
Abstract
The purpose of the study was to examine the P3a and P3b components of the event-related brain potential (ERP) in patients sustaining moderate to severe brain injury. Electrophysiological and behavioral responses were recorded in brain injured (N = 18) and healthy control (N = 21) participants during performance of an auditory 3-stimulus distractor paradigm. Auditory stimuli consisted of a series of repetitive standard tones (75 ms), occasionally interrupted by equiprobable target (25 ms) and distractor sounds (white noise). Tone duration discrimination accuracy was similar in patients and controls, but patients had prolonged reaction times to targets. The reaction time delay was paralleled by a prolongation of P3b latency to targets in the patient group relative to controls. The stimulus and task dependent modulation of ERP responses in the brain injury group was similar to that of controls in terms of the spatial distribution of ERPs over the scalp. However, the brain injury group had attenuated P3a and P3b amplitudes to distractor and target stimuli, respectively. The electrophysiological data suggest a deficit in the allocation of attentional resources to the processing of deviant stimuli in the brain injury group.
INTRODUCTION
The P3 event-related brain potential (ERP) component as seen in the oddball paradigm is not an unitary response and there is agreement on the existence of at least two classes of late positive potentials. Predictable and task-relevant targets typically generate small prefrontal P3s, and large parietal maximum P3s. However, separation of the two components tends to be difficult in the 2-stimulus paradigm because they often overlap in time within the same ERP waveform. The various subtypes of P3 are more easily dissociated in a modified oddball paradigm when a second type of deviant stimulus (distractor) is added to the conventional stimulus sequence of target and standard stimuli. Different types of distractors have been employed. One type of P3 (novelty P3) is observed when introducing unexpected, unique complex stimuli (e.g., environmental sounds) that are physically distinct from the target and standard stimuli. 1 Another procedure uses rare stimuli that differ from an equally rare target and common standard only along a single dimension, such as pitch. 2 A variant of this procedure uses highly salient (but not unique) non-target stimuli that are easily distinguished from targets and standards. This type of non-target is sometimes referred to as a “no-go” distractor, and often has a more centroparietal distribution than the novelty distractor. 3 The label P3a has been coined for the P3 evoked by unexpected “no-go” or novel stimuli in order to distinguish it from the P3b response elicited by targets. The P3a component typically has a shorter latency and a more anterior scalp distribution than the P3b and has been considered an index of frontal brain activity. 4
The P3a is often seen as a reflection of an early orienting response, but recent studies suggest that P3a amplitude is affected by the target-standard discrimination difficulty, and not by novel stimulus characteristics per se. 3,5,6 These results are in agreement with a recent review suggesting that the earlier mismatch negativity (MMN) reflects the initial detection of deviant events, whereas the P3a is associated with the evaluation of deviance for subsequent action. 7 The posterior maximum P3b reflects the allocation of attentional resources to stimulus evaluation 8 and memory processes involved in the storage of stimulus information. 9 Although there is no consensus on the functional significance of the P3, all models relate P3 to attention or working memory mechanisms. Because aspects of attention and memory are often affected in various brain disorders and injuries the P3 component has been used as an index of cognitive function in patients suffering from a range of neurologic and neuropsychiatric conditions.
P3a and P3b in Brain Injury
Most studies using oddball paradigms have demonstrated that patients with head injuries ranging from mild to severe have attenuated, and sometimes delayed, P3bs to target stimuli. 10,11 The possibility of differential effects of brain injury on various subtypes of P3s has received limited attention. Rugg et al. 12 examined a group of severely head-injured patients with an auditory novelty (environmental noises) oddball task. The target-elicited N2 and P3b did not differ between patients and controls, whereas the P3a elicited by the rare novel sounds had longer peak latency in the head injured group, suggesting that shifts of attention were delayed in severe head injury. A study by Kaipio et al. 13 showed enhanced P3a amplitudes to natural novel sounds in a group of severely head-injured patients compared to healthy controls. The data suggested that novel stimuli caused greater distraction in the head-injured group than in controls. Even studies of patients with mild head injury have reported significant ERP changes. Potter and colleagues 14 found no significant differences between a mild head injury group and controls in the P3b elicited by target stimuli or the P3a produced by novel stimuli. However, a greater negativity following the P3a in the mild head injury group suggested increased activation of components of the attention network. Solbakk et al. 15 used an auditory 3-stimulus task with target and distractor tones that differed along the dimension of tone frequency only. They reported that mild head injury patients and controls had comparable P3 latencies and scalp topographies, but the patient sample had impaired performance and attenuated P3 amplitudes to both classes of deviants relative to controls.
The few reports using distractor paradigms have yielded mixed results. This most likely relates to the use of paradigms with different types of distractors and patient groups differing in head injury severity and location. A series of studies conducted by Knight and coworkers 16 –20 on patients with circumscribed lesions due to unilateral cerebral infarctions have demonstrated that the P3a and P3b components are differentially sensitive to lesion location. The P3b to target stimuli is mainly affected by lesions in the temporoparietal junction and is insensitive to frontal or medial-temporal lesions. However, P3a to distractor stimuli is sensitive to frontal lesions as well as to hippocampal lesions, and to some extent to parietal lesions. Studies of patients with localized brain lesions converge with the intracranial as well as the scalp-recorded data in suggesting that the observed P3 response reflects the concerted activity of a distributed neural network, and the various elements of this network respond differentially to various stimulus and task parameters. Different functional aspects of this network may be disturbed in brain injured individuals, and the distinction between various subtypes of P3s may contribute to identify and characterize dysfunctional processes in these patients.
Present Study
A three-stimulus oddball paradigm consisting of target and standard tones of different duration, and highly salient non-target (distractor) stimuli, was used to obtain P3 ERPs in a group of patients with moderate to severe brain injury. The aim of the study was to examine whether P3a and P3b would be differentially affected by brain injury. It was hypothesized that patients would be relatively more distracted by perceptually salient non-target stimuli than healthy controls and that this might interfere with task performance. A previous study 21 using the present stimulus design with a large group of healthy participants has demonstrated that the paradigm is efficient for distinguishing the P3a and P3b components at the individual subject level. Because group studies mask individual differences, an illustrative case will be presented to show the effects of brain injury on P3 at the individual patient level.
METHODS AND MATERIALS
Participants
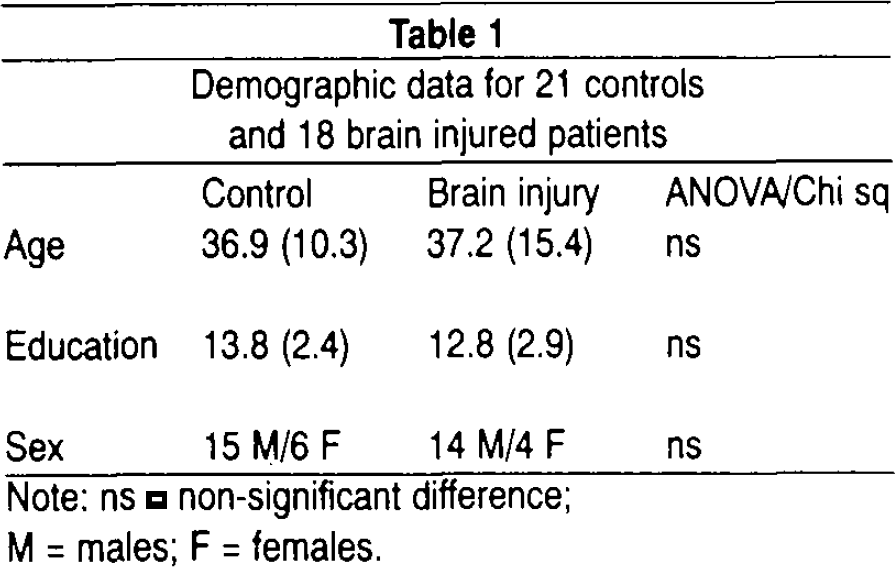
Table 1 summarizes subject characteristics. The brain injury group comprised 18 patients who had suffered moderate or severe brain damage due to traumatic head injury (N = 12) or cerebrovascular infarct (N = 6). The injury or disease was sustained at least 3 months before participating in the study. All patients had brain damage (mostly of moderate severity) confirmed by neuroradiological examination (CT/MRI). Ten patients had contusions or infarcts primarily located to frontal or frontotemporal brain regions. Neuropsychological reports concluded with mild-to-moderate cognitive dysfunction in all patients. Some of the patients had hemiparesis, but used their normal functioning hand for task performance. All patients were undergoing rehabilitation at the time of the study. The patients did not suffer from any known neurological or psychiatric disease prior to the injury.
Demographic data for 21 controls and 18 brain injured patients
Note: ns = non-significant difference;
M = males; F = females.
The healthy control group included 21 age and education matched subjects, with no history of substance abuse, psychiatric or neurological disease. The patient and control group did not differ significantly in age or years of education. All participants reported normal hearing.
Case Study
An 18 year-old right-handed female with 10 years of education participated in the investigation. She sustained a severe head injury due to a traffic accident 8 months prior to the study. She had a long comatose period (weeks), but gradually regained full consciousness. Initial neuroradiological examination showed a right-hemisphere epidural hematoma that was surgically removed, and left-hemisphere frontotemporal contusions. Recovery was complicated by heightened intracranial pressure and epileptic seizures in the post-acute phase. Neuropsychological testing conducted 8 months postinjury showed moderate impairment in most domains of cognitive functioning, but verbal abilities were largely preserved.
ERP Paradigm
A total of 360 tones were presented binaurally through calibrated headphones. Standards (72%) and targets (14%) were spectrally composite tones (80 dB) with a 500 Hz fundamental frequency and harmonics at 1000 Hz and 1500 Hz. Standard and target tones differed in duration in that standards had 75 ms duration and targets 25 ms duration. A low-probability (14%) distractor sound of 100 ms duration, 95 dB spectrally filtered white noise (24 dB/octave; in the interval < 250 and > 3000 Hz) was added to the stimulus sequence. All stimuli were presented with a 5 ms rise/fall time. The stimuli were delivered in a quasi-random order (fixed SOA = 1000 ms) to ensure that a target was never followed by another target, and that no target occurred among the 10 initial stimuli. Subjects were instructed to give an immediate button press when detecting the target tone, and to refrain from responding to all other stimuli. Button press and response latency from stimulus onset was registered. The task was preceded by a practice session to ensure that the participant could differentiate targets from standards, and make the required button press to targets. The choice of stimulus characteristics was influenced by a study by Tervaniemi et al. 22 aimed at identifying the auditory stimulus parameters (frequency, intensity, duration) that most reliably elicit early sensory and discriminatory processes preceding the P3 component. They concluded that a duration difference with a 75 ms duration deviant and a 25 ms duration target was optimal in terms of test-retest reliability.
EEG/ERP Recording
Continuous electroencephalographic (EEG) activity was sampled at a digitization rate of 500 Hz. The signals were amplified with a SynAmps system (bandpass 0.1–30 Hz). Neuroscan Inc. (Herndon, VA) software was used to present stimuli, and to register and analyze the EEG activity. Nineteen monopolar electrodes (Ag/AgCl) were manually placed according to the International 10–20 system at frontal (Fp1, Fp2, F3, F4), central (C3, C4), temporal (T3, T4), parietal (P3, P4, P7, P8), occipital (O1, O2), and midline (Fz, Cz, Pz) sites, and referenced to a left mastoid electrode. A ground electrode was attached to the forehead. Vertical and horizontal eye movement artifacts were recorded with bipolar leads at the supraorbital ridge and suborbital region of the left eye, and at the outer canthi of each eye. Electrode impedance was kept below 5 kΩ. EEG epochs were digitally bandpass filtered (0.1–15 Hz; 24 dB/octave), linearly detrended and baseline corrected (−100 to 0 ms). Statistical eye movement correction as described by Semlitsch et al. 23 was performed on single sweeps prior to averaging. Epochs with amplitudes exceeding ± 100 μV were rejected. Single sweeps from 100 ms pre- to 1000 ms post-stimulus were averaged for both classes of deviant tones.
ERP Data Analysis
Identification of peaks and computation of mean amplitudes from averaged ERP waveforms was performed with computer software. 24 P3b latency for the target tone was quantified as the time point of maximum amplitude on three midline sites (Fz Cz, Pz) in the window 250–600 ms post-stimulus. P3a latency for distractors was identified as the time point of maximum amplitude on the same electrodes within a 230–320 ms window. All P3 amplitude measures were locked to the time point of maximum amplitude at Pz for targets and Cz for distractor stimuli. The P3 peak elicited by target tones could not be accurately identified in some of the individual ERP waveforms, and sometimes overlapped with an earlier positivity in the P2/P3a time range. Thus, in addition to P3 peak analysis, mean amplitudes in successive time intervals corresponding to P3a and P3b were calculated. P3b for the target was represented by average amplitude in three time windows (Interval 1: 300–400 ms; Interval 2: 400–500 ms; and Interval 3: 500–600 ms), and P3a for the distractor was represented by mean amplitude in one time interval (200–300 ms).
Statistical Analysis
SPSS 6.1.3 for Windows (SPSS Inc.) was used for all statistical analyses. Comparison of group differences in demographic and performance (hit rate, false alarm rate, and reaction time) data was calculated using one-way Analysis of Variance (ANOVA) with group as between-subjects factor.
Repeated-measures ANOVA for mixed designs was used to test differences in ERP magnitude, latency, and topographic distribution between stimulus types and between groups. The overall ANOVA was defined by two repeated measure factors for the analyses of amplitude (peak amplitude for both deviants and mean amplitude for the distractor) and latency data obtained at midline electrodes: stimulus type (target vs. distractor), electrode (Fz, Cz, Pz), and one between-subjects factor (group). The ANOVA for the analyses of mean P3b amplitude to targets included the additional within-subjects factor of measurement interval (3 levels). Supplementary analyses for lateral electrodes yielded no significant effects and will not be reported. Significant interaction effects of group and repeated measure factors were decomposed using additional ANOVAs. Alpha was set to .05, and Greenhouse-Geisser epsilon corrected p-values, along with uncorrected degrees of freedom, are reported whenever there were more than two levels of a repeated measure factor.
RESULTS
Behavioral Performance
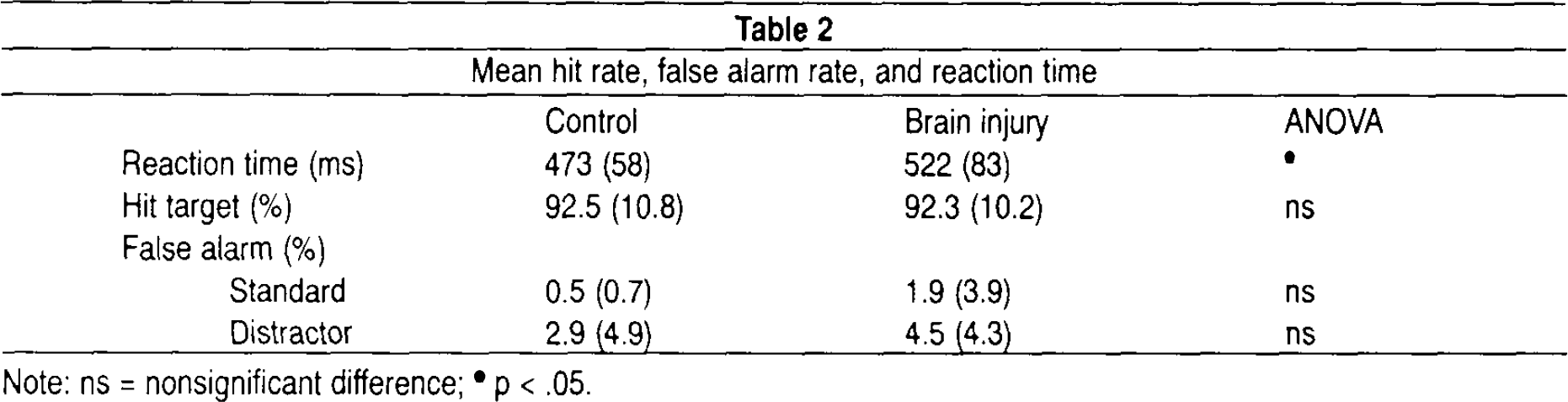
Mean reaction time, percent detected targets, and percent false alarms are reported in Table 2. The number of erroneous responses and false alarms to non-target stimuli was low in both groups, and yielded no significant group differences. Statistical analyses revealed significant group differences with regard to reaction time, F(1,37) = 4.96; p < .032. The brain injury group responded more slowly to targets compared to controls.
Mean hit rate, false alarm rate, and reaction time
Note: ns = nonsignificant difference;
p < .05.
Event-Related Potentials to Target and Distractor Stimuli
Figures 1 and 2 illustrate grand-average ERP waveforms for target and distractor tones, respectively. Mean ERP values are summarized in Table 3. P3b parameters to target tones were missing in 1 control subject and 1 patient due to lack of a defineable peak.
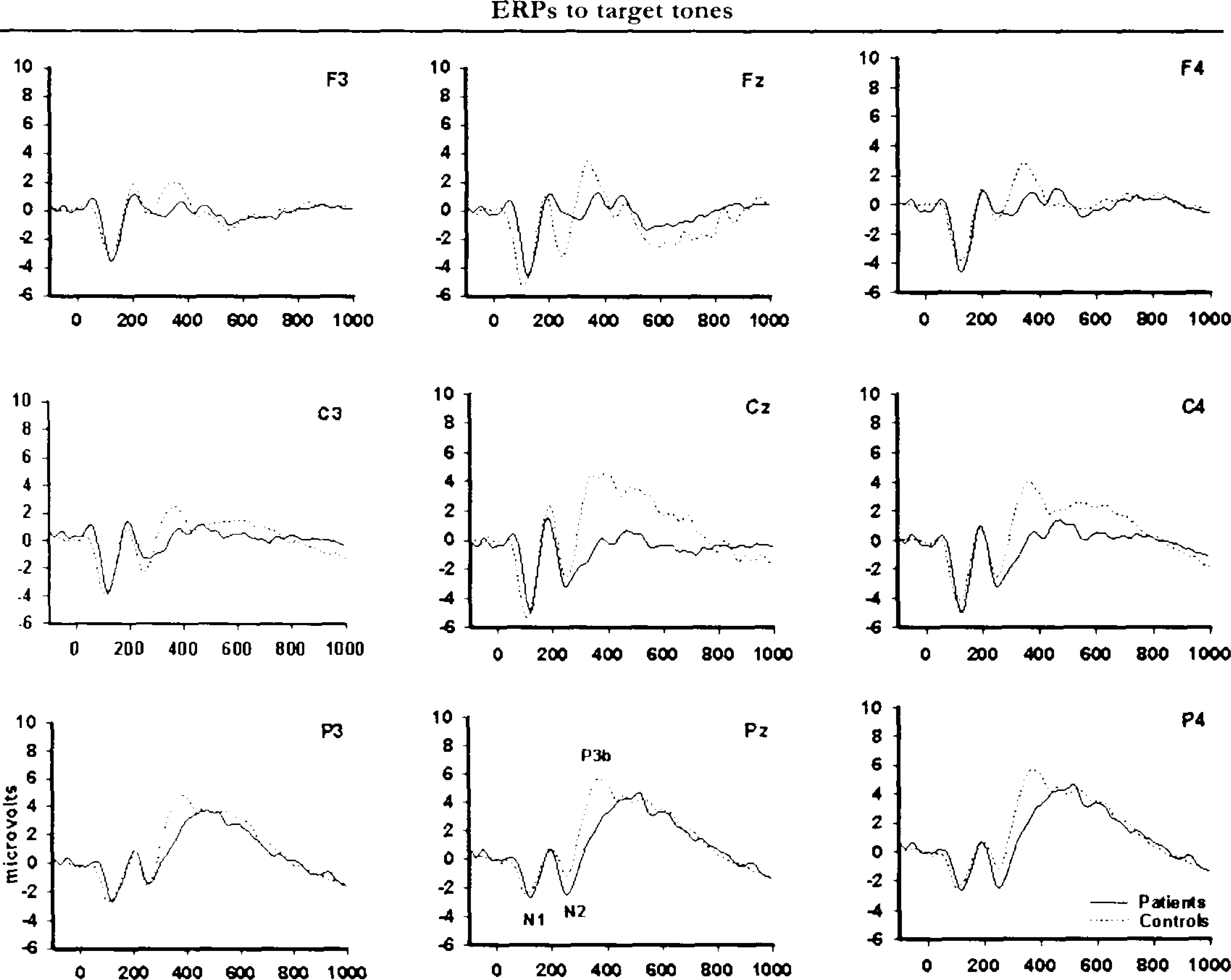
Grand-average ERPs for target tones. Waveforms are depicted for the control group (solid line), and the brain injury group (dotted line). Positive polarity is up.
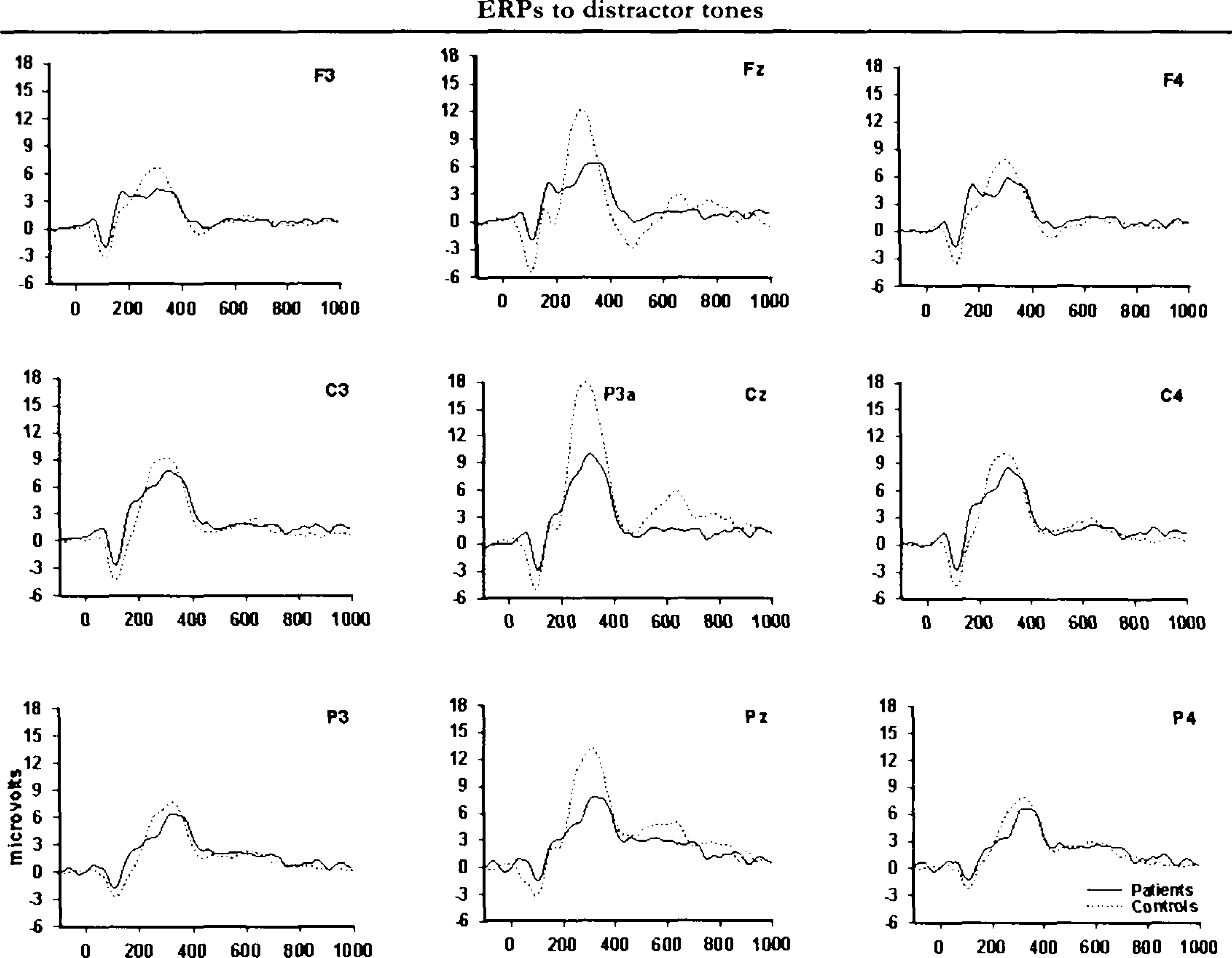
Grand-average ERPs for distractor stimuli. The brain injury group average is indicated as dotted line and the control group average is indicated as solid line. Positive polarity is up.
Mean (SD) ERP values for the target and distractor stimuli at midline electrodes
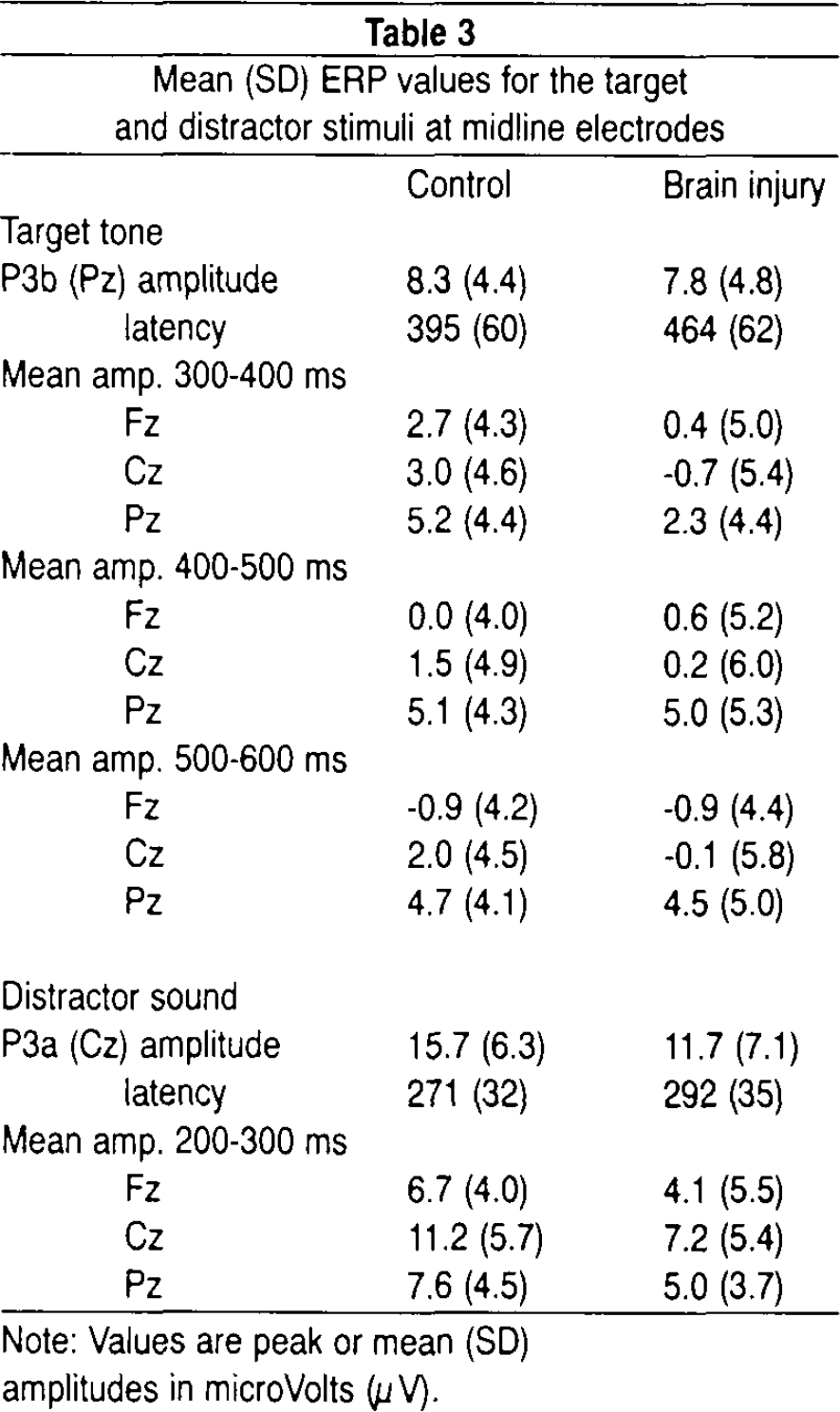
Note: Values are peak or mean (SD)
amplitudes in microVolts (μV).
Figures 1 and 2 indicate that the scalp distributions of the target and distractor stimulus P3s were not identical. The ERPs elicited by target tones had a prominent posterior maximum, whereas ERPs to distractor sounds had a central maximum and were more evenly distributed over frontal and parietal sites. A significant interaction of stimulus type and electrode, F(2,70) = 55.76; p < .001, ε = 0.86, in the overall analysis confirmed that target and distractor tones had distinct spatial distributions over the scalp. The group factor did not interact with the stimulus type differences in scalp topography. The expected gradual increase in P3b amplitude to targets in frontoparietal direction was present in all groups as well as a central maximum P3a for distractor stimuli.
Although the initial analysis did not reveal scalp topography differences between the groups, visual inspection of grand-average waveforms in Figures 1 and 2 suggests that brain-injured patients had a general lowering of P3s to both categories of deviant stimuli compared to controls. This impression was partially confirmed by statistical analyses. The peak-derived amplitude data showed no statistically significant group differences. However, analyses of mean amplitudes in defined time intervals (covering the P3a and P3b components, respectively) confirmed a significant interaction of group and interval, F(2,74) = 5.31; P<.009, for the target stimulus P3b. Post hoc analyses showed that the brain injury group had attenuated amplitudes compared to controls in the early portion (300–400ms) of the P3b complex, F(1,37) = 4.69; P<.037. The analyses also confirmed that the control group had more pronounced P3a to distractor sounds relative to the patient group, F(1,37) = 4.73; P<.036.
The overall analysis of peak latencies showed a significant main effect of Group, F(1,27) = 10.47; p < .003, on P3 latency that was modified by Group × Stimulus type interaction, F(1,27) = 5.50; p < .027. Follow-up ANOVAs showed that the brain injury group had longer P3b latency to targets relative to controls, F(1,27) = 11.73; p < .002. There was a small group difference in P3a latency that approached significance (p < .08).
Case Study
The patient with extensive left hemisphere damage due to a traumatic head injury correctly responded to 64% of the targets and had a 10% false alarm rate to distractor sounds. Inspection of the continuous EEG data with event and response markers showed that the low target detection rate was partly due to delayed responses occurring shortly after the onset of the subsequent standard stimulus. A general response slowing was also indicated by the prolonged reaction time (approximately 700 ms) to targets responded to within the allowed response window. Thus, the behavioral data show that the patient followed task instructions and correctly detected a reasonable number of targets, although reaction time was clearly delayed.
Visual inspection of the stimulus averaged ERP waveforms (see Figure 3) suggests that the distractor elicited a P3a of comparable latency as that of the control group, but with amplitude of a similar magnitude as the patient group at midline electrode sites. The most striking finding, however, is related to the topographic distribution of the P3a over the skull. Whereas the electrodes positioned at right hemisphere lateral sites show rather large amplitude P3as, the P3a at certain left hemisphere lateral frontotemporal locations (particularly F7 and T3) is virtually abolished. The topographic location of this effect corresponds with her left hemisphere brain lesion. The data also reveal an abnormality in the P3b response to target stimuli. A P3b component is hard to identify, but the late positivity observed at centroparietal sites may represent the ascending slope of a delayed P3b component. This is suggested by the scalp distribution of the ERP waveform as well as the reaction time data. Altogether, the ERP results indicate a general disturbance in target processing, and relatively more topography specific changes in distractor processing.

Average ERPs for target (dotted line) versus distractor (solid line) stimuli at selected electrode locations in a patient with severe left hemisphere damage.
DISCUSSION
The present study employed an auditory 3-stimulus distractor task to examine the P3a and P3b components in patients with moderate to severe brain injury and healthy controls. The results of the behavioral measures show that the brain injury group performed at a similar level as healthy controls with regard to response accuracy and false alarm rate on this relatively difficult tone duration discrimination task. Although discrimination accuracy was comparable to that of the control group, patients had prolonged reaction time to target tones. The behavioral response slowing was accompanied by a prolongation of P3b latency to targets, suggesting some delay in both stimulus evaluation and response production. P3b latency was delayed by approximately 70 ms, and reaction time by 50 ms compared with the control group.
In addition to the neurophysiological and behavioral indications of delayed responses to targets, the patient group had a significant amplitude attenuation of the early portion of the P3b complex, corresponding to the peak of P3b in the control group. Diminished P3b amplitude is a common finding in head injury of varying degrees of severity, whereas P3b latency prolongation is more frequently observed in severe head injury. 10 Reduced P3b amplitude may be related to loss of information due to inattention in conditions with stimuli that are difficult to discriminate. The present task is quite demanding in terms of stimulus discriminability. ERP paradigms requiring discrimination of tone duration are judged to be more difficult than pitch discrimination tasks as there can be no echoic replay of the stimuli in the former task. 25 Because this type of task requires vigilant attention for optimal performance, the P3 abnormality may index a deficit in focused sustained attention.
Interference from non-target information may have contributed to deficient target processing. However, the data do not lend support to this interpretation. In contrast, the reduced P3a amplitude to perceptually salient distractors observed in the grand-average waveforms of patients compared with controls suggests decreased processing of irrelevant sounds rather than greater distraction. Moreover, the lack of excessive false alarms to non-target stimuli provides behavioral support for the notion that interference from non-target information is not a likely explanation for the processing deficit observed in the brain injury group. In contrast to the target tone, the distractor deviates extensively from the immediate stimulus context and does not require vigilant attention for evoking a response. A distractor sharing elements of the physical features of the target and standard stimuli may have produced more interference with task performance than a distractor differing greatly from the task-relevant stimuli. 26 The results are in contrast with those of Kaipio et al., 13 showing enhanced P3a amplitudes to natural novel sounds in a group of patients sustaining severe closed head injury. However, Knight and coworkers 16,18 reported that dorsolateral prefrontal cortex lesions reduce the P3a component elicited by unexpected novel sounds.
Altogether, the behavioral and electrophysiological data suggest that the brain injury group suffer from a deficit in the allocation of attentional resources to task-relevant stimuli. A deficit due to information loss or a reduction in the level of attention is also suggested by the scalp topography data. The lack of significant group differences in the spatial distribution of ERPs suggests that the attributed stimulus significance did not vary between groups in cognitive terms, and that the neural generators mediating the ERP effects appeared to be similar in patients and controls. 27,28
Decreased availability and/or mobilization of attentional resources would be expected to compromise motor responses, leading to prolonged reaction times as seen in the brain injury group. The increased reaction time in the brain injury group might suggest that they compensated for a relative lack of attentional resources by engaging in extended stimulus evaluation. Although earlier ERP components such as N1, P2, and N2 were not measured in the present study, inspection of the grand-average ERP waveforms do not suggest a general slowing of information processing in the brain injury group. Reaction time studies usually indicate a general slowing of cognitive processes in traumatic brain injury, but studies using both behavioral and ERP data to measure processing speed have suggested that brain injury induces differential slowing of various stages of information processing. 29
Although brain injury did not have a differential effect on P3a and P3b in the present rather heterogeneous patient group, studies of Alzheimer's type dementia have reported differential sensitivity of the P3a and P3b components. 30–31 It is of course difficult to predict specific attention effects in a group of patients sustaining multiple focal injuries and/or diffuse injuries. In closed head injury it is common that several areas are simultaneously affected and that both localized and diffuse axonal injury occurs. 32 This was also the case in the present brain injury sample. Therefore, the aggregated results of the brain injury group may in fact include subgroups with different types of attention and memory deficits. A case was presented showing the effect of lateralized brain contusion on ERPs. In a patient with severe left hemisphere damage to the frontal and temporal lobes, P3a responses to distractor sounds were virtually abolished at left hemisphere lateral frontotemporal recording sites. The topographic specificity of the ERP effect corresponded well with lesion location evidenced on neuroradiological examination.
CONCLUSION
The brain injury group was sensitive to the task demands and to the cognitive significance of stimuli in this 3-stimulus distractor task. However, brain injury interfered with the speed and magnitude of electrophysiological and behavioral reactions to task-relevant target stimuli. Specifically, the patient group had reduced P3b amplitude and increased latency relative to a healthy control group. The ERP deficit in turn had adverse consequences for reaction time performance. Although P3a amplitude was diminished in the patient group, the results indicate that basic orienting and evaluation of unexpected and perceptually salient stimuli was largely preserved. The P3 amplitude data favor a resource limitation rather than a distraction interpretation of attentional dysfunction in this brain injury group. The addition of on-line physiological processing measures to the neuropsychological examination may contribute to identifying subtypes of attention deficits in individual patients as illustrated with the case report. ERPs clearly offer more precision than behavioral measures in the process of making a functional diagnosis. The development of standardized test procedures and normative studies is expected to further enhance the clinical utility of cognitive ERPs.
Footnotes
ACKNOWLEDGMENT
This research was funded by the Research Council of Norway.
We thank Dr. Magnus Lindgren for helpful comments on an earlier version of this article.