
Abstract
Reading speed and efficiency are achieved through the automatic recognition of written words. Difficulties in learning and recognizing the orthography of words can arise despite reiterative exposure to texts. This study aimed to investigate, in native Spanish-speaking late adolescents, how different levels of orthographic knowledge might result in behavioral and event-related brain potential differences during the recognition of orthographic errors. Forty-five healthy high school students were selected and divided into 3 equal groups (High, Medium, Low) according to their performance on a 5-test battery of orthographic knowledge. All participants performed an orthographic recognition task consisting of the sequential presentation of a picture (object, fruit, or animal) followed by a correctly, or incorrectly, written word (orthographic mismatch) that named the picture just shown. Electroencephalogram (EEG) recording took place simultaneously. Behavioral results showed that the Low group had a significantly lower number of correct responses and increased reaction times while processing orthographical errors. Tests showed significant positive correlations between higher performance on the experimental task and faster and more accurate reading. The P150 and P450 components showed higher voltages in the High group when processing orthographic errors, whereas N170 seemed less lateralized to the left hemisphere in the lower orthographic performers. Also, trials with orthographic errors elicited a frontal P450 component that was only evident in the High group. The present results show that higher levels of orthographic knowledge correlate with high reading performance, likely because of faster and more accurate perceptual processing, better visual orthographic representations, and top-down supervision, as the event-related brain potential findings seem to suggest.
Introduction
Oral reading fluency is conceived as the oral translation of text with speed and accuracy. It may serve as an indicator of overall reading competence 1 and is essential for progressive increases in academic achievement. Efficient, low-level word recognition facilitates the higher level, integrative comprehension processing of text that allows the individual to abstract the main idea and relate it to previous knowledge. In fact, the reading experience triggers adjustment processes that progressively allow visual word recognition to become a more automatic and effortless operation.
Orthographic difficulties have been related to developmental dyslexia,2-4 though previous research has focused more on reading than on writing processes of dyslexic students. Several authors assume that there is a failure to abstract orthographic regularities even after repeated exposure to text, as well as difficulty in acquiring automatic word recognition, both of which affect reading speed. 5 The velocity with which text is reproduced into spoken language is the most salient characteristic of skillful reading. 6 Also, in languages characterized by transparent orthographies, reading accuracy is usually high whereas reading speed is extremely low.7,8
Spanish is a transparent language with a regular orthography because of its high grapheme–phoneme correspondence. In contrast to other languages, phonological deficits in Spanish seem to represent a secondary problem, whereas reading speed has been reported as the core deficit in dyslexics.9-11
Studies involving the Spanish language seem to correspond to the notion established by several models of word reading that posit a direct access route from orthography to meaning, bypassing phonology.12-14 However, some authors have postulated that in languages with consistent alphabetic orthographies, children will rely heavily on grapheme–phoneme recoding while reading. 15 Probably because of the fact that in languages with these characteristics the consistency of the orthography is much lower in the direction from phoneme-to-grapheme than from grapheme-to-phoneme (eg, German, Spanish, etc), it has been reported that in dyslexic children the number of phonologically incorrect spellings is low, but they often produce spellings that are orthographically incorrect, indicating that they have not been able to develop an extensive, easily accessible orthographic lexicon.3,16 Also, the buildup of orthographic representations takes a considerable amount of time, 16 with the result that reading speed is very slow, with frequent errors in orthographic spelling; a condition that may persist into adulthood.
In this context, a recent longitudinal study using neuroimaging evaluated the brain activation patterns associated with performance of a rhyming judgment task in young people aged 9 to 15 years, who were tested again approximately 3 to 5 years later. That experiment found significant correlates between brain activation and reading skills, leading its authors to suggest a direct, predictable relation between orthographic analysis and subsequent reading proficiency. 17 In spite of growing evidence of the relation between reading fluency and orthographic abilities in children, few studies have explored this relation in late adolescence and adulthood. Recently, Shany and Breznitz 18 reported a lower activation of the visual association cortex—as indicated by the amplitude of the N170 component of the event-related brain potentials (ERPs)—in accuracy-disabled young adults compared with skilled control readers. In addition, Wimmer et al 19 used a functional magnetic resonance imaging experiment to examine phonological and orthographic decision tasks in adult dyslexics and controls, finding that the former showed lower brain activation in areas involved in visual–orthographic whole word recognition, but increased activity in regions primarily associated with silent articulatory processes, suggesting a grapheme–phoneme reading strategy, instead of whole word processing.
In another study, Mariol et al 20 analyzed ERPs in adults while performing a go/no-go lexical decision task. They presented single words (go trials) and pseudowords (no-go trials), whose orthographic conformation included the addition of double consonants either in legal or illegal positions. Those authors found electrophysiological differences between words and pseudowords later than 230 ms post-stimulus. Moreover, the lexical decision elicited significant changes that were visible in the frontoparietal P3 component—between 300 and 400 ms—and were interpreted as part of the orthographic-based lexical decision.
It seems plausible, then, to expect that the study of electrophysiological responses could be useful to further evaluate orthographic recognition skills and their relation to reading fluency. In this context, we designed the present study to examine more closely differences in the ERPs that underlie weak and strong orthographic abilities in late adolescents during detection of orthographic errors; that is, misspelled words produced by substituting single letters while maintaining identical phonology.
Methods
Participants
Forty-five healthy, right-handed participants were selected from a pool of 430 high school students attending any of 6 different high schools, according to their performance on 5 orthographic-knowledge tasks, and then divided into 3 groups as follows: High (15 participants; mean age = 17.69 years, SD = 0.75 years; total orthographic errors in the 15th percentile or lower); Medium (15 participants; mean age = 17.53 years, SD = 0.52 years; number of errors between the 30th and 70th percentiles); and Low (15 participants; mean age = 17.55 years, SD = 0.69 years; number of errors in the 85th percentile or above).
The 3 groups were matched according to age, handedness, and educational level (12th grade), though gender composition varied among groups (High, 8 males; Medium, 7 males; Low, 12 males). All subjects had a Wechsler Intelligence Scale III (2003) 21 global IQ of 90 or higher and were attending the final grade at a normal high school. None had any personal or family history of psychiatric, neurological, or degenerative illness, nor had they been diagnosed with attention deficit/hyperactivity disorder or any emotional disturbance or behavioral disorder, according to Diagnostic and Statistical Manual of Mental Disorders (fourth edition) criteria. All participants underwent neurological examination and baseline EEG with normal results. Ethical approval for all procedures was obtained prior to the study. Informed written consent was also obtained from all participants.
The tasks used to classify the participants were selected with the objective of depicting orthographic knowledge through the most representative writing exposure situations. All tasks were administered in one single session that lasted approximately one hour. The specific tasks applied were as follows: (1) Word completion: Participants were told that they would see a list of 36 words with letters missing, and that their task was to fill in the blanks to make complete words by choosing between 2 or 3 possible homophone letters (eg, no_ivo, c-s-z). Participants were given an unlimited amount of time to complete the task. (2) Letter dictation: Participants were instructed to write down a letter—narrative text—as it was spoken. The letter had 196 words adapted from a text included in a primary school textbook. (3) Word dictation: Participants were instructed to write down a list of 40 words as they were spoken. Each word contained 2 or 3 syllables and all were susceptible to pseudohomophone errors. (4) Text correction: In an expository text of 276 words, 22 words were replaced by pseudohomophone errors (eg, bacterias × vacterias). Participants were instructed to detect the orthographic errors in the text. (5) Free writing: Participants were asked to write an argumentative text on the topic of abortion. They had unlimited time to complete the assignment.
None of the 5 tasks was previously standardized. Internal consistency was evaluated using the Spearman–Brown split-half reliability coefficient in tasks 1 (word completion, r = 0.721), task 3 (word dictation, r = 0.870), and task 4 (text correction, r = 0.812). The Spearman correlations between performance on each task, and the total number of orthographic errors—with task performance effects partialed out—showed the highest correlation for task 3 (r = 0.805). In addition, task 1 (r = 0.705), task 2 (r = 0.700), and task 4 (r = 0.732) showed high correlations, whereas task 5 showed the lowest correlation (r = 0.367).
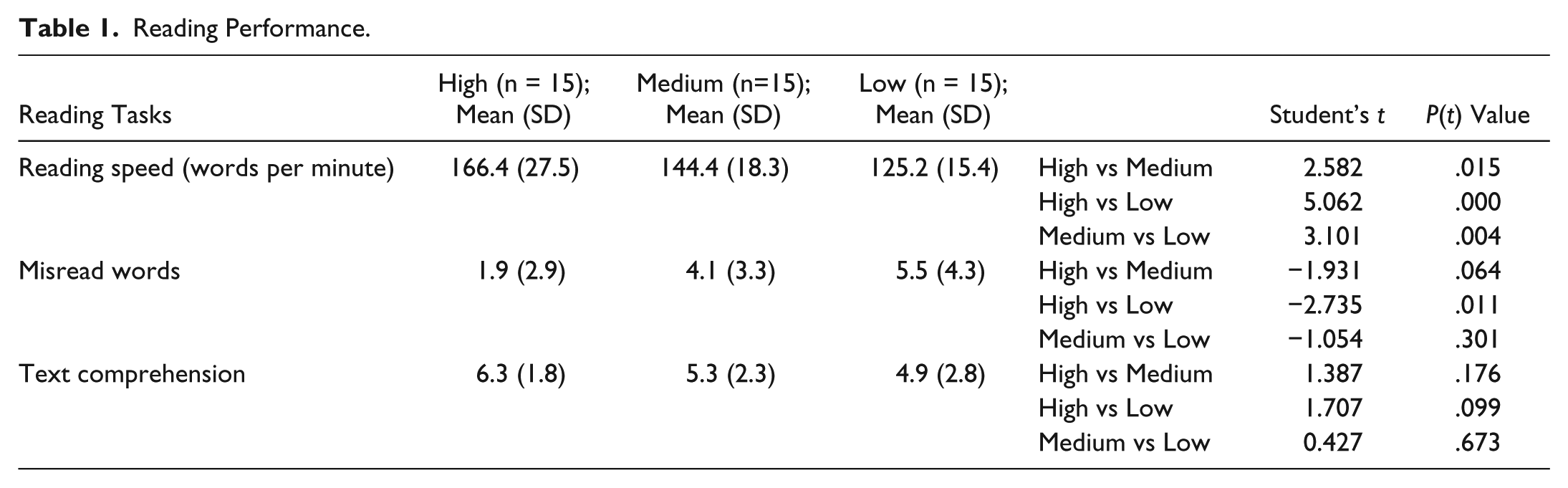
To evaluate the relationship between orthographic abilities and reading performance, all subjects were asked to read aloud an expository text of 504 words. During this exercise, the following parameters were measured: (1) reading speed, (2) number of misread words, and (3) text comprehension. Table 1 shows the results of the reading tests.
Reading Performance.
Experimental Task
All participants performed an orthographic verification task in which each trial consisted of the sequential presentation of an easily named picture from one of three counterbalanced categories (object, meal, or animal). The images appeared for 1000 ms on a 17-inch CRT monitor (screen resolution = 800 × 600; refresh rate = 60 Hz), followed by a 2- or 3-syllable word that matched the picture but was written correctly (W), or incorrectly (OM for orthographic mismatch). The words also appeared for 1000 ms with an interstimulus interval of 1000 ms, and were centrally projected in white lowercase Arial 60 letters against a black background, subtending a visual angle of 0.75°. Subjects were instructed to determine whether the word was correctly written, as quickly as possible, by pressing a keyboard key with their left or right index fingers, respectively. The key order was counterbalanced across conditions (correctly or incorrectly written words) and subjects.
Stimuli
A total of 144 picture–word pairs were used as stimuli with 72 being used in each experimental condition. The words were common, 2-syllable (57.9%) or 3-syllable (42.1%) nouns. The accompanying images were black and white pictures (250 × 162 pixels) at 640 × 480 dpi resolution, and a size of 5.8 × 8.3 cm. The pictures used as stimuli were selected from Snodgrass and Vanderwart, 22 based on their standardization for Spanish. 23
With respect to the words selected, 72 were correctly written (W), whereas the other 72 (OM) were replaced by pseudohomophones (ie, one letter was replaced to create a plausible orthographic error, while maintaining the same phonology of the real word; example in Spanish: corazón × corasón; similar to circus × sircus in English; see Figure 1). There were no significant differences in the number of syllables between conditions.
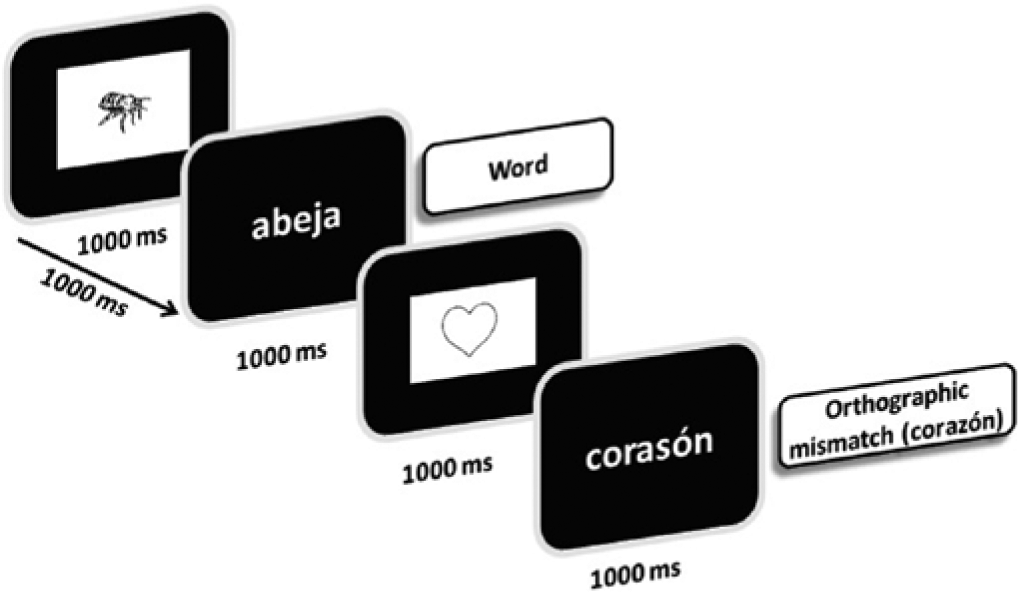
Flowchart of the experimental task used to obtain the event-related brain potentials.
Procedure
Subjects were tested in a sound-attenuated room. The list containing all the picture–word pairs from the 2 experimental conditions (W, OM) was semi-randomized and divided into 3 blocks to be administered with a brief rest interval. The presentation order of the blocks was appropriately counterbalanced among subjects, who were instructed to minimize eye and body movements during the experiment. All participants were adequately trained before performing the task.
Stimulus delivery, response collection, and data acquisition onset were all synchronized and controlled by the MIND-TRACER software (Neuronic S.A., Havana, Cuba, 2003).
Electrophysiological Methods
Recording
EEG activity was recorded from the Fp1, Fp2, F3, F4, F7, F8, C3, C4, P3, P4, O1, O2, T3, T4, T5, T6, Fz, Cz, and Pz scalp sites. All electro-oculograms (EOGs) were recorded from the outer canthus and infraocular orbital ridge of the right eye. Electrophysiological recordings were made using 10-mm diameter gold disc electrodes (Grass Type E5GH, Natus Neurology Inc, Middleton, WI) and Grass electrode cream. All recording sites were referred to linked mastoids. Inter-electrode impedances were less than 5 kohm at 30 Hz. EEG and EOG signals were amplified at a bandpass of 0.5 to 30 Hz (3-dB cutoff points of 6 dB/octave roll-off curves) with a sampling period of 5 ms on the MEDICID-03E system. Data were collected as 1.100-second epochs for all recording channels beginning 100 ms prior to stimulus onset (word or pseudohomophone). Single trial data were stored off-line for averaging and analysis.
Data Analysis
Behavioral Measures
Correct and incorrect responses were automatically marked on the EEG by the software; reaction times were recorded simultaneously.
Signal Averaging
Epochs of data on all channels were excluded from averages when the voltage in a given recording epoch exceeded 100 µV on any EEG or EOG channel. Epochs with artifacts were also rejected by visual inspection. Twenty artifact-free correct trials—randomly selected across each EEG record—were considered to obtain the individual ERP in each condition. Every individual ERP reached a standard deviation rate less than 1.1 and a residual noise level (RNL) less than 2.
Statistical Analysis
Two-factor analyses of variance (ANOVAs; Group × Condition) were performed to examine correct responses and reaction times during experimental task performance. Pearson correlations were used to explore the relationships between behavioral responses during the experimental task and reading performance.
According to the visual examination of the resulting group-averaged ERP waveforms, the main voltage variations were estimated to examine the electrophysiological data in several time windows. Split-plot ANOVAs (Group × Condition × Recording Site) were conducted on average voltages across each time window as the dependent variable. Greenhouse–Geisser corrections to the df were applied as needed, with the corrected probabilities reported. Additionally, post hoc Tukey’s honesty significant difference tests were done to explore any trends in the observed changes.
Results
Behavioral Results
On analyzing the number of correct responses, significant differences among groups (F2,42 = 22.6, P < .001, η2p = 0.518) and between conditions (F1,42 = 67.3, P < .001, η2p = 0.616), as well as a significant interaction between the 2 factors (F2,42 = 14.193, P < .01, η2p = 0.403) were found. Post hoc analyses demonstrated that both Medium (P < .05) and Low (P < .01) had lower numbers of correct responses than High, but only when processing orthographic mismatches. In general, the three groups achieved more correct responses when processing correct words than when orthographically incorrect words were shown (P < .01). Table 2 shows the behavioral performance of each group.
Behavioral Performance on the Experimental Task.
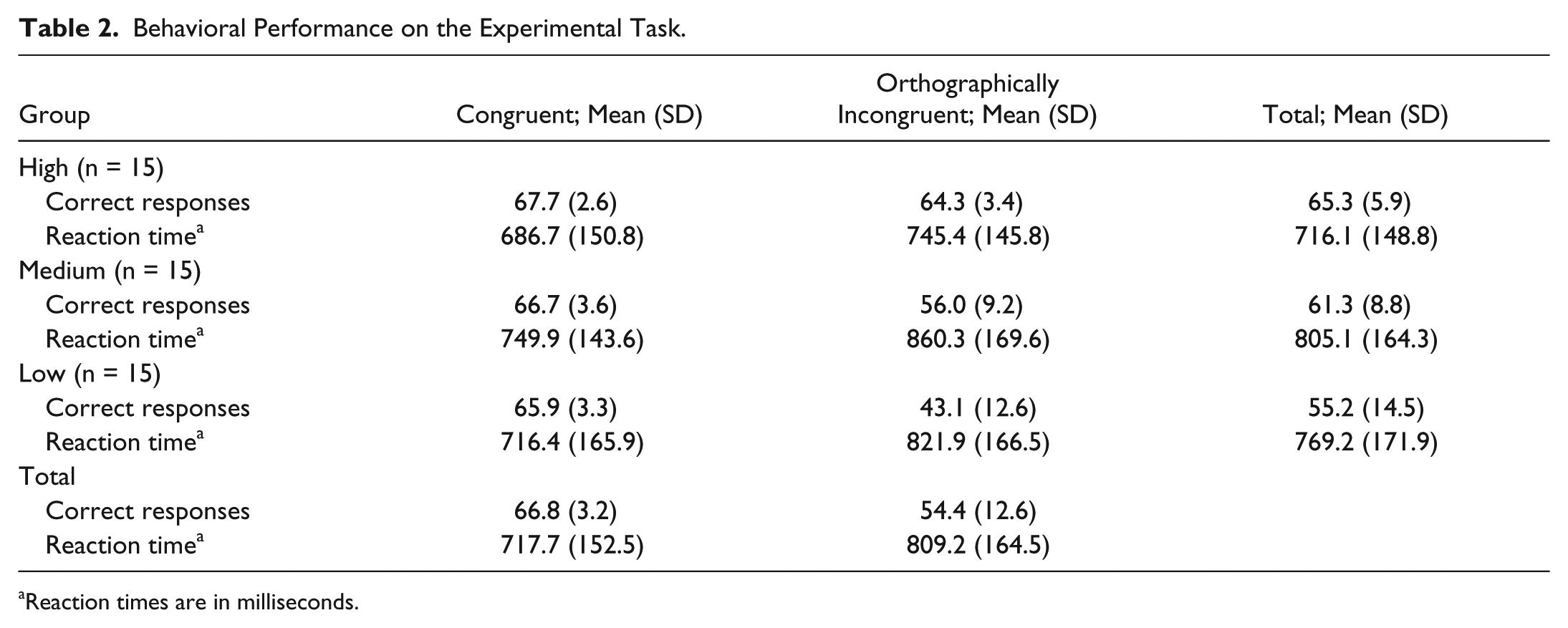
Reaction times are in milliseconds.
Pearson correlations showed that the number of correct responses while processing the orthographic errors correlated positively with reading speed—that is, the number of words read per minute (r = 0.469, P < .01)—whereas its correlation with the number of misread words was significantly negative (r = −0.376, P < .05). Thus, reading performance by subjects with poorer orthographic abilities was both less accurate and slower.
In addition, the 2-factor, split-plot ANOVAs showed significant differences in reaction times between conditions (F1,42 = 75.4, P < .001, η2p = 0.642); indicative of prolonged times when processing orthographic errors in all groups.
Electrophysiological Results
To analyze the electrophysiological results, 3-factor, split-plot ANOVAs (Group × Condition × Recording Site) were conducted on average voltages across each time window as the dependent variable.
Regarding the visual examination of the group-averaged ERP waveforms, 5 main ERP components were observed—P150, N170, P200, P450, and P600. Thus, 4 time windows were selected to evaluate the electrophysiological data, from 50 ms before to 50 ms after each maximum voltage main peak. The factor Recording Site was analyzed in locations where the main visually detected changes took place.
Because of its predominant topographical distribution, the analysis of P150 was performed at locations P3, P4, O1, O2, T5, and T6, and showed significant interactions between Condition and Recording Site (F2,42 = 2.64, P < .05), where post hoc tests showed that higher voltage amplitudes in correctly written words were reached at T5 and O1 (P < .05). The analysis of P150 latency revealed no significant change across conditions. Figure 2 shows the ERPs that correspond to the locations at which the analyses of P150 and N170 were performed.
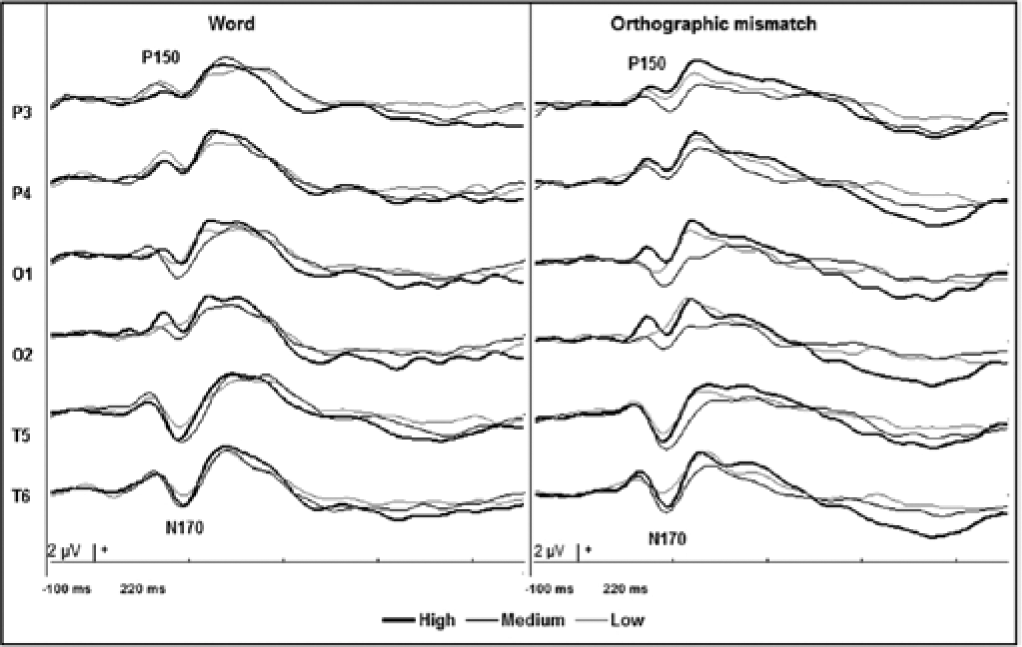
Grand-averaged, event-related brain potentials while performing an orthographic decision task at temporal–parietal and occipital recording sites.
The N170 component seemed to reach higher amplitudes over the posterior temporal areas, especially in the left hemisphere. The analysis of N170 showed a significant effect only for the factor Recording Site (F1,42 = 11.25, P < .001). However, the analysis of its latency showed a significant interaction between Condition and Recording Site (F2,84 = 3.43, P < .05), which suggests that N170 latency was more prolonged at T6 than at T5 during orthographic errors (P < .01). Poorly defined lateralization was evident for N170 in the Low group, which exhibited a tendency to reach lower voltage magnitudes while detecting correctly written words (see Figure 2).
The analyses of the subsequent P200, P450, and P600 components were conducted at locations F3, F4, C3, C4, Fz, Cz, and Pz, where the principal changes were discernible. The component P200 showed no statistically significant effects, probably because of individual variability. However, the analysis of P450—a positive component primarily distributed over the frontal–central areas—did show significant differences between conditions (F1,42 = 11.28, P < .01), indicating that significantly higher voltages were reached while processing orthographic errors. Also, a significant interaction between Group and Condition (F2,42 = 7.50, P < .01) was found. Post hoc analyses demonstrated that High reached lower voltages than Medium (P < .05) and Low (P < .05) under the correct words condition. When processing orthographic mismatches, only High showed significant differences between conditions (P < .01), by reaching higher voltages in the case of orthographic errors. The analysis of P450 latency showed no significant variations across conditions.
Finally, the analysis of P600 showed a significant effect for the factor Condition (F1,42 = 6.85, P < .001), with a significant interaction between Group and Recording Site (F2,84 = 1.99, P < .05). With regards to the post hoc analyses, they denote that Low and Medium had higher voltages over parietal areas compared to High (P < .05 and P < .05, respectively). Figure 3 shows the midline ERPs at which the P200, P450, and P600 components are observed.
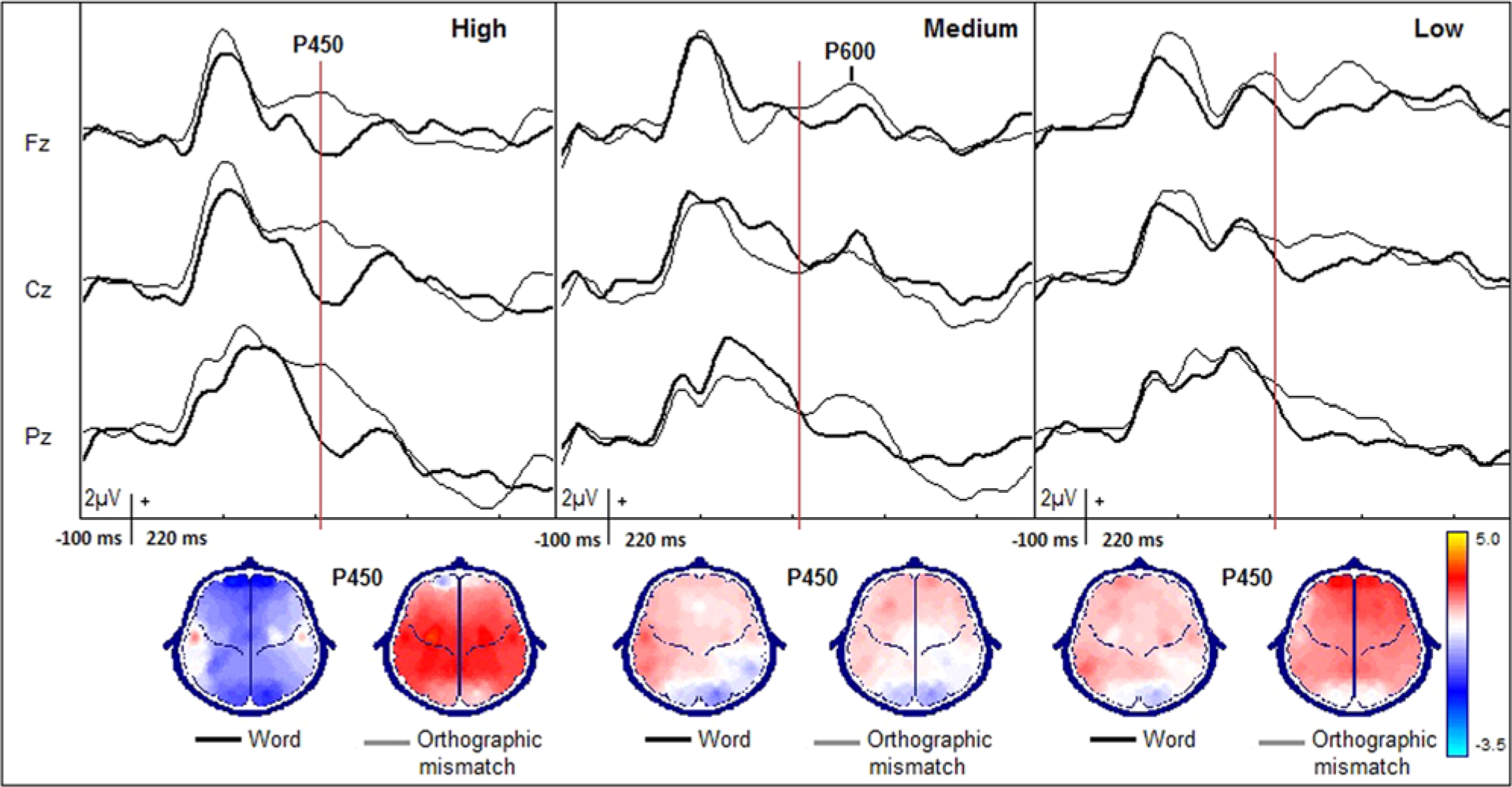
Midline grand-averaged, event-related brain potentials during performance of an orthographic decision task.
Discussion
In languages that use the Latin/Roman alphabet, the ability to read words “by sight” (ie, automatically) seems to rest on the ability to map letters and letter combinations to sounds. 24 Indeed, the development of insights into how letters and sounds correspond allows the learner to memorize differences between words that may not be visually very distinctive (eg, male, malt, mole). Given that reading and spelling build on, and rely on, the same mental representation of a word, the instructions and information needed to learn how to spell words could be used in both spelling and sight reading. Furthermore, it has been postulated that the spelling of a word makes the mental representation robust in memory and readily accessible for fluent reading. 25
As spelling proficiency develops, knowledge of both the consistencies and inconsistencies of the language is gained. In this context, spelling has been related to implicit memory and cognition, since the use of “rules” during spelling may involve “implicit knowledge of sound–spelling correspondences, orthographic regularities, frequencies, and statistical probabilities used during automatic retrieval, as well as explicit phonological knowledge and knowledge of orthographic conventions” (see Steffler 26 for an extensive review). In other words, limited orthographic knowledge might be an expression of inadequate language and reading development, which might limit, in certain aspects, the subsequent individual adaptation to higher educational demands. Based on this reasoning, we evaluated orthographic skills and their relation to reading fluency with the aim of more closely examining differences in the ERPS that underlie weak and strong orthographic abilities in late adolescents during detection of orthographic errors.
With regards to the nature of the task, and based on the specific group selected, behavioral results confirmed the expected differences in orthographical error detection. Also, the prolonged reaction times suggest that the increased difficulty caused by the appearance of a pseudohomophone correlated with greater processing effort; a finding that agrees with previous literature on this topic.27,28
Despite the fact that all participants were regular students and none had references of any previous reading disorders, there were significant differences among groups in reading performance, as the subjects in Low showed significantly slower reading speeds than those classified as High and Medium orthographic performers. Indeed, the Low group showed an average reading speed within the level recently reported for Mexican students attending the sixth grade. 29 Low reading speed is considered a core deficit in Spanish dyslexics; thus, our results suggest that poor orthographic knowledge could be related to reading problems.
In addition, the number of participants by gender was not equivalent between groups, because of the results from the earlier screening conducted to select the experimental samples. The higher number of males in the Low group seems to be consistent with previous reports of higher prevalence of reading disabilities in males.30,31
ERP Findings
The first main component—P150—showed a temporal–parietal–occipital distribution that could be related to early visual perceptual linguistic categorization that likely depends on the posterior fusiform gyrus. 32
Early changes in ERP, as reflected by P150, could represent a word superiority effect, as proposed by Coch and Mitra. 33 However, also as per these authors, it could be associated with orthographic fluency, probably depending on the availability of processing resources. In this context, it is reasonable to assume that the P150 component might index visual categorization processes, sensitive to orthographic regularity, as part of a visual route to an orthographic input lexicon. 34 The effect found in the P150 component—lateralization to the left—fits with these previous theoretical assumptions.
However, nonsignificant effects were found among groups. This could be attributed to an inter-individual variability effect in view of the fact that, as has been reported previously, 35 the voltage effects of P150 are relatively small. Alternatively, the lack of significant effects among groups could be because of the relative preservation of early perceptual processes in each one.
In general, the N170 component has been interpreted as a hallmark of visual orthographic specialization36,37 that may reflect increased visual processing expertise, 38 most likely in prelexical orthographic processing. 39 The present results seem to correspond well to previously reported findings of the N170 component, where source localization and imaging studies have shown that this early stage of perception processing occurs in the fusiform gyrus, and is lateralized depending on the nature of the stimuli (left side for words; right side for pictures).40,41 Accordingly, the N170 component was lateralized and also faster at the left side when orthographic errors were analyzed, probably reflecting sublexical perceptual processing. However, nonsignificant changes among groups were found, even though Low seemed to be less lateralized and to have lower voltage magnitudes while detecting correctly written words.
When evaluating the functional significance of later ERP components, an effect of the orthographic mismatch was obtained later, at around 450 ms, with orthographic errors eliciting larger amplitudes in comparison to correctly written words at frontoparietal sites. Regarding latency and topography, P450 resembles the extensively documented P3 component, which appears when a stimulus is perceived and attentional resource allocation 42 and memory operations 43 are involved (see Polich et al 44 for a review). This seems to be the case in the present study, with greater emphasis on the orthographic incongruent condition. When a task imposes greater demands on attention resources, the result is larger P3 amplitudes, but in the present experiment, only High was sensitive to these demands. This result could reflect the higher capacity of this group to differentiate an orthographic anomaly as a product of the comparison between the incoming visual stimuli and the expected mental template.
Another analog to the present P450 waveform was found in an experiment conducted to study the effect of word frequency on the processing of emotional words. 45 This component was interpreted as reflecting adjustments in attentional mechanisms to benefit lexical information processing. The P450 waveform could reflect a late stage of processing, 40 probably associated with recognition processes. In the present experiment, P450 was significantly greater in High while recognizing orthographic mismatches; a finding that would be compatible with the greater orthographic knowledge demonstrated by this group.
Finally, as per the findings from Mariol et al 20 described above, one could hypothesize that the P3 analogue would be greater when there are stronger mental representations of the expected word. In short, lower orthographic recognition in Medium and Low might be derived from a lower ability to construct adequate mental representations of the words during the reading experience.
Regarding the temporal dynamics and topographic distribution, one might relate the late positivity observed in Medium and Low to the P600 effect typically observed in previous studies with syntactic or semantic manipulations (see Gouvea et al, 46 Kolk and Chwilla, 47 and Kuperberg 48 for reviews). However, in the present experiment, semantic congruence remained intact despite the orthographical errors, and there were no evident syntactic inconsistencies. The recent finding of a P600-like component elicited by violations of derivational morphology in Spanish, 49 provides evidence to suggest that the detection of orthographic errors in subjects with lower orthographic abilities might depend, at least in part, on more unspecific analyses, such as those morpheme-based processes reflected by P600. Alternatively, one could conjecture that the P600 waveform might be a delayed analogue of the P450 described for High. However, the lack of significant differences in response times among groups gives little credence to this assumption.
Conclusions
Taken together, earlier findings and the results of this study suggest that orthographic accuracy in a transparent language, such as Spanish, seems to depend on a complex combination of factors that run from the earliest steps of low-level perceptual processes—including sublexical processing—to higher level neural substrates, where priming and other comparing–recovering mechanisms may play an important dynamic role, along with the integrity of the neural circuits that are essential for reading. The present study confirms that spelling skills and reading speed are closely related in Spanish. In addition, it offers insight on late adolescents with high and low orthographic knowledge. However, the small sample size might limit the scope of the present study, and more extensive research on the neural structures involved in reading and their functional interplay, is needed to clarify neural availability and connectivity, among other intriguing topics related to reading processes and associated developmental disorders.
Footnotes
Acknowledgements
We thank Paul Kersey and Vanessa Ruiz-Stovel for their revision of the English version.
Authors’ Note
The first two authors (AAG-G and FRG-V) contributed equally to this work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was partially supported by CONACYT Grant 80906 and the Neuroscience Institute (University of Guadalajara, Mexico).