
Abstract
Breast and cervical cancers remain significant causes of cancer-related deaths in women, necessitating the development of effective treatments. This study investigated the repositioning potential of procaterol as a chemosensitizer to increase the efficacy of cisplatin in HeLa and MCF-7 cells. On its own, procaterol showed limited cytotoxicity; however, it significantly inhibited cell viability when combined with low doses of cisplatin. Both colony formation and cell proliferation studies showed a strong inhibition of cell growth, especially in HeLa cells. Similarly, wound-healing studies showed greater inhibition of cell migration in the presence of both drugs than with either drug alone. Live/dead assays and elevated TNF-α (tumour necrosis factor-alpha) expression confirmed apoptosis. The increased generation of reactive oxygen species suggested that HeLa cells are under oxidative stress. Flow cytometric analyses showed that procaterol induced G0/G1 arrest, cisplatin induced G2/M arrest, and the combination induced both arrests while significantly decreasing S-phase populations. Procaterol sensitized cisplatin by increasing apoptosis and oxidative stress and inducing cell cycle arrest. As an FDA-approved compound, it can be used as a cost-effective adjuvant for treating breast and cervical cancer. Additional in vivo verification and clinical trials are required to establish the translational value of this new combination for the treatment of cervical and breast cancer.
Keywords
Introduction
Breast and cervical cancers are responsible for a significant portion of the world’s female cancer morbidity and mortality worldwide due to their high incidence, metastatic potential, and resistance to traditional therapies (Ramesh and Munjal, 2023). Cervical cancer ranked fourth among female cancers, with approximately 3.1% of all new cases and 56.6% mortality among those diagnosed in women (Kim and Kim, 2022). High-risk human papillomaviruses are the cause of the majority of cervical tumors; yet, in many low- and middle-income nations, vaccination, screening, and treatment are still inconsistent, which contributes to disproportionate mortality (Tanzi and Canuti, 2023). In 2020, female breast cancer accounted for approximately 11.7% of the total new cancer cases in the world, with a death rate of about 29.8% among the diagnosed female population (Sedeta et al., 2023; Shang and Xu, 2022). Risk factors for breast cancer include hormonal imbalance, genetic heterogeneity, and lifestyle factors. Although survival for both cancers has increased due to advancements in imaging, surgery, radiation, and systemic medication, persistent control is still limited by inherent heterogeneity, metastatic spread, and the evolution of drug resistance (Harbeck et al., 2019).
The first line of management of cervical and breast cancer usually consists of surgery, radiation, and stage-appropriate systemic therapy, which may include cytotoxic chemotherapy, targeted medicines, and hormonal therapy (Máté, 2022). Among several medicines, cisplatin, a platinum-based chemotherapeutic agent also known as cis-diamminedichloroplatinum(II), is most frequently used to treat various cancers, including sarcomas, carcinomas, lymphomas, and germ cell tumors. Cisplatin suppresses transcription and replication by forming intra- and interstrand DNA cross-links, which, in turn, cause apoptosis (Srivastava, 2018). However, the development of DNA-repair-mediated resistance and cumulative nephrotoxicity, neurotoxicity, and myelotoxicity limits the therapeutic relevance of cisplatin (Oun et al., 2018). Therefore, current research focuses on rational combination strategies that can increase the anticancer activity of cisplatin while enabling the use of lower, safer dosages.
Drug repurposing offers an effective path for developing combination treatments in oncology with established safety profiles that can rapidly enter cancer trials (Xia et al., 2024). β2-Adrenoceptor agonists, such as procaterol, long proposed for chronic obstructive pulmonary disease and asthma, have recently shown potential against cancer. Specifically, procaterol regulates oxidative stress, inflammation, apoptosis, and proliferation (Cole et al., 2015). It has also been shown in screening studies to be a potential inhibitor of checkpoint kinase-1 (CHK1), a master regulator of S-phase and G2/M checkpoints triggered by DNA damage (Giridhara Prema et al., 2024). CHK1 is a serine/threonine kinase involved in cell cycle arrest and an attractive drug target for anticancer drug development. In gastric cancer models, procaterol and cisplatin act synergistically to inhibit cell proliferation, consistent with CHK1 inhibition.
In this study, we assessed procaterol, a β-adrenoceptor agonist with a newly discovered inhibitory action on CHK1, as an inexpensive cisplatin chemosensitizer for breast and cervical cancer. We hypothesized that co-administering the drug would increase the effectiveness of cisplatin by enhancing apoptotic signaling, increasing reactive oxygen species (ROS) generation, and enforcing cell-cycle arrest. Therefore, in a series of in vitro tests, we evaluated the cytotoxic, antimigratory, and pro-apoptotic effects of single-agent versus combination treatment using HeLa (cervical) and MCF-7 (breast) cells. This study offers a feasible strategy to overcome resistance in cervical and breast cancer treatment, while also providing molecular evidence that procaterol increases the effectiveness of cisplatin.
Materials and Methods
Chemicals
Procaterol was procured from Sigma-Aldrich, USA; Dulbecco’s modified Eagle media (DMEM), fetal bovine serum (FBS), and 100× antibiotic/antimycotic solution from Gibco, Thermo Fisher Scientific USA, Cisplatin (Celon labs), DMSO (MP Biomedicals), MTT (Himedia), Crystal violet, HOECHST-33342, Acridine Orange (AO), ethidium bromide (EB), and 2′, 7′-dichlorofluorescein diacetate (DCFDA) from Sigma-Aldrich, USA. BCA (bicinchoninic acid) assay kit from Thermo Fisher Scientific, Massachusetts, USA. Antibodies were obtained from Cell Signaling Technology, Danvers, USA. Nitrocellulose membrane and ECL substrate were obtained from BioRad Laboratories, USA.
Cell culture
HeLa and MCF-7 cells were procured from the National Centre for Cell Science (NCCS), Pune, India. The cells were cultured in DMEM high-glucose medium supplemented with 10% FBS and incubated at 37°C and 5% CO2 in a humidified atmosphere.
Cell cytotoxicity assay
HeLa and MCF-7 cells were seeded at 5000 cells/well in a 96-well plate. The cells were exposed to various concentrations of positive control cisplatin, procaterol, and in combination (procaterol and cisplatin). The cytotoxicity study was carried out using the MTT assay in accordance with the protocol described in Suchitha et al. (G P et al., 2025) The HeLa cells were subjected to various concentrations of cisplatin (1–6 µg/mL) and procaterol (2–12 µg/mL) after a 24 h incubation period. MCF-7 cells were also exposed to varying concentrations of cisplatin (1–6 µg/mL) and procaterol (1–9 µg/mL). For the combination therapy, the IC50 of procaterol and varying concentrations of cisplatin (1–6 μg/mL) were used to evaluate whether procaterol would potentiate the anticancer activity of cisplatin, thereby allowing a reduction in cisplatin concentration and identifying potential synergistic effects. After 48 h of incubation, MTT dye was added, and the cells were incubated for 3 h at 37°C. The mixture of ethanol and DMSO (1:1) was used to dissolve the formazan crystals. Absorbance was measured at 570 nm and 650 nm using the multimode microplate reader (Multiskan Sky, Thermo Fisher Scientific). Cell viability and IC50 were determined compared with the untreated control, which was then used in subsequent experiments.
Combination index analysis
The Chou–Talalay methodology was used to investigate the interaction between procaterol and cisplatin quantitatively. Cell viability was used to determine Fa, and R software was used to calculate the combination index (CI) using the median-effect model. The following is how CI is interpreted: an additive effect is recognized if CI equals 1, antagonism is shown if CI is greater than 1, and synergism is observed if CI is less than 1.
Colony formation assay
HeLa and MCF-7 cells were seeded at a density of 500 cells/well in a six-well plate and treated with cisplatin, procaterol, and the combination (procaterol and cisplatin). The cells were cultured for 8–10 days to allow for growth. Following methanol fixation (Sigma-Aldrich, St. Louis), the colonies were stained with 1% crystal violet, and images were taken with Magvision Keer System. ImageJ Software (National Institute of Health, Bethesda, USA) was used to quantify the number of colonies (G P et al., 2025).
Wound healing scratch assay
HeLa and MCF-7 cells were seeded at a density of 4 × 105 cells/well in a six-well plate. After 85% confluence, a 200 µL pipette tip was used to create a scratch. The cells were treated with cisplatin, procaterol, and both in combination. Images were captured at 0, 6, 24, and 48 h using an inverted microscope (PrimoVert, Carl Zeiss, Germany). The migration distance was calculated using ImageJ software (National Institute of Health, Bethesda, USA) (G P et al., 2025).
Cell proliferation assay
In a six-well plate, HeLa and MCF-7 cells were seeded at a density of 3 × 103 cells/well. After 24 h of incubation, the cells were treated with cisplatin, procaterol, and in combination for 48 h. Following the removal of media, the cells were washed with PBS and stained with 3% crystal violet in 50% methanol for an hour at room temperature. To remove excess stains, the well was subsequently rinsed with MilliQ water. The plate was allowed to air dry, and images of stained cells were captured with an inverted microscope (Carl Zeiss Microscopy, Germany) at 4× (G P et al., 2025).
Live–dead assay
HeLa and MCF-7 cells were seeded at a density of 2 × 105 cells/well in a six-well plate and treated with cisplatin, procaterol, and in combination (procaterol and cisplatin). After 48 h of incubation, the media was removed, and the cells were stained with AO and EB prepared in serum-free media and incubated for 15 min in the dark at 37°C. Cells were imaged using ZOE™ Fluorescent Cell Imager (Biorad Laboratories, California, USA). The ratio of AO to EB-stained cells was calculated using ImageJ, NIH software (G P et al., 2025).
Assessment of ROS
HeLa and MCF-7 cells were seeded at a density of 2 × 105 cells/well in a six-well plate and treated with cisplatin, procaterol, and in combination (procaterol and cisplatin). After incubation for 48 h, the cells were stained with 5 µg/mL Hoechst-33342 and 25 µM DCFDA in serum-free media and incubated for 15 min in the dark at 37°C to detect intracellular ROS. ZOE Fluorescent Cell Imager was used to take images. Results were expressed as a percentage of ROS compared with the control (Giridhara Prema et al., 2024).
Cell cycle analysis
HeLa and MCF-7 cells were seeded at a density of 2 × 105 cells/well and treated with cisplatin, procaterol, and in combination. The cells were trypsinized and washed twice with 1× PBS. The resultant cell pellet was resuspended in hypertonic lysis buffer (1 mg/mL trisodium citrate, 0.1% Triton-X 100, 100 μg/mL RNase, 2 μg/mL of PI) and incubated in the dark for 20 min. Cells were immediately measured for red fluorescence using the Guava® easyCyte Flow cytometer, EMD Millipore, Massachusetts, USA (Giridhara Prema et al., 2024).
Western blotting
HeLa and MCF-7 cells were plated at a density of 1 × 106 cells in a 10 cm dish and incubated with cisplatin, procaterol, and in combination for 48 h. After cell harvest, proteins were isolated and estimated using the BCA method with BSA (bovine serum albumin) as a standard. The expressions of β-actin and TNF-α (tumour necrosis factor-alpha) were estimated using western blotting. Proteins were transferred from the SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis) gel to a nitrocellulose membrane, which was then blocked with 5% skimmed milk for 1 h. Membranes were incubated overnight at 4°C using primary antibodies with 3% BSA. Membranes were then incubated with respective secondary antibodies at room temperature for 2 h. Immunoreactive bands were developed in a gel documentation system using an enhanced chemiluminescent reagent. ImageJ software was used for densitometry analysis of band intensity; normalization was performed using a loading control, and fold change was calculated relative to control cells.
Statistical analysis
All experiments were conducted in biological triplicate, and data are presented as mean ± SEM (standard error of the mean). One-way ANOVA and GraphPad Prism software were used for data analysis, and p-values <0.05 were used to identify statistically significant results.
Results
Cell cytotoxicity assay
MTT assay demonstrated the cytotoxicity effect of procaterol in both HeLa and MCF-7 cells. In HeLa cells, procaterol reduced the cell viability at 8 µg/mL (IC50), whereas in MCF-7, the IC50 obtained at 7 µg/mL. Conversely, cisplatin had a marked effect on reducing cell viability, with an IC50 of 4 µg/mL in both cell lines. Significantly, cotreatment with procaterol (8 µg/mL) plus cisplatin (2 µg/mL) in HeLa cells increased cytotoxicity. Similarly, in MCF-7, procaterol (7 µg/mL) plus cisplatin (4 µg/mL) further enhanced cytotoxicity, consistent with a synergistic interaction. In combination with procaterol (8 µg/mL), even a concentration half the IC50 of cisplatin (2 µg/mL) produced about 50% cytotoxicity in HeLa cells, highlighting procaterol’s chemosensitizing ability. This finding suggests that therapeutic anticancer activity can be achieved at substantially lower cisplatin doses, potentially reducing dose-limiting toxicities while maintaining therapeutic efficacy (Fig. 1).
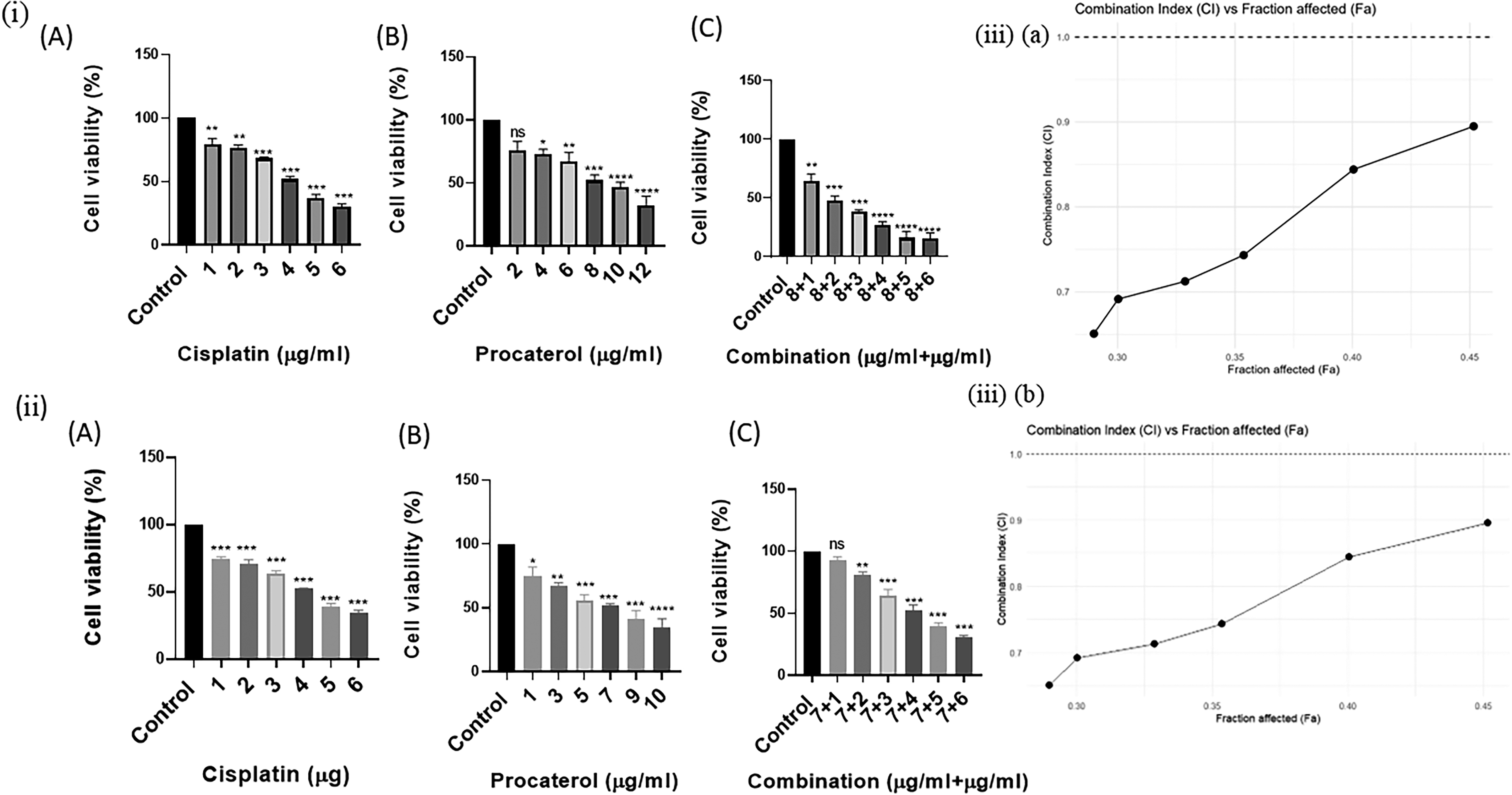
Combination index analysis
The type of interaction between procaterol and cisplatin was studied using a CI test. It was observed that the CI values obtained for different Fa concentrations in HeLa and MCF-7 cells were below 1, thus proving a synergistic interaction between the two compounds. Moreover, the CI value showed that the synergism between the two drugs increased with increasing Fa values, suggesting greater efficacy of the combination at higher levels of inhibition. Furthermore, the synergistic effect was comparatively stronger in HeLa cells than in MCF-7 cells, indicating a cell line-dependent response to the combination treatment (Fig. 1).
Colony-forming assay
The colony-forming assay was performed to determine the long-term proliferative potential of HeLa and MCF-7 cells treated with procaterol, cisplatin, or both. In HeLa and MCF-7 cells, procaterol treatment reduced colony number and size in both cell lines, but the reduction was more pronounced in HeLa cells, indicating greater cervical cancer cell sensitivity to procaterol-mediated inhibition. Cisplatin treatment also inhibited colony formation to a significant level in both cell lines, demonstrating its strong cytotoxic activity. Combination therapy elicited the greatest inhibition, with over 80–90% reduction in colony count. The observed synergistic inhibition of colony growth demonstrates that procaterol enhances the cytotoxicity of cisplatin, suggesting its potential as an adjuvant in the treatment of cervical and breast cancers (Fig. 2).
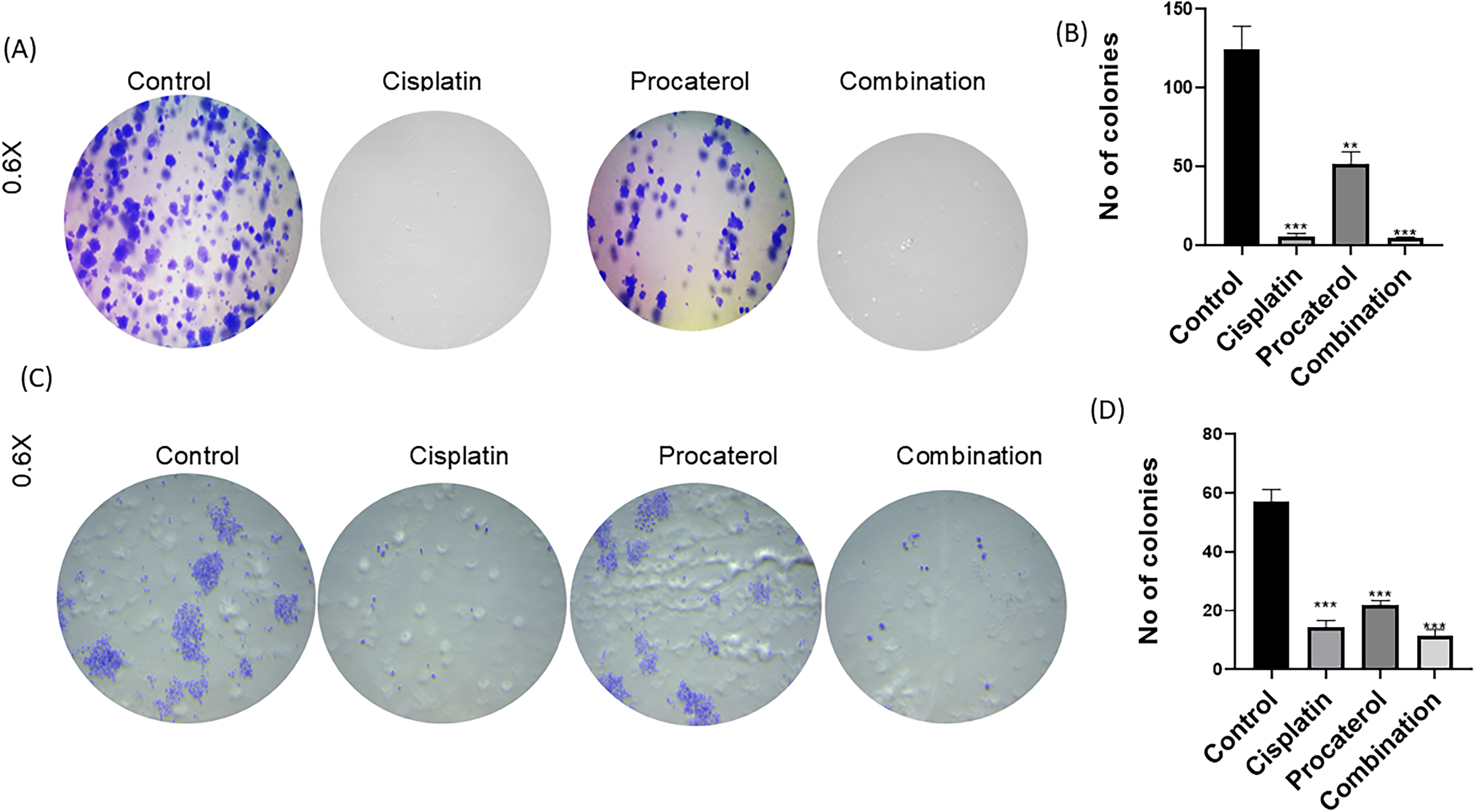
Procaterol and combination (procaterol + cisplatin) decrease colony formation on HeLa and MCF-7 cells.
Wound healing scratch assay
A wound-healing scratch assay was performed to measure the effects of cisplatin, procaterol and their combination on the migration of MCF-7 and HeLa cells. The wound closure rate was measured at 0, 6, 24, and 48 h in MCF-7 and HeLa cells; the control sample showed rapid wound closure, with cells migrating toward the scratched area over time. Procaterol treatment alone caused mild inhibition of wound closure when compared with the control. In HeLa cells, at 48 h, total wound closure was observed compared with the control. Both cell lines showed reduced migration in cisplatin-treated cells. Procaterol and cisplatin combination inhibited migration, with minimal wound closure observed in both HeLa and MCF-7 cells. The scratch remains largely unclosed even after the full incubation period, indicating a synergistic effect of the combination in inhibiting cell migration. The percentage wound closure was quantified to determine the effect of treatment on cancer cell migration (Fig. 3).
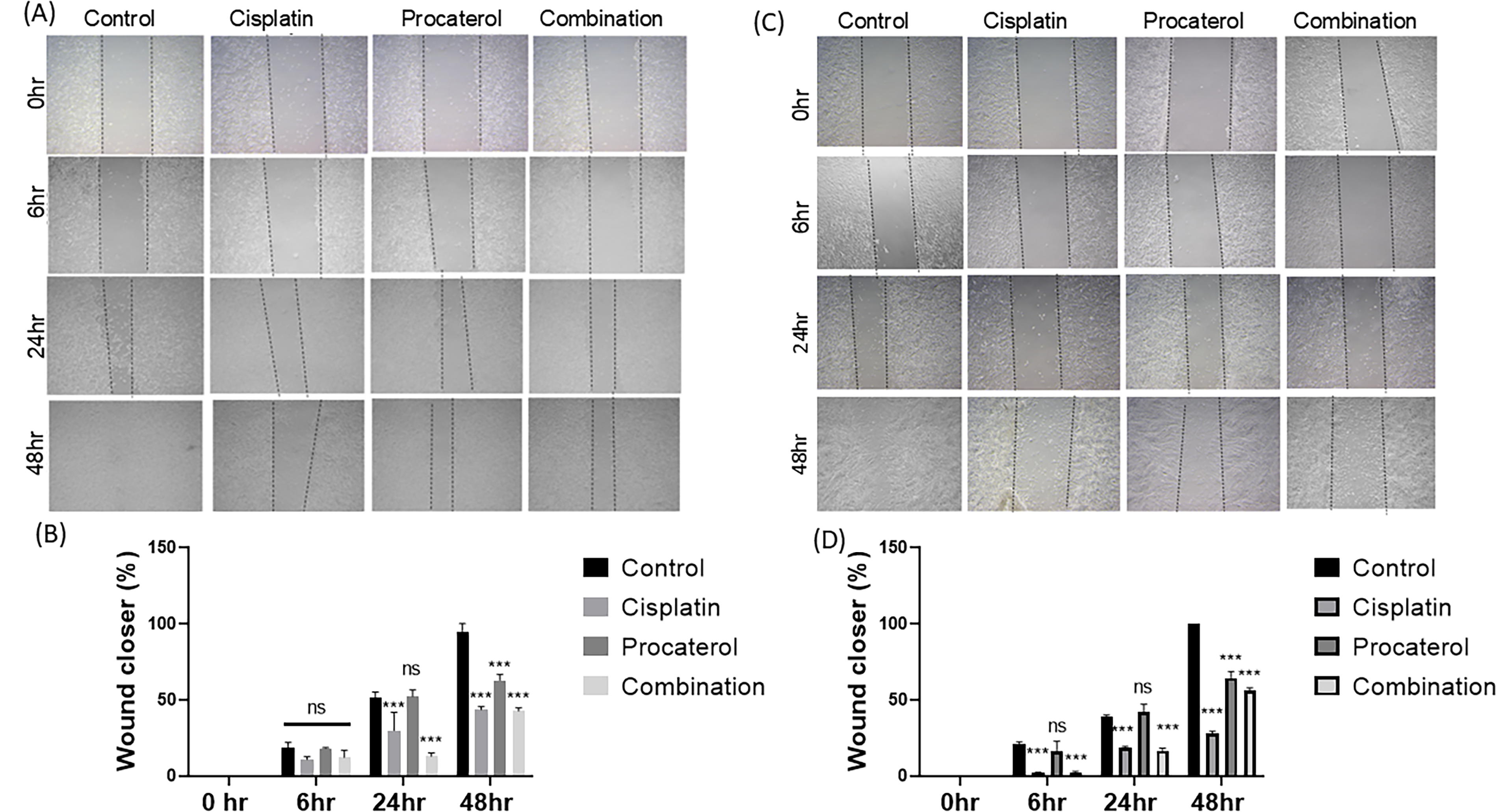
Procaterol and combination (procaterol + cisplatin) decrease colony migration ability of HeLa and MCF-7 cells.
Cell proliferation assay
The control groups of both cell lines showed sustained proliferation, indicating unrestricted growth potential. Procaterol alone caused a modest decrease in proliferation compared with the control group. The reduction was greater in HeLa cells, indicating greater sensitivity of cervical cancer cells to procaterol. Significant inhibition in cell growth was observed in cisplatin-treated cells for both cell lines. The combination treatment showed significant inhibition, as evidenced by a reduction in cell number over time. The inhibitory effect was greater in HeLa cells, resulting in almost complete suppression of proliferation (Fig. 4). The number of proliferated cells was quantified to determine the effect of treatment on cancer cell viability and proliferation.
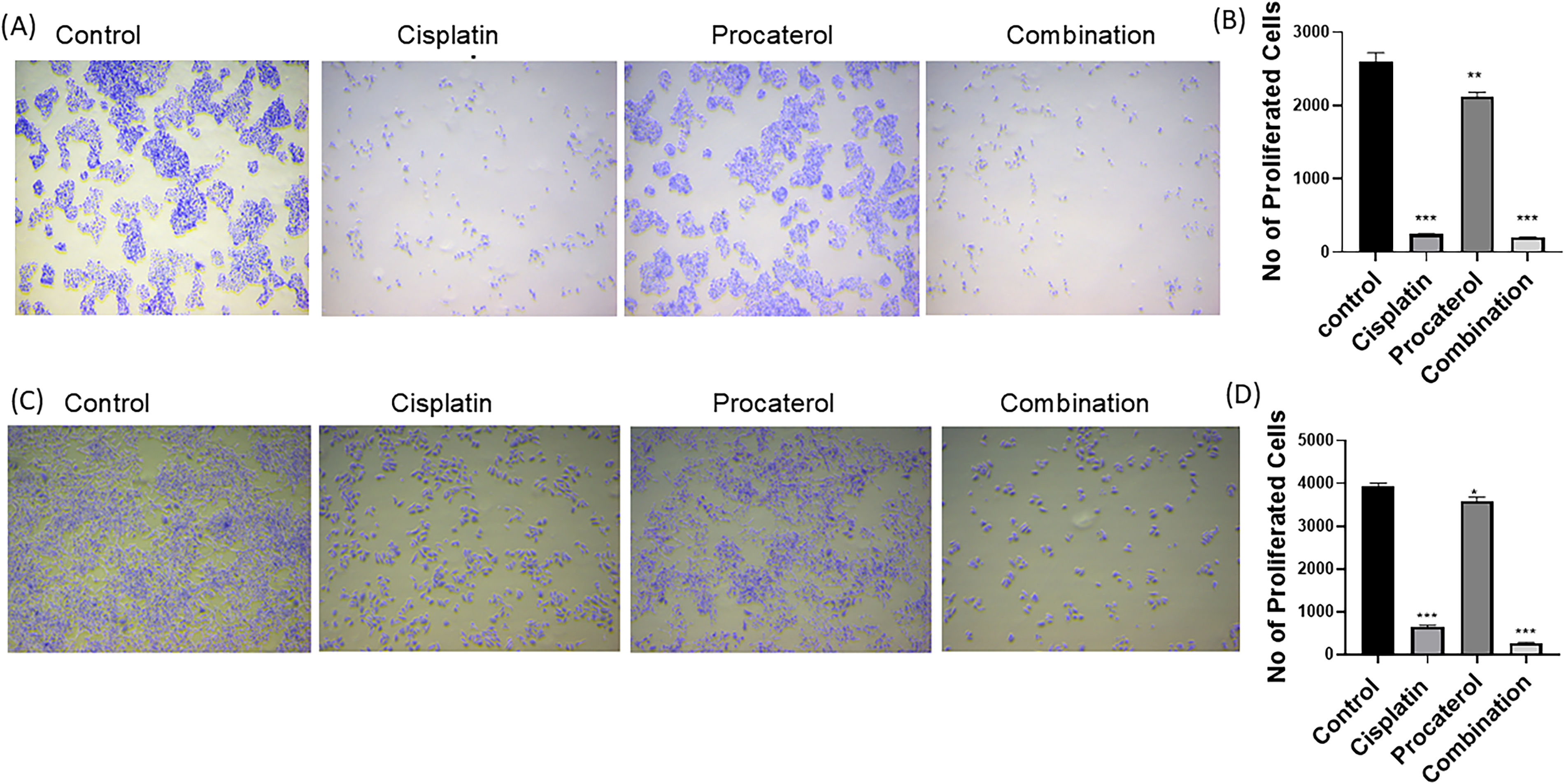
Procaterol and combination (procaterol + cisplatin) treatment reduces cellular proliferation in HeLa and MCF-7 cells.
Live–dead assay
Live–dead assay showed a significant decrease in the viability of HeLa and MCF-7 cells after treatment with the drugs alone or in combination. In the control group, the HeLa and MCF-7 cells exhibited predominantly green fluorescence, indicating cell viability. Treatment with procaterol alone created a higher red fluorescence signal, reflecting partial loss of viability, which correlated with moderate upregulation of TNF-α expression, confirming its role in apoptosis induction. Cisplatin therapy further increased red fluorescence, representative of increased cytotoxicity and a significant increase in TNF-α expression, validating its involvement in cisplatin-induced apoptosis. A combination of cisplatin and procaterol showed maximum cytotoxicity as evidenced by predominant red fluorescence and reduced green fluorescence, representing synergistic toxicity. This combination also showed elevated TNF-α expression. HeLa cells showed slightly elevated TNF-α expression compared with MCF-7 cells, suggesting greater sensitivity to TNF-α-mediated apoptosis. Taken together, these findings suggest that procaterol increases cisplatin-induced apoptosis by upregulating TNF-α expression, rendering the cells more vulnerable to chemotherapeutic-induced cell death (Fig. 5).
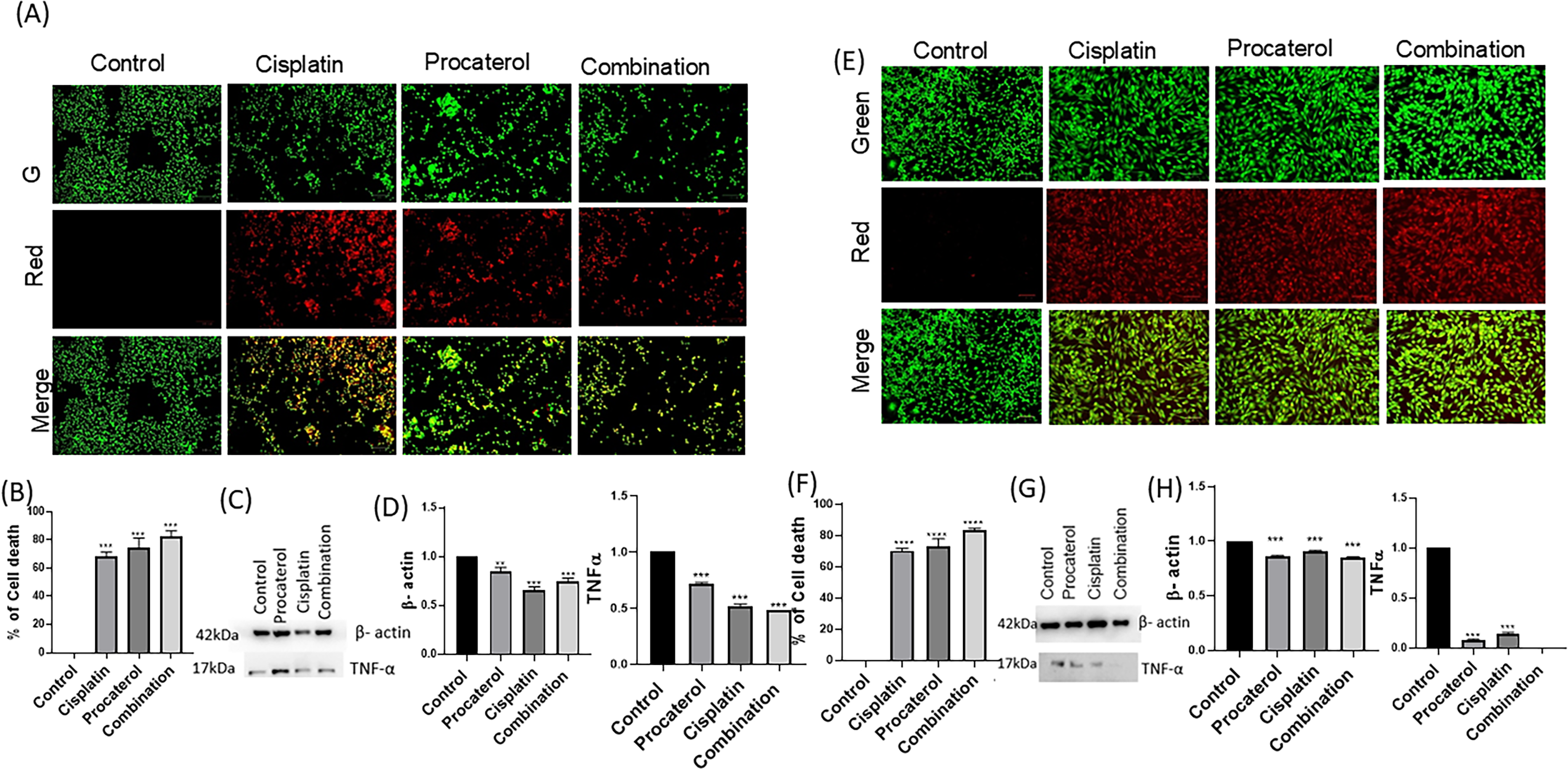
Procaterol and combination (procaterol + cisplatin) treatment induce apoptosis in HeLa cells.
Assessment of ROS
A ROS assay was used to assess oxidative stress levels in HeLa and MCF-7 cells following treatment. In the control group, minimal fluorescence intensity indicated low ROS production, suggesting that untreated cells maintained normal oxidative stability. Procaterol treatment slightly elevated ROS levels, as indicated by increased fluorescence, particularly in HeLa cells, suggesting a role in oxidative stress generation. Compared with procaterol treatment alone, cisplatin treatment led to a marked increase in ROS levels in both cell lines, suggesting its involvement in oxidative stress-induced cytotoxicity. Combination therapy achieved the highest ROS levels in both cell lines, indicating synergistic induction of oxidative stress. Our findings revealed that ROS accumulation was significantly higher in HeLa cells than in MCF-7 cells, suggesting greater sensitivity of cervical cancer cells to oxidative stress-induced injury (Fig. 6).
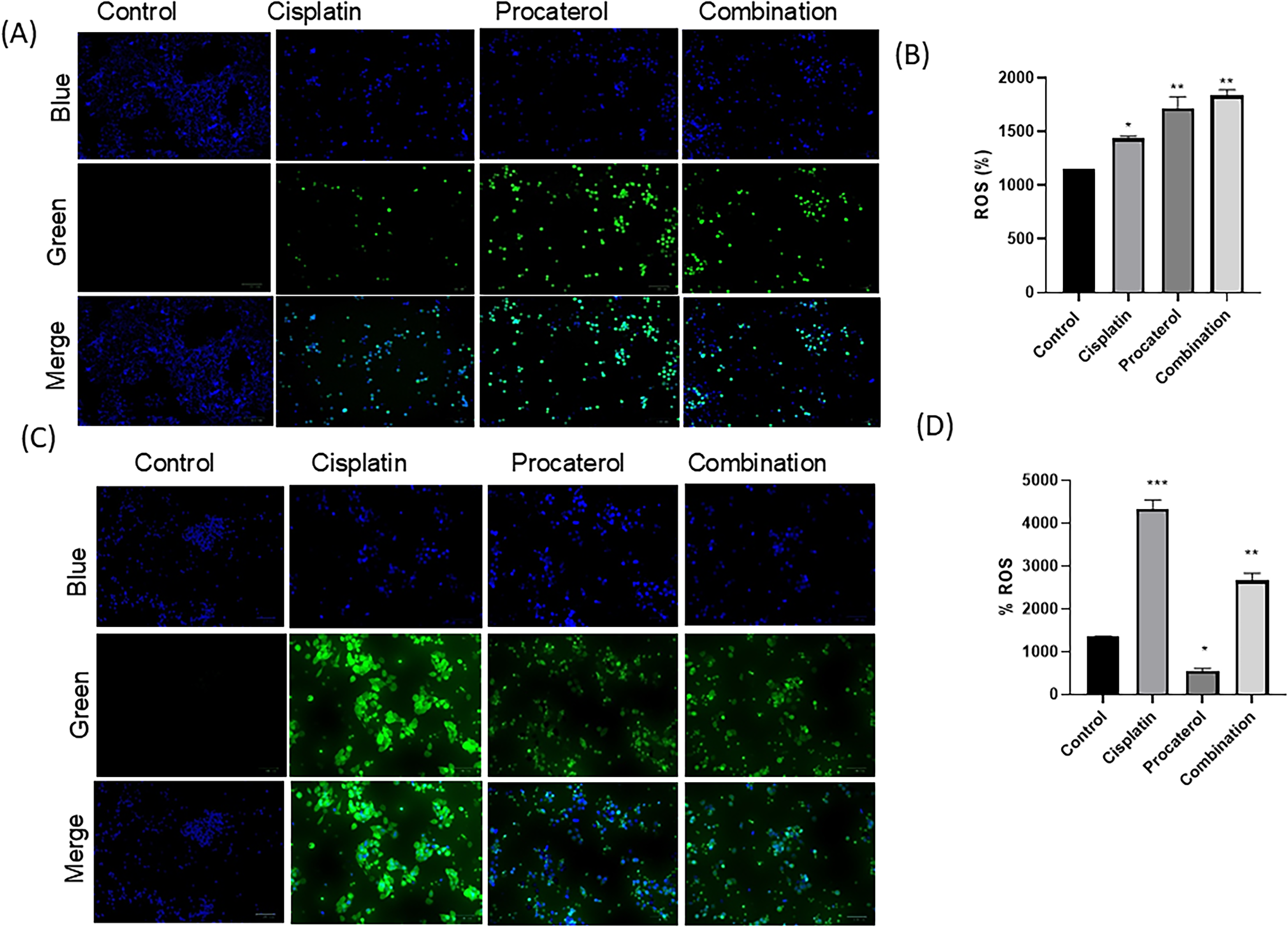
Combination (procaterol + cisplatin) treatment induces reactive oxygen species (ROS) in HeLa cells
Cell cycle analysis
Flow cytometry was performed to assess cell-cycle distribution. Both HeLa and MCF-7 cells in the control group had a normal cell cycle distribution, with the majority of cells in the G0/G1 phase, indicating active proliferation. Procaterol treatment led to a moderate increase in the G0/G1 population, followed by a decrease in the S phase, suggesting inhibition of cell cycle progression. HeLa cells were rendered more sensitive to procaterol-induced G0/G1 cell cycle arrest. Cisplatin treatment, on the other hand, resulted in a significant increase in the G2/M population, indicative of G2/M checkpoint arrest because of DNA damage and delayed mitotic progression. While this effect was observed in both the cells, MCF-7 cells demonstrated slightly higher G2/M arrest, indicating greater sensitivity to cisplatin-induced DNA damage. The combination treatment induced a two-phase arrest, resulting in extensive cell accumulation at G0/G1 and G2/M phases, along with inhibition of S-phase progression, indicating enhanced inhibition of cell cycle progression through synergistic action (Fig. 7).
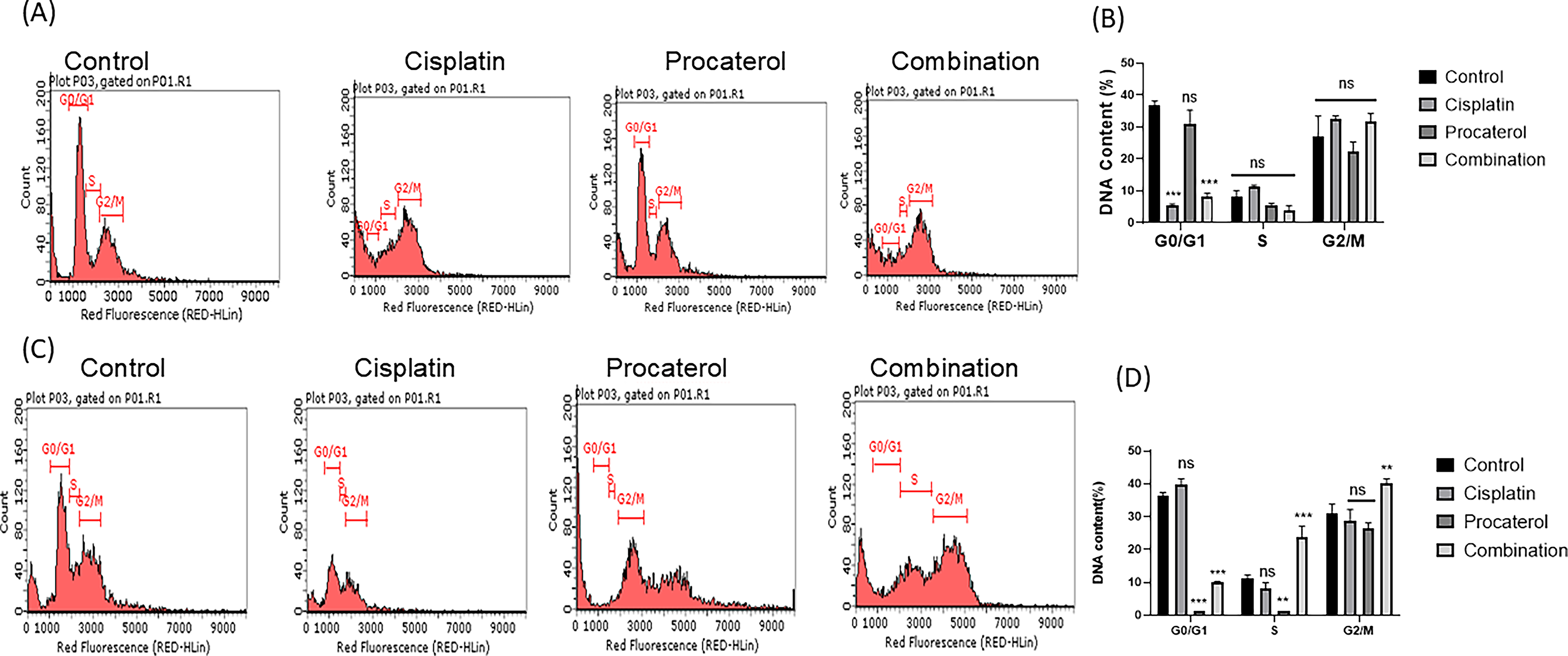
Discussion
Breast and cervical cancers remain a significant global health burden, with epidemiological data highlighting their unequal burden among women globally. Based on the American Cancer Society Cancer Statistics 2023; (Siegel et al., 2023) and GLOBOCAN 2020 estimates (Sung et al., 2021), breast cancer is now the most common malignancy diagnosed in women, representing 11.7% of all new cases of cancer worldwide, while cervical cancer is fourth among gynecologic cancers. They are a significant proportion of all cancer morbidity and mortality among women. In high-income nations, improvements in imaging, screening, surgery, radiotherapy, and systemic therapies have increased survival rates; however, in low- and middle-income areas, where access to comprehensive treatment options, early detection, and vaccination remains restricted, disparities persist (Sung et al., 2021; Bray et al., 2021; Holohan et al., 2013). Significantly, Bray et al. reported that cancer is increasingly becoming a major cause of premature death globally, with breast and cervical cancers playing a major role in this regard (Bray et al., 2021). These epidemiological facts highlight the urgent need for accessible and efficient therapeutic interventions to address global inequalities in cancer outcomes.
In this regard, drug repurposing, the process of discovering novel therapeutic applications of existing drugs, has been identified as a cost-effective, time-saving measure to speed up the treatment development (Burrell and Swanton, 2014). Specifically, the combination of conventional chemotherapeutics with β2-adrenoceptor agonists or oxidative stress inducers has been of interest as a means to enhance therapeutic gain without augmenting systemic toxicities (Xia et al., 2024; Cole et al., 2015). This is relevant to breast and cervical cancer, where chemoresistance and dose-limiting side effects of first-line drugs like cisplatin continue to be a problem for patients’ quality of life and survival. Furthermore, as metastasis remains the main cause of cancer-related death (Steeg, 2016), and disparities in access to treatment are also increasing (Prager et al., 2018), there is an urgent need for inexpensive interventions that can effectively be integrated into the diverse healthcare systems. Despite the success of modern targeted treatments, therapy resistance remains a major hurdle, particularly in highly aggressive neoplasms such as triple-negative breast cancer (Lev, 2020). Within this framework, drug repurposing represents a viable solution to overcome these therapeutic limitations (Pushpakom et al., 2019).
Besides being cost-effective, the biological rationale for drug repurposing has been strengthened by emerging insights into stress-related signaling pathways. β-Adrenergic signaling, for instance, has been recognized in tumor proliferation, in which chronic stress may lead to tumor initiation and angiogenesis (Thaker et al., 2006). In an interesting turn of events, recent evidence suggests that β2-adrenoceptor agonists, thus far used as bronchodilators, may be repurposed as anticancer agents. Notably, the potential to circumvent drug resistance and access cancer stem cells is key to maximizing therapeutic responses, and repurposed drugs that leverage these mechanisms hold great promise (Moitra, 2015). Recent experimental and in silico analyses have identified procaterol, a β2-adrenoceptor agonist, as a novel CHK1 inhibitor, a key mediator of DNA repair and chemoresistance (Qiu et al., 2018). (Liu et al., 2020) demonstrated that procaterol synergizes with cisplatin in gastric cancer cells by augmenting DNA damage, triggering apoptosis, and suppressing tumor growth by inhibiting CHK1 (Liu et al., 2020). This finding provides strong preclinical evidence that, in combination with cisplatin, procaterol can be a repurposed therapy capable of overcoming resistance and improving treatment response in cervical and breast cancers. However, the role of CHK1 in the current study is hypothetical, as no direct experimental validation was performed. Further studies are required to confirm the role of CHK1.
In the current study, we examined the interaction between cisplatin and procaterol, a β2-adrenoceptor agonist, on HeLa and MCF-7 cells. Our results revealed that procaterol significantly enhances the anticancer efficacy of cisplatin, resulting in increased cytotoxicity, apoptosis, oxidative stress, and cell cycle arrest. These results confirm and reinforce previous research on cisplatin combination drugs and underpin the prospects of procaterol as a repurposed anticancer agent. Interestingly, the combination therapy showed synergistic effects, with procaterol augmenting the efficacy of cisplatin and reducing inflammation. Compared with single-agent therapies, cotreatment with procaterol and cisplatin resulted in substantial reductions in cell viability and increased cell death in both cell lines. (Yoshida et al., 2009) reported that procaterol enhanced the anti-inflammatory effects of budesonide and provided better control of asthma when used in combination than when used individually (Yoshida et al., 2009). Procaterol peak plasma concentration (Cmax) ranges from 136 to 358 pg/mL (about 0.14–0.36 ng/mL) following oral ingestion of 50 µg of the drug, according to a pharmacokinetic study. Even lower levels near the quantitative limit are observed when procaterol is inhaled. This suggests that therapeutic doses of procaterol may result in sub-nanomolar plasma concentrations (Eldon et al., 1993; Kobayashi et al., 2010; Krogh et al., 2016). Whereas in the present study, procaterol was given to HeLa cells at 7 µg/mL and MCF-7 cells at 8 µg/mL (≈7000–8000 ng/mL). This is about four orders of magnitude higher than the plasma concentrations achievable in a clinic. This is a study limit in terms of clinical scenarios. However, it is widely accepted that higher drug concentrations are required to demonstrate a significant biological effect, since in vitro systems lack pharmacokinetic factors such as metabolism, tissue distribution, and plasma protein binding. To determine a dosage close to the IC50 required to exhibit a significant phenotypic effect in the cell line of interest, preparatory dose–response tests were used to determine the concentrations of the medications administered in the current investigation. However, more in vivo research would be required to determine whether cisplatin-like effects can be achieved.
β2-Adrenoceptor agonist procaterol enhances the therapeutic activity of cisplatin in cervical cancer cells, supporting the growing evidence for drug repurposing as a cost-effective and valuable strategy to improve cancer therapy. Previous reports have shown that β2-adrenoceptor agonists are cisplatin sensitizers. For instance, Bando et al. reported that procaterol significantly increased cisplatin accumulation by NSCLC cells, thereby enhancing cytotoxicity (Bando et al., 2000). Similarly, (Liu et al., 2020) and (Matsumoto et al., 2016) showed that β2-adrenergic receptor agonists potentiated cisplatin-induced apoptosis in NSCLC cells (Matsumoto et al., 2016). Our findings are consistent with recent literature emphasizing the therapeutic potential of combination therapies to counteract chemoresistance.
Rahimifard et al. also demonstrated that resveratrol combined with cisplatin induced gastric cancer cell senescence and apoptosis, highlighting oxidative stress and cell cycle arrest as key mechanisms (Rahimifard et al., 2022). Also, Ren et al. confirmed that cisplatin and resveratrol cotreatment triggered apoptosis by endoplasmic reticulum stress and G2/M arrest, providing additional evidence that combination regimens increase cisplatin efficacy through triggering stress-mediated apoptotic pathways (Ren et al., 2020). The cotreatment of cisplatin with AKT inhibitors greatly increases cytotoxicity in several cancer models, in the direction of PI3K/AKT signaling inhibition-mediated sensitization (Tao et al., 2016). In the present study, enhanced cytotoxicity was associated with high TNF-α expression, which correlated with increased apoptotic activity. This concurs with earlier research demonstrating that combination therapies strengthen apoptotic signaling, thereby sensitizing cancer cells to chemotherapy (Moitra, 2015). TNF-α is also a major activator of the extrinsic pathway of apoptosis, and its overexpression has been linked to caspase activation and enhanced cell death in cells treated with chemotherapeutic agents (Qiu et al., 2018). Interestingly, we also observed increased ROS production in HeLa cells treated with procaterol in combination, suggesting that oxidative stress is a major mechanism underlying therapeutic synergy. This observation is consistent with Rahimifard et al. (2022), who reported that cotreatment of gastric cancer cells with cisplatin and resveratrol increased ROS generation and cytotoxicity (Matsumoto et al., 2016). Differential responses to ROS occur among different cancer cell types. This difference highlights the complexity of oxidative stress regulation, and it may influence how cancer cells respond to combination treatments. Cell cycle analysis revealed procaterol-induced G0/G1 phase arrest, while cisplatin caused G2/M phase arrest. Notably, combination treatment caused dual-phase arrest with severe repression of the S phase, reflecting a cumulative inhibition of cell cycle progression. Such checkpoint interference is in line with findings that CHK1 inhibition potentiates cisplatin-induced DNA damage responses to cause enhanced killing of tumor cells (Xing et al., 2025). Recent evidence indicates that β2-adrenergic receptor activation regulates CHK1-dependent DNA damage signaling (Patil et al., 2013), and a computational model has identified procaterol as a potential CHK1 inhibitor (Mravec et al., 2020). This mechanistic overlap supports the hypothesis that procaterol’s inhibition of CHK1 is a mechanism by which it enhances its synergism with cisplatin, as demonstrated by (Tao et al., 2016), who showed that CHK1 inhibition enhanced cisplatin-induced checkpoint arrest and the DNA damage response (Tao et al., 2016). However, it still requires further experimental validation. The clinical relevance of our findings is the growing recognition of drug repurposing as a viable strategy for accelerating cancer drug development. Repurposed drugs possess well-established pharmacokinetic and safety profiles, minimizing both time and cost for clinical translation (Ioakeim-Skoufa et al., 2023). The concentration of procaterol employed in this study is higher than clinically feasible plasma levels, which limits its direct translational relevance. To confirm whether comparable effects can be achieved, more in vivo and pharmacokinetic research will be required. Moreover, using a single cell line per cancer subtype limits the scope of the results. Our data indicate that procaterol acts as an effective chemosensitizer, increasing cisplatin-induced apoptosis, ROS production, and cell cycle arrest, all of which are important for circumventing resistance mechanisms in cervical and breast cancers. In addition, emerging clinical evidence emphasizes the role of combination approaches in cancer treatment. (Gupta et al., 2018) showed that neoadjuvant chemotherapy and subsequent chemoradiation decreased mortality by 40% and recurrence by 35% at 5 years in patients with cervical cancer (Gupta et al., 2018).Collectively, these results underscore the therapeutic potential of repurposed drugs, such as procaterol, in combination regimens to improve treatment outcomes and overall survival. The present study identifies procaterol as an effective chemosensitizer that synergizes with cisplatin to increase apoptosis, oxidative stress, and cell cycle arrest, thereby enhancing its anticancer activity. Overall, the present findings provide preliminary insights, however, validation through in vivo studies, and detailed mechanistic investigation is required to establish the translational approach.
Conclusion
The present research highlights the potential of procaterol, a selective β2-adrenoceptor agonist, as an effective chemosensitizer that may modulate response in combination with cisplatin for the treatment of cervical and breast cancer. Although procaterol alone didn’t significantly affect cancer cell viability, its combination with cisplatin effectively augmented cytotoxic effects, as evidenced by enhanced apoptosis, increased ROS, and cell cycle arrest. The upregulation of TNF-α also underscores procaterol’s role in enhancing extrinsic apoptosis, contributing to the observed synergistic effect. Together, these results indicate that procaterol can efficiently enhance the anticancer efficacy of cisplatin, suggesting its potential to overcome drug resistance and reduce required drug doses, thereby minimizing drug-associated toxicities. With procaterol’s proven clinical safety profile, its repurposing presents an efficient and economical means of enhancing current chemotherapeutic regimens. Overall, this study is solely based on in vitro experiments; additional molecular analyses, in vivo work, and clinical trials are needed to confirm these findings and further investigate the therapeutic potential of this combination in the clinical treatment of cervical and breast cancer.
Authors’ Contributions
N.S.: Performed the experiments, analysis, and article drafting; G.P.S.: performed the experiments and analysis; T.S.K.P.: Conceptualized the study. S.D.: Conceptualized the study, designed the components, analyzed the data, and reviewed/corrected the article. All authors read and approved the final article.
Footnotes
Acknowledgments
The authors acknowledge the Department of Biotechnology National Facility grant for supporting the Center for Systems Biology and Molecular Medicine at Yenepoya (Deemed to be University), under the project “Skill Development in Mass Spectrometry-based Metabolomics Technology BIC” (BT/PR40202/BTIS/137/53/2023). The authors acknowledge the use of ChatGPT (OpenAI) to improve the English language and grammar of the article.
Author Disclosure Statement
The authors declare that there is no conflict of interest.
Funding Information
The authors received no specific funding for this work.