
Abstract
The influence of sensory modulation on the early stage of information processing was investigated with a somatosensory contingent negative variation (CNV) paradigm. Whether or not even a somatosensory input as well as auditory or visual stimulus to one hemisphere elicits the symmetrical “early CNV” was also examined. Eleven normal individuals (3 males, 8 females) performed a conventional CNV paradigm with a click sound as the warning stimulus (WS) and a red light flash as the imperative stimulus (IS). Nine individuals (5 males, 4 females) did the somatosensory CNV paradigm with paired electrical stimuli as WS and IS. The subjects were instructed to press a button in response to IS as fast as possible. The early CNV amplitude was smaller and P300 latency was longer in somatosensory paradigm than conventional paradigm. In addition, the latency of P100 in a somatosensory paradigm was longer than that of N100 in a conventional paradigm. These findings suggest that the initiation of early detection, reflected by P100, the initiation of cognition, reflected by P300, and orienting response, reflected by early CNV, are delayed in a somatosensory CNV paradigm. Furthermore, all event-related potentials (ERPs) evoked by somatosensory stimuli showed a bilateral symmetry.
INTRODUCTION
The contingent negative variation (CNV), first described by Walter 1 in 1964, has been considered to be a correlate of the arousal level, expectancy, and the preparation of information processing. In 1976, Rohrbaugh et al. 2 distinguished the early from the late component of the CNV by using a long interstimulus interval (ISI). Since then, the early CNV has been considered a correlate of the orienting response.
On the other hand, Desmedt et al. 3 suggested that the receiving somatosensory cortical areas are deployed on the brain surface, whereas the auditory or visual receiving areas are more deeply located. Therefore, early components of the somatosensory event-related potentials (ERPs) can be easily recorded from the scalp electrodes. In this respect, a somatosensory CNV paradigm can be applied to the investigation of the early stage of information processing including the early detection, cognition and the orienting response. 2 However, little is known about the effect of somatosensory stimuli used as the warning stimulus (WS) and the imperative stimulus (IS) on each ERP, such as N100, P300 to WS and each CNV (early and late CNV). Especially, there is little agreement as to the influence of the sensory modulation on the early CNV related to the orienting response. 4 Furthermore, the hemispheric asymmetries of each CNV are still unclear.
In the present study, we examined the changes of ERPs in conjunction with the onset of somatosensory orienting response. We also studied whether or not even a somatosensory input to the contralateral hemisphere elicits the symmetrical “early CNV” as well as the other sensory inputs (auditory or visual stimulus).
MATERIALS AND METHODS
Eleven right-handed normal individuals (3 males and 8 females; mean age = 23.55, SD = 4.83 years) performed a conventional CNV paradigm. Nine normal individuals, including 1 left-handed, and 4 right-handed males and 4 right-handed females (mean age = 26.5, SD = 5.94 years), did the somatosensory CNV paradigm. The latter group included the same 7 subjects of the former group and matched the former group in age, educational, social and economic backgrounds. This study was approved by the Ethic Committee of Hirosaki University Hospital and all subjects gave their written informed consent before study. The details of each paradigm areas follows:
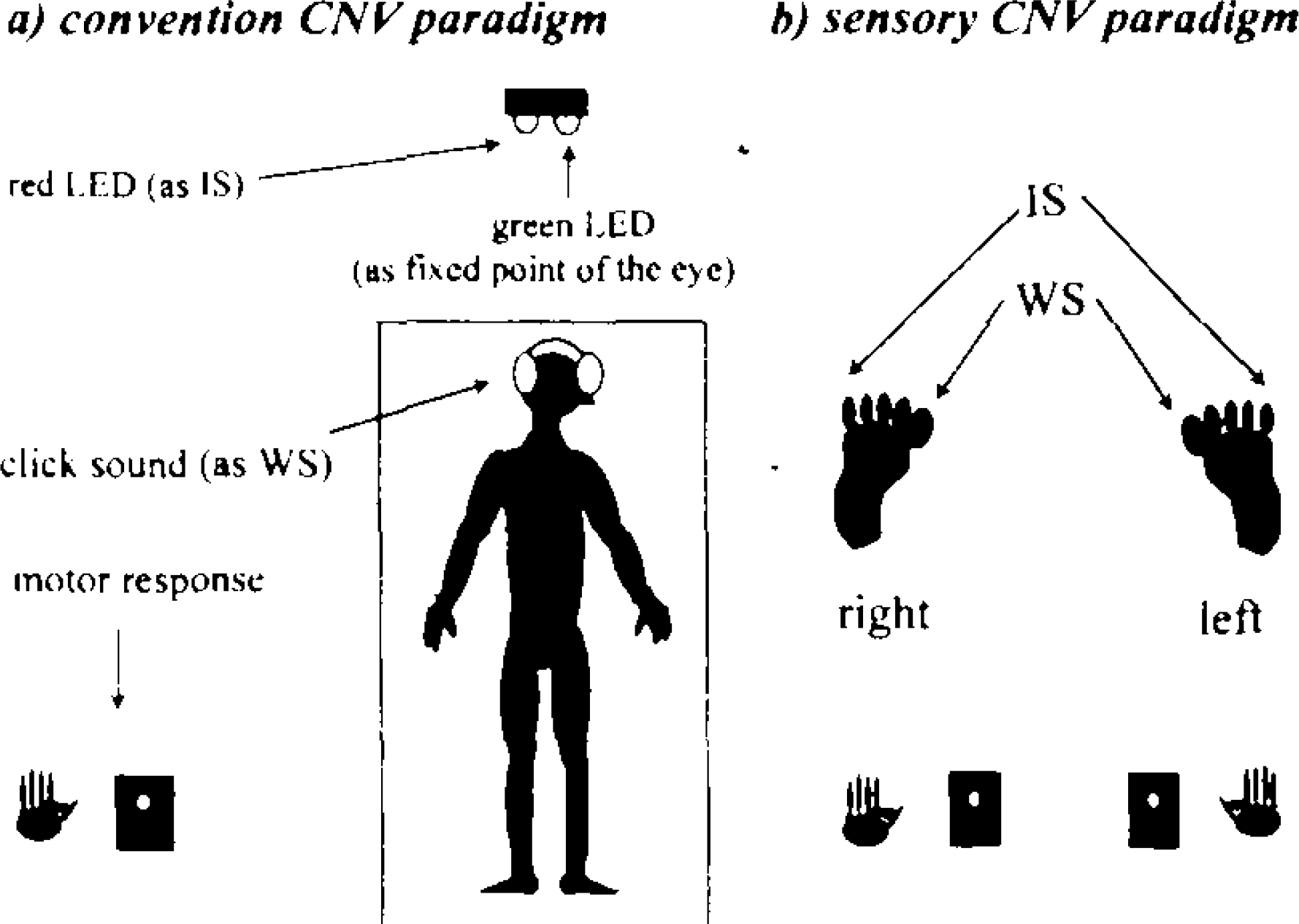
The block diagram of the present study.
Both paradigms consisted of an ISI of 2 s and an intertrial interval (ITI) ranging from 10 to 15 s at random. All subjects were required to refrain from blinking between WS and IS and to gaze at a gleaming green LED (à=565 nm) attached at the same point from which the red light flashes. The experiment was carried out in an electrically shielded sound-damped absolutely dark room. The EEG was recorded with Ag/AgCl electrodes at Fz, Cz, Pz, F3, F4, C3′ and C4′ (3 cm posterior C3 and C4 respectively) referred to linked earlobes. Vertical eye movements were measured by electrooculographic (EOG) recordings with electrodes placed above and below the right eye. An electrode was also placed on the right mastoid process to detect electrodermal activity (EDA) artifacts. EEG and EOG signals were recorded with a time constant of 5 s and a high-frequency cut-off of 60 Hz. Electrode impedance was maintained below 5 k Ω. Trials with premature (over 1000 ms) or erroneous responses as well as electrooculographic artifacts or electrodermal activity (either exceeded ± 75 μ V) were excluded from the analysis. RTs from IS onset to a button press were calculated only for these artifact-free trials, matched for each averaged wave. N100 and P300 were measured from 80 to 150 ms, and 250 to 500 ms following WS respectively. The early CNV was identified as the mean amplitude over a consecutive 200 ms, starting at 500 ms after WS, which included the peak amplitude of the early CNV, 2,5 and the late CNV was measured as the mean amplitude over the 200 ms preceding IS. The baseline was defined as the average EEG in a period of 1 s prior to the on set of WS.
The data of all ERPs and RTs were analyzed by the Wilcoxon rank-sum test.
RESULTS
Each trial number calculated for averaging was 20.36 ± 5.05 (mean ± SD) in the conventional CNV paradigm, 47.11 ± 12.47 in the somatosensory CNV paradigm with stimulation of the right toes and 44.78 ± 13.29 in the paradigm with stimulation of the left toes. Figure 2 shows the CNV recordings in both the conventional CNV paradigm and the somatosensory CNV paradigm with stimulation of the right toes. Figure 3 shows those in both the conventional paradigm and the somatosensory paradigm with stimulation of the lefttoes. The early CNV was small and P300 latency was delayed in the somatosensory CNV paradigm. In addition, instead of N100, P100 was recorded in the somatosensory CNV paradigm.
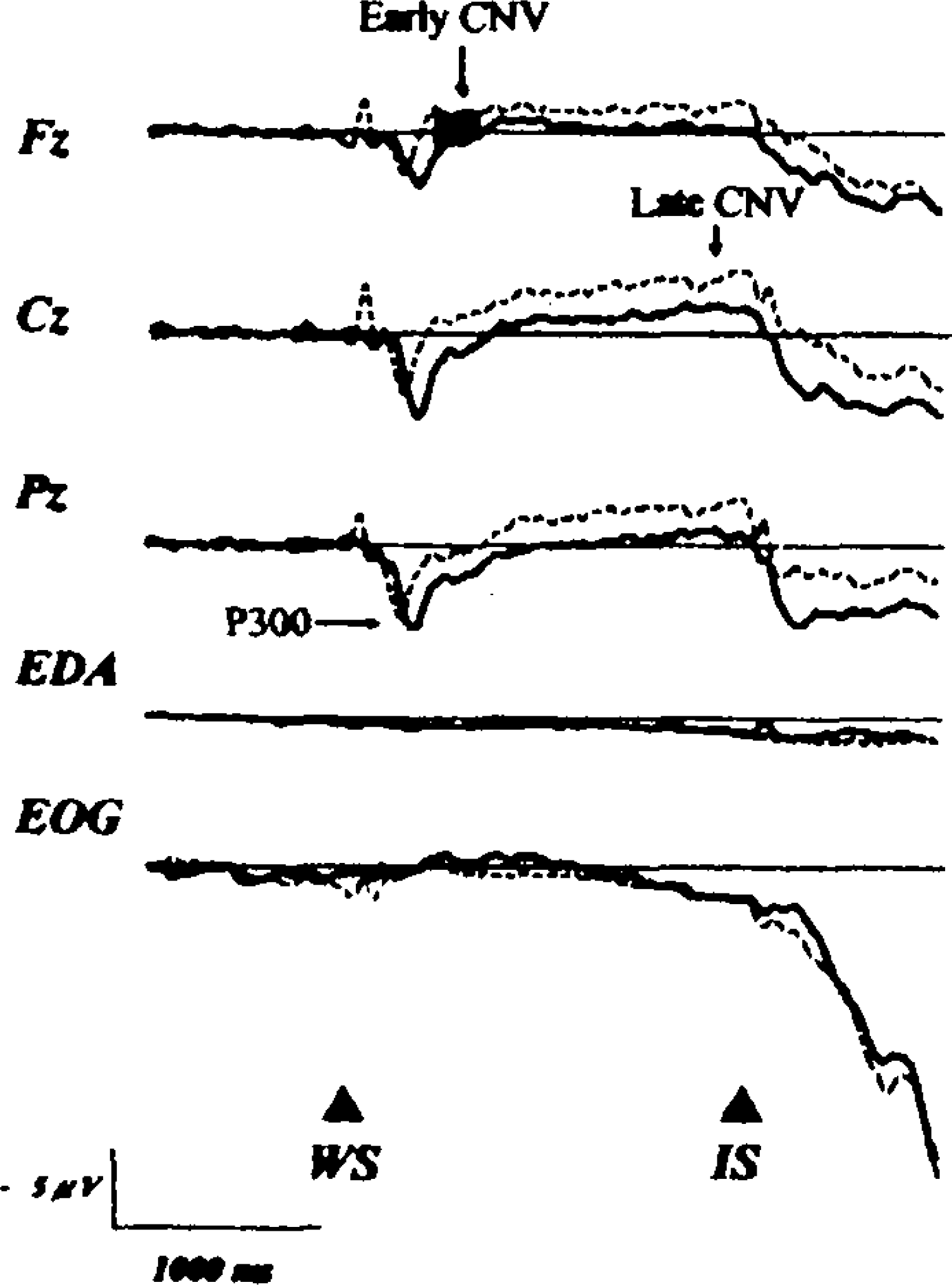
Grand-averaged waveforms, vertical EOG, and EDA in the conventional CNV paradigm (dotted line) and in the somatosensory CNV paradigm with right toes stimulation (thick line).
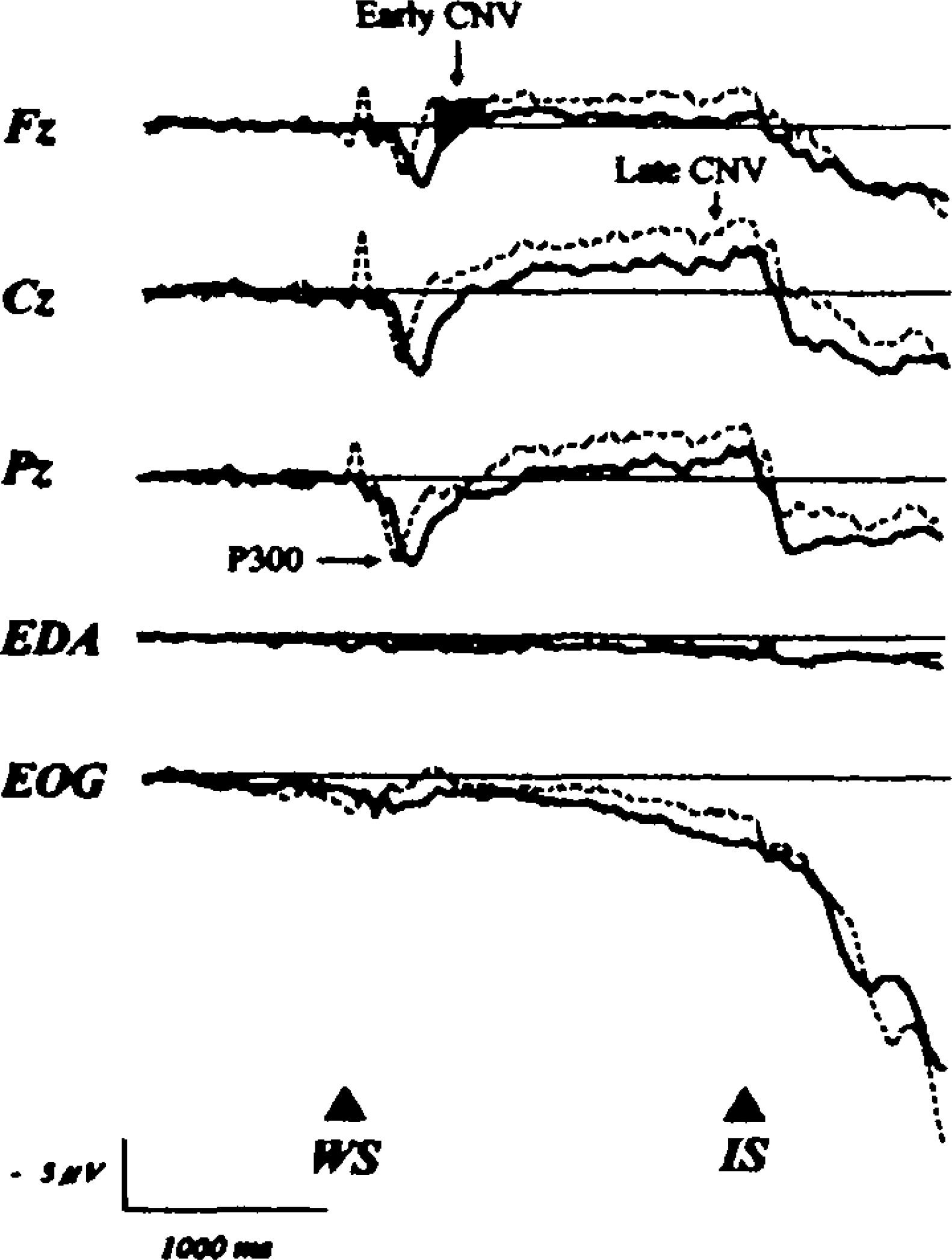
Grand-averaged waveforms, vertical EOG, and EDA in the conventional CNV paradigm (dotted line) and in the somatosensory CNV paradigm with left toes stimulation (thick line).
The early CNV amplitudes were significantly lower at Fz (two-tailed p < 0.03) and Cz (two-tailed p < 0.007) in the paradigm with stimulation of the right toes and at Fz (two-tailed p < 0.05) in the paradigm with stimulation of the left toes than in the conventional CNV paradigm (Figure 4).
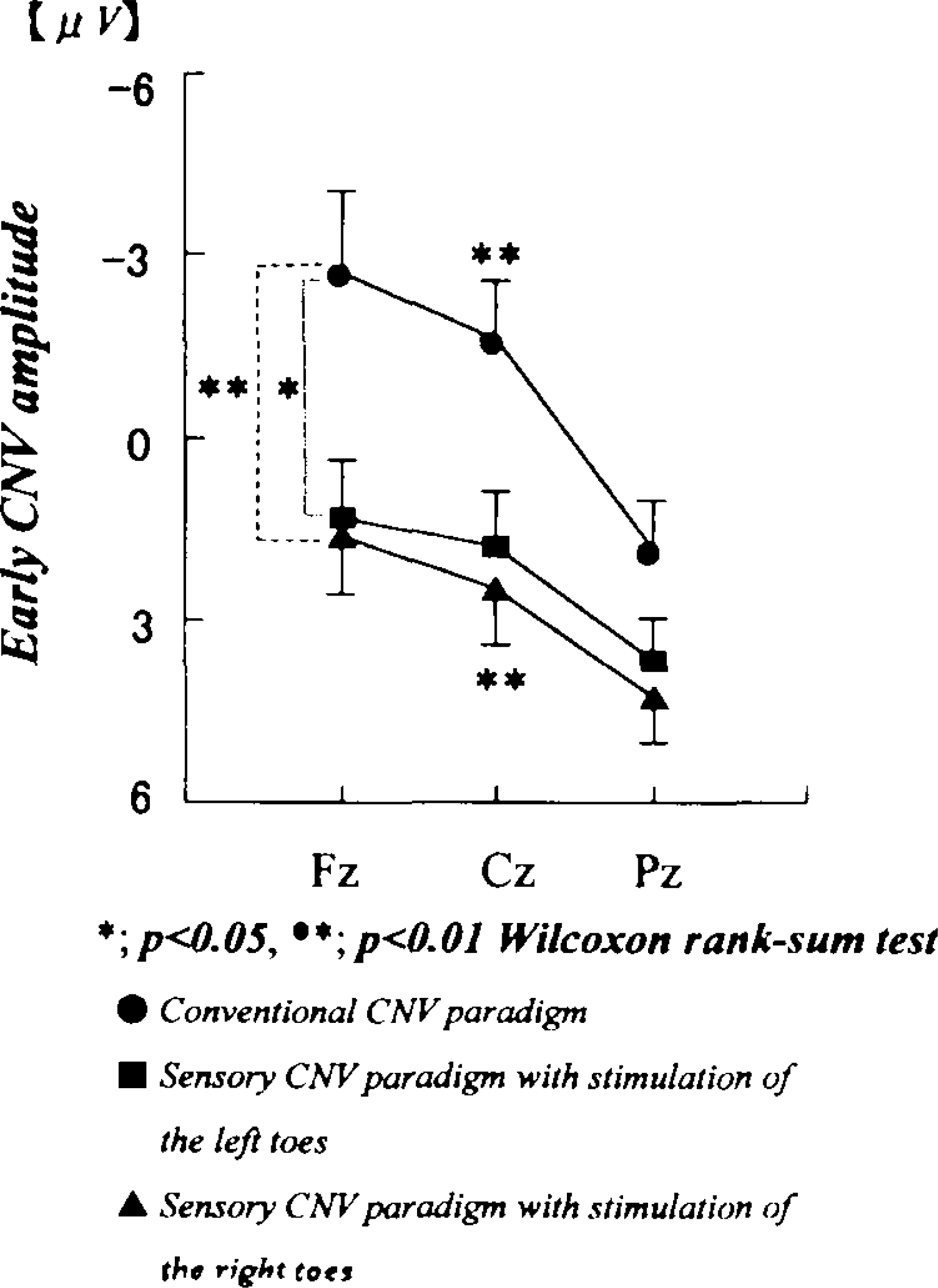
Differences in the early CNV amplitudes between the conventional CNV paradigm (closed circle) and the somatosensory paradigm with right-toes stimulation (filled triangle), and between the conventional paradigm and those of left-toes stimulation (filled square), respectively.
P300 was longer in latency at the midline electrode sites in the somatosensory CNV paradigm in the condition with stimulation of the right toes (two-tailed; Fz: p < 0.006; Cz: p < 0.004; Pz: p < 0.01), in those with stimulation of the left toes (two-tailed; Fz: p < 0.04; Cz: p < 0.04; Pz: p < 0.01) than in the conventional CNV paradigm (Figure 5).
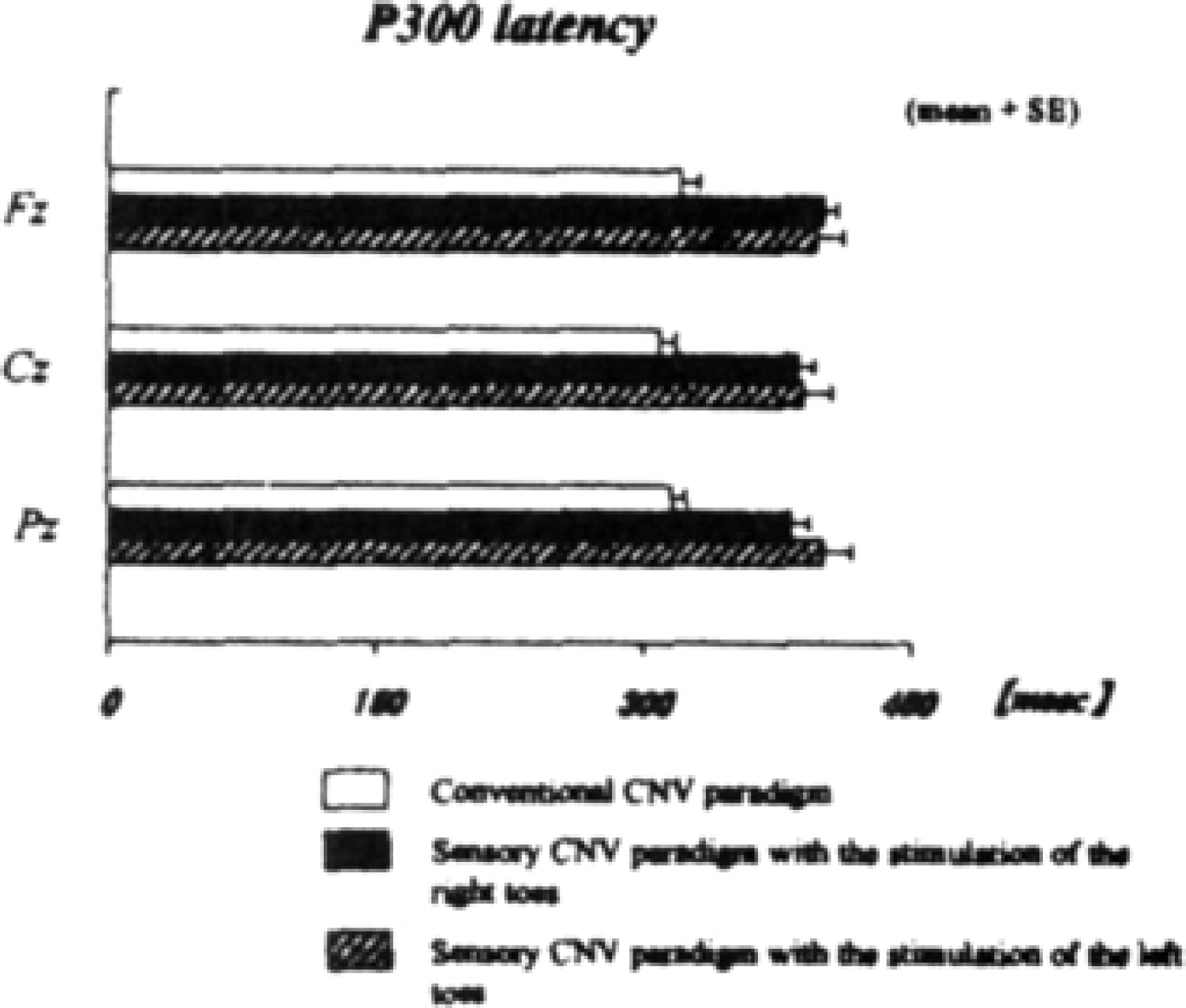
Differences in P300 latency between the conventional CNV paradigm (white bar) and the somatosensory paradigm with the right-toes stimulation (black bar), and between the conventional paradigm and those with left-toes stimulation (slashed bar), respectively.
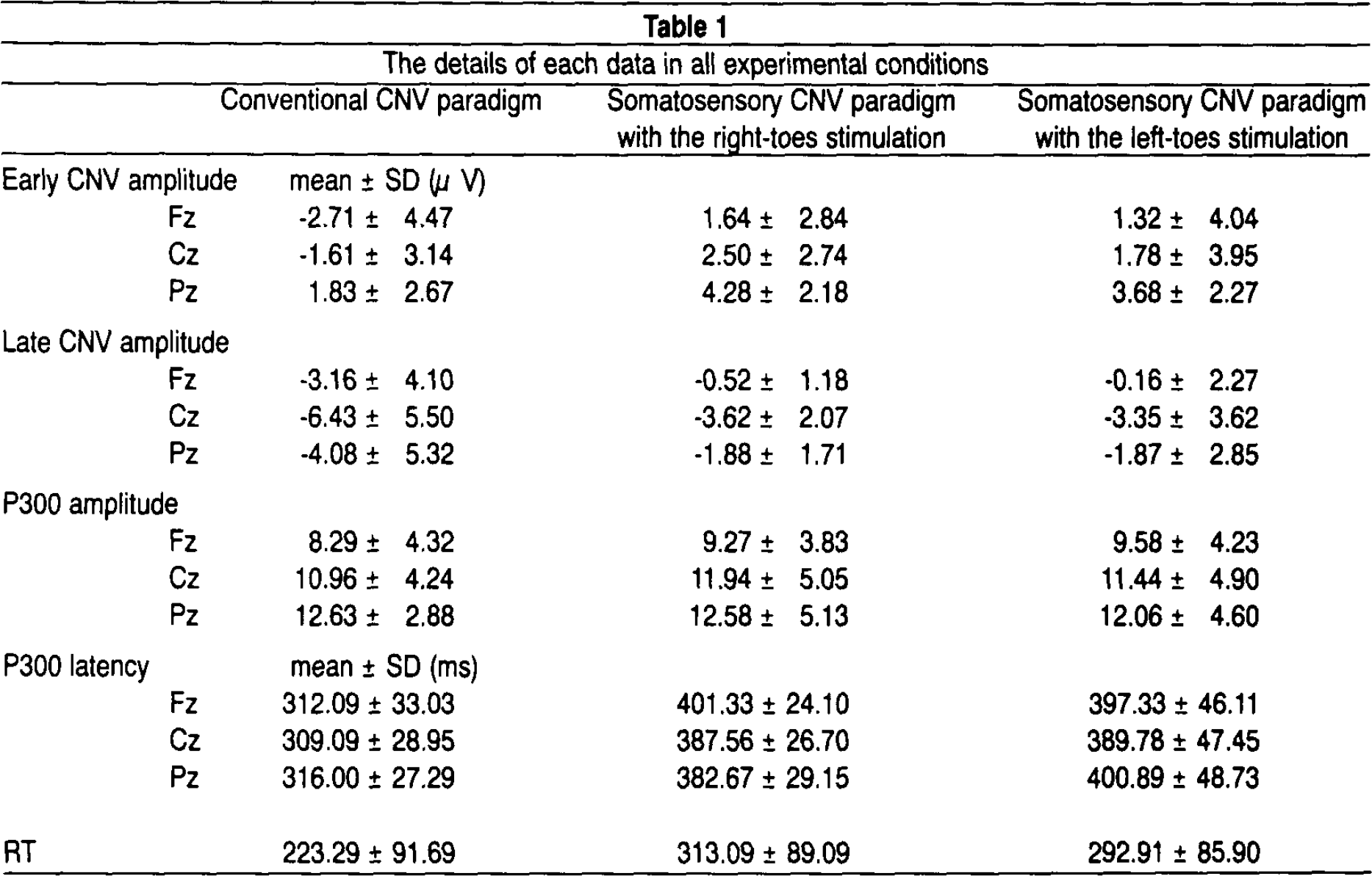
Both P300 and late CNV were not significantly different in amplitude between each paradigm. Furthermore, differences in each ERP (P300, early CNV and late CNV) in the somatosensory CNV paradigm between stimulation of the right toes and stimulation of the left toes were not significant. RT in the somatosensory CNV paradigm in both conditions of right and left toes stimulation appeared to be more prolonged than in the conventional CNV paradigm, but there was not a significant difference in RT between these groups. The details of each data in all experimental conditions are presented in Table 1.
The details of each data in all experimental conditions
DISCUSSION
Only a few studies have focused on the effect of somatosensory stimuli on each CNV component (early and late CNV) to date. Some previous studies have reported the modulation of somatosensory evoked potentials during the foreperiod of a forewarned RT task. 4,6 However, they did not employ somatosensory stimuli as WS and IS, therefore, the CNV evoked by a somatosensory stimulus was not examined in their studies.
In addition, the hemispheric asymmetries of each CNV are still unclear. Several studies have shown symmetries of the CNV, 7,8 whereas others have shown task-dependent 9 or-independent asymmetries. 10 Although Lutzenberger et al. 11 reported the hemisphere-specific processing of tactile tasks, they did not compare their findings statistically with data depending on other stimulus modalities.
On the other hand, Desmedt et al. 3,12 suggested that the receiving somatosensory cortical areas are situated on the brain surface, whereas the auditory or visual receiving areas are more deeply located. Therefore, the early components of somatosensory event-related potentials (ERPs) can be easily recorded from the scalp electrodes in comparison with other modalities. The somatosensory stimulus is inputted into the hemisphere contralateral to the stimulated toe and performing finger. These facts suggest that, a somatosensory CNV paradigm, with paired electrical stimuli as the WS and IS, is suitable for the investigation of hemispheric asymmetry of the CNV and of the influence of the sensory modulation on the early stage of information processing including the early detection, cognition and the orienting response. 2
In the present study, the early CNV was smaller in amplitude and P300 was longer in latency in the somatosensory CNV paradigm in comparison with the conventional CNV paradigm. The term “P350” used by Desmedt and Debecker, 13 instead of P300, is associated with the cognition of stimulus and considered as the time of closure of the stimulus evaluation process in the somatosensory task. The early CNV has been considered to be a correlate of the orienting response. 2 Thus, the initiation of cognition and orienting response is delayed in the somatosensory CNV paradigm.
Furthermore, the latency of P100 evoked by a somatosensory stimulus (peak latency at Fz: approximately 292 msec in the condition with stimulation of the right toes, 296 msec in those with stimulation of the left toes) was longer than that of N100 elicited by an auditory stimulus (peak latency at Fz: approximately 282 msec). P100 may index the classification and the identification of input stimuli. 3 Therefore, the initiation of early detection as reflected by P100 is delayed. This delay may be due to the longer transmission pathway from the toe to the cortex for the SEP signal than for the auditory signal. Similarly, this longer pathway can possibly explain why the P300 response is longer for the somatosensory signal than for the auditory signal.
There was no significant difference in each ERP (P300, early CNV and late CNV) on the sensory CNV paradigm between both hemispheres. This finding indicates that even a somatosensory input to the contralateral hemisphere was dealt with symmetrically, similar to other sensory inputs.
In conclusion, the present study revealed that a change in sensory modality in the CNV paradigm modified the early CNV component, yet did not produce the difference in the information processing of somatosensory stimulus between the hemispheres.
Footnotes
ACKNOWLEDGMENT
This study was supported by a Grant-in-Aid (09670972) for Scientific Research from the Japanese Ministry of Education, Science and Culture.