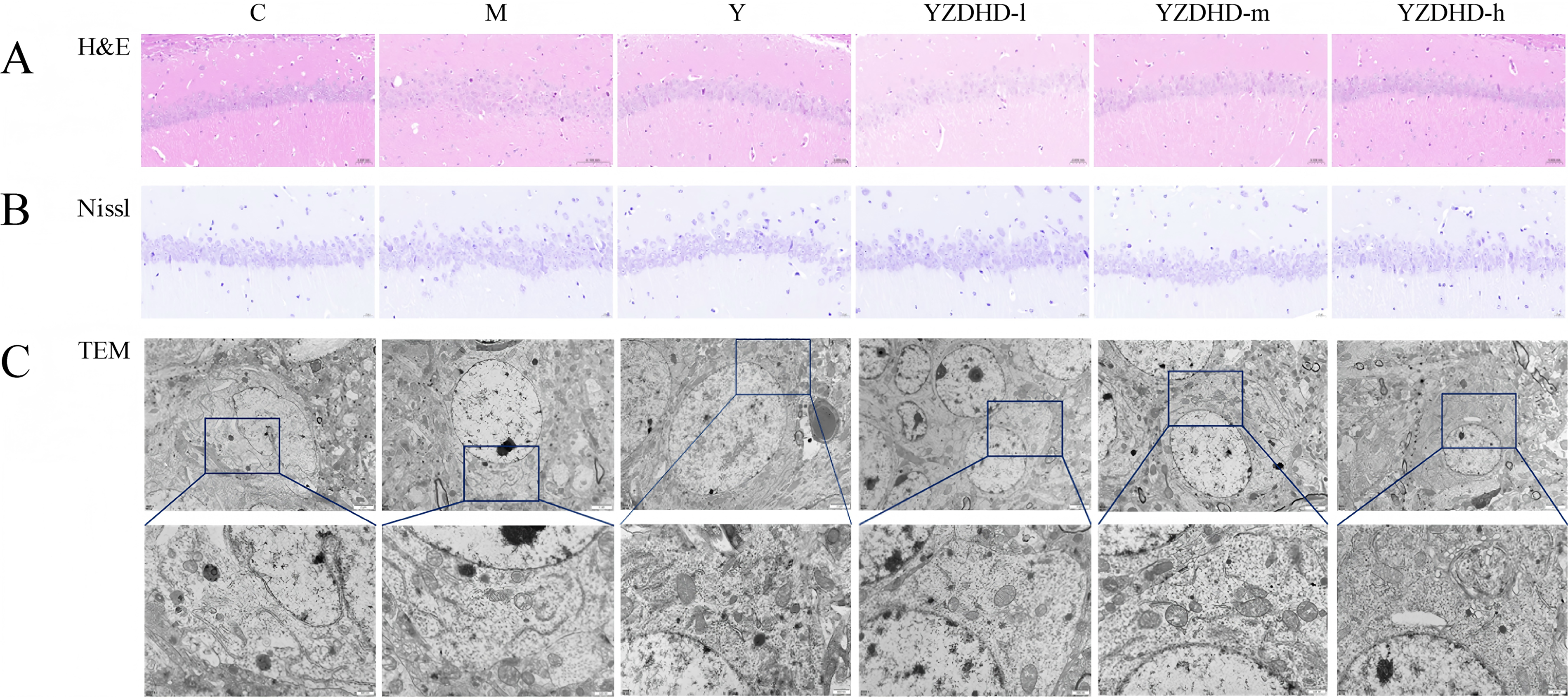
Histopathological and ultrastructural improvements in the hippocampus of D-galactose-induced MCI mice following YZDHD treatment.
Nissl staining
Neurons in C displayed normal morphology, regular arrangement, and abundant Nissl bodies with typical striped or granular patterns. M showed a marked decrease in Nissl body density, vacuolar degeneration, and nuclear pyknosis. Y exhibited partial restoration with residual localized vacuolation. YZDHD-d showed slight increases in Nissl density with persistent vacuolar change. YZDHD-m demonstrated pronounced recovery with fewer vacuolated areas, and YZDHD-h approximated the C phenotype with abundant Nissl bodies and intact nuclei (Fig. 5B).
TEM
On transmission electron microscopy, M exhibited a reduced number of autophagosomes, frequent single-membrane or ruptured vesicles with content leakage, and mitochondria showing swelling, disrupted cristae, and vacuolar degeneration. In contrast, C displayed intact double-membrane autophagosomes with well-preserved contents and normal mitochondrial architecture. YZDHD restored autophagosome abundance and membrane integrity; in YZDHD-m and YZDHD-h, autophagosomes presented complete double membranes with tightly enclosed cargo, accompanied by alleviated mitochondrial swelling and more orderly cristae (Fig. 5C).
Effects of YZDHD on the expression of PI3K, p-AKT/AKT, p-mTOR/mTOR, p62/SQSTM1, and LC3-II/LC3-I in MCI mice
Western blot analysis
Relative to M, C showed markedly lower PI3K, p-AKT/AKT, and p-mTOR/mTOR (all p < 0.01), accompanied by reduced p62/SQSTM1 (p < 0.05 or p < 0.01). Across YZDHD-d/m/h, PI3K, p-AKT/AKT, and p-mTOR/mTOR were significantly decreased (p < 0.05 or p < 0.01), the LC3-II/LC3-I ratio increased (p < 0.01), and p62/SQSTM1 declined (p < 0.01; Fig. 6A and B).
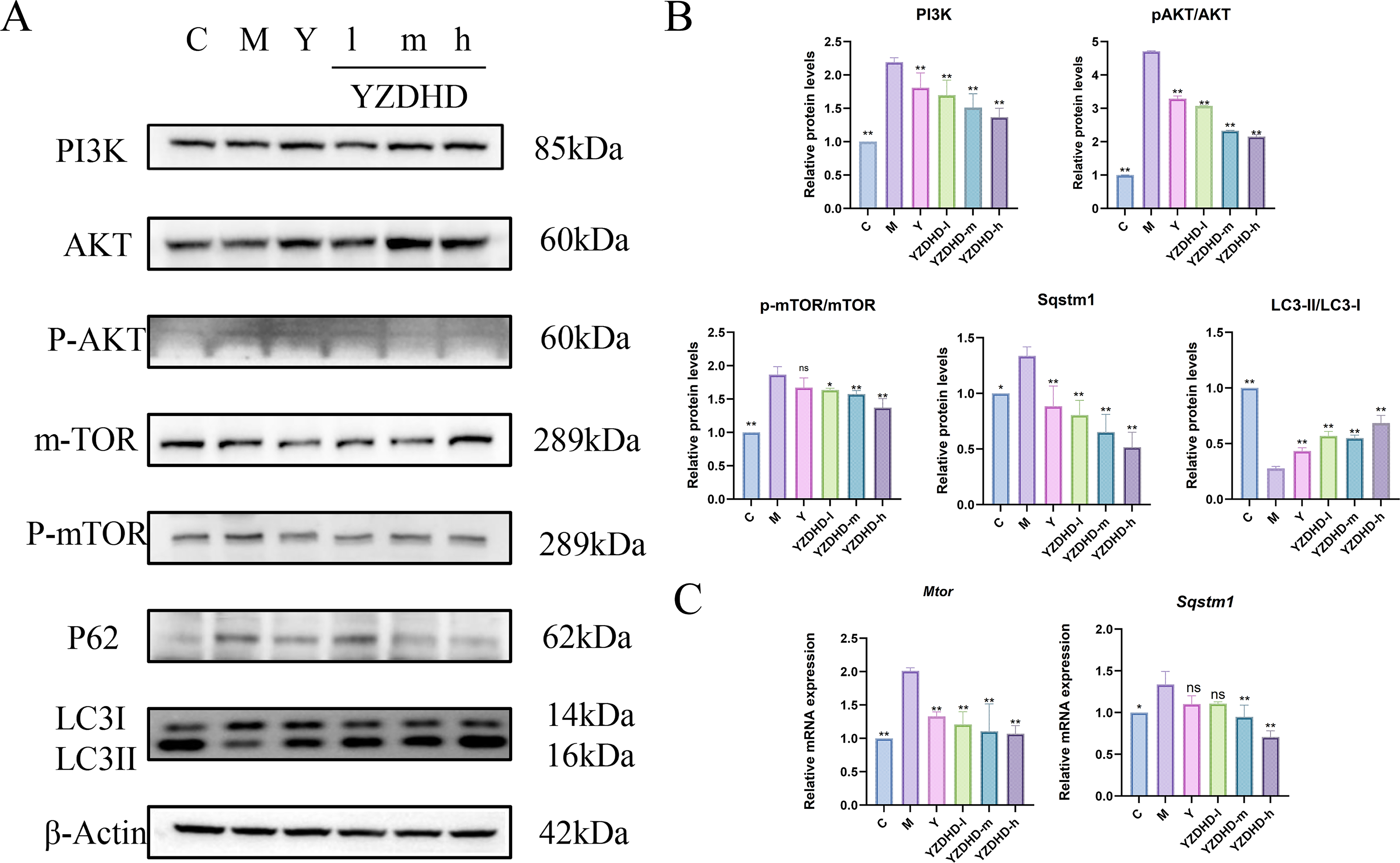
Effects of YZDHD on autophagy-related and PI3K–AKT–mTOR pathway proteins and gene expression in the hippocampus of MCI mice.
qRT-PCR analysis
Compared with M, C exhibited lower Sqstm1 mRNA (p < 0.05) and a pronounced reduction in Mtor mRNA (p < 0.01). In YZDHD-m and YZDHD-h, Sqstm1 mRNA was significantly decreased (p < 0.01), whereas Y and YZDHD-d showed no significant change (p > 0.05). Mtor mRNA was significantly downregulated in all YZDHD-d/m/h groups (Fig. 6C).
Discussion
MCI represents an intermediary stage between normal aging and dementia, sharing neuropathological features with Alzheimer’s disease while differing in severity and cognitive domain specificity. 25 Leveraging network pharmacology integrated with in vivo validation, our study indicates that YZDHD mitigates MCI-like phenotypes by rebalancing the PI3K–AKT–mTOR axis toward an autophagy-permissive state. A systems-level analysis highlighted 35 active compounds and 37 hub targets; the network also suggested ancillary involvement of MAPK and HIF-1 signaling. Molecular docking supported feasible compound–target engagement at AKT1, PIK3CA/PIK3CD, and mTOR. Across behavioral, histological, ultrastructural, and molecular outcomes, the pharmacological profile cohered with this mechanistic interpretation, providing convergent evidence rather than isolated findings.
Our BBB-aware screening, disease–target intersection, PPI modularity, and GO/KEGG enrichment prioritized nodes centered on PI3K–AKT–mTOR, with network-suggested signals pointing to MAPK and HIF-1. Docking results supported a testable hypothesis: YZDHD tempers upstream PI3K–AKT input and relieves mTOR-mediated autophagy repression, thereby improving neuronal proteostasis. In vivo findings tracked with this prediction—behavioral improvements were accompanied by histological and molecular signatures of autophagy restoration—supporting a model in which autophagy recovery is a proximate driver of cognitive benefit. Beyond this core mechanism, the network suggested immune–metabolic axes involving EGFR and TLR4, which remain hypothesis-generating and warrant targeted validation before firm conclusions can be drawn. 26
The PI3K–AKT–mTOR axis orchestrates neuronal survival, protein synthesis, synaptic plasticity, and—critically—autophagy. Previous studies indicate that activation of PI3K–AKT can promote plasticity and trophic responsiveness under specific conditions. 27 However, persistent mTORC1 activation suppresses autophagy initiation and contributes to proteostatic failure in neurodegeneration. 28 In disease-relevant states, dysregulated PI3K–AKT–mTOR signaling is tightly linked to abnormal tau phosphorylation (e.g., via GSK-3β) and impaired proteostatic clearance, ultimately damaging synapses and neurons.7,29 Pharmacological downregulation of mTOR restores autophagy and improves cognition across models by facilitating clearance of toxic species. 30 These findings are consistent with the proposed mechanistic framework. YZDHD attenuated the activation of AKT and mTOR pathways and improved autophagy-related activity, as evidenced by an increased LC3-II/LC3-I ratio and reduced p62/SQSTM1 expression. Collectively, the data indicate that YZDHD rebalances this pathway toward an autophagy-permissive state that supports neuronal proteostasis. Isoform-specific regulation may be relevant: the network and docking prioritized PIK3CA and PIK3CD, suggesting potential isoform-sensitive tuning of AKT input in vulnerable circuits.
Autophagy impairment is increasingly recognized as a bottleneck in MCI and related syndromes, with convergent evidence for defects in autophagosome biogenesis, trafficking, and lysosomal degradation.31,32 Because LC3-II accumulation may reflect either increased autophagosome formation or impaired downstream degradation when interpreted in isolation, LC3-II/LC3-I should be evaluated together with autophagic substrate markers such as p62/SQSTM1 and morphological evidence. Early in disease, autophagy aids clearance of misfolded proteins and supports neuronal survival; with progression, autophagic flux becomes impeded, undermining degradative capacity and network resilience. 28 The lipidation conversion of LC3-I to LC3-II reflects autophagosomal membrane elongation and autophagosome formation. p62/SQSTM1, as an adaptor protein linking LC3 and polyubiquitinated proteins, can enter autophagosomes together with autophagic substrates and is subsequently degraded in autolysosomes. Therefore, an increased LC3-II/LC3-I ratio accompanied by decreased p62/SQSTM1 levels generally suggests that autophagosome formation and substrate degradation may be improved, although this should be interpreted cautiously in the context of autophagic flux. 33 In the present study, the model group showed insufficient LC3 lipidation and accumulation of the substrate p62/SQSTM1, suggesting impaired autophagy-related processes. To further evaluate autophagy-related changes, we focused on the LC3-II/LC3-I ratio. The results showed that YZDHD significantly increased the LC3-II/LC3-I ratio and reduced p62/SQSTM1 levels, suggesting that YZDHD may promote autophagosome formation and improve autophagic substrate degradation. Together with the restoration of double-membrane autophagosomes observed at the ultrastructural level, these findings support a potential ameliorative effect of YZDHD on hippocampal autophagy abnormalities at both the molecular-marker and morphological levels. It should be noted that dynamic autophagic flux assays were not performed in this study; therefore, the present findings provide indirect evidence of improved autophagy-related processes rather than definitive proof that YZDHD restores complete autophagic flux. Future studies should further validate the regulatory effect of YZDHD on autophagic flux using lysosomal inhibitors or the mRFP-GFP-LC3 fluorescent reporter system.
Beyond PI3K–AKT–mTOR, the network suggested MAPK and HIF-1 involvement. Aberrant MAPK signaling—particularly ERK and JNK branches—can amplify neuroinflammation, oxidative stress, and tau phosphorylation, leading to synaptic dysfunction and cognitive decline. 34 The presence of MAPK3 among hub targets motivates further exploration of whether YZDHD moderates overactive inflammatory signaling. HIF-1 integrates hypoxic adaptation with glycolytic programming, mitochondrial homeostasis, and antioxidant defenses.35,36 Enrichment signals and docking-inferred interactions raise the possibility that YZDHD enhances metabolic resilience under stress. Immune–metabolic nodes such as EGFR and TLR4 were also highlighted. EGFR signaling contributes to synaptogenesis and glial homeostasis, and its modulation has been associated with improved recovery in brain injury models. 37 TLR4 is a key initiator of neuroinflammation; dampening TLR4-driven responses can improve cognition and histopathology in preclinical settings, but YZDHD’s influence here awaits dedicated validation. 38 JAK2/STAT3, another enriched axis, regulates glia-derived inflammatory outputs and represents an additional hypothesis-level mechanism for future investigation. 39 Among these predicted pathways, PI3K–AKT–mTOR signaling was prioritized for in vivo validation because it showed a direct mechanistic association with the autophagy-related abnormalities investigated in this study. Network topology analysis highlighted high-connectivity targets within this axis, including AKT1, PIK3CA/PIK3CD, and MTOR, while molecular docking further suggested favorable interactions between core YZDHD compounds and AKT1, PI3K isoforms, and mTOR. More importantly, PI3K–AKT–mTOR signaling provides an experimentally tractable link to the autophagy-related markers examined in this study, including p62/SQSTM1 and LC3-II/LC3-I. Therefore, we examined PI3K–AKT–mTOR pathway-related proteins to preliminarily explore its potential involvement in the effects of YZDHD. It should be noted that network pharmacology findings remain predictive, and whether MAPK, HIF-1, TLR4, and JAK2/STAT3 pathways are also involved requires further validation.
Individual constituents of YZDHD—such as tanshinones from Salvia, β-asarone from Acorus, Alpinia-derived compounds, and Cistanche polysaccharides—have been reported to lower oxidative stress and apoptosis, enhance cholinergic tone, promote neurotrophic support, and repair synaptic function. 40 However, single-ingredient strategies often face limitations due to pharmacokinetics and restricted target scope. Classical prescriptions employ multicomponent synergy to engage parallel and convergent nodes, increasing efficacy while reducing single-agent liabilities.41,42 Composite formulae such as Taohong Siwu Tang and Huanglian Jiedu Tang have shown additive or synergistic effects on synaptic plasticity, inflammatory balance, and oxidative injury with improved cognitive outcomes in models. 43 YZDHD, with core compounds and hub targets spanning PI3K–AKT–mTOR and network-suggested MAPK, HIF-1, EGFR, and TLR4, is structurally positioned to exert such network-level regulation.
Limitations and Perspectives
The D-galactose model reproduces salient features of MCI-like biology—oxidative stress, aberrant protein phosphorylation, synaptic and neuronal injury, and mild-to-moderate cognitive impairment—making it useful for early-stage therapeutic interrogation. 44 Our behavioral, histological, ultrastructural, and molecular endpoints were mutually coherent, supporting construct and face validity. Nevertheless, the model does not encompass the full spectrum of human pathology. Extending validation to aged cohorts and tau-focused transgenic lines would strengthen generalizability and determine whether YZDHD’s benefits translate across etiologically diverse contexts.
Standardization and chemical characterization remain important limitations of the present study. Although the YZDHD used in this study was prepared according to the 2020 edition of the Pharmacopoeia of the People’s Republic of China and provided by the hospital pharmacy, this study did not include an HPLC fingerprint or LC–MS/MS-based chemical characterization of the administered YZDHD batch. In addition, quantitative analysis of batch-defining chemical markers was not performed, and the brain pharmacokinetics or blood–brain barrier permeability of YZDHD were not evaluated. Future work should establish LC–MS-based fingerprints for YZDHD, define quantitative markers for quality control, and determine in vivo pharmacokinetics and brain distribution to support reproducibility and dose selection.
Interpretation of docking requires caution. Docking predicts affinity under idealized conditions but does not by itself establish on-target engagement in brain tissue. Biophysical binding assays, target occupancy measurements, and phospho-proteomics in relevant brain regions would be informative to substantiate direct interactions with AKT1, PI3K isoforms, and mTOR.
Clinical translation will require careful staging. Human MCI exhibits considerable heterogeneity in both trajectory and underlying biology, making it necessary to conduct biomarker-anchored trials, such as stratification based on plasma or CSF tau levels and imaging markers of neurodegeneration, to evaluate whether YZDHD can slow cognitive decline or modify disease-associated pathways. The design of such studies in early-stage patients, integrating clinical outcomes with mechanistic biomarkers, has the potential to provide decisive evidence of both therapeutic efficacy and biological mechanisms. 45 In this study, the low, medium, and high doses of YZDHD were 15.925, 31.85, and 63.7 g/kg/day, respectively. The medium dose was designed as the mouse-equivalent dose converted from the clinical adult dose using the body-surface-area method and a human-to-mouse conversion coefficient of 9.1, while the low and high doses represented 0.5-fold and 2-fold of this equivalent dose, respectively. All doses refer to crude-herb equivalents rather than purified extract weight. This dosing regimen shows a certain correspondence with clinical application and may provide a preliminary reference for future clinical translation and dose optimization. Meanwhile, considering that interventions aimed at antiaging or cognitive improvement usually require long-term administration, the long-term safety, effective dose range, and safety window of YZDHD should be further clarified through systematic toxicological evaluation, dose-escalation studies, and clinical investigations.
Conclusion
By integrating network pharmacology with experimental validation, we show that YZDHD improves learning and memory, preserves hippocampal structure, and restores autophagy-related markers in a D-galactose model of MCI. The data support a mechanism centered on rebalancing PI3K–AKT–mTOR signaling toward an autophagy-permissive state. The network also suggested ancillary pathways—including MAPK, HIF-1, EGFR, and TLR4—that broaden the hypothesis space for future investigation. These findings substantiate a multicomponent, multitarget therapeutic paradigm consistent with classical formula theory and highlight YZDHD as a promising candidate for early intervention in MCI, meriting deeper mechanistic and clinical evaluation.
Authors’ Contributions
Z.H., T.M., and S.L. contributed to the experimental design, data acquisition, and initial data analysis. Y.B. participated in methodology development, data curation, and statistical analysis. R.Q. assisted with literature review, visualization, and article editing. J.C. conceived the study, supervised the entire research process, and revised the article critically for important intellectual content. All authors reviewed and approved the final version of the article.
Supplemental Material
sj-docx-1-rej-10.1177_15491684261460820 — Supplemental material for Yizhi Dihuang Decoction Ameliorates Mild Cognitive Impairment by Restoring Autophagy via PI3K–AKT–mTOR Modulation: In Vivo Validation and Network Pharmacology Analysis
Supplemental material, sj-docx-1-rej-10.1177_15491684261460820 for Yizhi Dihuang Decoction Ameliorates Mild Cognitive Impairment by Restoring Autophagy via PI3K–AKT–mTOR Modulation: In Vivo Validation and Network Pharmacology Analysis by Zhihua Hao, Tianwei Meng, Songzhe Li, Yifan Bu, Rui Qie, and Jing Chen
Supplemental Material
sj-docx-2-rej-10.1177_15491684261460820 — Supplemental material for Yizhi Dihuang Decoction Ameliorates Mild Cognitive Impairment by Restoring Autophagy via PI3K–AKT–mTOR Modulation: In Vivo Validation and Network Pharmacology Analysis
Supplemental material, sj-docx-2-rej-10.1177_15491684261460820 for Yizhi Dihuang Decoction Ameliorates Mild Cognitive Impairment by Restoring Autophagy via PI3K–AKT–mTOR Modulation: In Vivo Validation and Network Pharmacology Analysis by Zhihua Hao, Tianwei Meng, Songzhe Li, Yifan Bu, Rui Qie, and Jing Chen
Supplemental Material
sj-docx-3-rej-10.1177_15491684261460820 — Supplemental material for Yizhi Dihuang Decoction Ameliorates Mild Cognitive Impairment by Restoring Autophagy via PI3K–AKT–mTOR Modulation: In Vivo Validation and Network Pharmacology Analysis
Supplemental material, sj-docx-3-rej-10.1177_15491684261460820 for Yizhi Dihuang Decoction Ameliorates Mild Cognitive Impairment by Restoring Autophagy via PI3K–AKT–mTOR Modulation: In Vivo Validation and Network Pharmacology Analysis by Zhihua Hao, Tianwei Meng, Songzhe Li, Yifan Bu, Rui Qie, and Jing Chen
Footnotes
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This work was supported by
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.