
Abstract
Methadone maintenance treatment (MMT) has been used as a treatment for opiate dependence since the mid-1960s. Evidence suggests that methadone binds to mu opiate receptors as do other opiates and induces changes in neurophysiological function. However, little is known, about how neural activity within the higher frequency gamma band (>30 Hz) while at rest changes in those stabilized on MMT despite its association with the excitation-inhibition balance within pyramidal-interneuron networks. Our study investigated differences in resting gamma power (37-41 Hz) between patients undergoing MMT for opiate dependence, illicit opiate users, and healthy controls subjects. Electroencephalographic data were recorded from 26 sites according to the international 10-20 system. Compared with the healthy controls subjects, people either undergoing MMT (mean difference [MD] = 0.32, 95% CI = 0.09-0.55, P < .01) or currently using illicit opiates (MD = 0.31, 95% CI = 0.06-0.56, P = .01) exhibited significant increased gamma power. The sLORETA (standardized low-resolution electromagnetic tomography) between-group comparison revealed dysfunctional neuronal activity in the occipital, parietal, and frontal lobes in the patients undergoing MMT. A more severe profile of dysfunction was observed in those using illicit opiates. Our findings suggest that long-term exposure to opioids is associated with disrupted resting state network, which may be reduced after MMT.
Introduction
Methadone is a synthetic opioid that has been used as a pharmacological substitute for the treatment of opiate dependence since the mid-1960s. Methadone elicits its pharmacodynamic effects by binding to mu opiate receptors, as do other opiates, but it has a much slower onset and longer duration of action partly because of its oral absorption.1,2 Methadone-related electroencephalographic (EEG) changes have been observed in the beta, alpha, theta, and delta frequency bands, reflecting the neurobiological underpinning of potential cognitive impairments associated with MMT. For example, in the study by Martin et al, 3 EEG recordings were conducted 2 weeks prior to MMT, during the administration of methadone, then after stabilization on 100 mg/d of methadone, and following 6 weeks abstinence from MMT. The study found reduced alpha activity and increased delta and theta activity during the induction and stabilization periods of MMT compared with the period before MMT. During the period of withdrawal from MMT, delta activity was significantly less than the periods either before MMT was initiated or after they were stabilized on 100 mg/d of methadone. A more recent study by Fingelkurts et al 4 demonstrated lower levels of alpha and beta activity and increased delta and theta activity in patients undergoing MMT compared with healthy volunteers. However, little is known about how neural activity within the higher frequency gamma band (>30 Hz) changes in patients stabilized on MMT.
Synchronous gamma activity is thought to reflect neural activity coordinating internal state for the purpose of cognitive demand.
5
It has been suggested that information processing involves multiple neurons and pathways distributed across different regions. To perform a particular task, there is a need to combine individual neuronal signals into a cognitive process.5,6 Therefore, gamma activity is closely correlated with cognitive function. Reduced gamma amplitude has been observed in Alzheimer’s disease
7
and people from densely affected alcoholic families,
8
whereas enhanced gamma amplitude has been found in those with epilepsy
9
and cigarette smokers.
10
It has been argued that a balance between GABAergic interneurons and excitatory glutamatergic pyramidal cells contributes to changes in gamma activity.
11
Therefore, the observed reduction in patients with Alzheimer’s disease might be related to the age-dependent loss of dopamine D2 receptor, which modulate both GABAergic and gultamatergic transmission.
7
For people at high risk of alcoholism, reduced gamma activity is thought to be associated with dysfunctional GABAergic mechanisms.
8
In contrast, increased gamma activity in epilepsy and smokers is related to decrease neuronal inhibition and excessive activation of the glutamatergic system.
12
Consistently, recent studies on gamma activity in schizophrenia have suggested that dysfunctions in gamma activity might account for the heterogeneity of symptoms. Decreased gamma activity has been found in patients with negative symptoms, for example, slowing of mental activity, while abnormal increased gamma activity has been found in patients with positive symptoms, for example, reality distortion.13,14 It has been suggested that decreased gamma activity might reflect a deficit in the GABAergic interneuron systems and increased gamma activity might be caused by both N-methyl-
Methods
Participants
All participants were free of a history of psychotic disorder, depression, cardiac disease, endocrine disorder, head trauma, neurological disease, and current pregnancy or breast-feeding, screened by the Composite International Diagnostic Interview (CIDI). The group undergoing MMT were recruited following recommendations from the MMT case managers at Auckland Community Alcohol and Drug Services (CADS), New Zealand. They were required to undertake the MMT for a minimum of 6 months and be stabilized on their current dose for at least 2 weeks. The MMT group consisted of 18 males and 14 females, with a mean age of 39.36 years (SD = 5.14). Their mean duration of education was 12.06 years (SD = 2.00); mean duration of opiate use was 10.03 years (SD = 6.08); mean duration of MMT 7.29 years (SD = 6.39); and current methadone dose 70.86 mg/d (SD = 40.61; range 8−180 mg/d). Opiate users were recruited from the Auckland Drug Information Outreach (ADIO) Trust Needle Exchange Services by advertisement or by word of mouth. They were not required to abstain from opiate use before testing. This group included 11 males and 6 females, with a mean age of 37.38 years (SD = 7.44); a mean duration of education 12.47 years (SD = 1.46) years; and a mean duration of opiate use of 11.41 years (SD = 8.60). A group of 25 healthy control subjects without histories of substance abuse was recruited by advertisements distributed in a range of local community centres such as the public library, shopping mall, cafés, or word of mouth. This group included 14 males and 11 females, with a mean age of 36.12 years (SD = 6.61) years and a mean duration of education 13.71 years (SD = 1.73). The 3 groups were matched for age, gender, and ethnicity.
Data Recording
Prior to commencing this research, ethical approval was granted by the Northern Regional X Ethics Committee of New Zealand and informed consent was given by all participants.
A QuickCap (Neuroscan 4.3) 40 sensor shielded cap was used to acquire EEG data from the cephalic sites. The 26 cephalic sites included Fp1, Fp2, Fz, F3, F4, F7, F8, Cz, C3, C4, FC3, FCz, FC4, T3, T4, T5, T6, Pz, P3, P4, O1, O2, and Oz electrode sites (10-20 international system). A further 14 channels recorded other data, for example, electrooculogram (EOG), heart rate (HR), muscle movements, events, and so on. Skin resistance was kept at <5 kohm. Scalp and EOG potentials were amplified and digitized continuously by a system (NuAmps, SCAN 4.3) having a frequency response from DC to 100 Hz (above which attenuating by 40 dB per decade), and a sampling rate of 500 Hz. EEG data were screened visually for artifacts, normal variants, and changes in alertness (the technician screening these data was blinded to group status).
All EEG recordings were conducted in a sound and light attenuated laboratory. Electrical impedance was always <5 kohm. Resting EEG data were collected in 2 states: eyes open (EO) and eyes closed (EC). In the EO condition, participants were asked to fixate on a red dot on the center of the computer screen for 2 minutes. In the EC condition, participants were asked to sit still with eyes closed for 2 minutes.
Data Reduction
The eye-blink correction algorithm took continuous EEG data and performed EOG artifact correction. Correction coefficients were calculated for both vertical and horizontal EOG data. The difference of channels HPHL (horizontal positive horizontal left) and HNHR (horizontal negative horizontal right), and the difference of channels VNVB (vertical negative vertical below) and VPVA (vertical positive vertical above) were calculated to create horizontal and vertical data arrays (EOGh and EOGv, respectively). The corrected EEG data were calculated by subtraction of vertical and horizontal EOG, each with an independent linear correlation coefficient.
The 2 minutes of EEG were divided into adjacent intervals of 4 seconds. Signals with a sharp edge (nonzero initial or final values) at the beginning and end of the epoch were excluded. Average power spectra were computed for 28 epochs during resting state EEG. Spectral power analysis was performed on each four-second interval by first applying a Welch window to the data, and then performing a fast Fourier transform. The 4 second windows are 50% overlapping, which gave 59 complete windows over the 2-minute recording period. Using fast Fourier transform, a fixed duration time window was uniformly applied to all frequencies and EEG data then reformatted into respective frequency spectral patterns. Epochs were rejected if the signal at 3 or more sites exceeded 100 μV for that particular epoch. If more than 50% of the epochs were rejected, then the entire analysis was considered faulty. Each power spectrum was detrended from 14 to 49Hz by subtracting the line of best fit prior to obtaining gamma power for the 37- to 41-Hz bandwidth, in order to remove any broad-spectrum EEG or electromyographic activity. This has been considered an effective approach to minimize the role of any electromyographic contaminants, and has been used by others.17-19
The power spectra were averaged for each electrode position and calculated for gamma (37-41 Hz) frequency range. Power data were log transformed to better approximate the normal distributional assumptions required by parametric statistical methods. Missing data were not replaced.
Data Analysis
Gamma power at 26 sites was divided into 4 regions by averaging within each region: frontal (Fp1, Fp2, F7, F3, Fz, F4, F8), central (FC3, FCz, FC4, C3, Cz, C4, CP3, CPz, CP4), temporal (T3, T4, T5, T6), and parietal-occipital regions (P3, Pz, P4, O1, Oz, O2). This lobe-based EEG regional analysis is supported by simultaneous EEG and MRI recordings; it has been shown that neuroanatomical measures of structural changes for the frontal, parietal, temporal, and occipital lobes are consistent with absolute EEG power calculated based on this regional division. 20 This regional division has also been used in previously published EEG research. 21-23
Group comparisons of spectral power were analyzed using 3-group (MMT vs opiate user vs healthy controls) repeated-measures analysis of variance, including recording condition (EO vs EC) and power within regions (frontal vs central vs temporal vs parietal-occipital) as the within-subjects factor. Education was entered as a covariate, since there was a significant group difference in education, F(2, 71) = 7.01, P = .002. The degrees of freedom were adjusted with a Greenhouse-Geisser correction when necessary. Significant main effects were followed-up with post hoc pairwise comparisons adjusted using a Sidak correction.
Localization of the differences in electrical activity of gamma band between groups was assessed by voxel-by-voxel unpaired t tests of the sLORETA, based on the log-transformed power of the estimated electric current density. 24 Statistical significance was assessed nonparametrically with a randomization test that corrects for multiple comparisons. 25 The results correspond to maps of t statistics of log-transformed data for each voxel, for corrected P < .01. It has been suggested that the statistical nonparametric mapping (SnPM) methodology corrects for all multiple comparisons, and does not require any assumption of gaussianity. 26 The voxels with significant differences were located in specific brain regions using sLORETA images: the red color means that patient subjects had an increased activation compared with the controls, and blue means a decreased activation when compared with healthy controls.
Results
There was a significant difference between groups on gamma power (F = 6.67, P = .002). Compared with the healthy controls subjects, people either undergoing MMT (mean difference [MD] = 0.32, 95% CI = 0.09-0.55, P < .01) or currently using illicit opiates (MD = 0.31, 95% CI = 0.06-0.56, P = .01) exhibited increased gamma power. The group differences were observed across the 4 regions during either the EC or EO conditions (Figure 1).
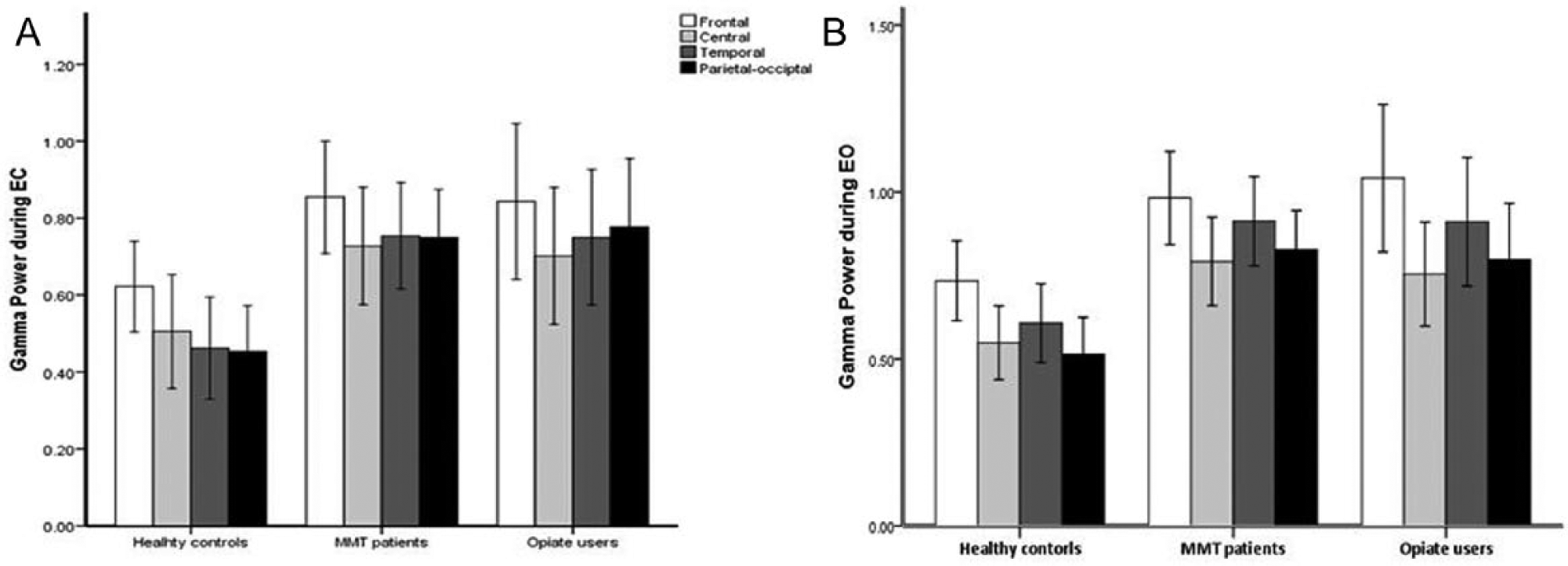
Regional gamma power by group status during the eyes closed (EC) and eyes open (EO) conditions.
The sLORETA between-group comparison revealed that relative to the healthy controls subjects, those undergoing MMT exhibited significant increased activation in the occipital lobe, and decreased activation in the parietal and frontal lobes during EO, and increased activation in the occipital lobe during EC (Table 1). In contrast, illicit opiate users showed significantly decreased activation in the parietal, occipital, frontal and limbic lobes during EC, but increased activation in the occipital lobe and decreased activation in the frontal lobe during EO, relative to the healthy control subjects. Compared with those currently using illicit opiates, patient undergoing MMT showed increased activation in the frontal, parietal, and occipital lobes during EO, and increased activation in the occipital and decreased activation in the parietal and frontal lobes during the EC (Table 1).
Group Comparisons (t Statistics) of Mean Current Density During the Eyes Open (EO) and Eyes Closed (EC) Conditions. a
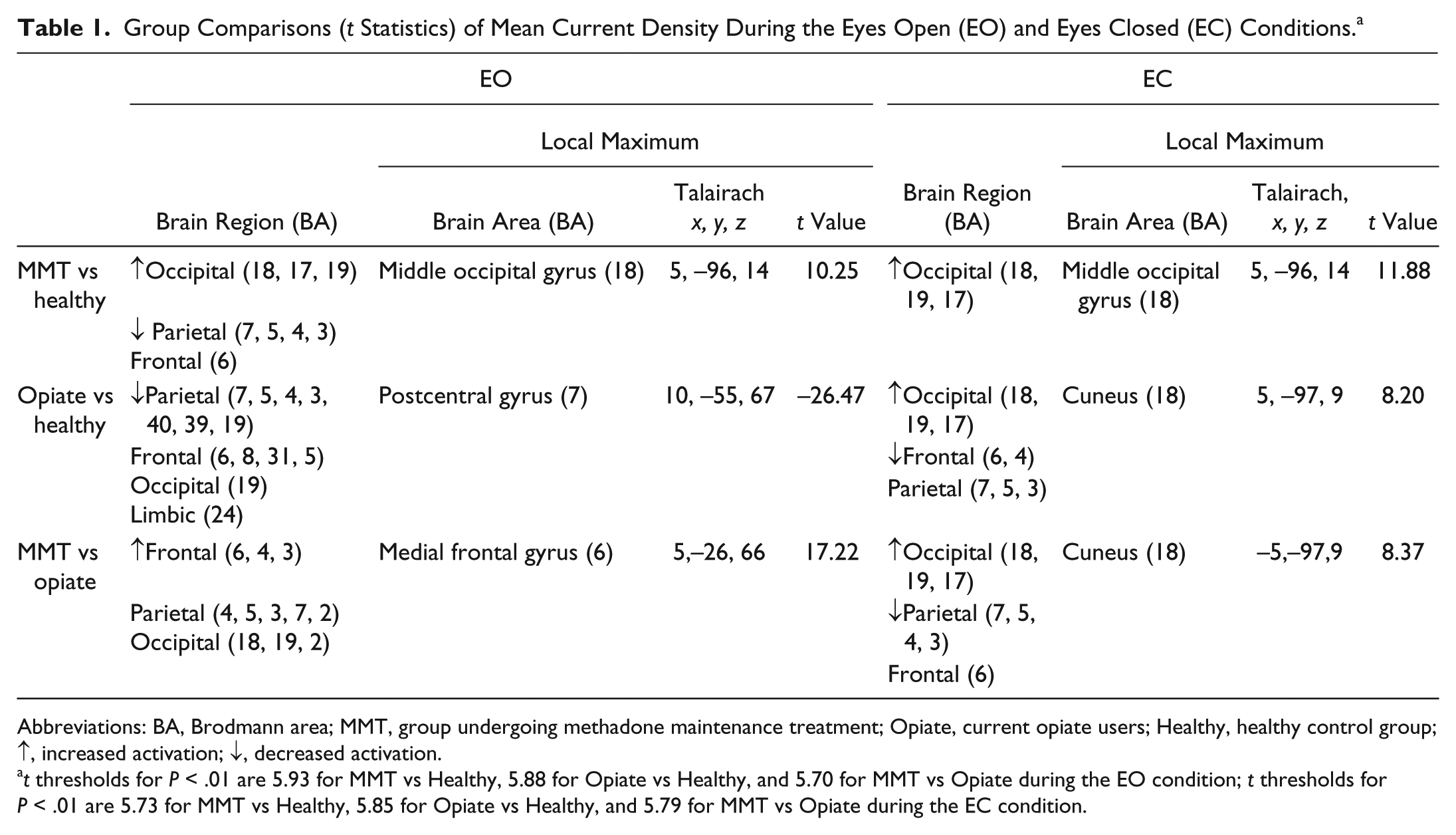
Abbreviations: BA, Brodmann area; MMT, group undergoing methadone maintenance treatment; Opiate, current opiate users; Healthy, healthy control group; ↑, increased activation; ↓, decreased activation.
t thresholds for P < .01 are 5.93 for MMT vs Healthy, 5.88 for Opiate vs Healthy, and 5.70 for MMT vs Opiate during the EO condition; t thresholds for P < .01 are 5.73 for MMT vs Healthy, 5.85 for Opiate vs Healthy, and 5.79 for MMT vs Opiate during the EC condition.
Furthermore, group comparison (t statistics) between patients undergoing MMT and healthy controls subjects showed a maximum difference in current density in the middle occipital gyrus in Brodmann area (BA) 18 during either EO (Figure 2) or EC (Figure 3). The maximum differences between opiate users and healthy controls subjects were located in the postcentral gyrus in BA7 during EO, and in the cuneus in BA18 during EC. The maximum differences between the MMT and opiate user group were located in the medial frontal gyrus (BA6) during the EO, and in the cuneus (BA18) during the EC.
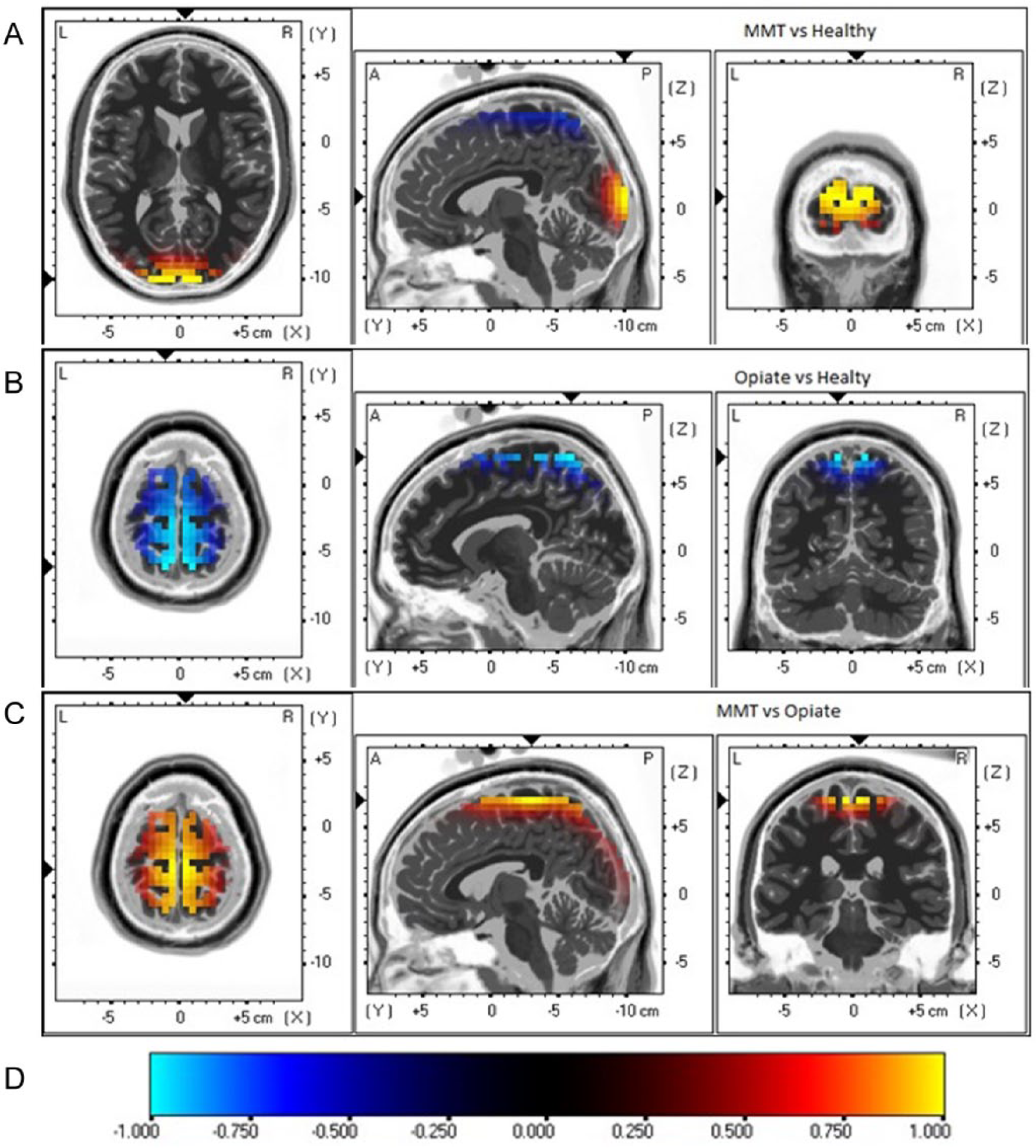
Images of maximum group differences on brain regional electrical activity during the eyes open (EO) condition using sLORETA. MMT, group undergoing methadone maintenance treatment; Opiate, current opiate users; Healthy, healthy control group.
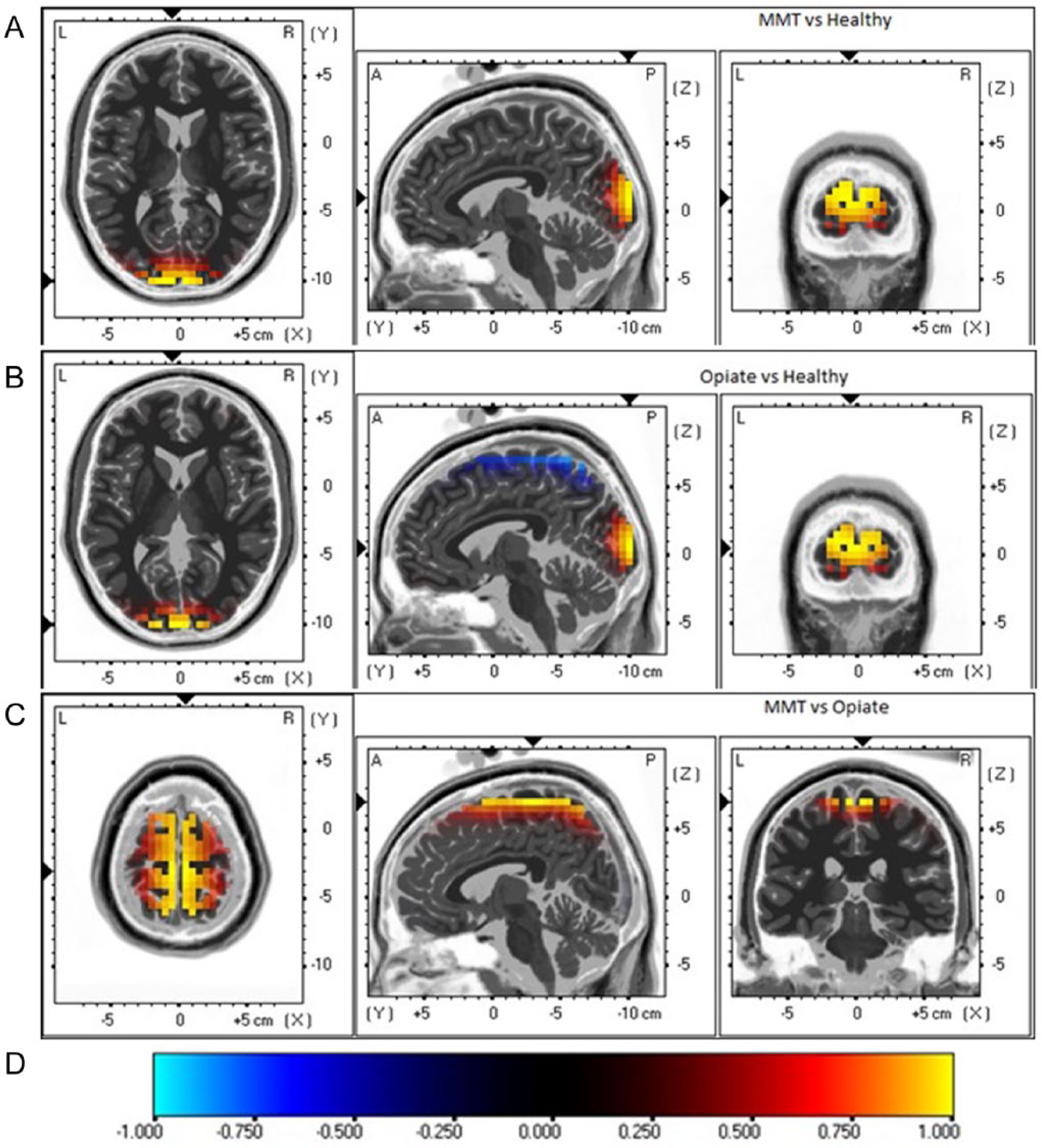
Images of maximum group differences on brain regional electrical activity during the eyes closed (EC) condition. using sLORETA. MMT, group undergoing methadone maintenance treatment; Opiate, current opiate users; Healthy, healthy control group.
Discussion
The current study investigated the effect of MMT on neurophysiological function by comparing the resting state gamma power of patients undergoing MMT with current opiate users and healthy volunteers. Our results are in good accordance with the majority of human EEG studies focusing on the effects of opiates on neurophysiological function,3,27,28 and indicate that long-term exposure to opioid-type drugs is associated with a disrupted resting state network. It has been suggested that resting state EEG represents the readiness or “potential” of the brain to allocate neural resources essential to cognitive function, for example, information processing and attention, 29 Spontaneous gamma activity that contribute a fraction of the total EEG power is driven by thalamocortical resonant synaptic interactions 11 and reflects an integration mechanism of the brain. 30 Disturbances of integration probably underlies a deficiency in coordination between and within areas involved in a task-relevant network, for example, binding distinct aspects of object perception into a coherent and unitary concept. 11 It has been shown that gamma activity becomes more apparent when subjects recognize a meaningful picture within random-dot patterns (autostereoscopic pictures). 31 Significant, 2- to 5-fold increases in gamma power have been observed at widespread posterior and central scalp sites during task of learning, reading, and subtraction. 32 It has been argued that a dysfunctional mental state can be produced by either increased or decreased of gamma activity. 15 Differences in gamma power between people with a history of opiate use in contrast to non-drug users may provide an index of modulation for cognitive function. Evidence shows that spontaneous steady-state and event-related gamma oscillations might be generated by the same neural circuits. 33 Cognitive impairments associated with opioids have been reported in many neuropsychological studies 34 ; for a review, see Wang et al. 35 Our findings and those of other neuropsychological studies are complementary in demonstrating abnormalities in people with opioid dependence. In the resting EC state, there is no external task-demand and the signal provides a baseline for arousal. The resting EO state, when the brain is activated with passive visual input and no active stimulus or task processing is required, a baseline for activation is provided, particularly for tasks involving visual processing. 36 The changes observed in the gamma band during the 2 recording conditions indicate the alternation of arousal and neural activation related to cognition associated with a history of opioid use.
The modulation of gamma-band activity has been considered to be related to both the GABAergic interneuron and thalamocortical activities. It has been suggested that when the GABAergic interneuron is activated, the postsynaptic interneuron potentials engage in ongoing mutual inhibition, producing synchronous gamma activity, without the need of the excitatory input from pyramidal cells. 15 Our results could be related to an opioid-induced synchronizing impact of GABAA-mediated inhibitory interneurons. These synchronous inhibitory postsynaptic potentials also disrupt the balance of synaptic excitation from pyramidal cells. An animal study has found that morphine produces burst firing in interneurons. 37 Furthermore, the thalamocortical arousal model suggests that some thalamocortical neurons may be involved in the facilitation of gamma oscillations. For example, a type of pyramidal cell termed chattering cells in the superficial layers of the cat visual cortex induce repetitive bursts at gamma frequencies. 38 Systemically administered opiates inhibit the entire thalamus and induce a shift of cell firing from tonic to bursting. 39 There is also the role of attention and arousal in the thalamocortical generation of gamma synchrony. 15 It has been shown that the power modulation of gamma activity is related to selective attention, whereas the frequency modulation of gamma activity is influenced by the arousal level. 40 The observed difference of gamma activity between groups may be related to tension experienced by subjects during the recording, which is likely to be greater in opiate addicts than healthy volunteers.
Alterations in the activity of the gamma band have been observed over extended areas of the brain in people with a history of opioid use which suggests the opioids induce widespread disruption. However, there are spatial differences between patients undergoing MMT and those currently using illicit opiates. Compared to healthy control subjects, an elevation of activity in the occipital cortex and reduced activity in the parietal and frontal cortices have been observed in those undergoing MMT, while an elevation of activity occurs in the occipital cortex and reduced activity occurs in the parietal, frontal, occipital, and limbic cortices in those using illicit opiates. It appears that regions affected by drug use are less disrupted in those undergoing MMT than those using illicit opiates. Methadone is a long-acting agonist with a half-life of 24 to 36 hours that relieves cravings and negates many of the acute symptoms associated with opiate withdrawal. 41 The pharmacological characteristics of methadone might also contribute to the observed difference in activity. It has been suggested that the deficits associated with drug dependence may not be permanent, and that the deficits induced should not be seen as a permanent “lesion effect” but recoverable limitations of neuroplasticity. 42 Therefore, recovery of brain function following the treatment of people with a history of opiate dependence is likely. The observed changes in the present study may be potential markers of treatment efficacy. Significant improvements in verbal learning, memory, and psychomotor speed have been reported following 2 months of MMT. 43 In our recent study, we also found cognitive performance was improved in those undergoing MMT compared with those still using illicit opiates. 44
There are some limitations of the present investigation that warrant consideration. People with a history of substance dependence have diverse lifestyles, histories of drug taking, histories of overdose, marked variance in nutritional status, and are also often polydrug users. These uncontrolled variables may introduce bias into the current results. Nevertheless, it should be noted that polydrug use is a common factor when undertaking research in those with a history of drug dependence; therefore, it would be exceptionally difficult to recruit people with opiate addiction who have never used other drugs. Furthermore, methadone plasma concentrations were not determined in the present study, which might differentially affect visual evoked responses and EEG recordings. Nevertheless, to the best of our knowledge, no other researchers have reported resting-state EEG activities in the gamma band of patients undergoing MMT. Our findings demonstrate the therapeutic value of substitution therapy for opiate addiction.
Footnotes
Author Contributions
Grace Wang and Bruce Russell designed the study. Grace Wang conducted the statistical analysis and wrote the first draft of the manuscript. Rob Kydd extensively edited and revised the final version of the manuscript. All authors contributed in a significant way to the manuscript and that all authors have read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this study was provided by the New Zealand Pharmacy Education Research Foundation (NZPERF). NZPERF had no further role in the design, analysis or publication of this report.