
Abstract
Monoaminergic challenge tests allow investigating central nervous changes in humans under acute depletion of specific neurotransmitters (5-HT, DA, NE). Along with studies using alpha-methyl-para-tyrosine test (AMPT) and phenylalanine/tyrosine depletion test (APTD), the tryptophan depletion test (ATDT) represents the currently most established human challenge tool for the assessment of brain serotonin functioning. Neurophysiological studies in healthy and clinical samples may contribute to the search for a non-invasive and reliable biological marker of monoaminergic vulnerability or dysfunction. In the design of these studies, various biochemical and methodological aspects have to be taken into account. This article focuses on electrophysiological methodology and results of monoamine depletion studies (i.e., electroencephalography, magnetoencephalography, polysomnography, auditory evoked potentials and startle response).
Keywords
INTRODUCTION
In biophysiological research of neurotransmission, important progress was achieved by the change from the categorical view of a decrease of monoamine being characteristic of specific diseases towards the dimensional perspective that monoamine dysfunction represents a manifestation of, e.g., a vulnerability to mood lability or cognitive dysfunction, in a spectrum of disorders. 1 Supposedly, monoamine dysfunction rather reflects the presence of a psychobiological risk factor, like a genetic predisposition, an environmental factor or commonly interplays of these. 2 Monoamine depletion studies enable exploration of vulnerability models, which are hypothesized for serotonergic (5-HT), norepinephric (NE) and dopaminergic (DA) neurotransmission. Current attempts to better quantify manipulation of monoamine neurotransmitters on mood, behavior and cognition (Figure 1), seek noninvasive parameters of the CNS, which are neurophysiologically based like electroencephalographic techniques.
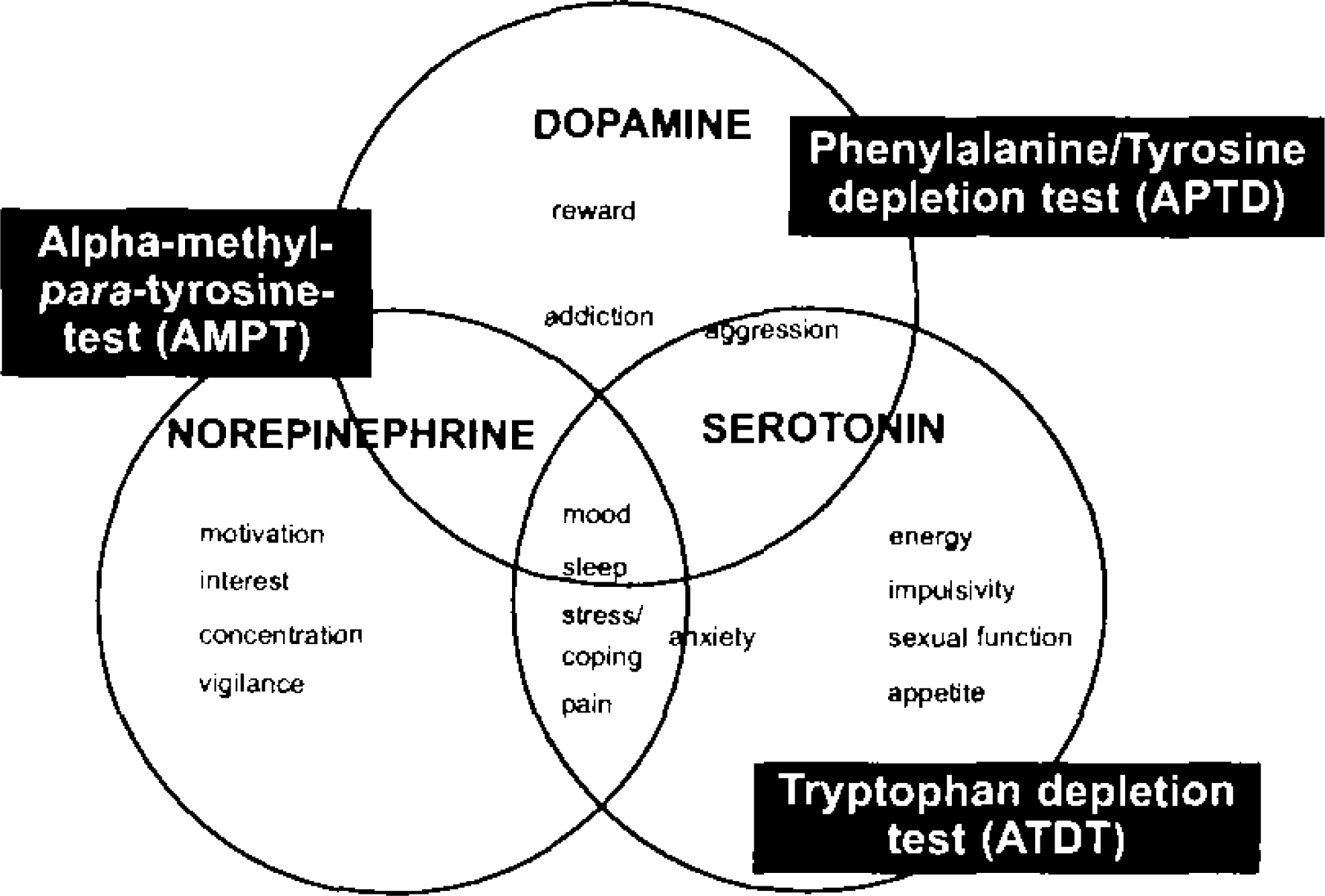
Dimensions of monoaminergic vulnerability. Modified from Healy and McMonagle 1997. 88
TRYPTOPHAN DEPLETION TEST (ATDT)
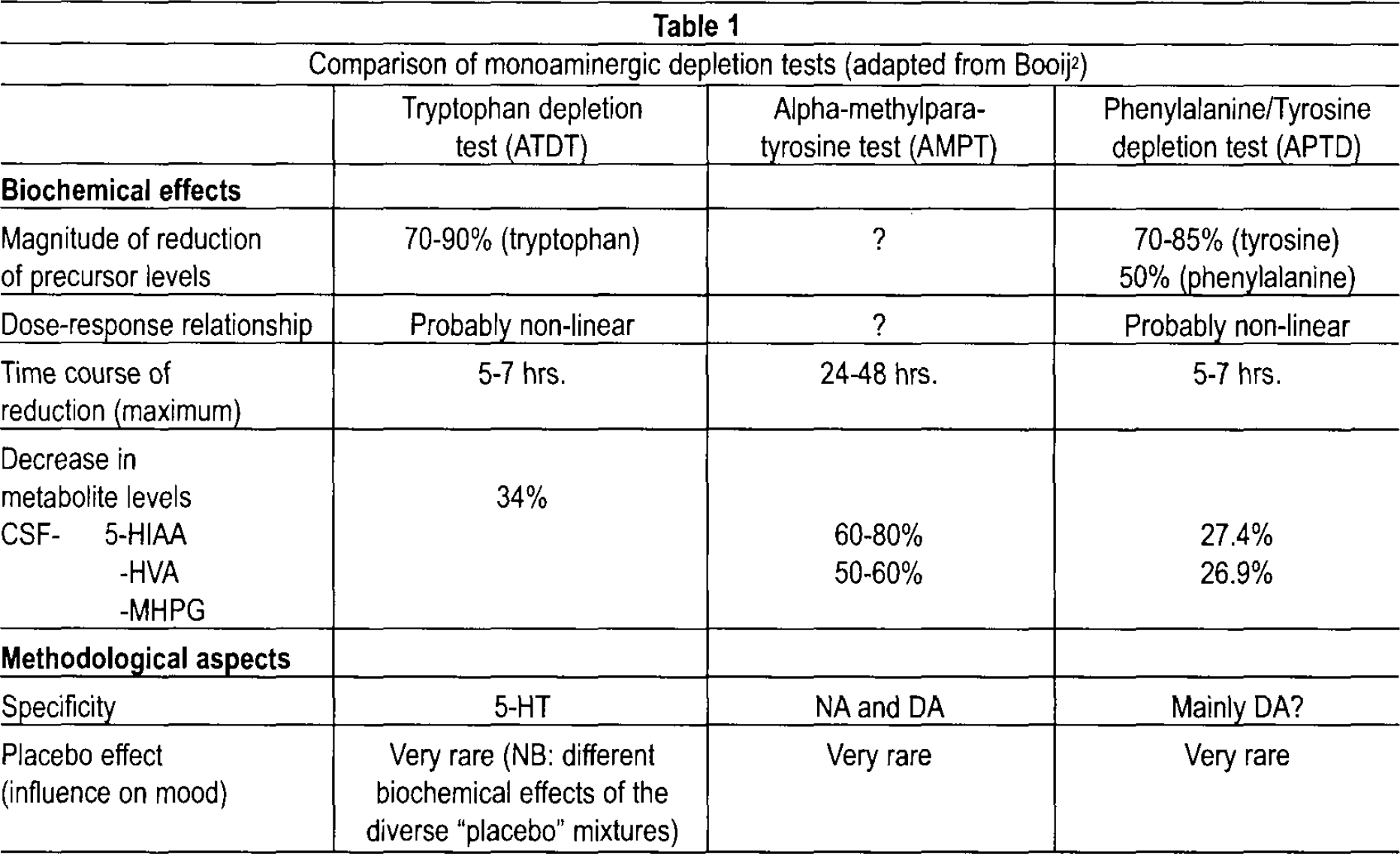
Following early animal studies 3 the tryptophan depletion test (ATDT) has become an established human challenge tool of the 5-HT system 4 for temporary reduction of tryptophan levels and central nervous 5-HT (Table 1). The method involves a highly concentrated and balanced amino-acid-drink with or without tryptophan, parallel to dietary regimens. Ingestion of 50–100 g of amino acid mixture without tryptophan leads to a transient decrease in plasma tryptophan of 70–90% within 4–6 hours when tryptophan from the human body itself is compensatory and more rapidly metabolized. As a precursor of 5-HT, tryptophan regulates with its free fraction — after having passed the blood-brain barrier in competition with other large amino acids — to synthesize 5-HT. 5,6 Levels of tryptophan in plasma and cerebrospinal fluid (CSF) are highly correlated. 7 Post mortem animal studies of ATDT showed a reduction of 50% of tryptophan in mediobasal hypothalamus, hippocampus and basolateral amygdala. 8 Single photon-(SPECT) as well as positron-emission tomography (PET), revealed reduced metabolism of central nervous 5-HT after ATDT in depressed patients. 9 In healthy volunteers the high 5-HT turnover was reduced markedly in the temporal cortex and putamen with a greater decrease seen in females. 10 Nowadays, this method has been widely explored clinically and is known to induce transient mood changes in vulnerable or previously depressed subjects, to reverse antidepressant effects, to be sensitive to predictors such as suicidality, recurrence or chronicity of depressive episodes, and female gender. 2,9,11
Comparison of monoaminergic depletion tests (adapted from Booij 2 )
EEG / Polysomnography
Electroencephalographic (EEG) recordings may be altered in response to acute challenge tests such as psychoactive or psychopharmacological substances like antidepressants. Knott et al. 12 found in quantitative EEG (QEEG) after ATDT a widespread increase of slow wave activity (delta amplitude), along with non-significant, highly variable theta increments and beta reductions. In this study, a fenfluramine-enhanced frontal EEG-pattern of beta activity was only detected in ATDT subjects. However, other studies 13 –15 did not find any EEG frequency difference between treatment conditions in ATDT. Still, these were pooled data of background segmented EEG preceding the delivery of acoustic stimuli (see below) as opposed to 5 minute resting EEG. 12 In the latter, our group found an increase in power in theta and alpha but decrease in beta bands in healthy subjects after ATDT (Norra et al. 47 ). QEEG investigations obviously need further confirmatory studies of that sort.
While slow wave activity is known to be associated to reduction in arousal and vigilance, two early polysomnographic (PSG) studies of ATDT were performed with normal sleepers, based on the model of monoaminergic/cholinergic regulation of NREM/REM sleep cycles. Bhatti et al. 16 reported decreased rapid eye movement (REM) latency in 11 healthy males, while a “control” drink containing only 25% of the ATDT experimental drink increased REM (time, %) compared with baseline. At the same time, Voderholzer et al. 17 demonstrated that ATDT led to decreased non-rapid eye movement (NREM) stage 2 sleep, and an increase of awake percentage and REM density.
The stimulating impact of ATDT on REM sleep underlined the 5-HT vulnerability in patients after remission from depression: Remitted depressives on SSRI maintenance treatment, showed a dose-dependent reduction of sleep latency and REM latency as well as an increase in REM (%, time, density) and total sleep time. 18 Remitted depressives on non-serotonergic antidepressive treatment, i.e., bupropion or cognitive behavior therapy, revealed a reduced NREM stage 2 (%) and pronounced REM activity (decrease in latency, increase in time and %). 19,20 Finally, in depressive patients ATDT reversed suppression of REM by phenelzine, a monoamino oxidase inhibitor. 21 Interestingly, no ATDT-induced mood changes in patients recovered from depression were observed in any of these studies. The only PSG study investigating effects of ATDT in non-depressed patients was performed with insomniacs, demonstrating an enhanced REM density and NREM stage 1 after ATDT, whereas NREM stage 2 was reduced. 22
In summary, sleep may serve as a sensitive neurophysiological indicator of monoaminergic functioning of CNS based on endogenous serotonergic challenge during regulation of sleep states.
Auditory evoked potentials (AEP) and magnetoencephalographic fields (MAEF)
In search of a biological marker for central 5-HT activity, characteristics like the high 5-HT innervation of the primary auditory cortex and hippocampus 23 as well as the 5-HT properties of tonic modulation of sensory thalamocortical processing 24 contributed to preferential investigations of AEP. Here, the concept of loudness dependence (LD) of AEP 25 gained major interest.
Neurophysiological effects of 5-HT depletion on auditory information processing were demonstrated in 1991 with an invasive animal model showing an enhancement of the N100 amplitude in hippocampal and cortical areas as opposed to lack of effects following dopamine (DA) depletion. 26 The negative correlation between the level of 5-hydroxyindolacetic acid (5-HIAA) in CSF and intensity dependence of sensory evoked potentials 27 was extended by investigating auditory stimulus intensity in animals, and also combined with manipulating 5-HT neurons in the raphe dorsalis. 28,29
For transfer of these findings to human studies, dipole source analysis of evoked potentials 30 has the advantage of noninvasively differentiating intracerebral sources for surface scalp-recorded waveforms. Two temporal sources were identified bilaterally for the auditory evoked N1/P2 component: a tangential dipole representing activity in the superior temporal plane where the primary auditory cortex is situated, and a radial dipole reflecting the evoked activity of the lateral superior temporal gyrus corresponding to secondary auditory areas. Hegerl and Juckel 25,29 postulated that augmentation of the N1/P2 in its tangential dipole activity with increasing stimulus intensity is associated with low 5-HT neurotransmission and vice versa. In challenge tests, source analysis is advantageous, because its test-retest-stability of auditory N1/P2 components is clearly superior to N1/P2 amplitudes of scalp electrodes.
Clinically, source analysis of loudness dependence auditory evoked potential (LDAEP) in patients with major depression 31 and other neuropsychiatric disorders like migraine, 32 alcohol 33 or chronic ecstasy abuse, 34 schizophrenia, 35 or borderline personality disorder, 36 thought to be characterized by 5-HT dysfunction, mostly indicated a strong LDAEP. Furthermore, pharmacological studies of enhanced stimulus intensity at baseline suggest a positive clinical response towards treatment with Selective-Serotonin-Reuptake-Inhibitors (SSRIs), lithium or other 5-HT agonists as opposed to clinical non-responders. 31,37 –42
However, the effect of central 5-HT manipulation by ATDT on LDAEP is not as prominent as expected by the paradigm. The hypothesis of an influence of 5-HT on the N1/P2 intensity dependence could not be confirmed by Dierks et al. 43 who applied AEP and dipole source analysis to a gender mixed group of 10 healthy subjects after ATDT, and he even measured some decrease of LDAEP on the right hemisphere. By analysis of scalp electrodes, Phillips et al. 44 investigated AEP stimulus intensity function in 13 males, followed by another study of 14 males by Massey et al., 45 both with negative results after ATDT. These findings were similar in a group of 18 females 46 with no differences in the central surface electrode sites. Applying source analysis to another healthy 16 female cohort, augmentation of LDAEP was more pronounced after ATDT, but findings were not significant from the control condition. 47 In a clinical sample of 14 bipolar patients, ATDT caused a decrease in the amplitude of N1/P2 (and P300) components of auditory ERP, 15 but the resulting LDAEP was not calculated. These patients were on lithium and did not show any mood changes; nevertheless, one might presume that the lithium treatment did not protect them from electrophysiological influences of ATDT, indicating some residing 5-HT trait vulnerability.
Furthermore, in different combined ERP paradigms under ATDT, no alterations in a three-stimulus “oddball” task regarding N1P2 and N2P3 (P300) components as well as contingent negative variation (CNV) magnitudes were found with healthy males, 13 nor any topographic ERP effects in episodic memory recall tasks. 48
Yet, the investigation with magnetoencephalography (MEG) showed significant influences of ATDT on the intensity of auditory stimulation, which is interesting because MEG appears to be specifically sensitive to tangential cortical activity. Kahkonen et al. 49 showed in 10 subjects that LD of N1/P2 magnetic dipole in the primary auditory cortex was decreased by ATDT at the contralateral hemisphere; furthermore, the highest stimulus intensity at 80 dB was detected to be stronger in N1m/P2m after control amino acid drink. They did not find any differences in concomitant ERP measurements. Although the authors conceded that the tangential component of N1m/P2m source in MEG may be different from the N1/P2 in EEG, their results were indeed contrary to the hypothesis of an increase of LD after ATDT and will have to be seen in some accordance to the previous EEG findings of Dierks et al. 43 They also speculate that prefrontal 5-HT neurons might influence inhibition of contralaterally presented acoustic stimuli.
These mostly negative ATDT findings for LDAEP as a biological indicator of central 5-HT activity will have to be put into perspective with respect to methodological differences regarding the amount of amino acid drinks and ATDT procedures, EEG localizing methods and analysis, and differences in subjects regarding gender, mood states, and personality traits assumed to be possibly linked to the 5-HT system and LDAEP like sensation seeking 50 or impulsivity. 36 However, despite strong depletion levels in all studies, a lowered 5-HT neurotransmission may contribute only to a limited extent to AEP, while interactions with other neurotransmitters like dopamine 51,52 have to be considered.
The debate is ongoing whether stimulation by selective 5-HT agonists may be a more specific challenge 53 ; preliminary results on the effect of single application of SSRI on LDAEP are diverse, 54,55 while longer treatment-related changes 31,35,39,41,42 seem to be replicated. Further exploration of LDAEP will show whether it in fact represents a brain electrical state or trait marker of 5-HT functioning.
Modulation of middle latency evoked potentials by ATDT was further investigated by combinations of EEG and its MEG counterpart. ATDT did not alter source dipoles (Pam, Nbm) of MAEF in a passive monaural auditory click tones (70dB) paradigm; it reduced Pam latencies and only tended to reduce Pam amplitudes. 56 Of note, interactions between gender (14 subjects, 7 females) with mixture and hemisphere were detected on the Nbm amplitude.
Manipulation of 5-HT was also investigated in the earliest “pre-attentive” auditory processing of mismatch negativity (MMN), and dipole modelling of MMN suggests that the assumed magnetic equivalent of supratemporal MMN subcomponent is generated in temporal lobe near the primary auditory cortices. 57 In ERP, ATDT seems to reduce the peak amplitude of deviant-tone N2 wave (representing task-irrelevant sound changes) and overlapping N2b subcomponents (representing active detection of sound changes). 58 In MEG, which is not sensitive to N2b detection, MMN peak latency was significantly increased over both hemispheres ipsi- and contralaterally to the ear stimulated while there were no alterations in MMN wave amplitudes, MMN source locations, or the subsequent P3a response. In summary, ATDT might delay automatic change detection in the auditory cortex and thus impair involuntary attention-shifting. However, these results are partly contrasted by a follow-up MMN study with increase of amplitudes to duration and frequency changes and decreased ipsilateral latencies of magnetic MMN to frequency changes after ATDT. 59 ATDT also modulated N1m latencies and decreased P2m source activity while P50 remained unchanged.
Effects of 5-HT depletion on attention were electrophysiologically demonstrated in a dichotic listening paradigm 60 where ATDT suppressed amplitude enhancement of gating P50 and the first orienting response N1 to attended tones of the overlapping processing negativity (PN) specifically generated during “selective attention,” but not P2 and following components. In MEG only, the N1m source amplitude tended to be increased by selective attention. Accordingly, as the early component of PN may be generated near or within the primary auditory cortex, 57 selective attention effects originating functionally near to the auditory cortex may also be modulated by 5-HT.
Startle
In acoustic response paradigms, the startle response (ASR) and prepulse inhibition (PPI) are often used (see review Koch 61 ). So far, most studies concentrated on dopaminergic pathways since DA agents disrupt PPI, and deficits in this “sensorimotor gating” are of major clinical impact in schizophrenia research. 62 However, disruption of PPI can be induced by both 5-HT stimulation 63 and pharmacological 5-HT depletion, 64 and be prevented by pre-treatment with 5-HT agents. 65 Regarding ATDT, the only comparable animal model showed an amplification of ASR in rats being fed an L-tryptophane-free diet for 13 days, and after addition of L-tryptophane their startle magnitude returned to control levels. 66
Applying ATDT in 13 healthy males, Philips et al. 44 did not observe changes in ASR but suppression of PPI, whereas a study of 15 healthy females revealed a reduction of ASR after ATDT without any significant influence on PPI. 67 Reflecting on these results, differences in species, in their complex neuroanatomy and pharmacology of 5-HT receptors, in methods, in gender and hormonal status have to be discussed. Knowledge of 5-HT mechanisms in human startle modulation presently is incomplete: 5-HT agonists or releasers 68,69 increased ASR and PPI, with prior application of SSRI (citalopram) enhancing ASR and attenuating the increase on PPI. 70 Conversely, single dosages of SSRI (fluvoxamine) did not affect ASR or PPI in healthy humans at all, 14 only long-term SSRI treatment (sertraline) in patients with major depression augmented ASR. 71
Thus, the variety of the more recent results from human studies weaken the assumed predictive value of the 5-HT influence on ASR as well as PPI disruption based on animal studies, 63 and changes were rather assumed to indicate alterations of neurotransmission other than just lowered 5-HT or a chronic 5-HT deficiency condition.
ALPHA-METHYL-PARA-TYROSINE DEPLETION (AMPT)
Depletion of catecholamines (CA), i.e., norepinephrine (NE) and dopamine (DA), can be obtained by administration of AMPT, given on a daily basis (1–4 g/day). After start of AMPT, levels of the CA metabolites 3-methoxy-4-hydro-xyphenylglycol (MHPG) and homovanillic acid (HVA) in urine and CSF decrease more than 50% within 48–72 hours, and return to baseline another 3–4 days after withdrawal of AMPT 2,72 (Table 1).
In clinical samples, patients remitted from depression on norepinephrine reuptake inhibitors (NRI; but not SSRIs) experienced a return of symptoms after AMPT, which also led to decreased brain metabolism in orbitofrontal, dorsolateral prefrontal and thalamic regions. 73 Thus, in recent combined studies, mood responses of patients to AMPT or ATDT were shown to depend on the pharmacological properties of the administered antidepressants. 74 However, in remitted drug-free patients with seasonal affective disorder, effects of light therapy were reversed by both ATDT and AMPT. 75 Moreover, relapse of symptoms like hypomania in bipolar patients with lithium-induced long-term remission was seen 24–48 hours after the last active AMPT dose, 76 and dysphoric responses in drug-free schizophrenic patients after AMPT correlated with changes in striatal DA function as measured by DA(2) receptor binding in (123)IBZM-SPECT imaging. 77
Electrophysiological effects of depletion of CA synthesis were investigated in the sleep-waking cycle in the cat: AMPT decreased waking, and increased paradoxical sleep (with reduced hippocampal kindling) as well as deep slow wave sleep. 78,79
In healthy subjects, AMPT caused some impairment of alertness and sleepiness but hardly any effects on mood. 2 In multiple sleep latency tests AMPT decreased sleep latencies to be reversed by additional administration of L-dopa, 80 while ATDT produced no altered response to AMPT. 81 Similar EEG studies in humans do not exist, but are of special interest, mirroring previous ATDT findings. Still, major limitations of CA depletion are the time consuming study design and the sedative effects of AMPT (requiring a sedative control drug!); moreover, the extent of effects on NE and DA after AMPT remain unclear.
ACUTE PHENYLALANINE/TYROSINE DEPLETION
Transient depletion of dopamine can also be achieved in the APTD paradigm. Here, similar to the ATDT procedure, the ingestion of a highly concentrated AA drink — but this time deficient in the CA precursors phenylalanine, and tyrosine — leads to a general stimulation of protein synthesis inducing a rapid lack of tyrosine and phenylalanine with decreased availability for transport through the blood brain-barrier due to competition with other large neutral AA (tryptophan, leucine, isoleucine and valine). 82 Subsequently, within 7 hours plasma levels decrease to a minimum (tyrosine to 70–85%, phenylalanine to 50%) (Table 1).
However, PET changes in [(11)C]raclopride binding potential (BP) in striatum, a measure of DA D2/D3 receptor availability, were shown to be increased for only 6% after APTD opposed to controls, 83 while d-amphetamine-stimulated DA release was (11.8 ± 11.9%) higher. 84 Regarding mood disturbances, there were no consistent reports in healthy individuals after APTD except for stress reactions to experimental procedure, 2,85,86 and also patients recovered from depression did not relapse. 87
Methodological differences in AA mixtures of APTD (in particular with higher variability of compositions than in ATDT studies), measurement instruments and study populations may account for inconsistencies in current findings. Possibly a higher dose-response relationship should favor APTD as a promising experimental method to investigate CA function especially parallel to similar ATDT study designs. Electrophysiological studies are awaited.
LIMITATIONS AND CONCLUSION
The roles played by 5-HT, NE, and DA, in mediating various cognitive and affective symptoms in humans may be elegantly explored in depletion tests. These methods are usually non-invasive, low-priced, with low placebo response and suitable for neurophysiological studies of monoaminergic vulnerability as well as drug effects. The value of monoaminergic depletion may also lie in the possibility to parallel different depletion methods in combined study designs in order to comprehensively investigate possible interacting neurotransmitters.
In comparison to the rather inconsistent findings in healthy subjects, results of depletion studies in individuals with assumed monoaminergic vulnerability or dysfunction, i.e., patients with neuro-psychiatric disorder, have shown to be correlated to the non-invasive electrophysiological measures. However, despite strong depletion effects, the assumed monoaminergic deficits are not always observable. As opposed to other selective challenges (e.g., with SSRI or NRI) possible causes may be lack of receptor specificity, a rather global manipulation in challenge tests (e.g., 5-HT system in ATDT), or interactions with other neurotransmitters systems (e.g., priority of DA system in APTD). Methodically, challenge studies are usually limited by the time-consuming procedure, variability of dose-dependent relationships and inter-individual monoaminergic levels, lack of test-retest-reliability, carry-over and order of treatments effects. Interactions with gender, personality, and genes could also contribute to heterogeneous findings.
In summary, potential electrophysiological markers for psychophysiological states or traits in humans are not yet investigated sufficiently apart from results of ATDT, which suggest that 5-HT predominantly plays a differential role in EEG activity and sleep regulation as well as in the modulation of early auditory cortical stimulus processing.