
Abstract
Background and Purpose
Radiation-induced ovarian injury (RIOI) compromises the clinical utility of pelvic radiotherapy and reduces fertility in young female patients. Dimethyl sulfoxide (DMSO) exhibits antioxidant property. Hearin, we aimed to explore the protective effect and underlying mechanism of DMSO on RIOI.
Materials and Methods
C57BL/6J female mice were treated with DMSO prior to undergoing lower abdominal radiation. Intragenerational and transgenerational fertility were assessed by mating experiment. Ovaries were harvested to evaluate follicle count and morphology. Granulosa cells were analyzed by immunohistochemical staining of TUNEL and Ki67. An ovary explant organ culture system was established to evaluate the effect of DMSO. Fluorescent probes were employed to evaluate mitochondrial mass and function. DNA double-strand breaks were detected by immunoblotting of γ-H2AX.
Results
DMSO preserved the fertility of irradiated mice and even safeguarded the reproductive capacity of their unirradiated female offspring. Histological analyses revealed that DMSO preserves the ovarian follicle reserve, including both primordial and developing follicles. Additionally, DMSO demonstrated radioprotective effects in ex vivo ovarian tissue. Moreover, DMSO reduced apoptosis and enhanced proliferation in granulosa cells. Mechanistically, DMSO alleviated radiation-induced oxidative stress and preserved mitochondrial function, as evidenced by increased mitochondrial mass, reduced oxidant levels, and enhanced mitochondrial membrane potential. Moreover, DMSO reduced the levels of DNA damage accumulation in vivo and in vitro.
Conclusion
Our data suggest that DMSO may offer a potential pharmacological treatment option for fertility impairment in women undergoing radiotherapy, which warrants further investigation in clinical settings.
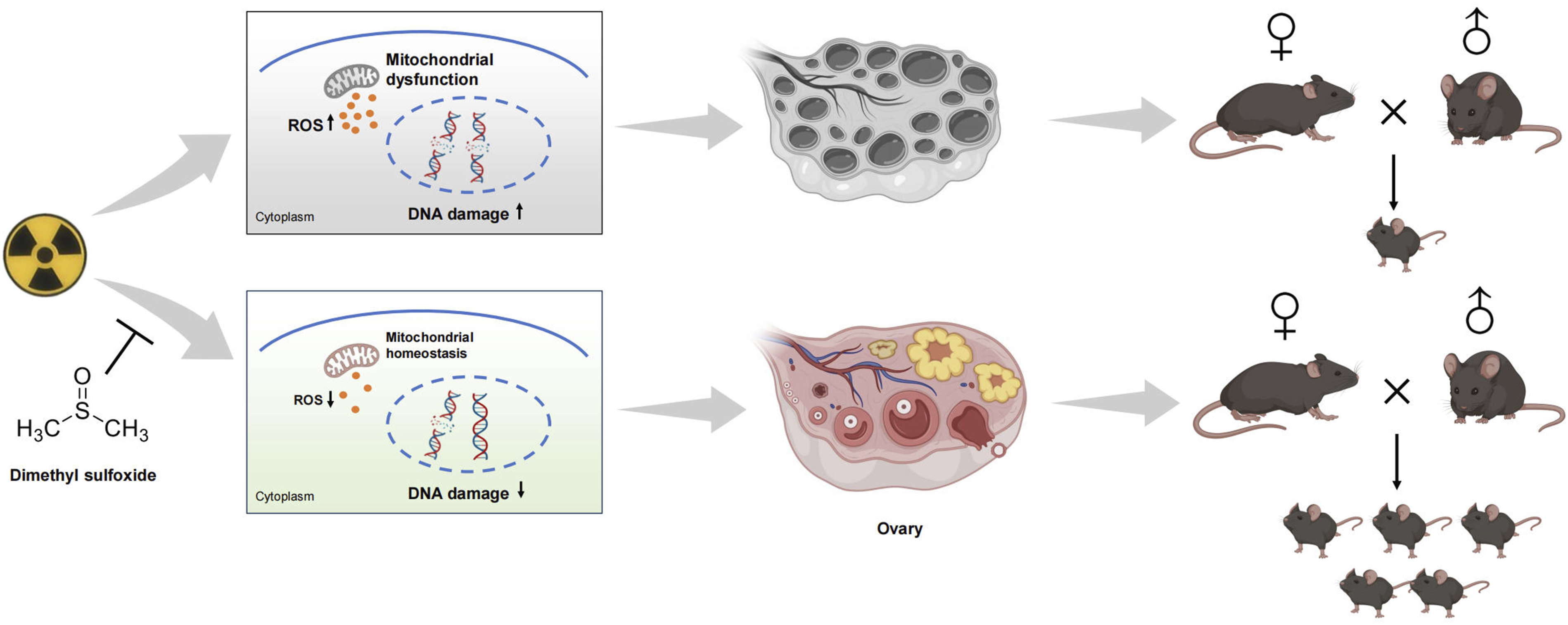
Introduction
The ongoing advancements in comprehensive cancer treatments, including radiotherapy, have significantly boosted survival rates among cancer patients and put forward higher requirements for the quality of life of tumor survivors. For adolescents and young women who need radiotherapy for lower abdominal or pelvic tumors, radiation toxicity to the reproductive system has increasingly attracted attention.1,2 Ovary is the most radiosensitive tissue in female reproductive system. The ovarian function in women is primarily reflected in the number of primordial follicles in the ovaries. Ionizing radiation consumes the follicular reserve in the ovaries, resulting in decreased ovarian steroid secretion, and/or premature ovarian failure (POF) and even infertility.3,4 Generally, cells with high mitotic activity and active DNA replication are more susceptible to radiation damage. However, oocytes seem to be an exception: although arrested at the diplotene stage of the first meiotic division, they are highly sensitive to radiations. 2 Adriaens et al estimated that the dose required to induce permanent ovarian failure would vary from 20.3 Gy at birth to 14.3 Gy at 30 years. 5 Furthermore, the damage to germ cells caused by radiation could be passed down through generations, and whether radiation-induced ovarian injury (RIOI) might affect the well-being of descendants, remains ongoing debate. 6
At present, there are several clinical treatment options to preserve female fertility, such as hormone replacement therapy, cryopreservation of oocytes/embryos/ovarian tissue, and pharmacological protection. Hormone replacement therapy does not fundamentally restore ovarian function. Oocyte collection typically requires several weeks of ovarian stimulation, making it unsuitable for women in need of urgent treatment. Cryopreservation of the ovarian cortical region, which contains 90% of the follicular reserve followed by autologous transplantation, is a potential strategy. However, the safety of this procedure is still a subject of debate.7-9 Technological advancements are therefore required for the pharmacological protection of ovaries from radiotherapy-induced damage.
The mechanisms of radiation-induced cell damage are typically categorized into direct and indirect effects. 10 Direct action refers the radiation energy directly ionizing and exciting biological macromolecules. Indirect action, conversely, refers to the deposition of energy into water molecules, leading to the production of reactive oxygen species (ROS), such as hydroxyl radicals (·OH) and superoxide anions (O2-), which subsequently target biological macromolecules. For low linear energy transfer (LET) radiations involving X and gamma ray, the radiation-induced cell damage is mainly mediated through indirect actions. 11 Both direct and indirect effects can only temporarily produce ROS; the persistent increase in cellular oxidative stress post-irradiation is primarily linked to mitochondrial dysfunction. Ionizing radiation impairs mitochondrial function and increases intracellular oxidative stress, resulting in excessive ROS that can damage the mitochondrial membrane and initiate mitochondrial apoptosis.12-14 Thus, mitigating cellular oxidative stress to maintain mitochondrial function is likely to ameliorate RIOI.
Dimethyl sulfoxide (DMSO) is a broadly utilized radical scavenger that has shown therapeutic benefits across a spectrum of human diseases.15-17 Recently, we reported that DMSO mitigates acute radiation induced toxicity in multiple organs, including the testis, hematopoietic system and oral mucosa.18-20 However, its potential in treating RIOI remains unexplored. In the present study, we investigated whether DMSO could protect mice from ovary injury. Our results demonstrated the radioprotective efficacy of DMSO against RIOI. Notably, DMSO maintains mitochondrial function, decreases cellular oxidative stress, and thereby preserves fertility in female mice by safeguarding the follicular reserve.
Materials and Methods
Mice
Female and male C57BL/6J mice (7–8 weeks or 1 week old) were obtained from SPF (Beijing) Biotechnology Co., Ltd., China. Mice were housed under controlled conditions (constant temperature) with free access to regular chow and sterile water. All procedures were approved by the ethics committee of the PLA Rocket Force Characteristic Medical Center (KY2023012) and followed the Guide for the Care and Use of Laboratory Animals.
Cell Culture
KGN cells (human granulosa cell line, AW-CCH252, AW-CELLS-H0191, China) were maintained in DMEM/F-12 (Gibco) containing 10 % fetal bovine serum (Gibco) and 1× Pen/Strep (Gibco) in 5 % CO2 at 37 °C.
Organ Culture and Drug ex Vivo Treatments
Ovaries from 1-week-old female mice were cultured on polycarbonate membranes (3422, Corning) in MEM (Gibco) containing 10 % fetal bovine serum (Gibco), 25 mM Hepes (Beyotime), and 1 × Pen/Strep (Gibco) in 5 % CO2 at 37 °C. On day 2, ovarian explants were irradiated with 3 Gy. Before irradiation, they were incubated for 2 h with or without 0.1 % DMSO (v/v). After irradiation, the medium was replaced. Oocyte survival was analyzed on day 5 via immunofluorescence.
Radiation Protocols and DMSO Administration
X-rays were generated using the XCELL 225 high energy X-ray irradiator (KUBTEC, USA) at the PLA Rocket Force Characteristic Medical Center (Beijing, China). For lower abdominal irradiation (LAI) at 0.72 Gy/min (225 kV, 13.2 mA) and 55 cm from the source, mice received 5 Gy (LAI). Mice were pre-treated with vehicle (saline) or DMSO (Alfa Aesar) at 5 g/kg via intraperitoneal injection 1 h before irradiation. KGN cells were irradiated with 4.5 Gy and cultured with or without 0.1 % DMSO (v/v) for 2 h prior to irradiation, with medium replacement after radiation.
Assessment of Female Fertility
The fertility of F0 mice was evaluated 15 days post-irradiation. Two females per group were housed with one mature male (n = 5-8/group) for 30 days. Pregnant females were isolated, and litter size (F1) and gestation time (mating to childbirth) were recorded. Pup mortality within 2 weeks was expressed as a percentage. Pregnant females (%) were calculated as the ratio of pregnant females to total females per group. Litter size was the mean of litters from bred females in each group. F1 female fecundity was assessed using the same method.
Hematoxylin and Eosin (H&E) Staining and Follicle Count
Ovarian tissue was fixed, embedded in paraffin, and sectioned at 4 µm thickness using a microtome. Hematoxylin and eosin staining was used to identify ovarian structure, and slides were examined under an optical microscope. Follicle numbers were counted every fifth slice. Ovarian morphology and follicle counts were observed as described previously, 21 follicles were classified into atretic, primordial, primary, secondary, and antral follicles.
Apoptosis Assay
Germ cell apoptosis in ovarian paraffin sections was detected using a TUNEL assay kit (C1098, Beyotime) following the manufacturer instructions. The percentage of TUNEL-positive cells was calculated from five random follicles per section.
Immunohistochemistry (IHC)
Ovary tissue sections were dewaxed with xylene, hydrated with gradient alcohol, and perforated with 0.1 % Triton-X100 after antigen retrieval and endogenous peroxidase elimination using H2O2. Sections were stained with Ki67 antibody (ab16667, Abcam). Specific staining was detected using an IHC kit (PV9001, ZSGB-BIO) and a diaminobenzidine detection kit (ZLI-9019, ZSGB-BIO) according to the manufacturer instructions. Hematoxylin was used for nuclear counterstaining.
Immunofluorescence (IF)
Ovarian explants were fixed in 4 % paraformaldehyde, embedded in paraffin, and sectioned at 4 μm. Fluorescence staining was performed on ovarian sections using the Tyramide signal amplification kit (G1259-50T, Servicebio) according to the manufacturer instructions. Primary antibodies p63 (ab124762, Abcam) and DDX4 (ab13840, Abcam) were used. Images were captured with a fluorescence microscope (Olympus) and quantification was performed in images after appropriate thresholding using the ImageJ software.
Western Blotting Assay
Mouse ovary tissues or cells were lysed with RIPA buffer. Protein concentrations were measured using a BCA kit (P0009, Beyotime). Western blotting was performed according to standard protocols. The antibodies used were: Cleaved caspase-3 (25128-1-AP, Proteintech), γ-H2AX (05-636, Millipore), and GAPDH (10494-1-AP, Proteintech).
ROS Assay
The ROS level in KGN cells was measured using a ROS Assay kit (S0033S, Beyotime) according to the manufacturer instructions and detected by flow cytometry (BD FACSCalibur).
Mitochondrial Function Assays
In accordance with the manufacturer instructions, label the cells using Mito-Tracker Green (C1048, Beyotime), MitoSOX Red (HY-D1055, MCE), and JC-1 (C2003S, Beyotime) probes respectively, and stain the cell nuclei with Hoechst 33342 (C1028, Beyotime). Fluorescence intensity was detected utilizing a Leica confocal microscope (MICA, Leica), and quantification of images was performed following appropriate thresholding using ImageJ software.
Statistical Analysis
Statistical analysis and image rendering were performed using GraphPad Prism free trial. Data were presented as mean ± standard deviation (SD). Animal and cell experiment data were analyzed using t-tests for differences, with p < 0.05 considered significant.
Result
DMSO Preserves Fertility in Mice After Irradiation
we firstly evaluated the effect of DMSO on female fertility. Mice were intraperitoneally injected with 5 g/kg DMSO 1 h prior to 5 Gy lower abdominal irradiation (LAI). Irradiated female mice were then mated with males 14 days later. All unirradiated female mice gave birth to mice, whereas only 62.5 % of female mice in the IR group became pregnant. Compared with the non-irradiated group, the time interval from mating to childbirth in the IR group was extended by 1.3 times, and the litter size was reduced by 49.1 %. In contrast, all DMSO-treated female mice were pregnant, with a shorter conception time and a larger litter size (Figure 1B). We observed that irradiated female mice gave birth to dead infants, an effect partly mitigated by DMSO treatment (Figure 1C). We further raised these female infants and mated them with males. Interestingly, despite neither group of offspring being exposed to radiation, female mice from the DMSO-treated group gave birth to more pups (Figure 1D). These results suggest that DMSO preserves female fertility and the reproductive capacity of their female offspring. Effect of DMSO on fertility of irradiated female mice.
DMSO Maintains the Reserve of Ovarian Follicles
Primordial follicles reside in the ovary and transition to developing and subsequently mature follicles, processes that directly determine the female reproductive life cycle.
22
We assessed morphological changes in ovarian tissue. Treating with DMSO alone exhibited normal ovarian morphology without pathological changes (Figure 2B). Radiation significantly reduced the population of follicles at various developmental stages, particularly primordial follicles, and increased the number of atretic follicles. In contrast, DMSO treatment led to significant improvements: the number of healthy follicles increased at each stage, and fewer atretic follicles were observed (Figure 2C). Effect of DMSO on pathological changes in irradiated ovaries.
DMSO Mitigates Radiation-Induced Follicular Developmental Impairment
Granulosa cells are the main follicular cell type that provides the structural support and microenvironment required for oocyte development.
23
We performed TUNEL staining in ovary sections. DMSO treatment resulted in a 24 % and 25 % decrease in TUNEL-positive follicular cells on days 3 and 7 post-radiation, respectively, compared to ovaries subjected to radiation alone (Figure 3A and B). Similarly, Treatment of irradiated ovaries with DMSO markedly decreased Cleaved Caspase-3 expression (Figure 3C and D). Ki67 expression in granulosa cells marks the initiation of follicle growth.
24
We observed that radiation exposure nearly depleted Ki67-positive follicular cells in developing and mature follicles. However, DMSO pretreatment significantly reversed this depletion, restoring Ki67 expression (Figure 3E and F). These results indicate that DMSO mitigates radiation-induced follicular damage by inhibiting granulosa cell apoptosis and maintaining granulosa cell proliferation. Effect of DMSO on apoptosis and proliferation of ovarian cells after radiation exposure.
DMSO Prevents Follicles Elimination Induced by Irradiation in ex Vivo Ovarian Culture
To investigate the direct effects of DMSO on oocytes following irradiation, we utilized an ex vivo whole-ovary culture system.25,26 We used ovaries from 1-week-old females, a stage characterized by a predominance of primordial oocytes (marked by p63) and a smaller population of oocytes in growing follicles (marked by DDX4) (Figure 4A). After exposure to 3 Gy of radiation, we observed a significant decrease in the survival of both primordial and developing oocytes. Compared to the irradiated (IR) groups, DMSO treatment improved the survival of primordial oocytes by 1.6 times and developing oocytes by 1.4 times (Figure 4B-D). DMSO mitigates radiation-induced damage to ovaries in an explant culture system.
DMSO Prevents Radiation-Induced Mitochondrial Dysfunction
Mitochondria are considered major cytoplasmic targets for oxidative stress and serve as precursors to cellular damage.12,27 To investigate whether DMSO affects radiation-induced mitochondrial dysfunction in KGN cells, we stained the cells with Mito-Tracker, JC-1, and MitoSOX probes. Compared with radiation treatment alone, radiation combined with DMSO treatment could significantly reduce mitochondrial dysfunction, as evidenced by increased mitochondrial mass, enhanced mitochondrial membrane potential, and decreased levels of mitochondrial oxidants (Figure 5A and B). Moreover, we monitored intracellular ROS levels using the DCFH-DA probe. The results showed that intracellular ROS levels were significantly elevated 24 h after IR, while the DMSO pre-treatment group exhibited decreased ROS levels compared to the IR-only group (Figure 6A and B). Effect of DMSO on mitochondrial function following radiation exposure. Effect of DMSO on radiation-induced ROS and DNA damage
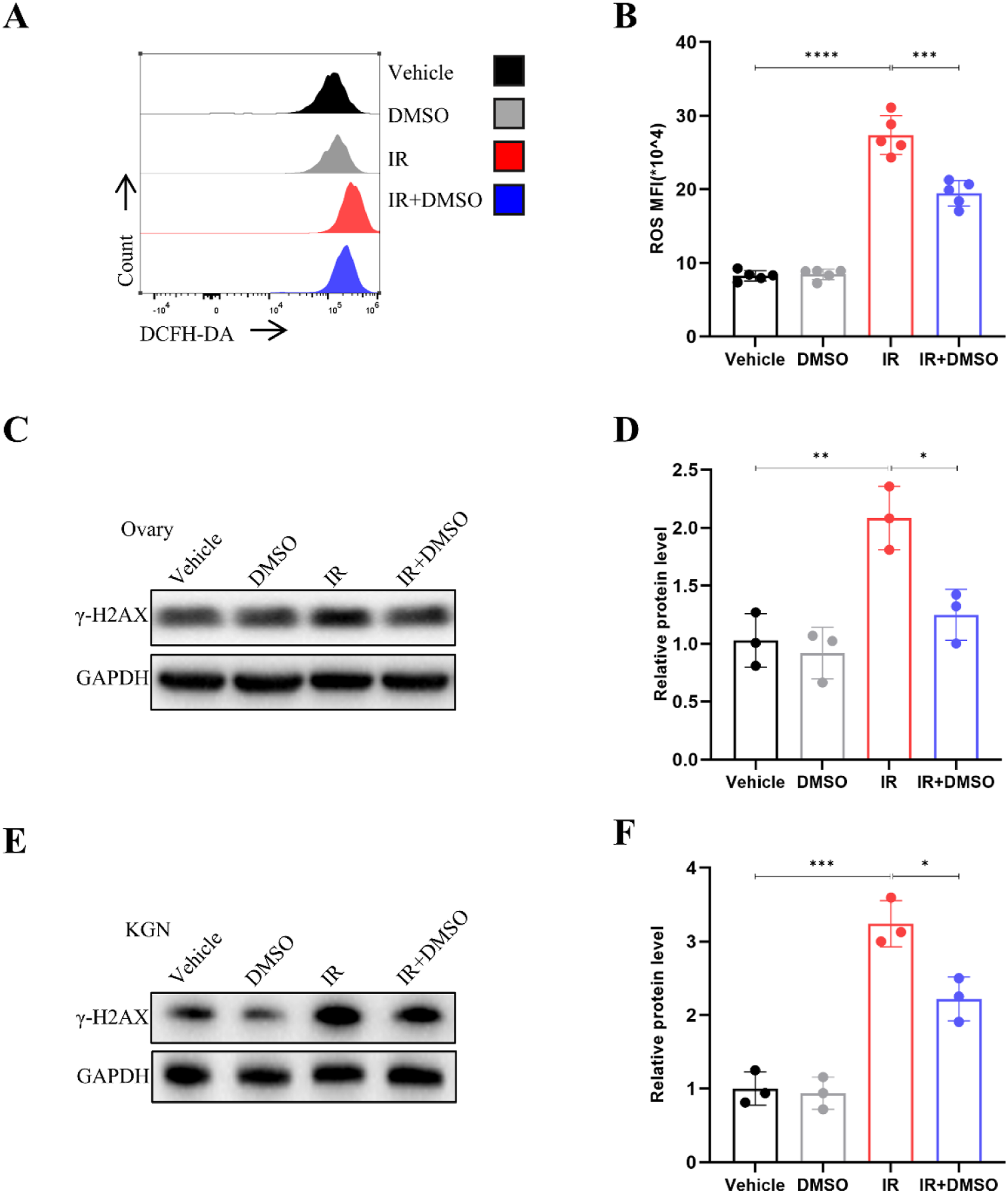
DSMO Alleviates DNA Damage Accumulation
γ-H2AX foci at later phases following radiation are considered surrogates for unresolved DSBs. 27 To clarify determine DMSO can reduce DNA damage, we examined unrepaired DNA DSBs in the ovaries by γ-H2AX immunostaining. As expected, DMSO dramatically decreased γ-H2AX expression in the ovaries of treated mice 3 days post-radiation (Figure 6C and D). We further analyzed γH2AX expression in KGN cells in vitro. Consistently, immunoblotting revealed that DMSO treatment (0.1 %, v/v) reduced γ-H2AX expression under irradiation(Figure 6E and F).
Discussions
To date, radiotherapy remains a common treatment for tumors. However, due to the ill-defined margins of malignant tumors and the movement of organs, damage to healthy tissues from radiation is often unavoidable. 28 For young female patients, whole abdominal or pelvic irradiation may expose the ovaries to radiation, potentially leading to ovarian failure and impacting fertility. Wallace et al. estimated that the dose required to destroy 50% of primordial follicles (LD50) would be less than 2 Gy. 2 Moreover, the non-renewable nature of the oocyte pool makes it particularly vulnerable. 29 Therefore, minimizing the toxic effects of radiotherapy on the ovaries and reducing the risk of premature ovarian failure (POF) and infertility is crucial. In this study, we utilized a mouse model of radiation-induced ovarian damage to explore the protective effects of dimethyl sulfoxide (DMSO) against ovarian injury. By evaluating fertility and performing histopathological assessments, we aimed to contribute a fresh pharmacological insight into the preservation of fertility.
Over the last decades, numerous approaches have been employed to prevent radiation-induced premature ovarian failure (POF) and preserve fertility. Techniques such as oocyte cryopreservation and embryo cryopreservation require assisted reproduction, including ovulation induction and in vitro fertilization.30,31 On the other hand, ovarian tissue cryopreservation involves surgical procedures and subsequent autologous transplantation. 32 Gonadotropin-releasing Hormone (GnRH) analogues, both agonists and antagonists, are potential treatments for infertility caused by chemotherapy. However, according to clinical practice guidelines, GnRH analogues do not shield the ovaries from the gonadotoxic effects of radiotherapy, and so they are not suggested for female patients scheduled to receive pelvic, abdominal, or total body irradiation.33-35 Indeed, oocyte and embryo cryopreservation are not suitable for girls and adolescents. 9 Consequently, the development of pharmacological protections, which are simpler procedures that can be used alone or in conjunction with fertility cryopreservation, holds significant clinical importance. DMSO’s radioprotective effects on various organs have been previously demonstrated.18-20 Our study clearly indicates that a single dose of DMSO administered within a short period prior to radiation (1 hour) is effective. This is particularly crucial because for patients, the sooner radiotherapy can be initiated after diagnosis, the better, and fertility must be rapidly and effectively preserved concurrently.
It’s worth noting that the DMSO dose examined in this study (5 g/kg) is equivalent to approximately 44 ml for a 60 kg individual. 36 Clinical studies have shown that DMSO does not induce any side effects when administered to humans at doses of 20–60 ml per day, which bodes well for translating this research into clinical practice.37,38 Moreover, DMSO is stable at room temperature and cost-effective. Therefore, DMSO emerges as a promising candidate for further development in human radiotherapy applications.
The fertility assessment experiment showed that irradiated females exhibited a significant decrease in both conception rates and litter size, with DMSO capable of providing partial relief. Female fertility is closely related to the size of the ovarian primordial follicle reserve, which forms during fetal development and completes shortly after birth. 29 In vitro ovarian culture experiments indicated that ionizing radiation led to a substantial depletion of the primordial follicle pool, likely due to direct damage to these early-stage follicles. Among the developing follicles, not all categories are affected equally by radiation; there was a significant decrease in the number of preantral follicles in irradiated ovaries, while the population of antral follicles remained unchanged compared to control ovaries. This aligns with previous research showing the high radiosensitivity of early follicular stages and the low radiosensitivity of more mature follicles.39,40 The reduction in the count of secondary follicles may stem from either the depletion of primordial follicles or the normal recruitment process of secondary follicles. In addition to follicular depletion, irradiated ovaries also exhibited a higher incidence of atretic and unhealthy follicles. DMSO treatment helps to preserve the primordial follicle pool, reducing the number of abnormal follicles and thereby extending the lifespan of the irradiated ovaries.
Whether the damage of germ cells caused by radiation can be inherited across generations has been controversial. Meredith et al. examined 130 children born to parents who were cleanup workers or exposed to occupational and environmental ionizing radiation following the Chernobyl nuclear accident. They discovered that the total de novo mutations in children did not increase when their mothers had been exposed to ionizing radiation prior to conception (mean = 19 mGy, range = 0-550 mGy). 41 Wang et al. indicated that DNA double-strand breaks in oocytes can either be properly repaired or eliminated, thus preventing transgenerational effects from maternal DNA damage. 42 However, some cohort studies have shown that female adolescent and young adult cancer survivors received radiotherapy are at higher risk of adverse birth outcomes, including preterm birth and low birth weight.43,44 Our study observed that female mice exposed to radiation exhibited a higher rate of stillbirths. Interestingly, the female offspring of irradiated mice, despite not being exposed to radiation themselves, showed a significant decrease in litter size when mated with males. Our study supports that the radiation damage of oocytes can be inherited across generations, but the specific mechanism warrants further investigation.
Oxidative stress is considered to be the most important pathogenic injury mediating the toxic effect of radiation on normal tissues. Ionizing radiation (IR) triggers water radiolysis, leading to a substantial generation of ROS that overwhelms antioxidant defenses, disrupts redox balance, and ultimately results in damage to DNA, lipids, and proteins. 11 DNA double-strand breaks (DSBs), primarily induced by hydroxyl radicals (·OH), are the most severe type of damage among the genomic toxicities caused by IR. 45 DMSO, a widely used radical scavenger, consistently demonstrates a reduction in DSBs in our current work, both in vivo and in vitro, aligning with previous studies.18,19 Mitochondria, the main sites of oxidative phosphorylation and ATP formation in cells, are also the primary source of endogenous oxygen free radicals, thus playing a crucial role in the indirect effects of radiation damage. 46 Studies have confirmed that inhibiting mitochondrial dysfunction has a therapeutic effect on radiation injury.12,47,48 In our study, DMSO preserves mitochondrial function, and it is ineffective when administered after irradiation. We speculate that DMSO mitigates the consumption of intracellular reductive substances by scavenging ROS produced by radiation, preserving redox balance, and indirectly protecting mitochondria, rather than directly, which warrants further investigation.
In conclusion, our research illustrates that DMSO attenuates radiation-induced ovarian damage in a mouse model. DMSO specifically curbs the accumulation of DNA damage, maintains mitochondrial function, and prevents granulosa cell apoptosis, thereby preserving the primordial follicle reserve. This suggests that DMSO could offer novel insights into pharmacological strategies for maintaining the fertility of female cancer patients. However, the precise mechanism by which DMSO acts on radiation-induced ovarian injury (RIOI) and its regulatory effects on the cytotoxicity of radiotherapy across various cancer models are not yet fully understood and warrant further investigation.
Supplemental Material
Supplemental material - DMSO Protects Against Radiation-Induced Ovarian Injury by Preserving Mitochondrial Function and Alleviating DNA Damage
Supplemental material for DMSO Protects Against Radiation-Induced Ovarian Injury by Preserving Mitochondrial Function and Alleviating DNA Damage by Jie Zhou, Renjun Peng, Zhongmin Chen, Yiying Zhang, Jing Dai, Suhe Dong, Huijie Yu, Sinian Wang, Qisheng Jiang, Wei Li, Bowen Shi, Fengsheng Li, Wei Wang in Dose-Response
Supplemental Material
Supplemental material - DMSO Protects Against Radiation-Induced Ovarian Injury by Preserving Mitochondrial Function and Alleviating DNA Damage
Supplemental material for DMSO Protects Against Radiation-Induced Ovarian Injury by Preserving Mitochondrial Function and Alleviating DNA Damage by Jie Zhou, Renjun Peng, Zhongmin Chen, Yiying Zhang, Jing Dai, Suhe Dong, Huijie Yu, Sinian Wang, Qisheng Jiang, Wei Li, Bowen Shi, Fengsheng Li, Wei Wang in Dose-Response
Footnotes
Acknowledgement
I would like to express my gratitude to my fellow students Yanqing Liu, Xinglei Song.
Author Contributions
W.W., F.S.L. and R.J.P. designed the study. J.Z., R.J.P., Z.M.C., Y.Y.Z., J.D., S.H.D., H.J.Y., S.N.W., W.L., performed laboratory experiments. J.Z. and R.J.P., analyzed the data and prepared the manuscript. F.S.L. and Q.S.J. directed final version of all contents. All authors reviewed and approved the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Military Research Program (HQKY22002).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.