
Abstract
Tic disorder (TD) is a common neurodevelopmental disorder in children. Acute histamine deficiency may be involved in the pathogenesis of TDs. In this study, we demonstrate that histamine depletion exacerbates tic-like behaviors in a rat model of TD. Sprague–Dawley rats were divided into control, TD model, TD plus saline, and TD plus α-fluoromethylhistidine (α-FMH) groups. TD was induced by intraperitoneal iminodipropionitrile, and α-FMH was administered intracerebroventricularly. Behavioral assessments showed that α-FMH treatment significantly increased stereotyped and motor behavior scores compared with TD groups (p < 0.05). Immunohistochemistry revealed marked microglial activation in the striatum of TD groups, which was exacerbated by histamine deficiency. Striatal IL-6 and TNF-α levels were elevated in all TD groups; IL-6 was significantly higher in the α-FMH group than in TD groups (p < 0.05). Striatal dopamine receptor D2 (DRD2) expression was also upregulated in TD groups, with the highest levels observed after histamine depletion (p < 0.05). Liquid chromatography-tandem mass spectrometry result showed that glutamate levels were significantly reduced in the α-FMH group compared to the TD-NaCl group (p < 0.05). No significant differences were observed in the levels of dopamine, serotonin, or acetylcholine between the TD-NaCl and TD-α-FMH groups. Overall, our findings suggest that acute histamine deficiency aggravates TD-related behaviors by promoting neuroinflammation and upregulating DRD2 expression, highlighting the therapeutic potential of targeting histamine signaling in TD.
Introduction
Tic disorder (TD) is a common neurodevelopmental disorder in children. The prevalence of TD in the general population has been reported to range from 0.04% to 0.16% (Yilmaz and Jankovic, 2025). Tourette syndrome (TS) is a severe form of TD, typically emerging before the age of 18 and with a significantly higher incidence in males than in females. TD substantially affects children’s learning, daily life, psychological well-being, and social adaptation, and its clinical management remains challenging because the pathogenesis of TD is unclear (Frey, 2025).
An increasing number of studies have emphasized the role of immunological involvement in TD. The association between TD and group A Streptococcus (GAS) infection has been recognized for decades. In a cross-sectional analysis, patients with TS exhibited a higher frequency of GAS infection, elevated anti-streptolysin O titers, and a higher frequency of anti-basal ganglia antibodies compared to the control group (Martino et al., 2011). In addition, a clinical entity characterized by the acute onset of tics, accompanied by obsessive–compulsive symptoms, behavioral changes, and personality regression in children following streptococcal infection has been described over the past few decades. This condition is termed pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections. Pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections typically occurs after an infection, most commonly caused by GAS. The underlying mechanism involves brain inflammation resulting from the immune system mistakenly attacking healthy brain cells, leading to autoimmune processes that disrupt central nervous system function. Affected children often exhibit abrupt and dramatic behavioral changes, including obsessive–compulsive disorder, anxiety, and tics or other abnormal movements (Swedo et al., 2012). On the other hand, patients with TS have been found to exhibit increased inflammatory activity, with elevated serum levels of tumor necrosis factor-alpha (TNF-α) and interleukin-12 (IL-12) compared to control subjects (Leckman et al., 2005). In another study, Yeon et al. (2017) recruited 29 patients without comorbid obsessive–compulsive disorder, the majority of whom were not on medication. The levels of six pro-inflammatory cytokines in the blood were compared between patient and healthy control groups. Among the cytokines assessed, IL-12 p70 and TNF-α levels were significantly elevated in the patient group relative to healthy controls. Furthermore, patients with anti-streptolysin O titers below 200 or Yale Global Tic Severity Scale scores between 10 and 19 also exhibited higher levels of IL-12 p70 or TNF-α. These findings suggest that inflammatory pathways play a significant role in the pathogenesis of TD.
Aberrant histaminergic function has also been proposed as a contributing factor in TD. Ercan-Sencicek et al. (2010) performed a genome-wide analysis of DNA samples from a two-generation pedigree with a high prevalence of TD and identified a rare nonsense mutation (W317X) in exon 9 of the histidine decarboxylase gene (Hdc). This discovery provided the first evidence linking histamine to TD pathogenesis. Hdc knockout mice exhibit repetitive behaviors and neurochemical features resembling TD and have since been recognized as a potential model for studying the pathophysiology of the disorder. Baldan et al. (2014) reported reduced striatal histamine levels in Hdc knockout mice and found that amphetamine treatment exacerbated TD-like stereotyped behaviors in these mice. Moreover, intracranial infusion of histamine alleviated these behavioral symptoms, further supporting the involvement of histamine in TD pathology. Rapanelli et al. (2017) demonstrated that specific ablation or chemogenetic silencing of histaminergic neurons in the tuberomammillary nucleus (TMN) of the hypothalamus resulted in markedly increased grooming behavior, a form of repetitive behavioral pathology, as well as elevated neuronal activity markers in both the dorsal striatum and medial prefrontal cortex. Infusion of histamine directly into the striatum reversed these behavioral abnormalities, confirming that acute histamine deficiency contributes to the observed effects.
The present study aimed to investigate whether acute histamine deficiency leads to immune abnormalities and inflammatory responses, which may contribute to the development of TD. To this end, we intracerebroventricularly administered α-fluoromethylhistidine (α-FMH) to rats to induce acute histamine depletion in the brain. Based on this model, we establish a TD animal model using 3,3′-iminodipropionitrile (IDPN) to explore whether acute histamine deficiency is accompanied by neuroinflammatory changes during neurotransmitter dysregulation, thereby contributing to the development of TD through both mechanisms.
Materials and Methods
Animals
Sixty specific pathogen-free male Sprague–Dawley rats, aged 3–5 weeks, were used in this experiment. All animals were purchased from Fuzhou Shichen Biotechnology Co., Ltd. and housed in the Animal Experiment Center of the 900th Hospital. The animal room environment was maintained under controlled conditions: temperature at 25 ± 2°C, relative humidity at 60% ± 5%, and a 12-h light/dark cycle. The rats had ad libitum access to autoclaved food and water. All animal experiments were approved by the Experimental Animal Ethics Committee of 900th Hospital of PLA Joint Logistic Support Force (Approval No. 2023-47). All animal experiments were conducted in strict accordance with the ARRIVE guidelines.
Animal grouping
After 7 days of acclimatization, rats were randomly divided into four groups (n = 15 per group): the normal control group (Control group), the TD model group (TD group), the TD model group receiving intracerebroventricular (ICV) injection of normal saline (TD-NaCl group), and the TD model group receiving ICV injection of α-FMH (Santa Cruz Biotechnology, Dallas, USA) (TD-α-FMH group).
Lateral ventricle cannulation and drug administration
Rats were anesthetized with isoflurane (Hengfengqiang, Jiangsu, China) administered via a continuous inhalation anesthesia machine and then secured in a stereotaxic apparatus (Ruiwode Life Technology, Shenzhen, China). The surgical site was shaved and disinfected to expose the skull. With bregma and lambda aligned on the same horizontal plane, a burr hole was drilled at the following coordinates relative to bregma, according to the Paxinos and Watson rat brain atlas: 1.0 mm posterior and 1.5 mm lateral to the right. A guide cannula was then implanted into the right lateral ventricle, at a depth of 4.0 mm from the skull surface. Following surgery, rats were housed individually for a 7-day recovery period. On postoperative days 8, 11, and 14, the TD-α-FMH group received an ICV infusion of 5 μg α-FMH solution at a rate of 1 μL/min using a micro-injection pump. The dose of 5 μg α-FMH per injection was chosen based on previous studies demonstrating that this dose achieved effective central histamine depletion in rats without nonspecific toxicity (Benetti et al., 2015; Rani et al., 2021). This three-dose regimen ensured sustained histamine depletion throughout the experimental period. The TD-NaCl group received an equivalent volume of sterile 0.9% NaCl into the lateral ventricle at the same time points.
TD model
TD was induced in rats according to the method described by Diamond (Diamond et al., 1982). Starting on day 9 post-cannulation, rats in the TD, TD-NaCl, and TD-α-FMH groups received daily intraperitoneal (i.p.) injection of IDPN (Sigma-Aldrich, Darmstadt, Germany) at a dose of 300 mg/kg for 7 consecutive days. The normal control group received daily i.p. injections of 0.9% NaCl (1 mL/kg) for the same duration. On day 8 post IDPN injection, stereotypic and locomotor behaviors were assessed and scored by two investigators blinded to the group assignments, using established rating scales for tic-like behaviors. Rats with scores ≥2 for both stereotypic and locomotor behaviors were used for experiments (Al Kadasah et al., 2009; Diamond et al., 1982). All rats in the TD, TD-NaCl, and TD-α-FMH groups achieved behavioral scores ≥2 for both stereotyped and motor behaviors and were included in the study. The exact timing of all procedures was shown in Figure 1.
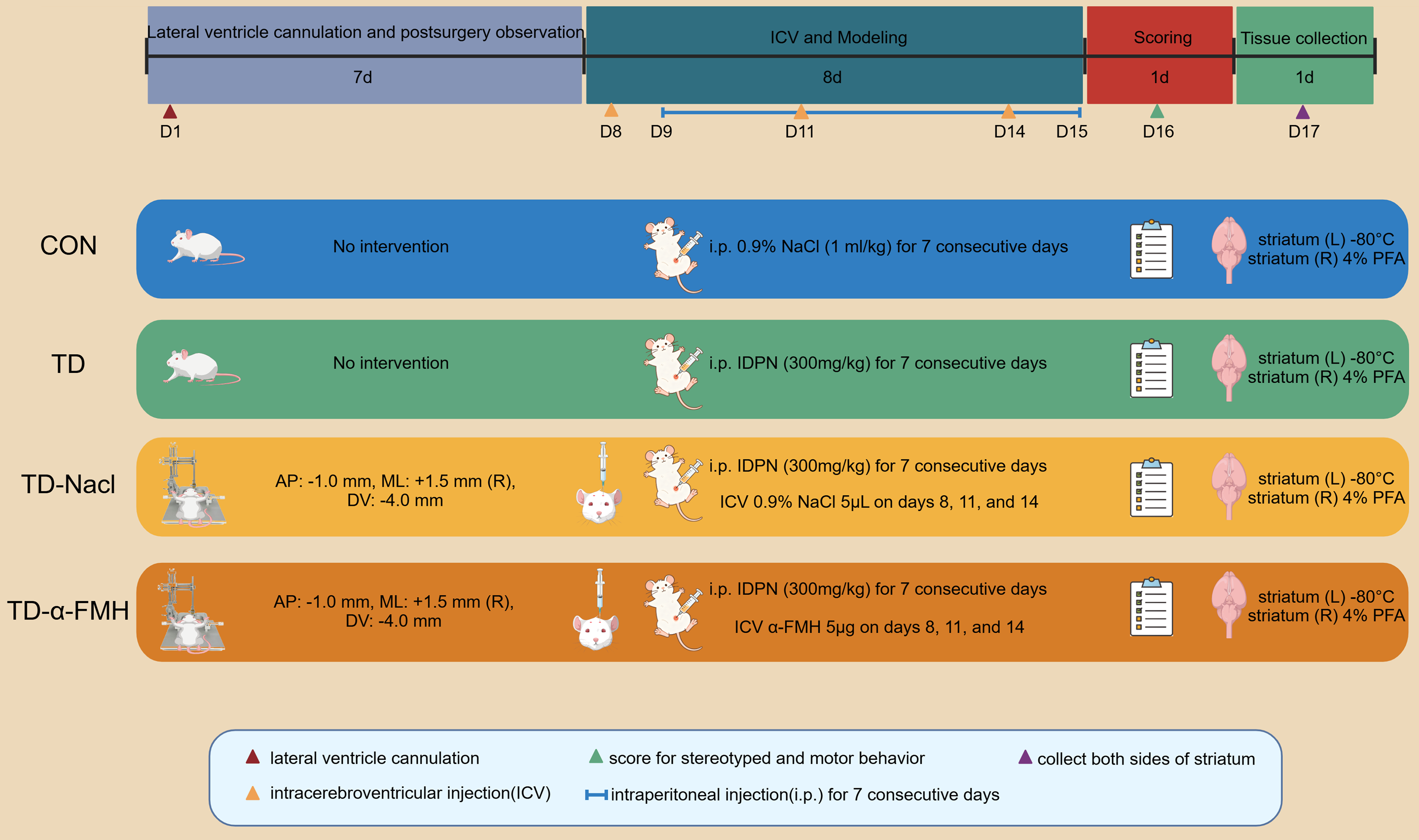
Experimental procedure. AP, anterior-posterior; Con, normal control group; DV, dorsal-ventral; ICV, intracerebroventricular; i.p., intraperitoneal; L, left; ML, medial-lateral; R, right; TD, TD model group; TD-NaCl, TD model group receiving ICV of normal saline; TD-α-FMH, TD model group receiving ICV of α-FMH.
Brain tissue collection and preparation
After behavioral assessment, rats were anesthetized and transcardially perfused with normal saline. The whole brain was removed. The left striatum was dissected, immediately frozen in liquid nitrogen, and stored at −80°C for subsequent specific enzyme-linked immunosorbent assay (ELISA) and liquid chromatography–tandem mass spectrometry (LC–MS/MS) analyses. The right striatum was dissected and fixed in 4% paraformaldehyde, followed by routine paraffin embedding and sectioning for immunohistochemical staining. The right striata from five randomly selected rats in each group were used for immunohistochemistry. The left striata of six rats randomly selected from the TD-NaCl and TD-α-FMH groups were used for LC–MS/MS. The left striata of the remaining nine rats in the TD-NaCl and TD-α-FMH groups, and those of nine rats randomly selected from the Control group and TD groups were used for ELISA. Due to insufficient tissue mass, some samples did not meet the requirements for ELISA assay.
Immunohistochemical staining
For Iba-1 staining, five rats were randomly selected from each group. Paraffin-embedded tissue sections (4 μm thick) were deparaffinized in xylene and rehydrated through a graded ethanol series. Antigen retrieval was performed using EDTA buffer, followed by blocking of endogenous peroxidase activity. Sections were incubated with an anti-Iba-1 polyclonal antibody (Proteintech Group, Inc. Chicago, USA), followed by a reaction enhancer and a horseradish peroxidase-conjugated goat anti-rabbit IgG polymer. Immunoreactivity was visualized using 3,3′-diaminobenzidine, and sections were counterstained with hematoxylin. After dehydration through an ethanol series, sections were cleared and coverslipped with neutral balsam. Iba-1 expression in the striatum was observed and images were captured under a light microscope.
ELISA
Striatal tissue samples were homogenized and centrifuged. The supernatants were collected, and the levels of TNF-ɑ, IL-6, and dopamine receptor D2 (DRD2) were measured using ELISA kits (Meike Biotechnology, Yancheng, China) following the manufacturer’s instructions. Absorbance was measured using a SPECTROStar microplate reader (BMG Labtech, Ortenberg, Germany).
LC–MS/MS
LC–MS/MS analysis of the six striatal tissue samples from the TD-NaCl and TD-α-FMH groups was performed by Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China).
Approximately 40 mg of each striatal tissue sample was mixed with 500 µL pre-cooled (−20°C) 70% methanol in a 2-mL centrifuge tube. The mixture was vortexed for 3 min and then centrifuged at 13,523 g for 10 min at 4°C. The supernatant was transferred to a new 1.5-mL centrifuge tube and incubated at −20°C for 30 min, which allows residual soluble proteins or lipids to precipitate. Following another centrifugation step under the same conditions, 200 µL of the supernatant was collected into a sample vial and stored at −20°C until analysis.
The sample extracts were analyzed using an ultra-performance LC–MS/MS system (UPLC, ExionLC AD; MS, QTRAP® 6500+). Chromatographic separation was carried out on a Waters ACQUITY UPLC HSS T3 C18 column (100 × 2.1 mm i.d, 1.8 µm) maintained at 40°C. The mobile phase consisted of 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B), at a flow rate of 0.35 mL/min. The gradient elution program was as follows: 0–8 min, 5–95% B; 8–9.5 min, 95% B; 9.6–12 min, 5% B. The injection volume was 2 µL.
Mass spectrometric detection was performed using an electrospray ionization source operated in both positive and negative ion modes. The ion source temperature was 550°C. The ion spray voltages were set at 5500 V for positive mode and −4500 V for negative mode. The curtain gas pressure was 35 psi. Data acquisition was performed in multiple reaction monitoring mode. The ion pairs (precursor ion → product ion) for each target analyte were optimized, along with their respective declustering potentials and collision energies.
All neurotransmitter concentrations were normalized to the weight of the striatal tissue sample. The lower limits of quantification (LLOQ) for each analyte were as follows: dopamine (LLOQ = 5 ng/mL), serotonin (LLOQ = 5 ng/mL), glutamate (LLOQ = 5 ng/mL), and acetylcholine (LLOQ = 10 ng/mL).
Statistical analysis
Statistical analyses were performed using SPSS software (version 26.0) and GraphPad Prism (version 8.0). Measurement data are presented as mean ± standard deviation (x¯ ± s) for normally distributed variables, or as median with interquartile range [M (P25-P75)] for non-normally distributed variables, or as median with interquartile range [M (P25-P75)] for non-normally distributed variables. Comparisons between two groups were conducted using the independent samples t-test for data meeting normality and homogeneity of variance assumptions; otherwise, the Mann–Whitney U test was used. Comparisons among multiple groups were performed using one-way analysis of variance (ANOVA), followed by the least significant difference (LSD) t-test for post hoc pairwise comparisons. A p value < 0.05 was considered statistically significant.
Results
Behavioral assessment
Rats in the TD, TD-NaCl, and TD-α-FMH groups all exhibited stereotyped and motor behavior scores ≥2, indicating successful establishment of the TD model. One-way ANOVA revealed significant differences among the four groups in both behavioral scores (stereotyped behavior, p < 0.001; motor behavior, p < 0.001). Further pairwise comparisons showed that rats in the TD-α-FMH group had significantly higher scores for both stereotyped and motor behaviors compared to those in the TD and TD-NaCl groups (p < 0.05). No significant difference was observed between the TD and TD-NaCl groups (p > 0.05) (Fig. 2).
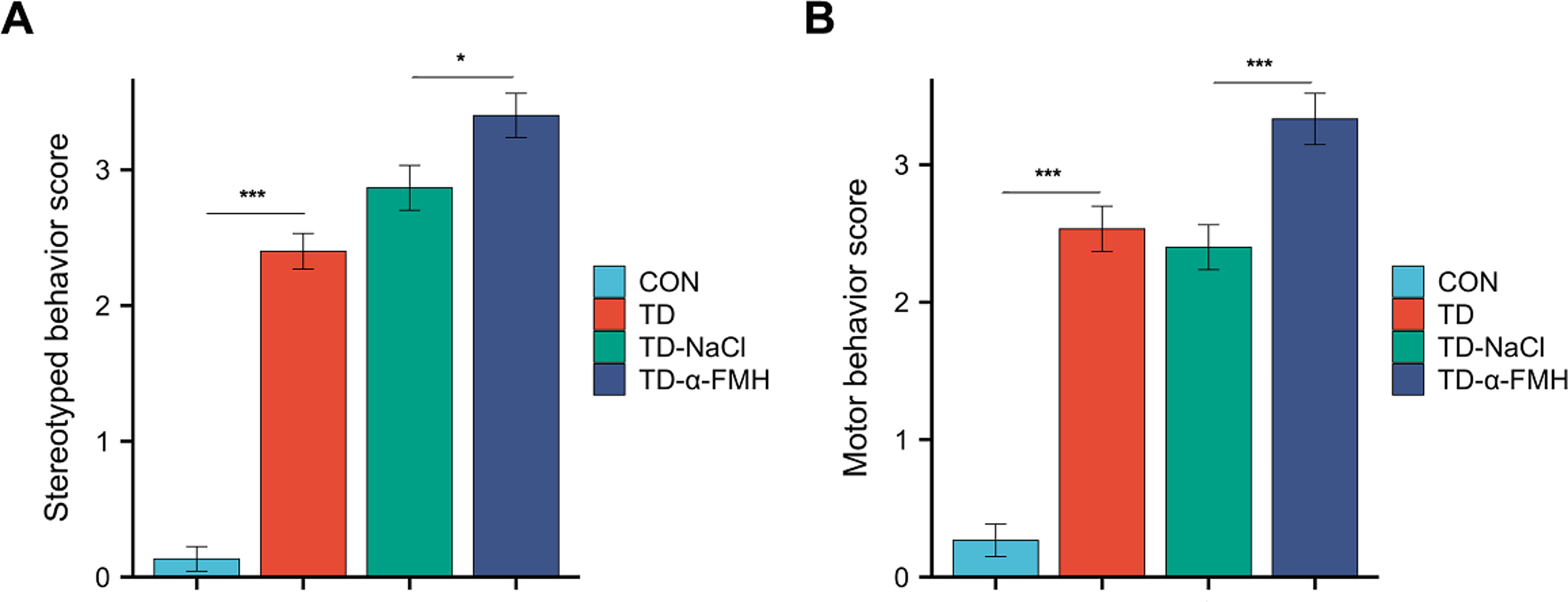
Comparison of stereotyped
Microglial morphology in the striatum
Immunohistochemical staining for Iba-1, a microglial marker, was performed to assess microglial activation in the striatum. In the Control group, Iba-1-positive cells displayed a resting, ramified morphology with small cell bodies and fine processes. In contrast, the TD and TD-NaCl groups exhibited a significant increase in the number of Iba-1-positive cells, which showed enlarged cell bodies and thickened, increased processes, displaying an amoeboid-like morphology and indicative of an activated state. The TD-α-FMH group also showed an increased number of Iba-1-positive cells compared to the Control group, comparable to the TD and TD-NaCl groups. However, the microglia in the TD-α-FMH group displayed a distinct morphology characterized by notably fewer processes and branches (Figs. 3 and 4).
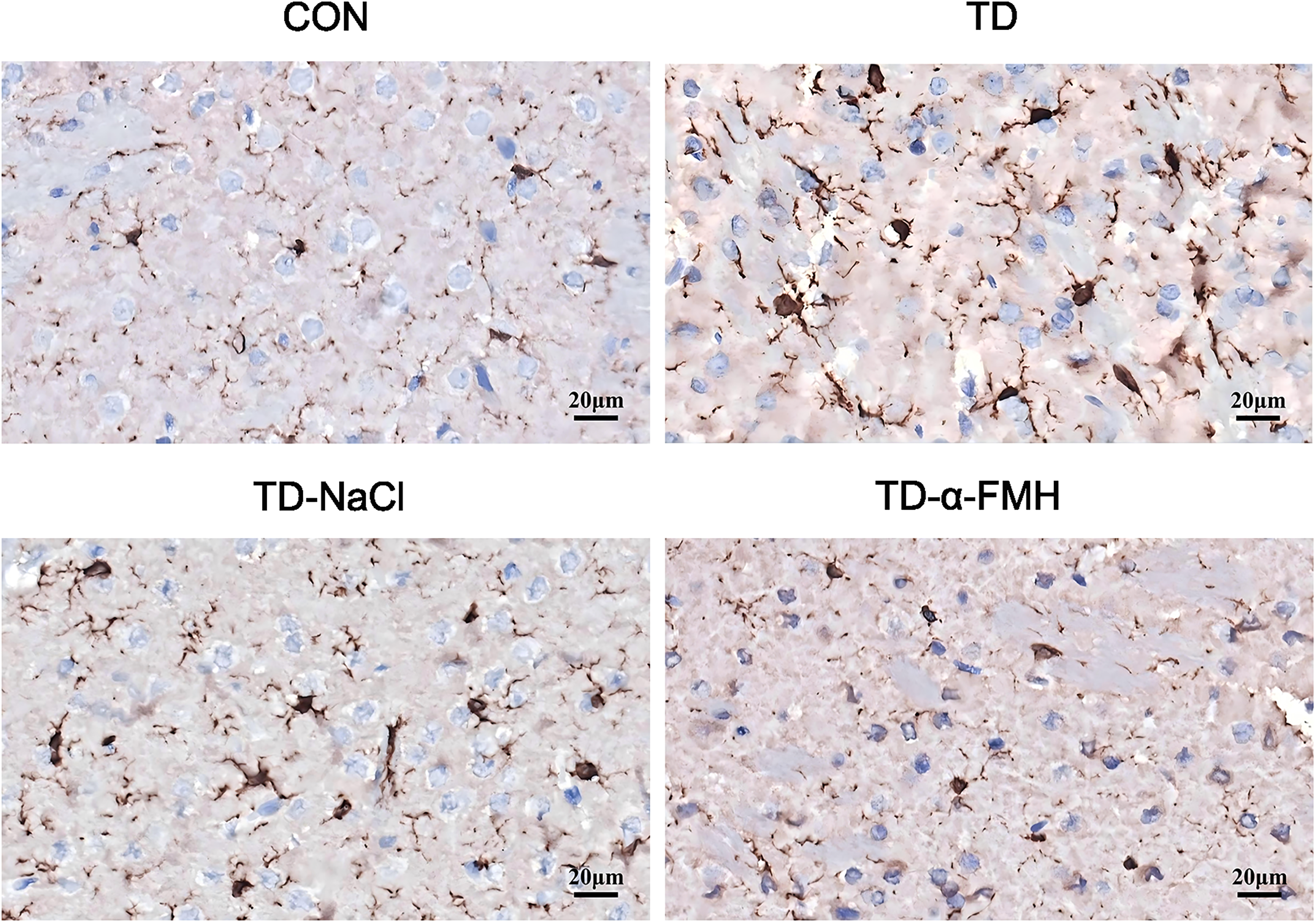
Representative images of Iba-1 immunohistochemical staining in the striatum (200 × magnification). Con, normal control group; TD, TD model group; TD-NaCl, TD model group receiving ICV of normal saline; TD-α-FMH, TD model group receiving ICV of α-FMH. Scale bar = 20 μm.
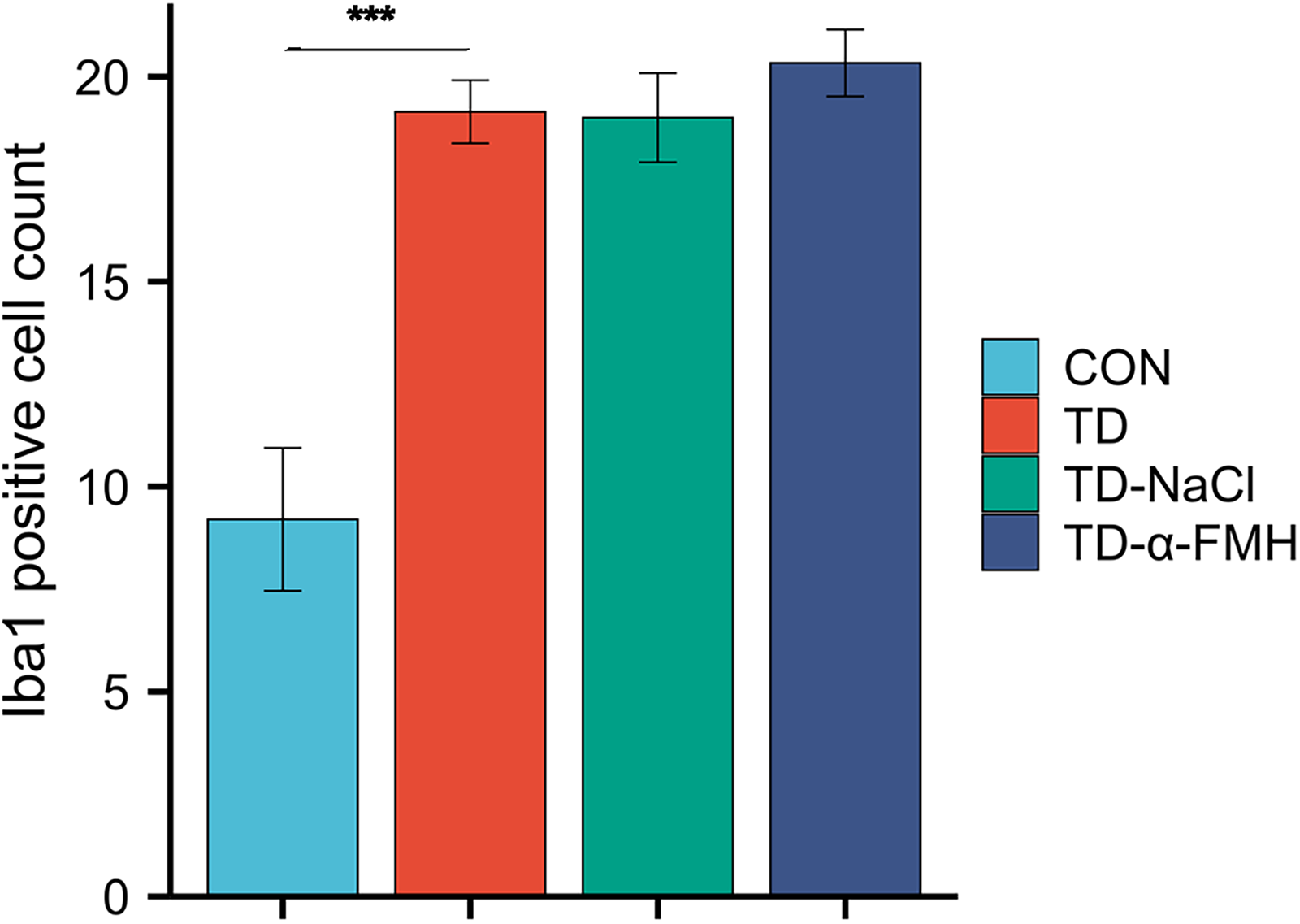
Quantitative analysis of Iba-1-positive cells in the striatum (200 × magnification). Con, normal control group; TD, TD model group; TD-NaCl, TD model group receiving ICV of normal saline; TD-α-FMH, TD model group receiving ICV of α-FMH. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Proinflammatory cytokine levels in the striatum
Levels of IL-6 and TNF-α in the striatum were significantly elevated in the TD, TD-NaCl, and TD-α-FMH groups compared to the control group in striatal tissues (p < 0.05 for all). Critically, the TD-α-FMH group exhibited a significantly greater increase in IL-6 levels compared to both the TD and TD-NaCl groups (p < 0.05). The TNF-α level in the TD-α-FMH group was also elevated relative to the TD and TD-NaCl groups, but this difference did not reach statistical significance (p > 0.05) (Fig. 5).
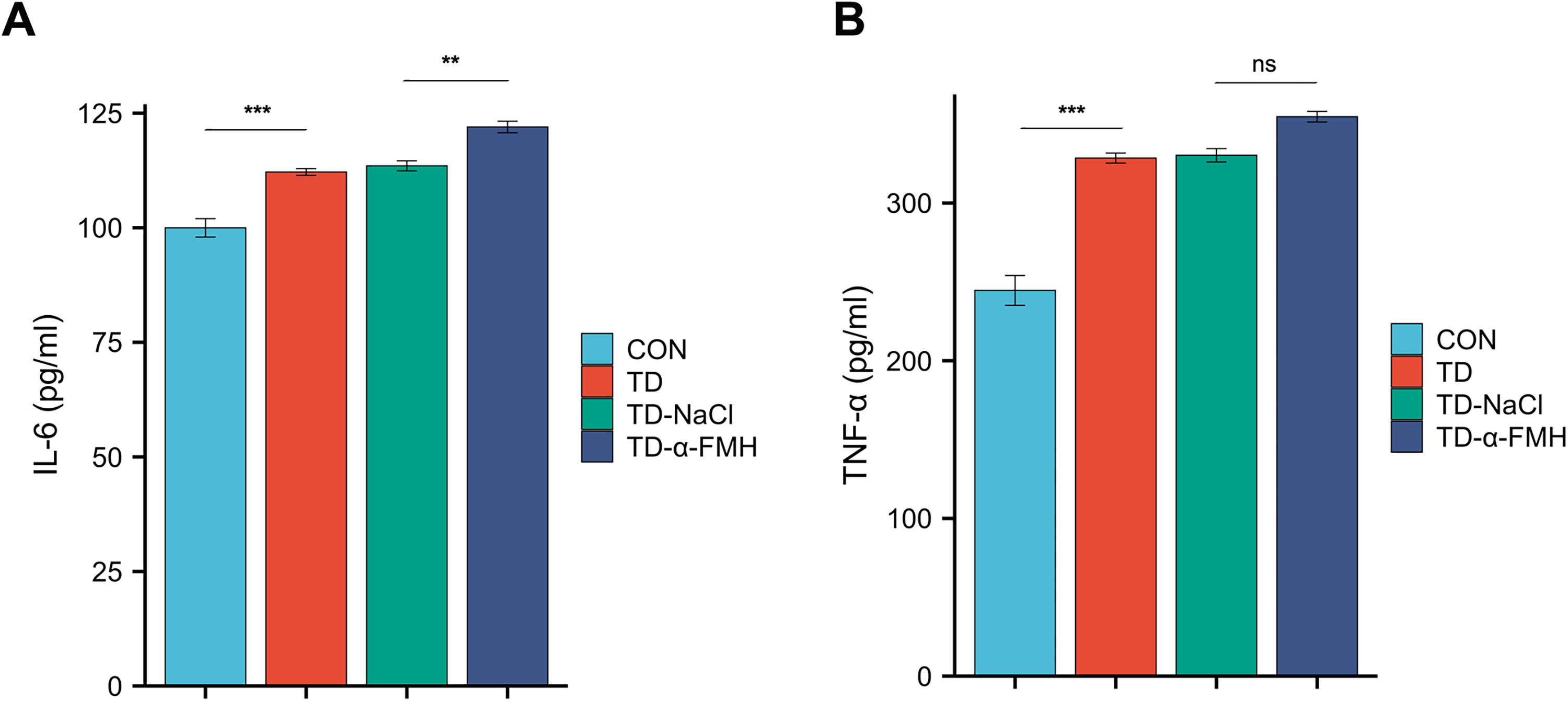
Comparison of IL-6
Dopamine D2 receptor (DRD2) expression in the striatum
Compared to the control group, DRD2 expression in the striatum were significantly increased in the TD, TD-NaCl, and TD-α-FMH groups (p < 0.05 for all). Notably, DRD2 levels in the TD-α-FMH group were significantly higher than those in the TD and TD-NaCl groups (p < 0.05), indicating that acute histamine deficiency further upregulated DRD2 expression in the TD model (Fig. 6).
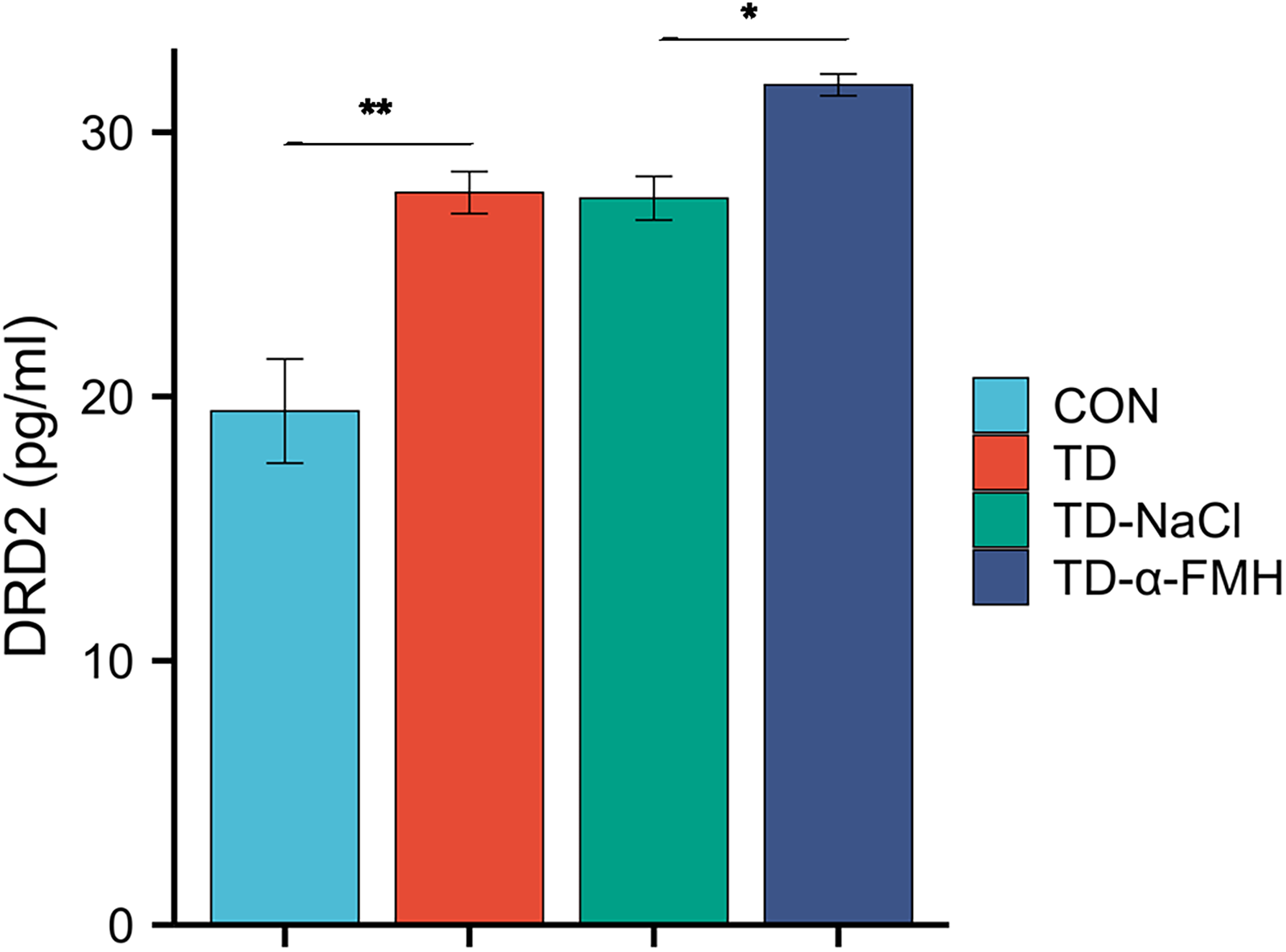
Comparison of striatal DRD2 levels. Con, normal control group; TD, TD model group; TD-NaCl, TD model group receiving ICV of normal saline; TD-α-FMH, TD model group receiving ICV of α-FMH n = 8 for each group. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Neurotransmitter levels in the striatum
Neurotransmitter levels in the striatum of the TD-NaCl and TD-α-FMH groups were measured using LC-MS/MS. The levels of key neurotransmitters in the striatum were compared between the TD-NaCl and TD-α-FMH groups using a Mann–Whitney U test. Among the four neurotransmitters quantified, glutamate levels were significantly lower in the TD-α-FMH group compared to the TD-NaCl group (p < 0.05). In contrast, no significant differences were detected between the two groups in the levels of dopamine, serotonin, or acetylcholine (p > 0.05 for all) (Fig. 7).
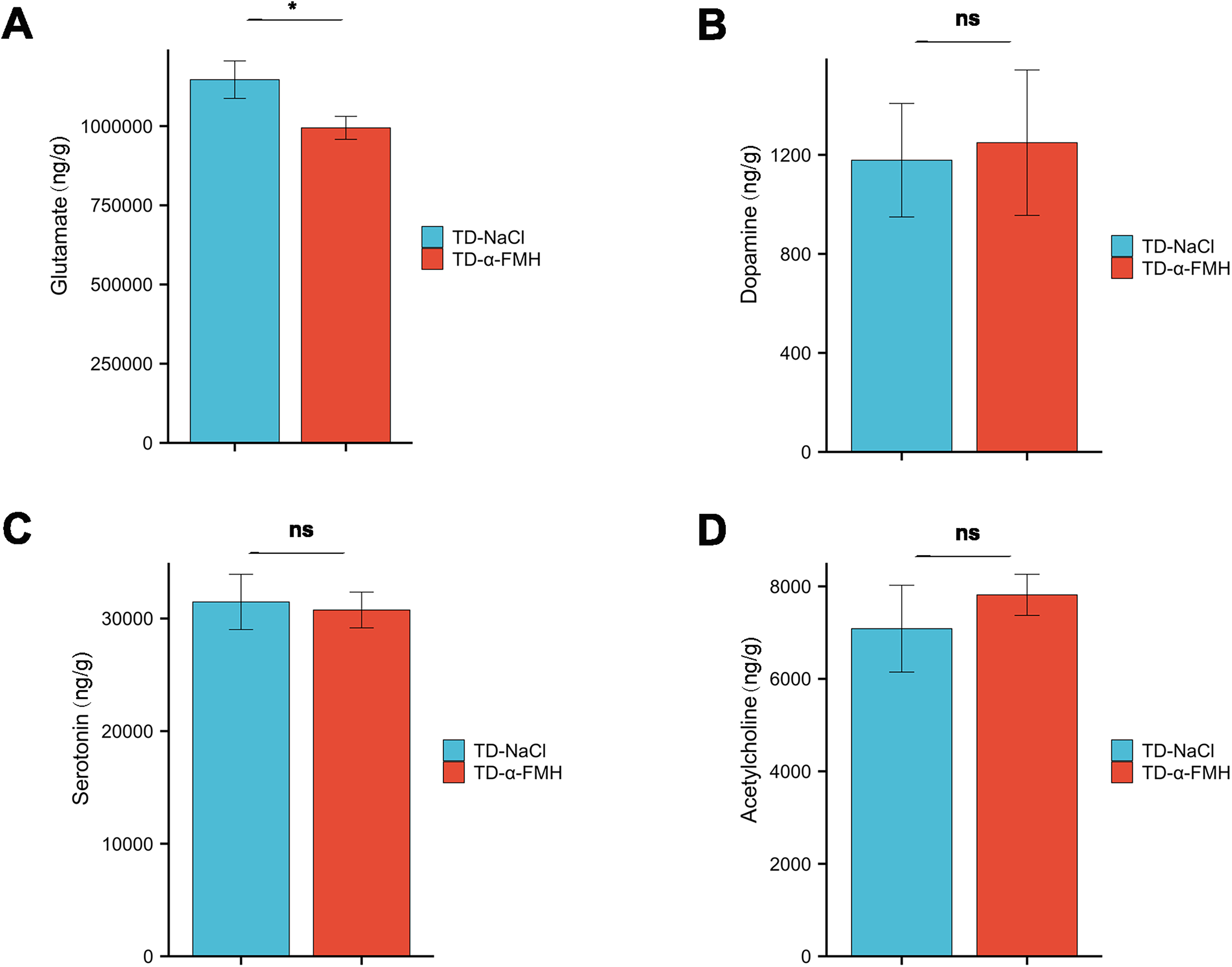
Striatal glutamate
Discussion
This study investigated the effects of acute histamine deficiency on neuroinflammation and neurotransmitter systems in the IDPN-induced rat model of TD. Our findings provide evidence that intracerebroventricular administration of α-FMH, an irreversible inhibitor of HDC, was associated with exacerbated tic-like stereotyped and motor behaviors in IDPN-treated rats. This behavioral worsening correlated with distinct morphological changes in striatal microglia, a significant elevation in the proinflammatory cytokine IL-6 and DRD2, and a selective reduction in striatal glutamate levels. These results offer novel insights into the complex pathophysiology of TD by linking histaminergic dysfunction to both neuroinflammatory and neurotransmitter abnormalities, supporting the histaminergic system as a potential therapeutic target for this disorder.
Our findings aligned with previous studies using genetic models of histamine deficiency. Baldan et al. (2014) demonstrated that Hdc knockout mice exhibit spontaneous repetitive behaviors and neurochemical alterations resembling TD, which were exacerbated by amphetamine treatment and alleviated by intracranial histamine infusion. Similarly, Rapanelli et al. (2017) showed that selective ablation or chemogenetic silencing of histaminergic neurons in the TMN increased pathological grooming behavior in rodents, a phenotype reversed by direct striatal histamine infusion. By using a pharmacological approach to acutely deplete histamine in a well-validated TD model, our study extended these findings by demonstrating that reduced central histamine levels, even in the absence of developmental genetic modifications, were sufficient to worsen TD-like symptoms. This suggests that histaminergic tone may act as a critical modulator of symptom severity, and that fluctuations in histamine signaling may contribute to the characteristic waxing and waning course of TD patients.
A key observation of our study was the association between acute histamine deficiency and amplified neuroinflammatory responses in the striatum. While IDPN administration alone induced significant microglial activation and elevated levels of IL-6 and TNF-α compared to controls, histamine depletion in the TD-α-FMH group resulted in a more pronounced inflammatory profile. Notably, microglia in the TD-α-FMH group displayed a distinct activated morphology characterized by enlarged cell bodies and reduced branching, a phenotype typically associated with a pro-inflammatory state, suggesting that histamine deficiency pushes microglia toward a more pronounced dysregulated phenotype (Ajoolabady et al., 2025; Mirarchi et al., 2024; Smith et al., 2012). Furthermore, IL-6 levels were significantly higher in the TD-α-FMH group compared to both the TD and TD-NaCl groups, while TNF-α showed a similar trend that did not reach statistical significance, possibly due to the late time point of measurement (day 8 post-IDPN), as TNF-α was often an early responder in inflammatory cascades (Riazi et al., 2008; Kuno et al., 2005).
Histamine may exert a tonic regulatory effect on microglial activity under physiological conditions. In its absence, the inflammatory response to a trigger like IDPN becomes dysregulated, leading to excessive release of pro-inflammatory cytokines such as IL-6. It was important to note, however, that our study did not establish a causal relationship between microglial activation and behavioral exacerbation. The observed IL-6 elevation might originate from other cell types, including astrocytes or neurons, and the morphological changes in microglia may represent an epiphenomenon rather than a causal factor. Future studies using pharmacological inhibitors of microglial activation (e.g., minocycline, PLX3397) or cell-specific genetic ablation will be necessary to definitively determine the role of microglia in histamine deficiency-induced TD exacerbation.
The significant upregulation of striatal DRD2 in all TD model groups is consistent with the well-established dopamine hypothesis of TD, which posits that hyperactive dopaminergic transmission, often mediated by supersensitive or overabundant D2 receptors, is a core pathophysiological mechanism (Gilbert et al., 2006; Yoon et al., 2007). The finding of our study is that acute histamine deficiency further increased DRD2 levels beyond those observed in the TD model alone, suggesting a regulatory interaction between the histaminergic and dopaminergic systems in the striatum.
Two potential mechanisms may underlie this observation. First, histaminergic neurons from the TMN project widely to the striatum, where they form synapses on medium spiny neurons (MSN) expressing DRD2 (Zhuang et al., 2018). Histamine can modulate MSN excitability via H1 and H2 receptors (Haas et al., 2008), while H3 receptors function as inhibitory heteroreceptors on dopaminergic terminals (Schlicker and Kathmann, 2017). Activation of striatal H3 receptors has been shown to increase dopamine release (Alfaro-Rodriguez et al., 2013). Therefore, histamine deficiency would remove this tonic inhibitory control, potentially leading to disinhibited dopaminergic transmission. The observed upregulation of DRD2 could represent a compensatory postsynaptic response to increased synaptic dopamine levels, although our LC–MS/MS analysis did not detect a significant difference in total striatal dopamine levels between the TD-NaCl and TD-α-FMH groups. This discrepancy may be explained by high inter-individual variability, rapid dopamine turnover, or the inability of total tissue measurements to distinguish between synaptic and intracellular dopamine pools.
Second, the heightened neuroinflammatory response in the TD-α-FMH group may itself drive DRD2 upregulation. Inflammatory cytokines, including IL-6, have been shown to alter the expression and function of dopamine receptors and modulate the sensitivity of the dopaminergic system (Zalcman et al., 1994). This interplay between inflammation and dopamine signaling represents a critical nexus in TD pathophysiology that warrants further investigation, as it may explain how immune triggers such as streptococcal infections contribute to tic onset or exacerbation in susceptible individuals.
Our neurotransmitter analysis revealed a selective reduction in striatal glutamate levels in the TD-α-FMH group compared to the TD-NaCl group, with no significant changes in dopamine, serotonin, or acetylcholine. This finding points to a specific disruption of glutamatergic signaling following histamine deficiency. Glutamate is the primary excitatory neurotransmitter in the brain, and corticostriatal glutamatergic projections play a central role in motor control and habit formation, processes that are dysfunctional in TD (Albin and Mink, 2006). The observed decrease in glutamate levels may be a direct consequence of DRD2 upregulation. DRD2 receptors are not only expressed postsynaptically on striatopallidal MSN of the indirect pathway but also presynaptically on corticostriatal glutamatergic terminals, where they act as inhibitory autoreceptors to reduce glutamate release (Surmeier et al., 2009). The significant upregulation of DRD2 in the TD-α-FMH group could therefore lead to enhanced presynaptic inhibition of glutamate release, resulting in the lower total tissue levels we detected. This provides a coherent mechanistic link between histamine deficiency, DRD2 upregulation, and glutamatergic dysfunction in TD.
Several limitations of this study should be acknowledged. First, as noted above, we did not directly test the causal role of microglial activation in the observed behavioral exacerbation. While our results demonstrate an association between altered microglial morphology, elevated IL-6 levels, and worsened tic-like behaviors, further experiments are required to establish causality. Second, all experiments were conducted in male rats. This choice was made to minimize confounding effects of hormonal fluctuations associated with the female estrous cycle, which can modulate neuroinflammatory responses and neurotransmitter signaling in initial mechanistic studies. However, this limits the generalizability of our findings to female animals and human females. Given the well-documented male predominance of TD and evidence of sex differences in neuroinflammatory and dopaminergic systems, future studies should include both male and female rats to assess potential sex-specific effects of histamine deficiency on TD pathogenesis. Third, we only assessed behavioral and molecular changes at a single time point (day 8 post-IDPN), which corresponds to the peak of tic-like behaviors in this model. This provides only a snapshot of the acute effects of histamine deficiency and does not allow us to determine whether the observed exacerbation of TD symptoms is transient or persists over time. Longitudinal studies assessing changes at multiple time points of the disorder will be necessary to characterize the temporal evolution of these effects. Four, intracerebroventricular administration of α-FMH depletes histamine throughout the entire brain, preventing us from identifying the specific brain regions or neural circuits where histamine loss exerts its most critical effects. The histaminergic system projects from the TMN to multiple brain regions involved in TD pathophysiology, including the striatum, cortex, and thalamus (Panula and Nuutinen, 2013). Future studies using region-specific knockdown of Hdc or chemogenetic manipulation of histaminergic projections will be required to dissect the circuit-level mechanisms underlying our findings.
Five, the LC–MS/MS analysis measured total tissue neurotransmitter levels, which do not distinguish between synaptic, vesicular, and metabolic pools. Techniques such as in vivo microdialysis will be necessary to measure real-time changes in neurotransmitter release and reuptake dynamics in response to histamine depletion. Finally, we only assessed a limited panel of inflammatory mediators (IL-6, TNF-α) and dopamine-related proteins (DRD2). A more comprehensive profiling of cytokines, chemokines, and other neurotransmitter receptors would provide a more complete picture of the molecular changes induced by histamine deficiency.
Conclusions
In conclusion, this study demonstrates that acute histamine deficiency is associated with exacerbated tic-like behaviors in the IDPN-induced rat model of TD. This effect correlates with a distinct pro-inflammatory microglial phenotype, significant upregulation of striatal DRD2, and reduced glutamate levels. These findings suggest that histamine deficiency may contribute to TD pathogenesis by promoting heightened neuroinflammation and disrupting the delicate balance between dopamine and glutamate signaling in the striatum. By elucidating the role of histamine as a key integrator of neuroinflammatory and neurotransmitter systems, our study opens new avenues for understanding the pathophysiology of TD and developing novel therapeutic strategies targeting the histaminergic system.
Data Sharing Statement
All data generated or analyzed during this study are included in this published article. All supporting data are available from the corresponding author on reasonable request.
Authors’ Contributions
J.Z. and Q.L.: Data collection, article writing, statistical analysis and organization; S.C. and R.S.: Data analysis and article revision; G.X.: Conceptualization, article revision, finalization, and funding acquisition.
Footnotes
Acknowledgments
Special thanks to all participants and researchers for their strong support of this study.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by Natural Science Foundation Project of Fujian Province (