
Abstract
Objectives
Ferroptosis is a distinct form of programmed cell death characterized by iron (Fe)-dependent oxidative stress and the accumulation of intracellular reactive oxygen species. Cisplatin (CPT), a chemotherapeutic agent, can induce ferroptosis in glioma cells and act synergistically with erastin (EST). The total extract of the fungus Sanghuangporus vaninii (TSV) has anticancer and antioxidant activities. This investigation aimed to assess the properties of TSV on CPT-induced inflammatory response and ferroptosis-related changes in glioma cells.
Methods
LNZ308 glioma cells were exposed to CPT and EST, followed by treatment with different TSV concentrations. Cell viability was assessed. Intracellular reactive oxygen species (ROS), glutathione peroxidase 4 (GPX4), glutathione (GSH), labile iron pool (LIP), LPO, interleukin-1 beta (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α) were examined, and cell migration was assessed using a scratch assay.
Results
TSV (25 µg/mL) exhibited limited toxicity to LNZ308 cells but failed to reverse the CPT-induced loss of cell viability. TSV markedly reduced ROS levels, LIP, and LPO, while restoring GSH and GPX4 levels. Moreover, it diminished IL-1β, TNF-α, and IL-6 in CPT-treated LNZ308 cells in a dose-responsive manner. L-ascorbic acid diminished ROS levels during oxidative stress. TSV also promoted scratch closure in both untreated and CPT-treated LNZ308 cells.
Conclusion
The results indicate that TSV modulates CPT-induced oxidative, inflammatory, and ferroptotic responses in LNZ308 glioma cells in vitro, supporting further investigation of its potential as an adjunct to chemotherapy.
Introduction
Glioblastoma (GBM) is the most common and aggressive primary malignant brain tumor in adults, with an annual incidence of approximately 3.5 cases per 100,000 individuals. 1 Despite therapeutic advances, the prognosis for patients with GBM has remained largely unchanged over the previous decade, with a median overall survival of 15–20 months and a 5-year survival rate of 5–7%. 2 The prevailing standard of care is maximal safe surgical resection followed by radiotherapy with concomitant and adjuvant temozolomide. 3 Novel therapeutic strategies, including autologous tumor- and dendritic cell-based vaccines, are currently under investigation; however, their efficacy remains limited, and survival benefit have not been established across these approaches. 4 The molecular processes underlying treatment resistance are diverse. The infiltrative characteristics of GBM render complete excision unfeasible. Hypoxic and perivascular niches support glioma stem cell maintenance and may contribute to recurrence and treatment resistance. 5 Furthermore, the blood–brain barrier and the brain–tumor barrier limit the delivery of antineoplastic agents into the brain. 6 ATP-binding cassette transporters facilitate the active efflux of therapeutic drugs from the brain parenchyma, leading to suboptimal treatment responses. 7 Additional therapeutic hurdles include intra- and inter-tumoral heterogeneity, dysregulated signaling pathways, and an immunosuppressive microenvironment. 7 Consequently, innovative, effective, and safe therapeutic alternatives are urgently required. 8
The concept of targeting GBM using phytocompounds has a long history. In this context, curcumin, isothiocyanates, resveratrol, and epigallocatechin-3-gallate (EGCG) have shown antitumor activity in preclinical GBM models.9-11 Recent pharmacological studies have demonstrated that Sanghuangporus spp. Exhibit anticancer, anti-inflammatory, antioxidative, antifungal, and immunomodulatory effects. Extracts from the medicinal fungus Phellinus rimosus exert cytotoxic and apoptotic effects on human HCT116 colon cancer cells. 12 The total ethanol extract of Phellinus igniarius induces apoptosis in SGC-7901 gastric cancer cells via a mitochondrial-dependent mechanism. S. vaninii extracts reduce the viability of B16-F10 murine melanoma cells at doses of 50–750 μg/mL. 13 Yu et al. found that polysaccharides derived from Phellinus gilvus can decrease the proliferation of murine melanoma cells and inhibit forestomach carcinogenesis. 13
Normal development, homeostasis, and the avoidance of hyperproliferative diseases such as cancer depend on cell death. 14 In addition to apoptosis, several forms of regulated cell death have been characterized, including necroptosis, pyroptosis, autophagy-dependent cell death, and ferroptosis. 15 Ferroptosis is a distinct form of programmed cell death characterized by iron-dependent oxidative stress and impaired cellular redox homeostasis. Ferroptosis’s potential as a cancer treatment is drawing increasing attention. 16 Ferroptosis suppresses tumor development, improves the efficacy of chemotherapy and radiation, and boosts antitumor immune responses, highlighting its potential in cancer therapy. 17 Recent studies suggest that ferroptosis is involved in GBM progression and treatment response. 18 CPT can induce ferroptotic cell death in several cancer cell types, and the ferroptosis inducer erastin can enhance CPT cytotoxicity by inhibiting system Xc− and depleting intracellular glutathione. 19 Accordingly, ferroptosis induction may enhance treatment efficacy and help overcome treatment resistance. 20
The total extract of Sanghuangporus vaninii (TSV) is a complex fungal preparation containing multiple bioactive compounds, including polysaccharides, polyphenols, flavonoids, and triterpenoids. 21 TSV may modulate signaling pathways implicated in oxidative stress, inflammation, and ferroptosis, including p53, nuclear factor erythroid 2-related factor 2, and the system Xc−/GSH/GPX4 axis.22-24 In addition, S. vaninii polysaccharides have shown anti-allergic effects in vivo. Nonetheless, the role of TSV in modulating CPT-induced ferroptosis-related changes in glioma remains unclear. This study investigated the effects of TSV on CPT- and EST-induced ferroptosis-associated oxidative stress and inflammatory responses in human LNZ308 glioma cells.
Materials and Methods
Preparation of TSV
The dried fruiting body of Sanghuangporus vaninii was obtained from Chun’an Qiandao Lake Sangdu Edible Mushroom Professional Cooperative (Hangzhou, China). The mature fruiting bodies were sliced and dried at 50 °C for 48 h in an oven. Water-soluble components were extracted with boiling water (1:10 w/v) for 2 h, then filtered. The filtered extracts were concentrated by rotary evaporation and lyophilized to obtain powders. The powders were stored at −20 °C until use.
For the cellular experiments, TSV powder (100 mg) was dissolved in 1 mL of ultrapure water, vortexed thoroughly, and centrifuged at 10,000 rpm for 10 min. The solution was sterilized by filtration through a 0.22 μm membrane and aliquoted for storage. The resulting 100 mg/mL stock solution was subsequently diluted to the desired concentration in the culture medium.
Cell Culture
In vitro experiments were performed using human glioma LNZ308 cells. LNZ308 cells were acquired from the cell bank of the Typical Culture and Preservation Centre of the Chinese Academy of Sciences, Shanghai, China, in accordance with ethical review criteria. LNZ308 cells were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum and 100 U/mL penicillin/streptomycin. All cells were maintained at 37 °C in a humidified atmosphere containing 5% CO2. The culture medium was changed regularly, and cells were passaged according to their growth conditions.
Cell Viability Assays
Cell viability was determined using the MTT assay, which measures the reduction of MTT, a yellow tetrazolium salt, into purple formazan crystals by NAD(P)H-dependent oxidoreductase enzymes.
25
LNZ308 cells (1.0 × 104 cells/well) were cultured in 96-well plates for 24 h. Fresh medium containing PBS vehicle or TSV (6.25–100 µg/mL) presence or absence of CPT (5 µg/mL) was added, and the cells were incubated at 37 °C for 24 h. MTT (5 mg/mL in PBS, 20 µL per well) was added to each well and incubated for 4 h. Dimethyl sulfoxide (150 µL) was added to each well, and the mixture was gently shaken for 20 min. Absorbance was measured at 570 nm using a microplate reader (Bio-Rad PR 4100, USA). Viability was determined using the formula:
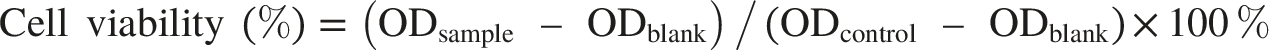
Measurement of Reactive Oxygen Species (ROS)
Intracellular ROS concentrations were assessed using an oxidation-sensitive H2DCFDA fluorescent probe. 25 LNZ308 cells (1.0 ×103 cells/well) were planted in a 96-well plate and treated with LAA (50 µg/mL) or TSV (3.13–18 µg/mL) with CPT (5 μg/mL) at 37 °C for 24 h. The cells were rinsed twice with PBS. Without treatment served as the control; the CPT-alone-treated group as the negative control; and the CPT and LAA-treated groups as the positive controls. Then, the cells were incubated with 20 μM H2DCFDA to detect intracellular ROS, and nuclei were counterstained with DAPI (2.5 µg/mL) for 30 min at 37 °C. The greenish fluorescent of oxidized dichlorofluorescein (excitation at 488 nm and emission at 530 nm) and the blue fluorescence of DAPI (excitation at 350 nm and emission at 465 nm) were imaged using a fluorescence microscope (Olympus, Tokyo, Japan). Fluorescence intensity (FI) was examined using a spectrofluorometer (SynergyH5, BioTek Instruments, Winooski, VT, USA) at 525 nm emission and 485 nm excitation.
Measurement of Membrane LPO
Membrane LPO was distinguished using a Liperfluo fluorescent probe. LNZ308 cells (1.0 ×105 cells/well) were treated with TOL (50 µg/mL) or TSV (3.13–18 µg/mL) with CPT (5 μg/mL) for 24 h. Without any treatment served as the control, the CPT alone-treated groups served as the negative control, and the CPT along with TOL-treated groups served as the positive control. The cells were stained with Liperfluo (1 μM) in DMSO at 37 °C for 20 min, then rinsed thrice with HBS. The cells were separate from the wells using EDTA/trypsin solution, transferred to a 1.5 mL microtube, and collected by centrifugation at 400× g for 10 min using an EBA 200 centrifuge with a fixed-angle rotor (Andreas Hettich GmbH & Co. KG, Tuttlingen, Germany). 26 The cells were analyzed by flow cytometry, and Liperfluor fluorescence was measured at 500–550 nm emission and 488 nm excitation.
Measurement of Mitochondrial LPO
Mitochondrial LPO was assessed using the MitoPeDPP fluorescent probe. LNZ308 cells (1.0 × 105 cells/well) were treated with TSV (3.13–18 µg/mL) or TOL (50 µg/mL) in the presence of CPT (5 μg/mL) and EST (50 µM) at 37 °C for 24 h. Without any treatment served as the control; the CPT alone-treated group served as the negative control; and the CPT along with TOL or EST-treated groups served as the positive control. The treated cells were stained with MitoPeDPP at 50 µM for 15 min at 37 °C in the dark, then washed with buffer. The cells were analyzed by flow cytometry, and MitoPeDPP fluorescence was measured at 452 nm excitation and 470 nm emission.
Assessment of Apoptosis
Apoptosis was assayed using the Annexin V–AbFluor/DAPI apoptosis detection kit, as described in a previous report. 27 LNZ308 cells (1.0 ×104 cells/well) were subjected to EST alone (10 µM) or CPT (5 µg/mL) with TSV (0-12.5 µg/mL) for 24 h at 37 °C and harvested by trypsinization. Without any treatment served as the control, the CPT alone-treated groups served as the negative control, and the CPT along with EST-treated groups served as the positive control. After washing thrice with PBS, the cells were suspended in Annexin V binding buffer solution and stained with Annexin V–AbFluor (5 µL) and DAPI (2.5 µL) for 20 min. The stained cells was investigated within 45 min using flow cytometric analysis at 617 nm for DAPI/517 nm for AbFluor™ 488 emission and 491 nm excitation wavelengths.
Analysis of Inflammatory Cytokines by ELISA
The 96-well plates were coated overnight at 4°C with capture monoclonal antibodies against TNF-α (2 µg/well) and IL–6 (2 µg/well) in PBS pH 7.4. The plates were rinsed thrice with Tween–20 (0.05%) containing PBS, and blocked with 2% (w/v) bovine serum albumin. LNZ308 cells (2.0 ×103 cells/well) were subjected to EST (10 µM) or CPT (5 µg/mL) with TSV (3.13–12.5 µg/mL) for 24 h. Then, the culture medium was gathered and diluted 1:20 with the dilution solution for the quantification of secreted TNF-α and IL-6. IL-6 and TNF-α standards (100 µL each) were added to the antibody-coated wells for 2 h at 30 °C. After washing with PBS, HRP-conjugated antibodies against TNF-α (0.5 µg/mL) and IL-6 (0.25 µg/mL) were added to each well plates. The plates were gently shaken and incubated for 1 h at 30 °C. After washing thrice with PBS, 200 µL of tetramethylbenzidine substrate containing 3% H2O2 was added to each well plates, and after 20 min, using 2 N H2SO4. Absorbance was determined at 450 nm using a microplate reader (PR4100, Bio-Rad, USA). 28 Results are presented as mean ± standard deviation from four independent experiments.
Scratch Assay
Different concentrations of TSV (3.13–25 µg/mL) were prepared in sterile culture medium. LNZ308 cells (3.0 × 102 cells/well) were incubated in a 6-well plate until 80% confluence. Straight scratches were made with a 1 mL pipette tip, and the cell layers were washed with PBS. The cells were exposed to CPT (5 μg/mL) and subjected to TSV (3.13–25 µg/mL) or LAA (50 µg/mL) for 24 h. Without treatment served as the control; the CPT-alone-treated group served as the negative control; and the CPT- or LAA-treated groups served as the positive controls. Cells were imaged using an inverted microscope (Olympus, Tokyo, Japan) at 0, 24, and 48 h after treatment. The scratch area was examined using ImageJ software version 2.16.0. 29
Statistical Analysis
All experimental data were subjected to statistical analysis utilizing GraphPad Prism 9.0 software (GraphPad Software, USA). Results are presented as the mean ± standard deviation (SD), with a sample size of n = 4. One-way and two-way analyses of variance (ANOVAs) were used for group comparisons, depending on the experimental design. Significance levels: *p < 0.05, **p < 0.01, ***p<0.001, ns, not significant. Error bars in graphs denote the standard deviations to illustrate variability within the data. This statistical method validates the dependability of the experimental results.
Results
Effects of TSV Against CPT-Induced Cytotoxicity
TSV at 50–100 µg/mL substantially reduced cell viability (Figure 1). The IC50 of TSV was 42.5 µg/mL, indicating moderate cytotoxicity. CPT at 10 µg/mL reduced viability to 75% and yielded an IC50 of 30.8 µg/mL. TSV did not reverse the CPT-induced reduction in cell viability under the tested conditions. Accordingly, TSV concentrations below 25 µg/mL were used in subsequent experiments because they produced limited cytotoxicity when administered alone. Cell viability of TSV. Dose response curve of TSV on LNZ308 viability with and without CPT treatment. Data were stated as means ± SD (n = 4)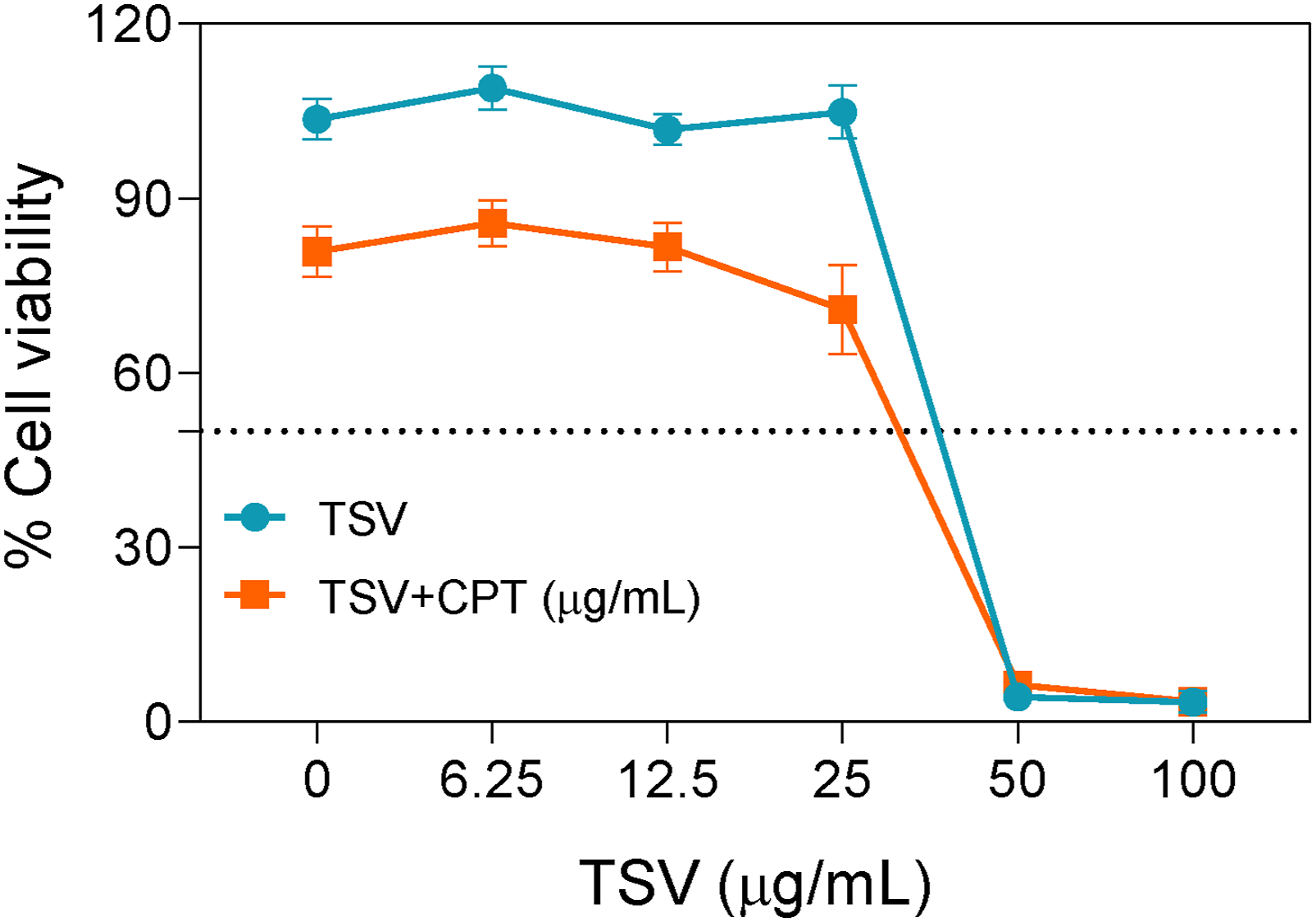
TSV Reduces CPT-Induced Oxidative Stress
Treatment of LNZ308 cells with CPT (5 µg/mL) markedly elevated ROS levels relative to control cells (Figure 2A), whereas TSV (3.13–12.5 µg/mL) reduced this effect. DAPI fluorescence revealed altered nuclear staining patterns in CPT-treated cells, with these changes less pronounced after TSV treatment. Quantitative FI analysis showed that TSV and LAA (50 µg/mL) significantly reduced ROS levels relative to the CPT-only group in a dose-responsive manner, with 12.5 µg/mL producing the greatest effect (Figure 2B and C). ROS levels in LNZ308 cells were induced by CPT and subjected to TSV or LAA. (A) Fluorescence imaging of ROS levels. (B) DCF intensity. (C) ROS production. Data were stated as means ± SD (n = 4). **P<0.01, ***P<0.001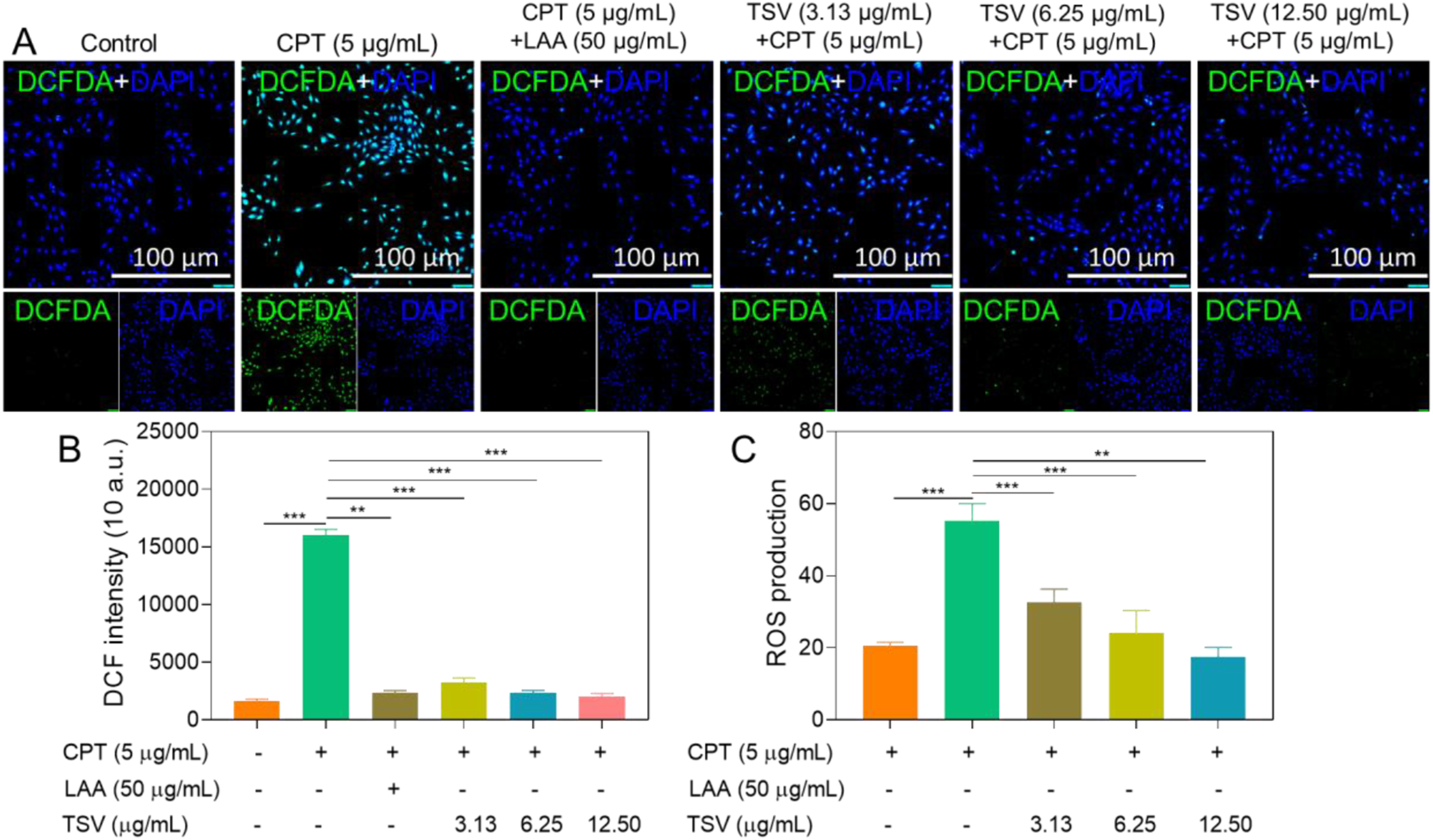
TSV Inhibits LPO
LPO levels are shown as representative FI peaks in Figure 3A and quantified as relative FI in Figure 3B. CPT (5 µg/mL) suggestively augmented LPO levels in LNZ308 cells compared to control cells. In contrast, TSV and TOL (100 µg/mL) reduced this effect in a dose-responsive manner, and the results indicate that TSV reduces CPT-induced lipid peroxidation in LNZ308 cells. LPO levels in LNZ308 cells were induced by CPT and subjected to TSV or TOL. (A) Flow cytometry image of LPO levels. (B) LPO ratio quantification. Data were stated as means ± SD (n = 4). **P<0.01, ***P<0.001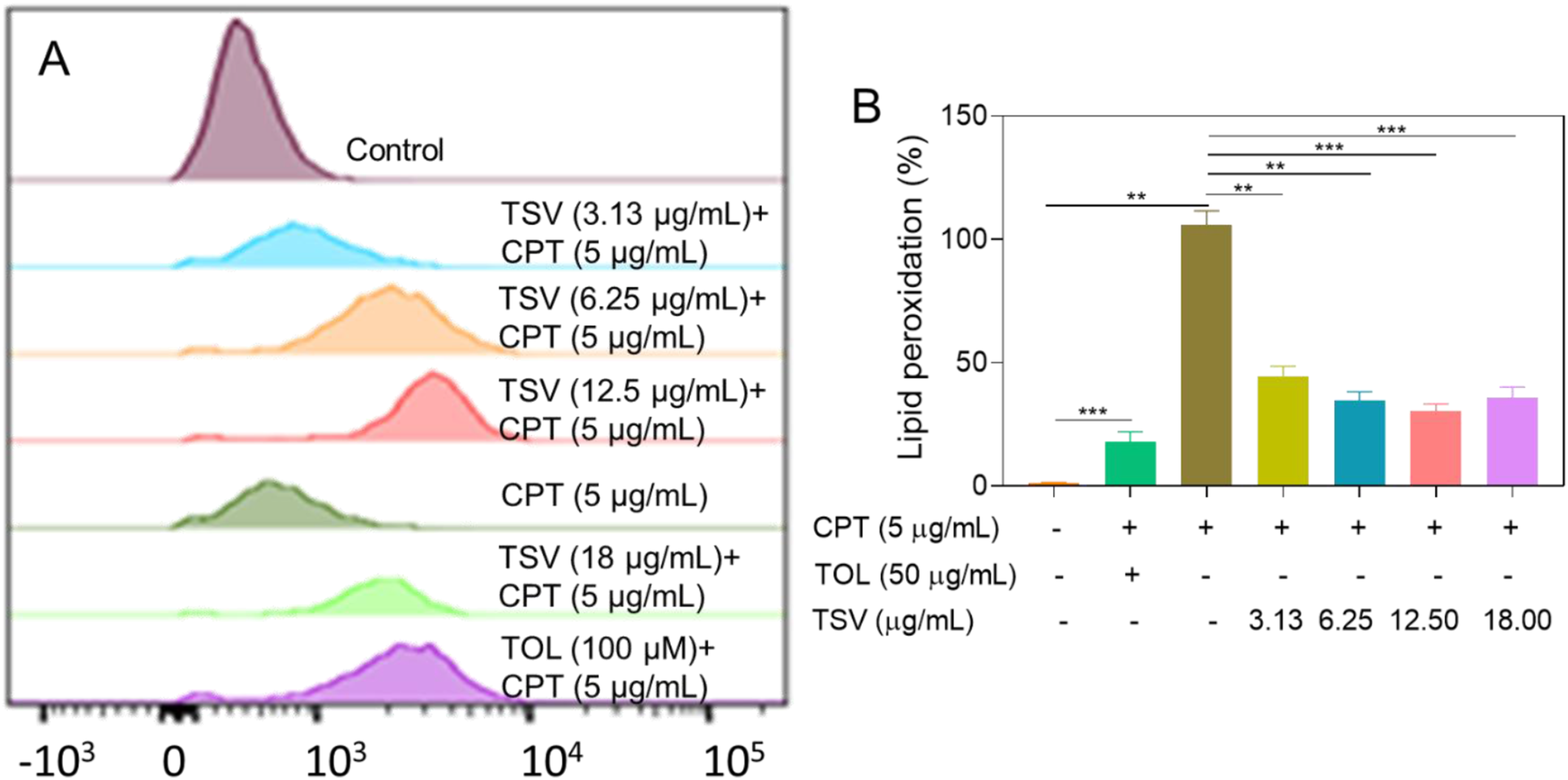
The analysis of MitoPeDPP fluorescence indicated increased mitochondrial LPO (Figure 4A). Green FI was highest in LNZ308 cells treated with CPT (5 µg/mL) and 50 µM EST compared to control cells (Figure 4B). TSV (3.13–12.5 µg/mL) and TOL (100 µM) reduced mitochondrial LPO. These findings indicate that TSV and TOL inhibited LPO induced by CPT and EST in LNZ308 cells. Mitochondrial redox-active iron (MitoPeDPP) levels in LNZ308 cells were induced by CPT and subjected to TSV, TOL, or EST. (A) Fluorescence imaging of MitoPeDPP. (B) MitoPeDPP intensity. Data were stated as means ± SD (n = 4). *P<0.05, **P<0.01, ***P<0.001, compared with control group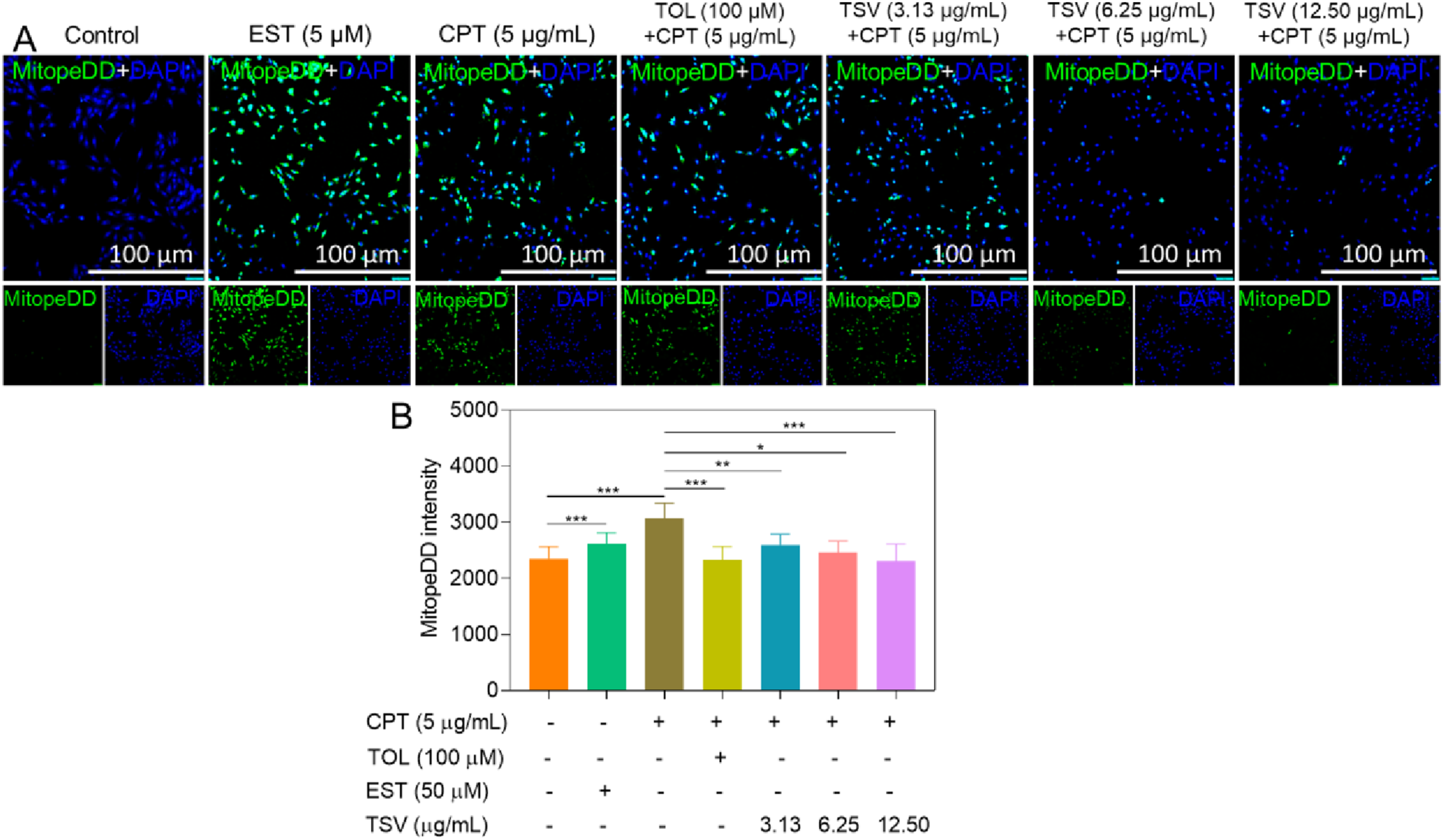
Effect of TSV on the LIP
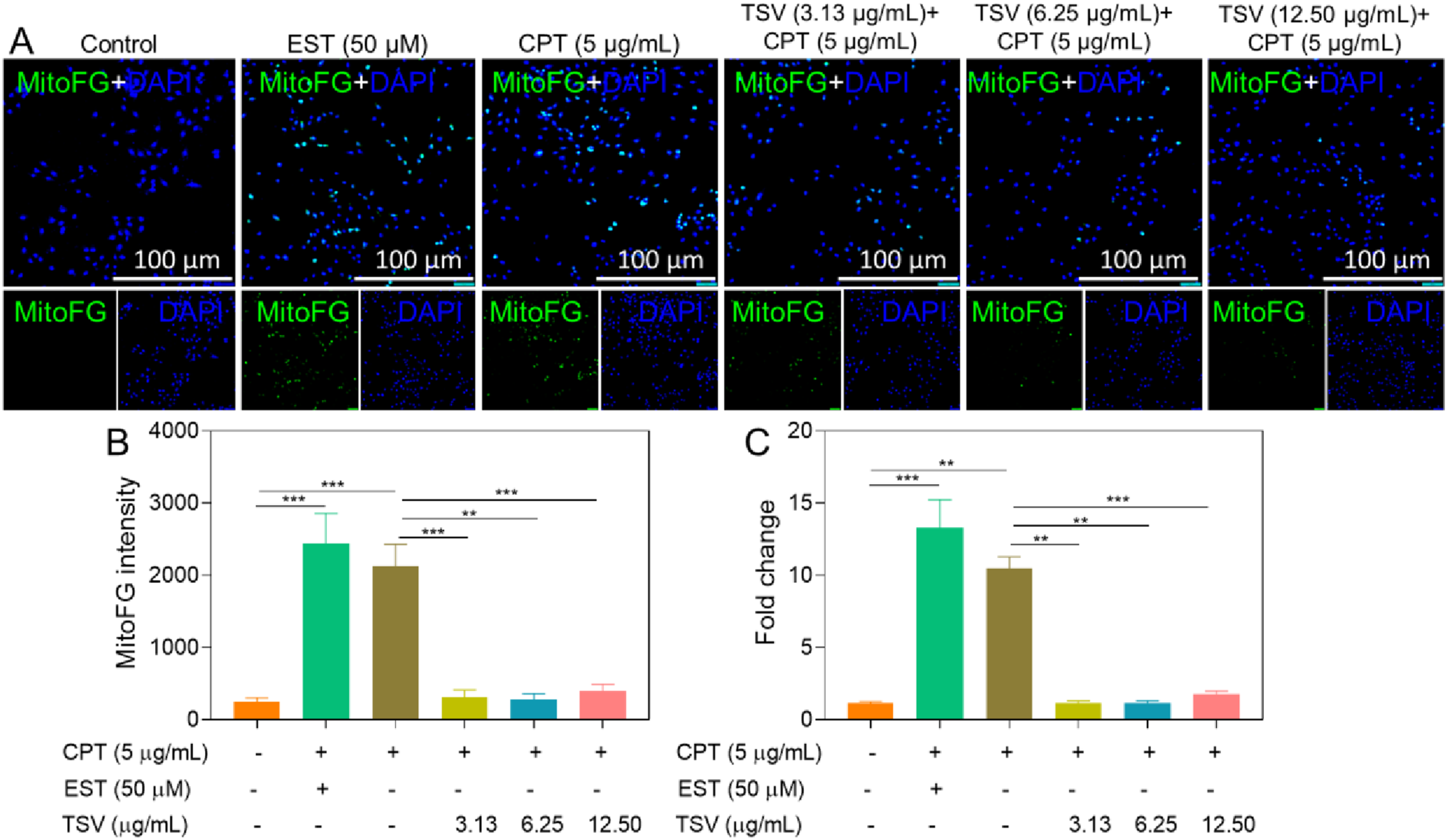
CPT (5 µg/mL) increased the intracellular LIP in LNZ308 cells relative to control cells; TSV (3.13 and 6.25 µg/mL) and 100 µM deferoxamine (DFO) attenuated this increase (Figure 5A and B). Furthermore, combined treatment with TSV and DFO did not reduce LIP further compared with either agent alone. These findings indicate that TSV and DFO attenuated the CPT-induced increase in LIP. CPT (5 µg/mL) and EST (50 µM) significantly reduced mitochondrial LIP, whereas TSV treatment attenuated this reduction (Figure 6A and C). Cytosolic LIP (cLIP) levels in LNZ308 cells were induced by CPT and subjected to TSV without/with DFO chelation. (A) Monotherapy of cLIP. (B) Combinational therapy of cLIP. Data were stated as means ± SD (n = 4). **P<0.01, ***P<0.001 Mitochondrial FerroGreen (MitoFG) levels in LNZ308 cells were induced by CPT and subjected to TSV. (A) Fluorescence imaging of MitoFG. (B) MitoFG intensity. (C) Fold change of MitoFG. Data were stated as means ± SD (n = 4). **P<0.01, ***P<0.001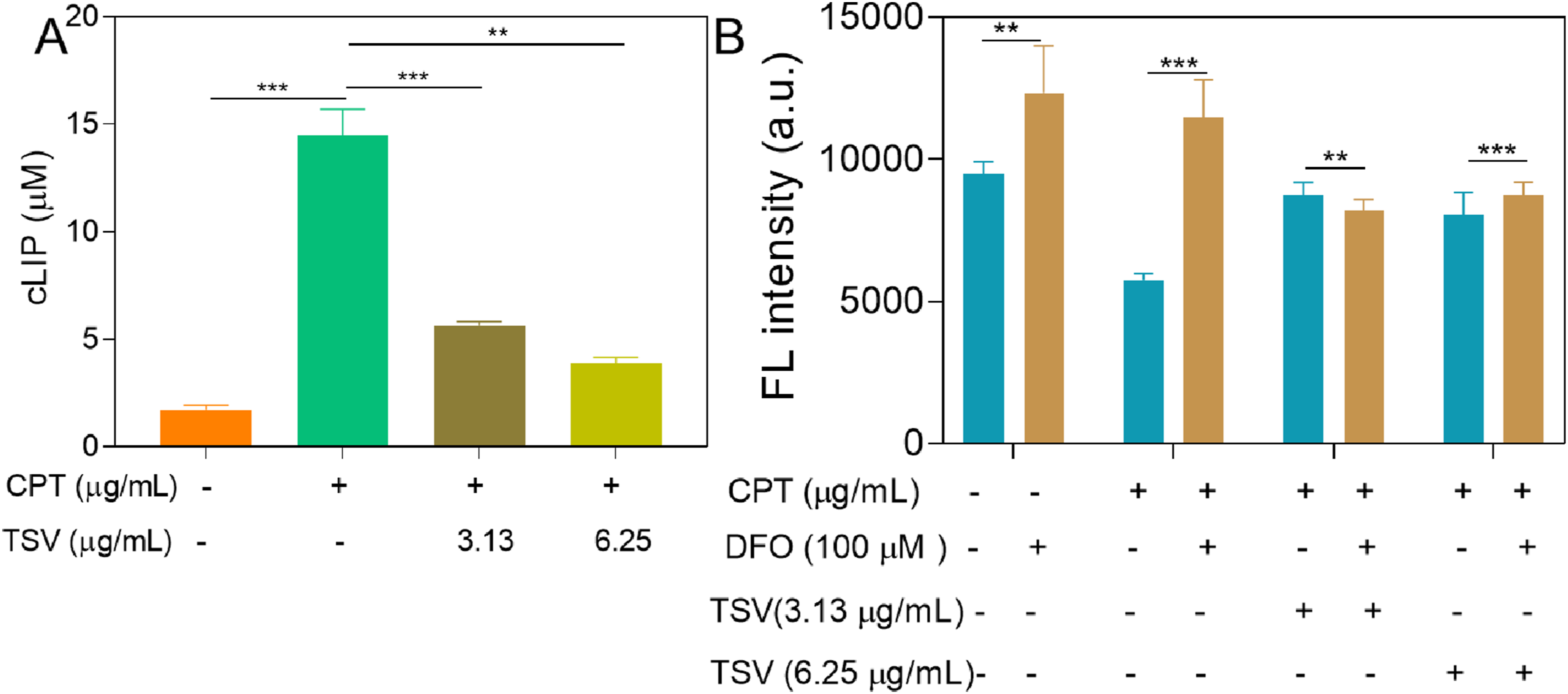
TSV Restores GSH and GPX-4 Levels
CPT (5 µg/mL) and EST (50 µM) significantly decreased GSH and GPX4 levels in LNZ308 cells, and TSV (3.13–12.5 µg/mL) attenuated these decreases (Figure 7A and B). These findings suggest that TSV partially restored the GSH–GPX4 antioxidant defense system under CPT- and EST-induced stress, consistent with the attenuation of ferroptotic stress. GSH and GPX-4-specific activity in LNZ308 cells was depressed by CPT or EST and subjected to TSV. (A) GSH activity. (B) GPX-4-specific activity. Data were stated as means ± SD (n = 4). *P<0.05, **P<0.01, ***P<0.001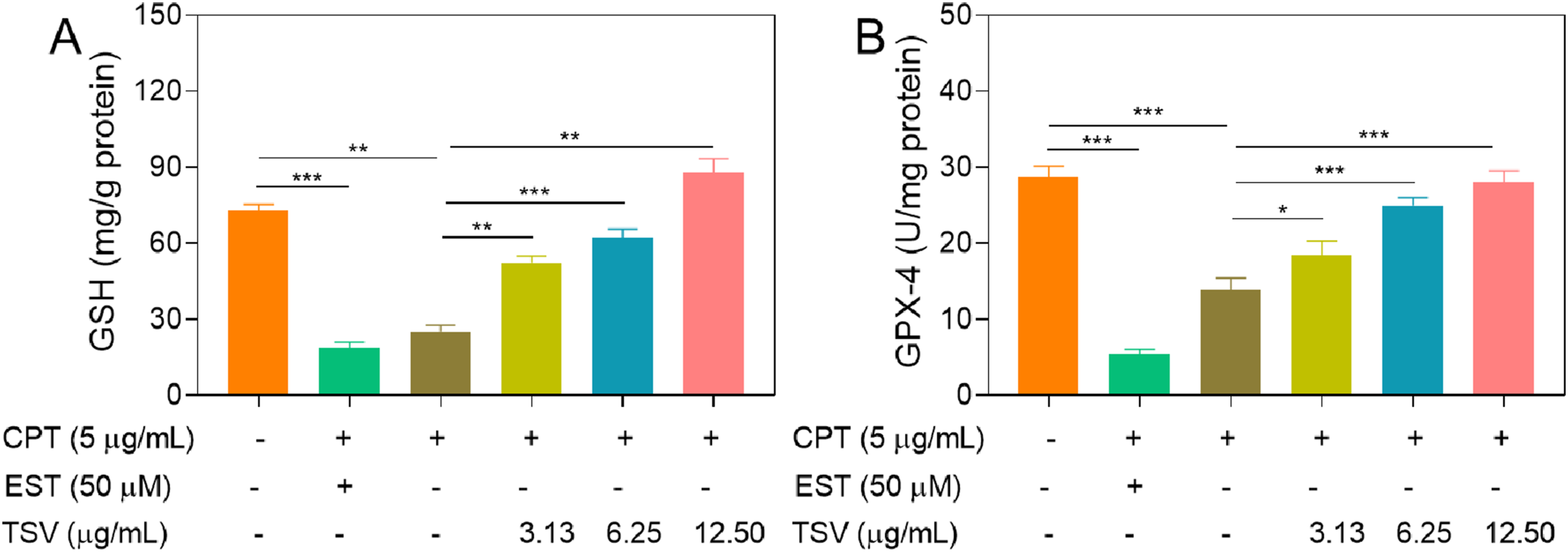
Effects of TSV on Ferroptotic Cell Death
CPT (5 µg/mL) induced ferroptosis-related changes, including increased cell death, ROS production, LPO, and redox-active Fe, along with decreased GSH and GPX4 levels. Flow cytometry analysis showed that CPT (5 µg/mL) and EST (10 µM) increased ferroptotic cell death, whereas TSV attenuated this effect (Figure 8A). Quantitative analysis showed that CPT and EST increased ferroptotic cell death, whereas TSV significantly reduced this effect (Figure 8B). LNZ308 cell viability was induced by EST or CPT and subjected to TSV. (A) The percentage of apoptosis ratio. (B) Cell death ratio. Data were stated as means ± SD (n = 4). ***P<0.001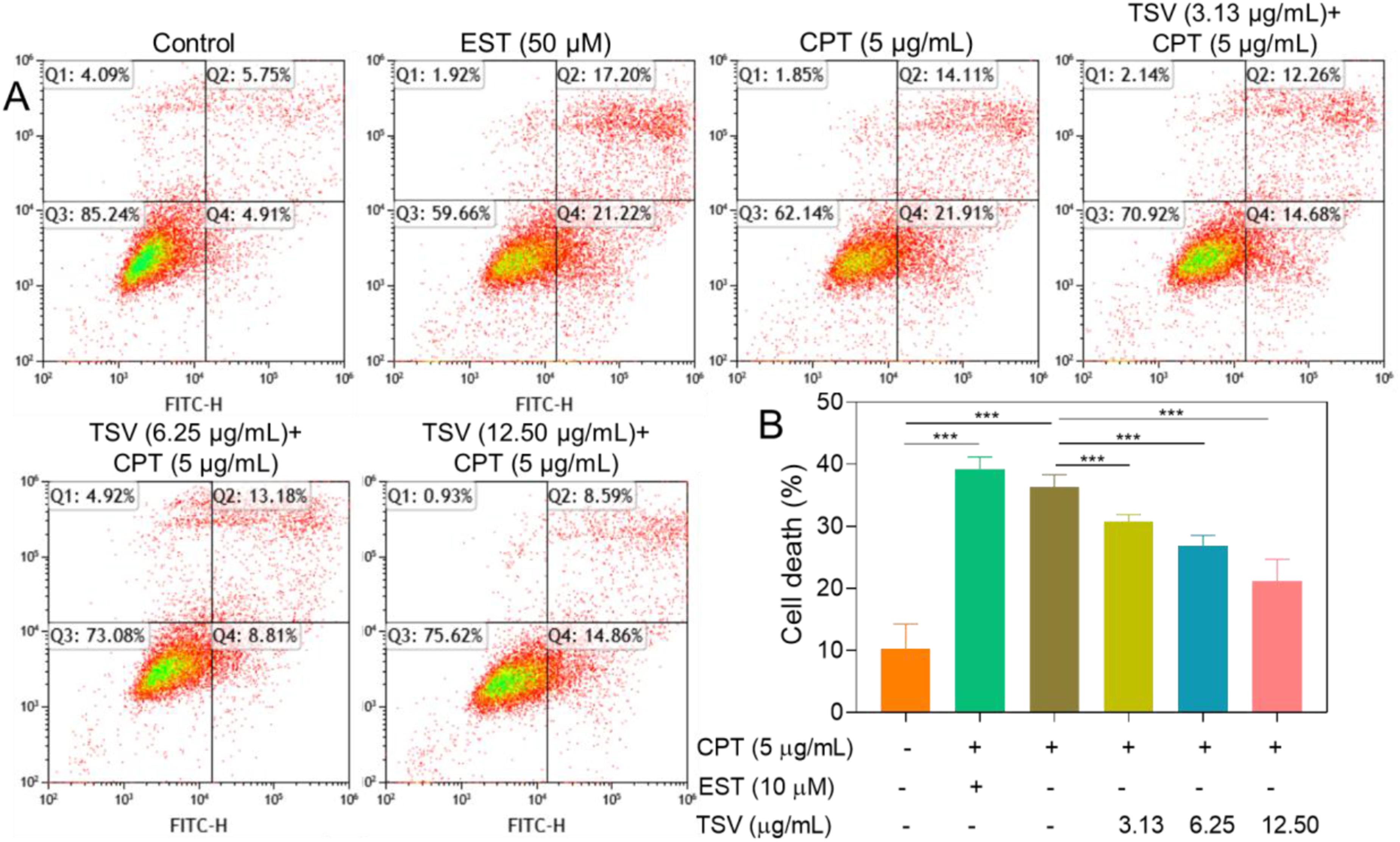
Effect of TSV on Cytokine Secretion
The anti-inflammatory effect of TSV was assessed by measuring the levels of the pro-inflammatory cytokines TNF-α and IL-6. CPT (5 µg/mL) markedly elevated TNF-α and IL-6 secretion, and TSV (3.13–18 µg/mL) and LAA (50 µg/mL) attenuated this increase in a dose-responsive manner (Figure 9A and B). The results suggest that TSV exerts anti-inflammatory effects in CPT-treated LNZ308 cells. TNF–α and IL–6 production from LNZ308 cells was induced by CPT and subjected to TSV. (A) TNF–α production levels. (B) IL–6 production levels. Data were stated as means ± SD (n = 4). *P<0.05, **P<0.01, ***P<0.001
Effects of TSV on Scratch Closure
The scratch assay results showed that CPT reduced the scratch closure of LNZ308 cells, while TSV mitigated this effect in a dose- and duration-responsive manner compared with CPT-treated cells (Figure 10). These findings suggest that TSV partially counteracted the CPT-associated impairment of scratch closure in LNZ308 cells. The migration ratio of LNZ308 cells was induced by CPT and subjected to TSV. (A) Microscopic image. (B) Wound closure ratios. Data were stated as means ± SD (n = 4). *P<0.05, **P<0.01, ***P<0.001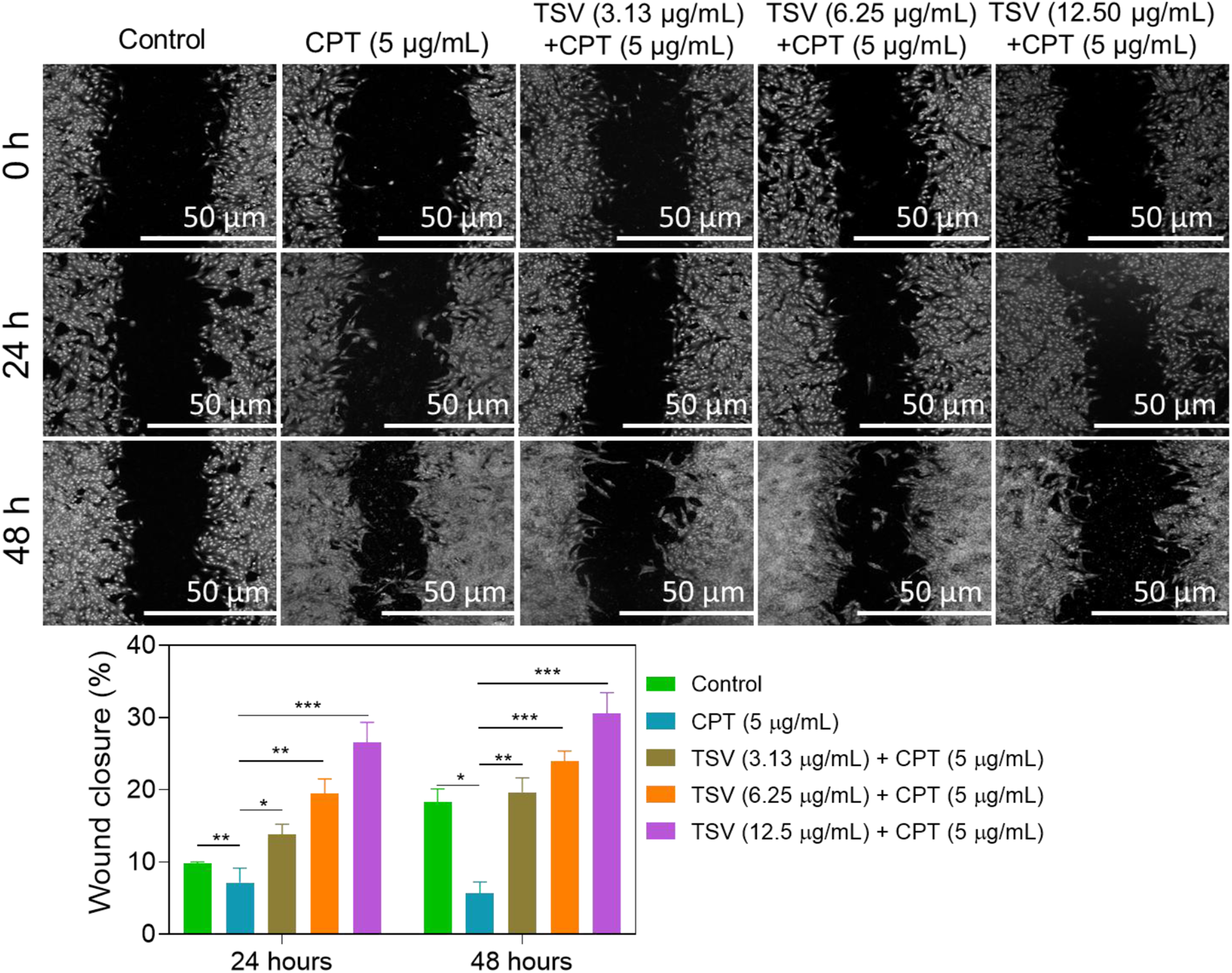
Discussion
Glioblastoma remains the most aggressive glioma in adults. The current standard of care is maximal safe resection followed by radiotherapy (typically delivered over approximately 6 weeks) with concurrent and adjuvant temozolomide. 30 Tumor-treating fields, when added to maintenance temozolomide, have improved survival in selected cases, and supportive care improves symptom control and patient quality of life. 31 Despite multimodal therapy, the prognosis remains poor, with a median overall survival of 12–15 months and a 5-year survival rate below 10%. Neurological and cognitive symptoms are common in glioma and are clinically relevant because they contribute to morbidity and may also correlate with outcomes. 32
S. vaninii is an edible and medicinal fungus whose fruiting bodies contain multiple bioactive compounds, including polysaccharides, polyphenols, flavonoids, and triterpenoids. The phenolic-rich extracts of S. vaninii have been reported to contain multiple phenolic compounds, including protocatechualdehyde, protocatechuic acid, caffeic acid, and (+)-taxifolin; these extracts exhibited antioxidant, anti-inflammatory, and antiproliferative activities. TSV attenuated ferroptosis-induced liver injury by reducing malondialdehyde and activating nuclear factor erythroid 2–related factor 2 (NRF2)–GPX4 signaling, whereas polysaccharide fractions decreased IL-1β, IL-6, and TNF-α levels in LPS-stimulated macrophage models. These extracts have also shown antiproliferative activity in several cancer cell models, including HepG2, A375, and SiHa cells, and a water-soluble polysaccharide fraction showed modest antiproliferative activity in MCF-7, HeLa, SHG-44, and SMMC-7721 cells.33-36
Our results revealed that TSV at the concentrations tested exhibited cell toxicity in LNZ308 cells; however, higher doses (50 and 100 µg/mL) decreased cell viability (IC50 = 42.5 µg/mL). Other studies have shown that S. vaninii has antiproliferative and pro-apoptotic activity in other tumor models; however, those studies used different extract types, compositions, and concentration ranges, limiting direct comparison with our cellular model.
In the present study, L-ascorbic acid reduced ROS levels, indicating that the assay could detect oxidative stress. Consistent with this finding, S. vaninii preparations have shown antioxidant activity, and total or polyphenol-rich S. vaninii extracts attenuated oxidative stress in non-glioma models, with involvement of the NRF2–GPX4 pathway.13,30-33
We found that TSV attenuated CPT- and EST-induced ferroptosis-related changes in LNZ308 cells by reducing cytosolic and mitochondrial ROS, LIP, and LPO products, while partially restoring cellular GSH and GPX4 levels.37-39 These biochemical changes are consistent with the attenuation of oxidative and ferroptotic stress. CPT can induce ferroptosis-related oxidative stress, and EST is a canonical system Xc− inhibitor that promotes glutathione depletion and LPO. This study demonstrated that CPT and EST increased Annexin V-positive cells, whereas TSV treatment mitigated this effect. Because Annexin V detects phosphatidylserine externalization and is not specific to apoptosis alone, the results suggest that TSV reduced the proportion of Annexin V-positive cells under oxidative stress rather than definitively establishing apoptosis inhibition.40-42
S. vaninii preparations have shown antiproliferative and pro-apoptotic activity in other tumor models. A phenolic-rich extract induced HepG2 cell apoptosis through a caspase-dependent mitochondrial pathway, an aqueous fruiting body extract induced cell cycle arrest and apoptosis in A375 melanoma cells, and a polysaccharide fraction promoted apoptosis and reduced migration or invasion in MCF-7 cells via p53-related signaling. 43
A proposed summary of the observed effects of TSV on CPT- and EST-associated oxidative stress, inflammatory signaling, and ferroptosis-related changes is shown in Figure 11. The results show that TSV promotes scratch closure in CPT-treated LNZ308 cells. TSV was observed to enhance EGFR and ERK activity, promote cell migration and proliferation in the LNZ308 cell line, and aid wound healing. Alongside flavonoids and phenolics, there are active constituents in the crude extract of TSV, which may exhibit noteworthy biological activity in CPT-challenged LNZ308 cells. Phenylboronic acid-Hyaluronic acid-TSV nanoparticles demonstrated promising anti-inflammatory, ROS-scavenging, and antioxidant characteristics in LNZ308 cells, along with tissue-remodeling and wound-healing capabilities in a mouse model. Furthermore, ECG reduced hydrogen peroxide generation and the cellular activation signals associated with cell death in ultraviolet-exposed LNZ308 cells in a concentration-responsive manner. Similarly, EGCG may decrease inflammation and tachyphylaxis in dexamethasone-treated LNZ308 cells by downregulating the expression of C-C motif ligands, TGF-β, IL-18, and IL-6.
44
Plant flavonoids, including quercetin, facilitate antioxidant wound healing and anti-inflammatory effects and influence glioma cancer migration in LNZ308 cells through the NF-κB and ERK1/2 MAPK pathways.
42
The TSV mechanism ameliorates CPT-triggered glioma tumor cell damage by regulating inflammation and ferroptosis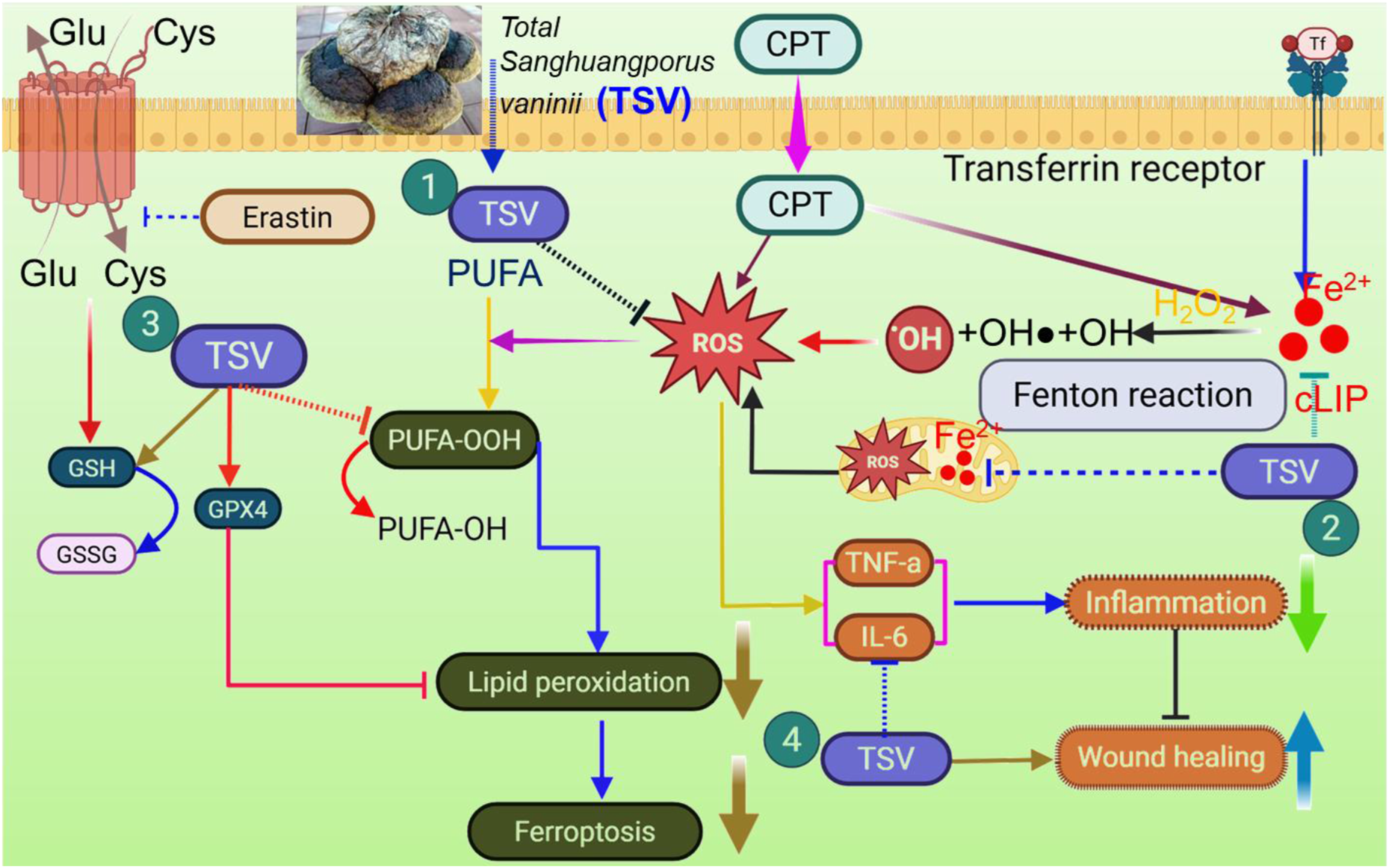
This study has limitations. First, the study used a single in vitro glioma model, limiting its generalizability. Second, the crude TSV preparation was not comprehensively characterized chemically; therefore, the active constituents responsible for the observed effects remain undefined. Third, the properties of TSV on IL-6, TNF-α, and other cytokines relevant to tumor biology were not examined due to cost constraints. Fourth, underlying signaling pathways and the effects of active components were not examined due to limited laboratory resources.
Conclusions
TSV modulated CPT-induced oxidative stress, inflammatory responses, and ferroptosis-related pathways in LNZ308 cells in vitro. Specifically, TSV reduced ROS, lipid peroxidation, and LIP, partially restored GSH and GPX4 levels, decreased IL-6 and TNF-α levels, and mitigated the CPT-induced reduction in scratch closure. However, TSV did not restore overall cell viability under CPT treatment. These findings demonstrate the biological activity of TSV in this in vitro glioma model. Future studies should identify the active constituents of TSV and the underlying pathways, evaluate additional glioma models, and determine whether these effects translate into therapeutically relevant antitumor activity.
Supplemental Material
Supplemental Material - Exploring sustainable tourism indicators (STIs) for the U.S. with secondary data
Supplemental Material for Total Sanghuangporus vaninii Extract Ameliorates Cisplatin-Induced Glioma Cell Death by Regulating Ferroptosis and Inflammation by Yankai Xu, Jiesi Zhou, Xiaobin Zhou, Yong Li, Senyuan Yang, Zebin Xue in Dose-Response
Footnotes
Author Contributions
All authors read and approved the final version of the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data supporting this study’s findings are available from the corresponding author upon reasonable request.
Supplemental Material
Supplemental material for this article is available online.
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.