
Abstract
Dietary restriction (DR) has been extensively investigated for its capacity to extend lifespan and mitigate age-associated pathologies. In the context of ionizing radiation (IR), DR—implemented through caloric reduction, selective nutrient restriction, or fasting-based regimens—has emerged as a significant modulator of both cellular and systemic responses to radiation exposure. Growing evidence suggests that DR influences radiation outcomes in a highly context-dependent manner, exerting either protective or sensitizing effects depending on tissue type, metabolic status, timing of intervention, and radiation conditions. This review synthesizes current knowledge regarding the mechanisms through which DR modulates responses to IR, with particular emphasis on oxidative stress regulation, inflammatory signaling, mitochondrial homeostasis, DNA damage responses, cellular quality-control pathways, and carcinogenic processes. We further discuss how distinct dietary paradigms interact with nutrient-sensing and stress-adaptive networks to influence radiation sensitivity and tissue recovery. Emerging evidence indicates that the timing, intensity, and composition of dietary interventions critically shape tissue responses to IR, underscoring the importance of considering nutritional status in radiation risk assessment and therapeutic planning. By integrating established findings with emerging mechanistic insights, this review highlights the translational potential of DR for improving outcomes in radiation oncology and reducing radiation-induced injury.
Keywords
Note on Terminology
Dietary Restriction (DR), in the context of this review, refers to the deliberate reduction of total dietary amount, caloric intake, or specific dietary components such as macronutrients or particular amino acids, without causing malnutrition or nutritional deficiencies. DR encompasses various nutritional regimens, including overall dietary reduction, caloric restriction (CR), selective restriction of specific nutrients (such as proteins or amino acids), and fasting-based approaches such as intermittent fasting (involving regular periods of fasting interspersed with periods of normal intake) or time-restricted feeding (limiting daily food intake to a specific time window). These interventions aim to maintain metabolic balance while activating stress-responsive and homeostatic pathways. DR has been shown to modulate key cellular mechanisms involved in oxidative stress, DNA repair, and inflammation, which are critically relevant to responses to ionizing radiation (IR). Although pharmacological agents that mimic CR are not classified as DR, their potential to reproduce some beneficial effects of CR will also be briefly addressed in this review.
1. Introduction
1.1. Dietary Restriction: Biological Foundations and Systemic Adaptation
Dietary restriction (DR) broadly describes nutritional strategies that reduce overall caloric intake, selectively limit specific macronutrients or amino acids, or modify feeding schedules, while preserving essential micronutrient sufficiency and preventing malnutrition. This framework encompasses caloric restriction (CR), protein restriction (PR), amino acid restriction (AAR), and fasting-based approaches including intermittent fasting (IF) and time-restricted feeding (TRF).1,2 Since the pioneering experiments by McCay and colleagues in the 1930s first demonstrated lifespan extension in rodents subjected to reduced caloric intake, 3 DR has become the most robust and evolutionarily conserved non-pharmacological intervention shown to extend lifespan and improve healthspan across diverse species. Research spanning yeast, invertebrates, rodents, and non-human primates indicates that DR does more than reduce energy consumption; it induces systemic remodeling of nutrient-sensing and stress-response pathways, leading to improved metabolic efficiency, enhanced cellular maintenance, and delayed progression of age-associated pathologies.4-6
Mechanistically, DR modulates conserved metabolic signaling networks. Central features include suppression of insulin/insulin-like growth factor-1 (IGF-1) signaling and the mammalian target of rapamycin (mTOR) activity, together with activation of AMP-activated protein kinase (AMPK) and sirtuin pathways.7-9 These coordinated shifts reorient cellular physiology away from growth and biosynthesis toward repair, stress adaptation, and quality control. Such reprogramming intersects with several hallmarks of aging, particularly mitochondrial dysfunction, genomic instability, impaired proteostasis, and chronic inflammation, thereby positioning DR as a systemic regulator of longevity-associated biology.10,11
1.2. Dietary Restriction as a Modulator of Ionizing Radiation Responses
Exposure to ionizing radiation (IR) provokes complex biological injury characterized by DNA double-strand breaks, oxidative damage, mitochondrial disruption, inflammatory cascades, and regulated cell death. These processes contribute to both acute tissue toxicity and delayed consequences such as fibrosis, degenerative changes, and carcinogenesis.12,13 Notably, many of these radiation-induced alterations overlap with molecular features of aging, including genomic instability, impaired protein homeostasis, mitochondrial dysfunction, and sustained inflammation. This convergence suggests that interventions enhancing cellular resilience and maintenance may influence radiation responses. Within this framework, DR represents a metabolic intervention with potential to reshape IR outcomes through several complementary mechanisms: reduction of baseline oxidative stress, reinforcement of antioxidant defenses, enhancement of DNA repair and cellular quality-control pathways, and modulation of inflammatory and immune signaling that drives late radiation injury.14-16 Importantly, DR effects appear context-dependent. Under nutrient limitation, normal cells often adopt a quiescent and stress-tolerant phenotype, whereas cancer cells—frequently constrained by oncogenic signaling and metabolic rigidity—may lack equivalent adaptive capacity.17,18 This divergence underlies the concept of differential stress resistance, whereby DR may confer protection to normal tissues while simultaneously increasing tumor susceptibility to radiation-associated stress.
1.3. Purpose, Scope, and Novelty of This Review
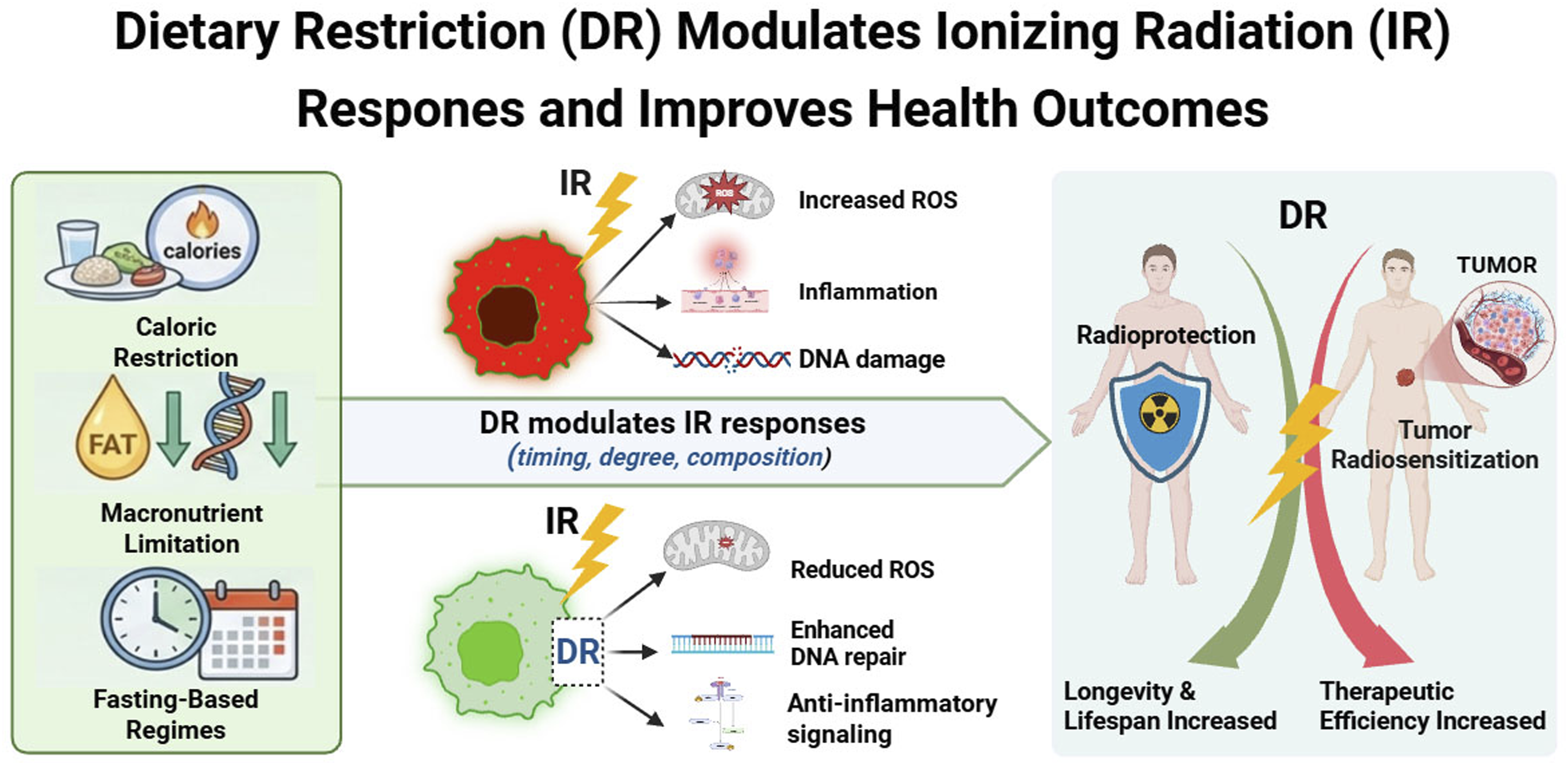
Although studies linking DR to radiation biology are expanding rapidly, findings remain fragmented across aging research, metabolism, oncology, and radiobiology. The present review integrates these domains by positioning DR as a systems-level metabolic intervention with emerging relevance to IR. We (i) outline principal DR regimens and their shared conceptual basis, (ii) synthesize molecular and systemic mechanisms through which DR reinforces cellular maintenance programs, and (iii) critically evaluate available evidence for both radioprotective and radiosensitizing effects in normal and malignant contexts. By connecting nutrient-sensing regulation with radiation injury and therapeutic response, we propose DR as a metabolic framework capable of enhancing radiation resilience while improving therapeutic index. Key knowledge gaps and priorities for translational development are also discussed. A schematic overview is provided in Figure 1. Conceptual framework linking dietary restriction to ionizing radiation responses. Dietary restriction (DR), encompassing caloric, protein, amino acid, and fasting-based regimens, induces systemic reprogramming of conserved nutrient-sensing pathways, including suppression of IGF-1 and mTOR signaling and activation of AMPK and sirtuin networks. These shifts enhance cellular maintenance programs—reducing oxidative stress and inflammation while promoting DNA repair, autophagy, and metabolic flexibility. Because many radiation-induced injuries overlap with hallmarks of aging, DR-mediated adaptations modulate responses to ionizing radiation. In normal tissues, DR promotes stress tolerance and radioprotection, whereas metabolically constrained tumor cells exhibit reduced adaptive capacity, resulting in radiosensitization. This differential stress response highlights DR as a potential metabolic strategy to improve radiation resilience and therapeutic index. Created in BioRender. Dechsupa, N. (2026) https://BioRender.com/wambtfp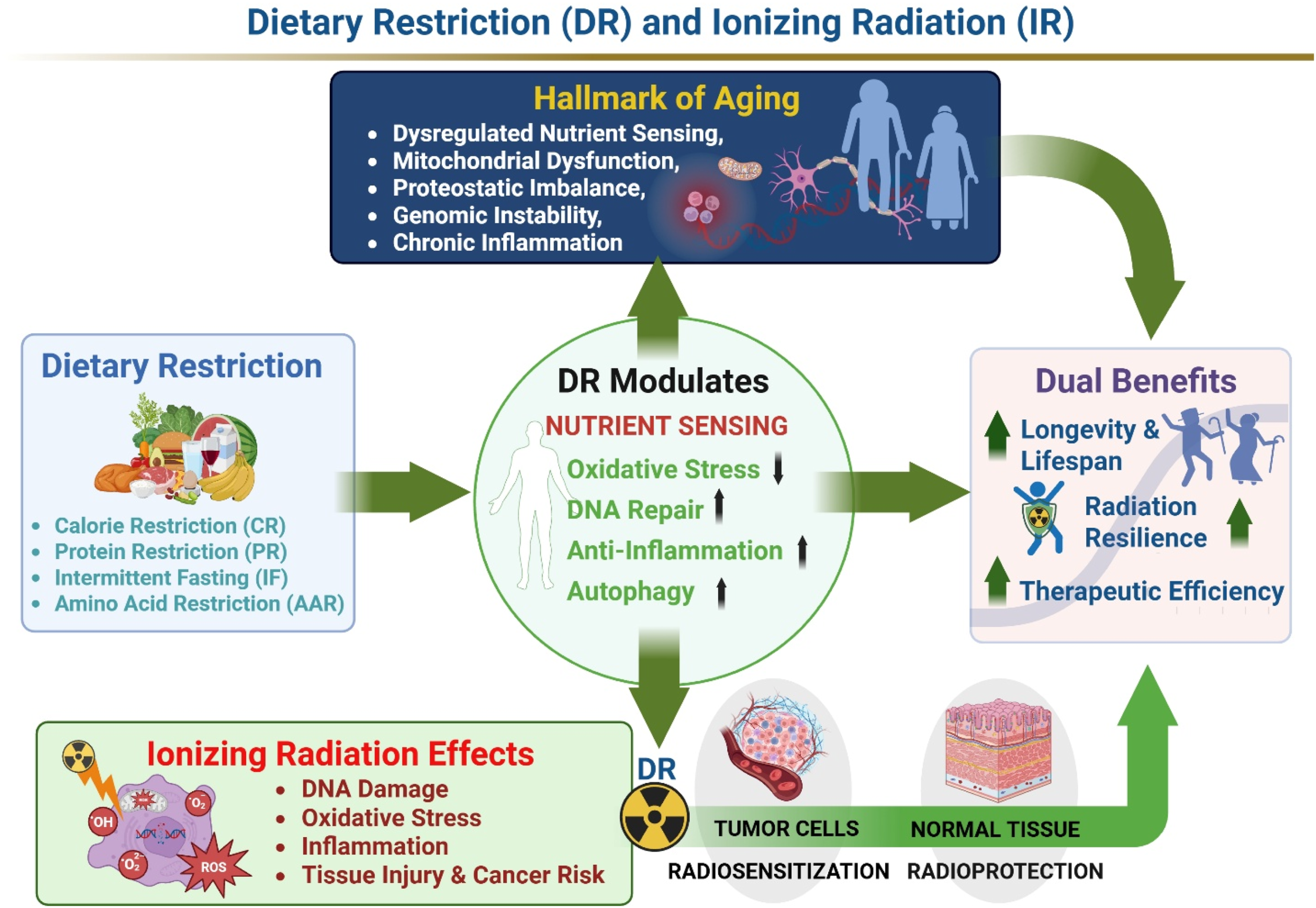
2. Biological Effects and Mechanisms of Dietary Restriction
2.1. Overview of Dietary Restriction Regimens
Dietary restriction (DR) encompasses a spectrum of nutritional interventions that reduce energy intake, modify nutrient composition, or alter feeding patterns without causing malnutrition. Major DR paradigms include caloric restriction (CR), protein or amino acid restriction (PR/AAR), and fasting-based approaches such as intermittent fasting (IF) and time-restricted feeding (TRF). Although these strategies differ in implementation, they consistently induce overlapping metabolic and cellular adaptations associated with improved stress resistance and metabolic homeostasis.
CR, typically defined as a sustained reduction in caloric intake while maintaining adequate micronutrient supply, remains the most extensively studied DR modality and has been associated with lifespan extension and delayed onset of age-related disorders across multiple experimental systems.4,19 In contrast, PR and AAR primarily target nutrient composition rather than total energy intake, highlighting the importance of amino acid signaling in regulating metabolism, stress responses, and longevity-associated pathways.20,21 Restriction of specific amino acids, particularly methionine and branched-chain amino acids, has been shown to reproduce several metabolic adaptations observed under CR, including improved insulin sensitivity, reduced oxidative stress, and altered nutrient-sensing signaling. These findings emphasize that modulation of amino acid availability itself, particularly through mTOR- and insulin/IGF-1–related nutrient-sensing pathways, can independently influence metabolic adaptation and stress resistance.22-24
Fasting-based regimens, including IF and TRF, differ from continuous dietary restriction by emphasizing periodic cycles of feeding and fasting. These approaches promote metabolic switching from glucose utilization toward fatty acid oxidation and ketone body production, thereby activating adaptive stress-response pathways and enhancing metabolic flexibility. This adaptive metabolic reprogramming is thought to contribute to enhanced cellular resilience and improved tolerance to metabolic and oxidative stress.25-28 Importantly, several beneficial effects of IF and TRF appear to occur independently of overall caloric reduction, suggesting that temporal regulation of nutrient availability itself exerts significant biological effects. 29
In addition to restrictive paradigms, emerging nutrient-directed strategies aimed at restoring metabolic balance and supporting tissue recovery have also gained interest. Such approaches, including “dietary inversion” concepts involving selective replenishment of amino acids, micronutrients, and antioxidant-associated nutrients, may complement restrictive interventions by mitigating treatment-associated metabolic dysfunction and normal-tissue injury.30-33
Despite their methodological differences, these dietary and nutrient-directed interventions converge on several conserved biological responses, including suppression of insulin/IGF-1 and mTOR signaling, activation of AMPK and sirtuin pathways, enhancement of autophagy and mitochondrial quality control, and attenuation of oxidative and inflammatory stress. Because these interconnected nutrient-sensing and stress-adaptive pathways critically influence radiation-induced tissue injury and recovery, they provide an important mechanistic framework for understanding how DR modulates responses to ionizing radiation.
2.2. Integrated Nutrient-Sensing and Stress-Response Networks Relevant to IR
Dietary restriction (DR) exerts its biological effects through coordinated modulation of conserved nutrient-sensing and stress-response pathways that regulate cellular metabolism, energy balance, and adaptive stress responses. 34 Rather than functioning independently, signaling networks involving insulin/IGF-1 (IIS), mTOR, AMPK, and sirtuins operate as an integrated regulatory system that dynamically adjusts cellular priorities according to nutrient availability.35-37 Under nutrient-rich conditions, IIS and mTOR signaling promote anabolic metabolism, protein synthesis, and cellular proliferation.38-40 In contrast, nutrient limitation suppresses growth-promoting pathways while activating AMPK- and sirtuin-dependent programs associated with metabolic adaptation, autophagy-mediated quality control, mitochondrial maintenance, and cellular stress resistance.41-44
Collectively, these signaling adaptations shift cellular physiology away from growth-oriented processes toward maintenance and repair. Reduced IIS and mTOR activity limit anabolic demand and enhance cellular quality-control mechanisms, whereas AMPK activation promotes energy conservation, fatty acid oxidation, and mitochondrial homeostasis.41,45,46 Concurrently, increased NAD+ availability during nutrient limitation enhances sirtuin activity, linking metabolic status to transcriptional reprogramming, antioxidant defense, and genomic maintenance.47,48 These adaptive responses also contribute to preservation of genome integrity and enhanced tolerance to oxidative and genotoxic stress.49,50 Extensive crosstalk among these pathways coordinates cellular responses to energetic and oxidative stress and contributes to improved metabolic flexibility and stress tolerance. 8
Importantly, many of these pathways directly intersect with biological processes implicated in ionizing radiation (IR) responses.51,52 Radiation exposure induces oxidative stress, mitochondrial dysfunction, inflammatory signaling, and DNA damage, all of which are influenced by nutrient-sensing networks regulated during DR. 53 This mechanistic overlap provides a biological basis for the capacity of DR to modulate radiation sensitivity, tissue recovery, and long-term radiation-associated pathology.
2.3. Cellular Maintenance and Stress-Adaptive Processes Influencing Radiation Responses
The coordinated nutrient-sensing adaptations induced by DR ultimately converge on a set of cellular maintenance and stress-response processes that are highly relevant to radiation biology. These include regulation of oxidative stress, preservation of mitochondrial integrity, modulation of inflammatory signaling, enhancement of DNA damage surveillance and repair capacity, and activation of autophagic quality-control pathways. Together, these mechanisms promote cellular resilience under conditions of metabolic and environmental stress. 11
One of the most consistent consequences of DR is improved redox homeostasis. Nutrient limitation enhances mitochondrial efficiency, reduces excessive reactive oxygen species (ROS) production, and strengthens endogenous antioxidant defenses through activation of stress-responsive transcriptional programs.54-57 At the same time, DR supports mitochondrial quality control through enhanced mitophagy and metabolic remodeling, thereby limiting accumulation of dysfunctional mitochondria that contribute to oxidative injury and inflammatory activation.58-61 DR also modulates inflammatory and immune signaling pathways implicated in both aging and radiation injury. Reduced nutrient availability suppresses chronic pro-inflammatory signaling, partly through inhibition of NF-κB activity and improved metabolic regulation, while promoting cellular programs associated with stress adaptation and tissue maintenance.62,63 In parallel, DR enhances genomic stability by supporting DNA damage surveillance and repair pathways and by reducing oxidative burden that contributes to DNA lesions.64-67 These protective adaptations are closely linked to activation of autophagy, a conserved cellular recycling process that removes damaged proteins and organelles while maintaining metabolic homeostasis under stress conditions. 68
Because oxidative stress, mitochondrial dysfunction, chronic inflammation, impaired DNA repair, and defective autophagy are central determinants of radiation-induced tissue injury and carcinogenesis, 69 these DR-responsive pathways provide a mechanistic framework through which dietary interventions may influence radiation sensitivity and therapeutic outcomes. These mechanistic interactions form the basis for growing evidence that DR can differentially modulate radiation responses in both normal and malignant tissues, as discussed in the following sections.
3. Impact of Dietary Restriction on Responses to Ionizing Radiation: Phenomena and Mechanisms
DR can modify biological responses to IR across multiple scales—from acute tissue injury and hematopoietic recovery to long-term carcinogenesis and therapeutic outcomes in radiotherapy. Rather than reiterating the core nutrient-sensing and cellular stress-response mechanisms detailed in Sections 2, this section synthesizes the radiation-facing evidence and highlights where dietary state reshapes canonical IR injury pathways, including oxidative and inflammatory cascades, DNA damage processing, tissue microenvironmental remodeling, and immune competence. For clarity, the literature is organized into (i) calorie-restricted diets (CRD), which primarily reduce total energy intake, and (ii) other restriction approaches (ORA), encompassing fasting-based regimens, low-carbohydrate/high-fat strategies, and targeted nutrient modulation. We first summarize classical observations establishing diet–radiation interactions, then map contemporary evidence and delineate mechanistic principles and translational boundaries for each category.
3.1. Classical Findings of Diet–Radiation Interactions
Early diet–radiation studies provided foundational evidence that nutritional state modulates radiation tolerance, tissue recovery, and late carcinogenic outcomes. In the 1970s, studies in irradiated rodents indicated that dietary composition could strongly influence survival, as exemplified by marked survival differences under altered electrolyte balance regimens. 70 Clinical observations also suggested a dietary role in modulating radiation injury. In a cohort of 44 children who received whole-abdominal radiation therapy at the Institut Gustave-Roussy between 1961 and 1972, 5 of 14 long-term survivors (36%) developed severe delayed radiation enteritis with small bowel obstruction. Histological findings included villus blunting, lymphatic dilatation, and inflammatory infiltrates. Remarkably, all affected patients showed sustained clinical and radiologic recovery under a fractionated, low-residue, low-fat, gluten-free, and milk-free diet, with no recurrence of enteritis. 71 Parallel work in animal models established that DR influences hematopoietic outcomes following radiation exposure, including bone marrow death and recovery. Regarding tumorigenesis, the most consequential late effect of radiation, the classical studies of Tannenbaum and colleagues in the 1940s provided convincing evidence that CR suppresses spontaneous tumor development.72-76 However, it was not until the 1980s that Gross and Dreyfuss demonstrated that restricting food intake could also suppress radiation-induced tumors in animal models, marking a major milestone.
In Sprague-Dawley rats subjected to fractionated total-body X-irradiation (1.5 Gy × 5 weekly), tumor incidence rose dramatically, from 22% to 93% in females and from 5% to 59% in males. 77 In rats fed ad libitum (AL) (5–6 pellets/day), all irradiated animals (14 females, 9 males) developed tumors or leukemia (100%), with average onset at 13.7 months in females and 13.4 months in males. By contrast, in rats maintained on a restricted diet (2 pellets/day), only 9/29 females (31%) developed tumors at 18.2 months, and 1/15 males (6.7%) developed a tumor at 9 months. In full-fed females, most tumors were benign, whereas in males, 1 leukemia and 8 tumors developed, 6 malignant and 1 benign. In the restricted group, female tumors were 4 malignant and 5 benign, and the single male tumor was a sarcoma. These findings clearly demonstrated that DR strongly suppressed radiation-induced tumorigenesis in both sexes. 78 Two years later, the same group extended their work to C3H(f) mice, a strain in which spontaneous leukemia is rare (<0.5%) but markedly increased by total-body irradiation. 79 Under the same fractionated gamma-irradiation regimen (1.5 Gy × 5 weekly), AL-fed mice (4.5–5.4 g/day) exhibited a high incidence of leukemia: 31/58 females (53.4%) and 24/50 males (48%), with an average onset at 8 months. In contrast, restricted-fed mice (2 g/day, then 3 g on alternate days) showed dramatically lower incidence, with only 2/55 females (3.6%) and 1/36 males (2.8%) developing leukemia at average ages of 9 and 12 months, respectively. Leukemia was predominantly lymphatic or lymphoblastic, with extensive infiltration of the thymus, lymph nodes, spleen, liver, kidneys, and bone marrow, and frequent leukemic involvement of the peripheral blood. Together, these results demonstrated that DR can strongly suppress both radiation-induced tumorigenesis and leukemia in rodents, although the underlying mechanisms remain to be elucidated.
These classical findings led to later experimental studies that expanded the scope of DR-mediated radiation effects beyond classical DR, including CRD, LHF, and ORA interventions. Emerging evidence indicates that dietary composition, restriction timing, and intermittent fasting protocols can influence radiation sensitivity in both normal and cancerous tissues. These studies reveal context-dependent effects, showing that DR can either mitigate or, under specific conditions, enhance IR-induced cellular damage, emphasizing the importance of diet type, duration, and biological context. These studies also lay the groundwork for subsequent mechanistic studies.
3.2. Calorie-Restricted Diets (CRD) and IR
3.2.1. Introduction
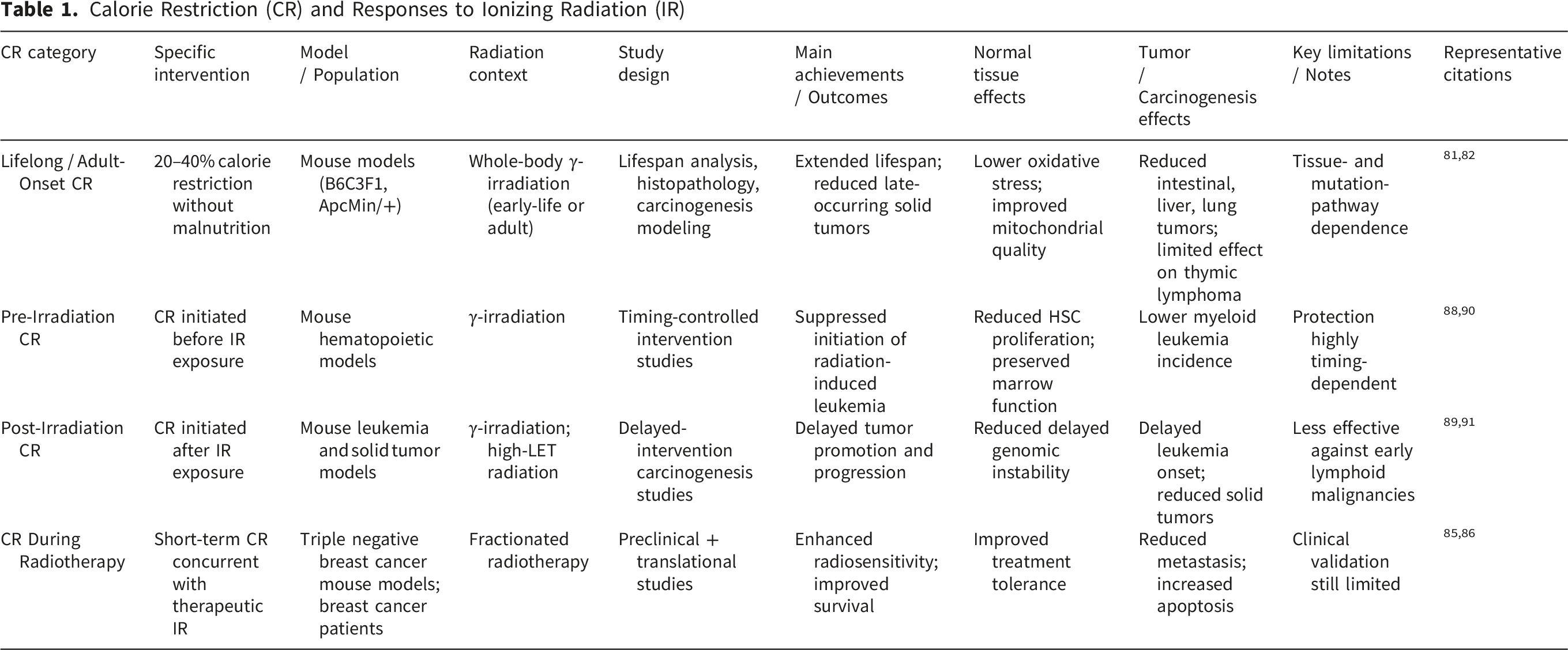
Calorie restriction (CR) has been extensively investigated as a modifier of biological responses to IR, particularly in relation to carcinogenesis, immune function, and tissue resilience. Epidemiological and experimental evidence consistently shows that early-life radiation exposure markedly increases lifetime cancer risk, especially for intestinal, hepatic, and lymphoid malignancies,80-83 with younger organisms displaying heightened susceptibility due to developmental differences in proliferation and DNA damage responses. 84 Within this context, CR has emerged as one of the most robust non-pharmacological interventions capable of modifying radiation-associated carcinogenesis. Preclinical and translational studies demonstrate that CR can enhance radiotherapy efficacy while preserving systemic immune competence. In aggressive tumor models such as triple-negative breast cancer, CR synergizes with IR to suppress metastatic progression, reduce lung tumor burden, and improve survival through downregulation of IGF-1R signaling and increased apoptosis. 85 CR also remodels post-irradiation immune responses, limiting regulatory T-cell expansion, increasing CD8+: Treg ratios, and enhancing CD8+ T-cell activation. 86 Complementary experimental work further shows that CR can amplify immune responsiveness to low-dose radiation, including enhanced splenic T-cell proliferation in a genotype-dependent manner. 87
Strong protective effects of CR against radiation-induced carcinogenesis have been documented across multiple animal models. Early studies demonstrated marked reductions in radiation-induced leukemia incidence, 79 with subsequent work confirming delayed onset and improved survival in myeloid leukemia models.88,89 The timing of CR relative to irradiation is a critical determinant of outcome, as pre-versus post-irradiation restriction differentially affects tumor initiation and promotion. 90 Even mild dietary restriction can enhance radioadaptive responses and improve hematopoietic recovery following low-dose priming irradiation. 91
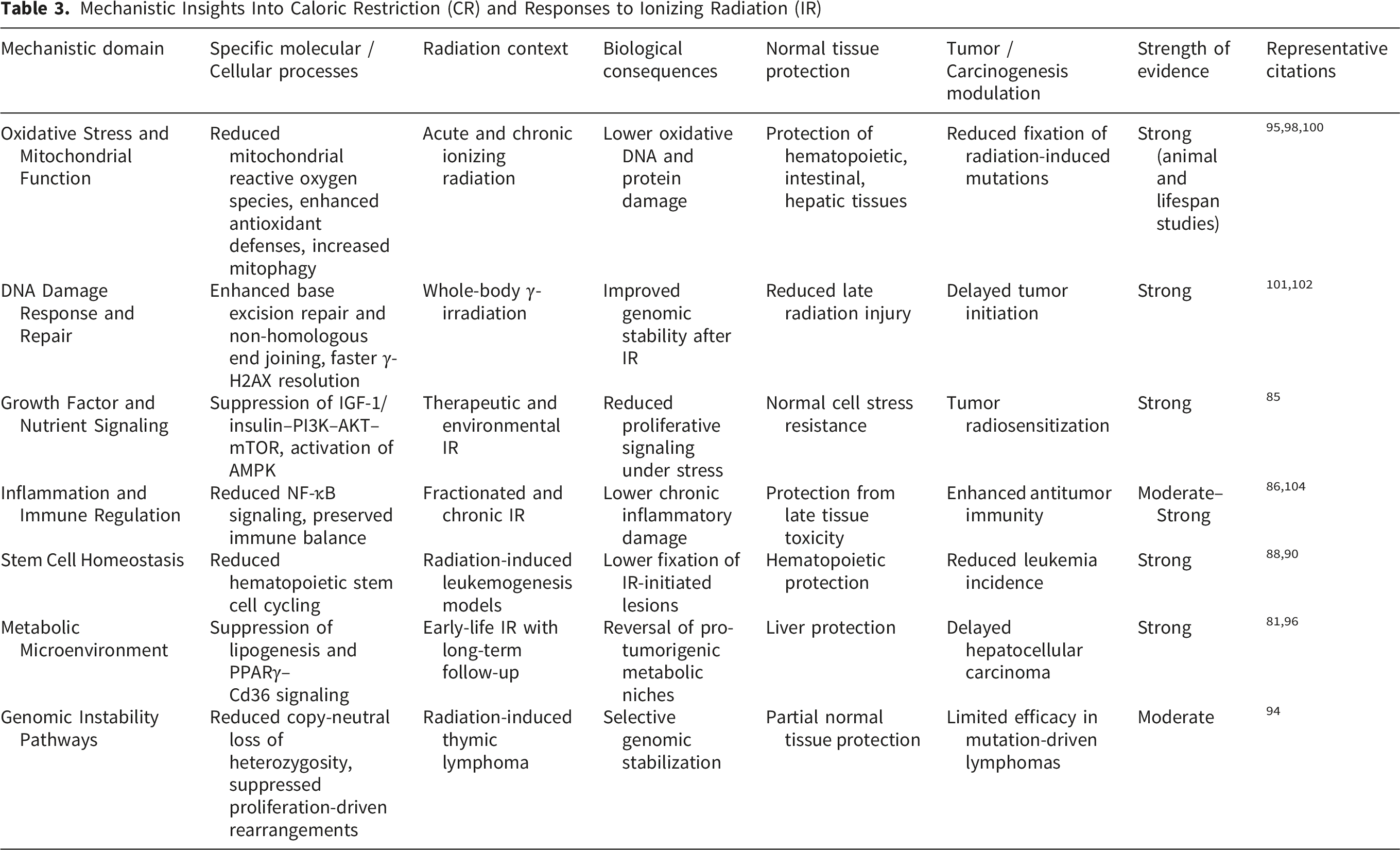
Mechanistic and genomic studies indicate that CR preferentially mitigates proliferation-associated genomic instability rather than mutation-driven oncogenesis. Radiation-associated lymphoid malignancies arise through diverse molecular routes involving Trp53, Ikzf1, Cdkn2a, and Pten alterations.92-94 CR selectively reduces copy-neutral loss of heterozygosity and oxidative mutation signatures, including G:C→T:A transversions linked to ROS damage, highlighting mechanistic specificity. 95 Metabolic studies further demonstrate that CR reverses radiation-induced hepatic lipid accumulation and delays hepatocellular carcinoma development via modulation of the PPARγ–Cd36 pathway. 96 Quantitative carcinogenesis modeling similarly shows that CR reduces age-specific mortality from solid tumors by increasing the number of rate-limiting steps in tumor progression rather than reversing radiation-initiated mutations. 97
Collectively, these findings establish CR as a biologically potent yet context-dependent modifier of radiation responses. By limiting proliferation-driven genomic instability, correcting metabolic dysregulation, and enhancing immune and tissue resilience, CR can mitigate many radiation-associated late solid tumors. However, its efficacy remains limited against early-onset, mutation-driven lymphoid malignancies, underscoring the need for mechanism-informed and context-specific translational strategies.
3.2.2. Mechanistic Synthesis (IR-Facing Mechanisms)
CR has been interrogated using a diverse and rigorously controlled set of experimental paradigms designed to define how reduced energy intake modulates cellular, tissue, and organismal responses to ionizing radiation (IR). In rodent models, CR is typically implemented as a 20–40% reduction in caloric intake without malnutrition, with macronutrient and micronutrient composition matched to ad libitum controls. This design enables isolation of energy restriction–specific effects while avoiding confounding nutritional deficiencies. Across studies, CR has been evaluated using integrated survival analyses, histopathology, molecular and biochemical assays, mutation-reporter systems, genomic profiling, metabolic phenotyping, and functional tissue assessments, providing a comprehensive framework to interrogate oxidative stress, DNA damage responses, inflammation, metabolism, stem-cell dynamics, and long-term carcinogenesis risk.
One of the most reproducible and mechanistically coherent achievements of CR research is the attenuation of IR-induced oxidative stress through improvements in mitochondrial function and redox homeostasis. CR consistently lowers basal metabolic flux and mitochondrial ROS production, enhances antioxidant defenses (including superoxide dismutase and catalase activity), stabilizes mitochondrial membrane potential, and promotes mitophagy, thereby limiting the accumulation of dysfunctional, ROS-generating mitochondria following irradiation.98,99,100 These biochemical adaptations translate into reduced oxidative damage to lipids, proteins, and nucleic acids and are supported by mutation-reporter studies demonstrating suppression of radiation-associated oxidative base substitutions, particularly G:C→T:A transversions linked to 8-oxoguanine formation. 95
CR further enhances genomic stability by improving DNA damage surveillance and repair following irradiation. Experimental studies in γ-irradiated rodents report reduced DNA double-strand break burden, accelerated resolution of γ-H2AX foci, and increased activity of DNA repair pathways, including non-homologous end joining and base excision repair.101,102 These effects are mechanistically linked to CR-induced metabolic and stress-response signaling, including AMPK activation, mTOR suppression, and increased NAD+ availability, which collectively support genome maintenance and cellular resilience under radiation-induced stress. 103 In parallel, CR suppresses chronic inflammation by attenuating NF-κB signaling and pro-inflammatory cytokine production, mitigating inflammatory cascades that contribute to late radiation toxicity and tumor promotion.104,105
Long-term carcinogenesis studies provide robust functional validation of these mechanisms while revealing important tissue-, pathway-, and age-dependent effects. In ApcMin/+ mice, CR instituted after irradiation, whether initiated during juvenile or adult stages, significantly suppressed the growth and progression of spontaneous and radiation-induced intestinal tumors. 82 In B6C3F1 mice exposed to IR during infancy, adult-onset CR extended lifespan by approximately 20% and markedly reduced the incidence of late-occurring tumors, including HCC, lung tumors, Harderian gland tumors, hemangiomas, and other non-thymic neoplasms, effectively reversing several radiation-associated increases in tumor burden. 81 In contrast, CR exhibited limited efficacy against early-onset thymic lymphoma (TL), highlighting the existence of CR-resistant carcinogenic pathways.
Genomic analyses have clarified the mechanistic basis for these divergent outcomes. Radiation- and chemically induced lymphoid malignancies arise through distinct oncogenic routes involving point mutations, structural rearrangements, and loss of tumor suppressor function. ENU-induced TLs frequently harbor point mutations in Trp53, Ikzf1, and Kras, often retaining the wild-type allele and suggesting dominant-negative or gain-of-function effects. 92 Radiation-induced TLs, by contrast, commonly exhibit Pten inactivation through diverse structural alterations, including missense and nonsense mutations, insertions, and focal deletions, leading to downstream Akt activation. 93 Lifelong CR preferentially suppresses proliferation-associated genomic instability, such as copy-neutral LOH, but does not prevent certain mutation-driven events; CR-group tumors instead show enrichment of dominant-negative Ikzf1 isoforms or point mutation–mediated inactivation. 94 These findings indicate that CR effectively limits cell-division–dependent genomic instability while allowing some mutation-centric oncogenic pathways to persist.
Metabolic and quantitative modeling studies further extend these insights. Early-life irradiation accelerates hepatic lipid accumulation and promotes a steatohepatitis-like microenvironment via activation of the PPARγ–Cd36 fatty acid uptake pathway, thereby facilitating HCC development; CR reverses these metabolic derangements, ameliorates hepatic inflammation, and delays tumor onset. 96 Multistage carcinogenesis modeling applied to lifespan tumor data demonstrates that CR reduces age-specific mortality from solid tumors and HCC regardless of irradiation status by increasing the number of rate-limiting carcinogenic steps, whereas irradiation accelerates transitions between steps. These findings indicate that CR primarily modifies host susceptibility and tumor promotion rather than directly counteracting radiation-initiated mutational events. 97
Collectively, this body of work establishes CR as a powerful, mechanistically grounded modulator of radiation responses. Through coordinated suppression of oxidative stress, enhancement of DNA repair and mitochondrial quality control, attenuation of inflammatory and metabolic tumor-promoting microenvironments, and limitation of proliferation-associated genomic instability, CR confers multi-level protection against IR. Importantly, these benefits extend to adulthood for many solid tumor types but remain limited for certain early-onset, mutation-driven lymphoid malignancies. Together, these achievements define CR as a biologically potent yet context-dependent strategy for modifying radiation sensitivity and carcinogenic risk, supporting its consideration within targeted, mechanism-informed approaches to radiation exposure and cancer prevention.81,82,92-97,100,101
3.2.3. Main Conclusions
Across preclinical and translational studies, CR consistently emerges as a potent modifier of radiation responses. In radiotherapy models, CR enhances tumor control by suppressing IGF-1R signaling, increasing apoptosis, and reducing metastatic burden, while simultaneously strengthening anti-tumor immunity through reduced Treg expansion and increased CD8+ T-cell activity.85,86 CR also amplifies immune responsiveness to low-dose radiation in a genotype-dependent manner. 87
CR profoundly reduces susceptibility to radiation-induced carcinogenesis, particularly hematopoietic malignancies. Studies demonstrate decreased incidence and delayed onset of radiation-associated leukemia, with protection linked to sustained suppression of stem-cell proliferation and reduced fixation of radiation-initiated lesions.79,88,89 Timing is critical, as pre-irradiation CR primarily affects initiation events, whereas post-irradiation CR more strongly modulates tumor promotion and microenvironmental processes. 90 Even mild restriction can enhance radioadaptive responses. 106
CR also confers broad protection against late solid tumors following irradiation by reducing oxidative DNA damage, correcting metabolic dysregulation, and modulating tissue microenvironments. These effects include suppression of ROS-associated mutation signatures, 95 reversal of radiation-induced hepatic steatosis, 96 and reduction of tumor mortality through delayed carcinogenic progression. 97 Nevertheless, CR is not universally effective. Early-onset lymphoid malignancies driven by mutation-centric mechanisms, including Trp53, Ikzf1, and Pten alterations, show relative resistance to CR-mediated protection.92-94 Thus, while CR substantially modifies long-term radiation risk through integrated metabolic, genomic, and immune mechanisms, its efficacy remains tissue- and pathway-dependent, underscoring the need for biomarker-guided and combinatorial strategies in radiation-exposed populations.
3.3. Other Restriction Approaches (ORA) and IR
3.3.1. Introduction
Radiotherapy (RT) is widely used in cancer management but is constrained by tumor radioresistance and dose-limiting toxicity to normal tissues, particularly in rapidly proliferating compartments such as the GI tract, oral mucosa, skin, and salivary glands. Interpatient heterogeneity in metabolic health, immune competence, microbiota composition, and baseline nutritional status further contributes to variable radiation responses and clinical outcomes.107,108
ORA—including fasting and fasting-mimicking diets (FMD), ketogenic/low-carbohydrate high-fat (LHF) regimens, intermittent fasting, and selective restriction/modulation of nutrients—have emerged as adjunct strategies intended to reshape radiation responses through systemic metabolic shifts and tumor–host asymmetries. Ketogenic/LHF diets induce ketosis and favor fatty acid oxidation, a state proposed to alter tumor energetic vulnerability while maintaining normal tissue function. 109 In parallel, fasting, intermittent fasting, and FMD activate stress-response programs that enhance cellular maintenance in normal tissues, whereas oncogene-driven tumors may fail to mount equivalent protective adaptations, resulting in increased DNA damage and radiosensitization. 110 Beyond whole-diet interventions, ORA also includes targeted nutrient modulation (e.g., amino acids, fatty acids, micronutrients) that can influence redox balance, nucleotide metabolism, mitochondrial dynamics, and immune interactions. Emerging evidence further implicates the tumor-associated microbiome and host metabolic context as actionable mediators of ORA effects on radiation response. 111
3.3.2. Main Achievements
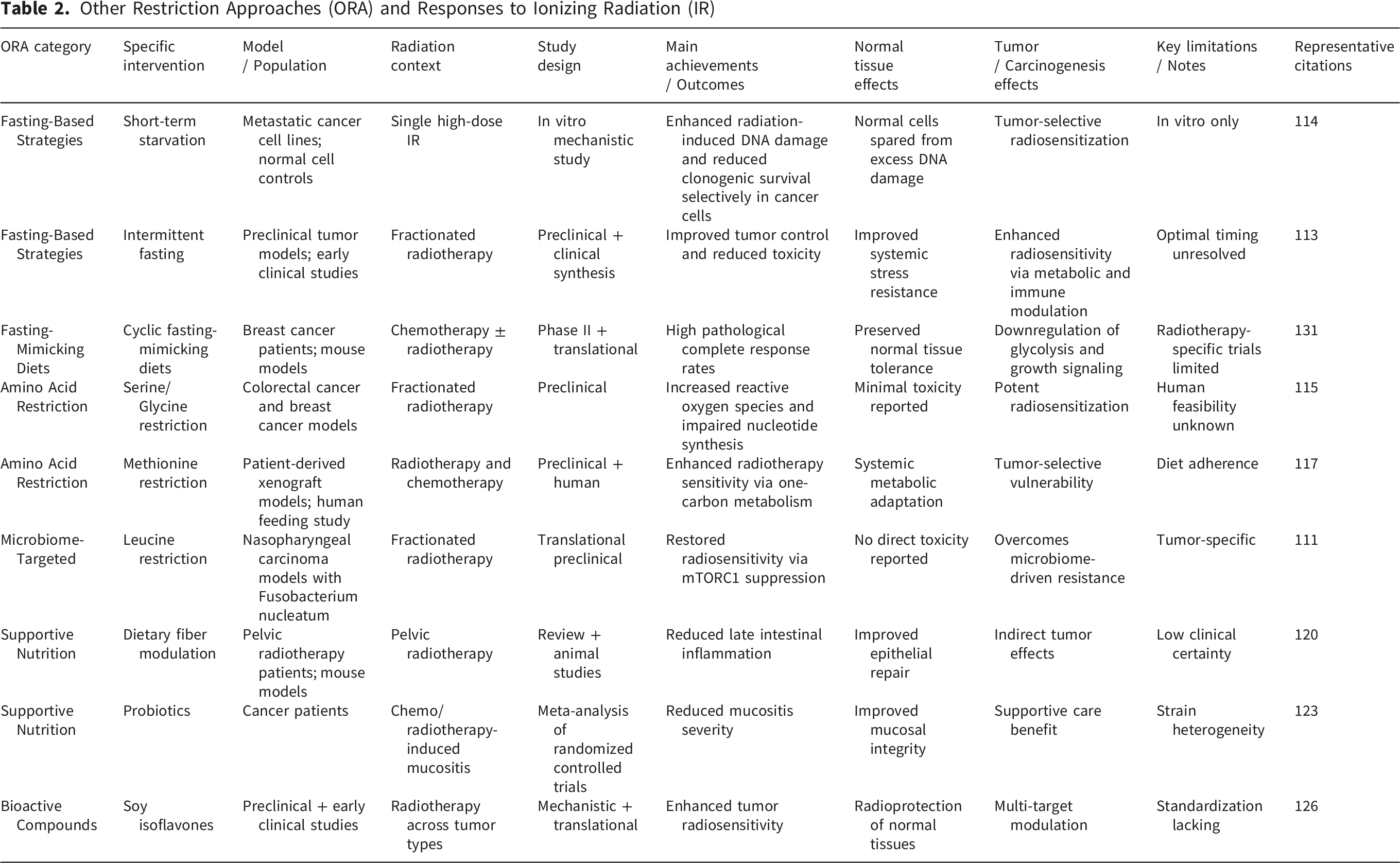
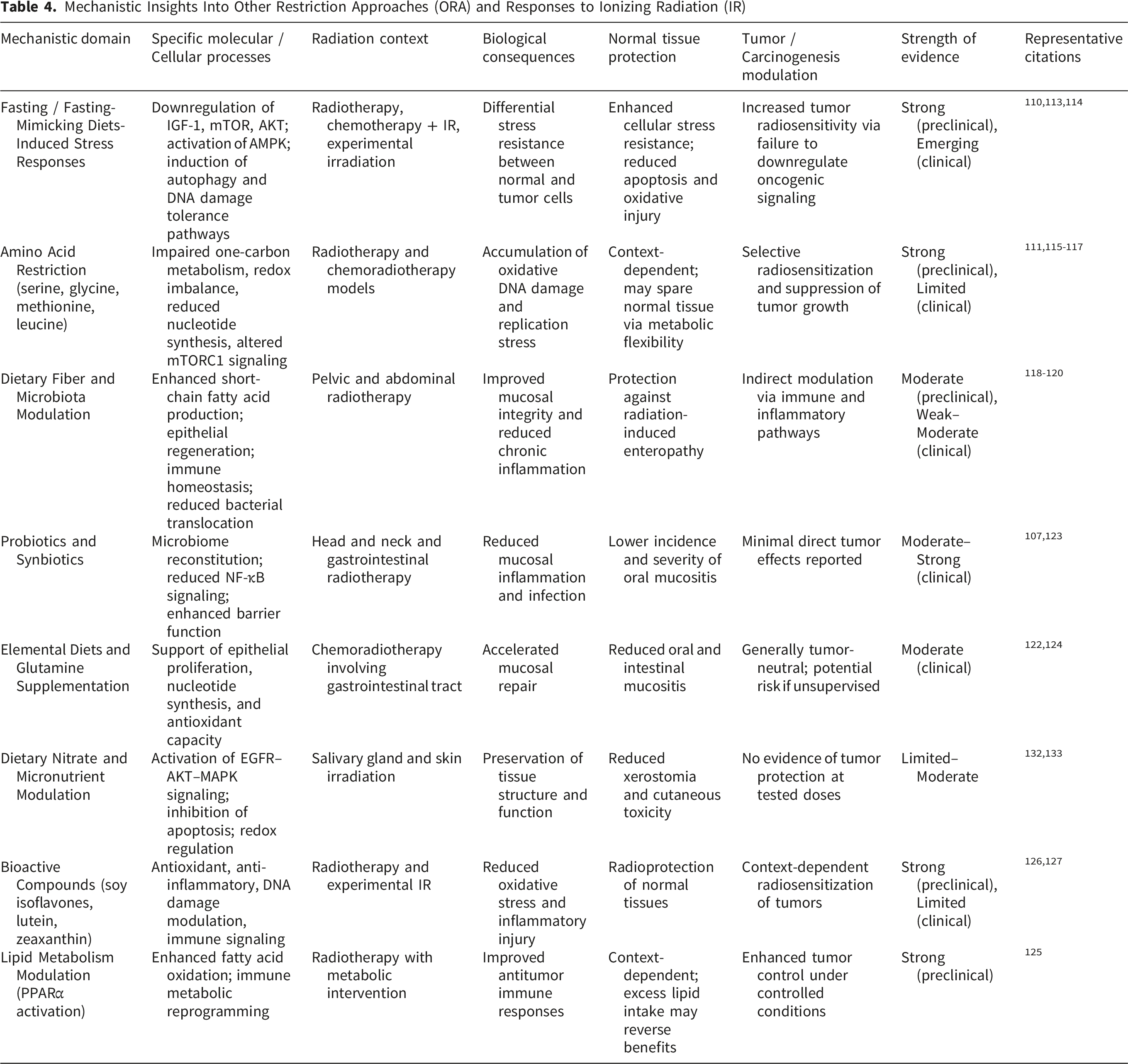
Research on ORA, encompassing fasting-based strategies, targeted nutrient restriction or supplementation, microbiome-modulating diets, and bioactive dietary compounds, has yielded several key advances in understanding how dietary modulation intersects with RT efficacy and toxicity. Collectively, these studies demonstrate that ORA can influence radiation responses at multiple biological levels, including tumor metabolism, normal tissue resilience, immune regulation, and host–microbiome interactions.
3.3.2.1. Feasibility, Safety, and Patient-Centered Outcomes
Across clinical and translational studies, ORA interventions have generally demonstrated acceptable feasibility and safety when implemented during RT or chemoradiotherapy. Systematic evaluations indicate high patient acceptability and completion rates for dietary interventions overall, although adherence varies substantially depending on intervention intensity and clinical context. 112 While survival benefits have not been consistently demonstrated, multiple studies report improvements in treatment tolerance, symptom burden, and quality-of-life parameters, supporting ORA as supportive adjuncts rather than replacements for standard oncologic care. Fasting and FMD protocols have emerged as particularly promising within this category. Reviews of early clinical studies indicate that short-term fasting or FMD cycles are feasible and generally well tolerated, with evidence of reduced treatment-related toxicity and preserved patient well-being.110,113 These findings establish a practical foundation for further clinical investigation of metabolically restrictive strategies during RT. Notably, while fasting-mimicking diets (FMD) are supported by consistent preclinical evidence, the available clinical data remain encouraging but heterogeneous. This apparent contrast reflects differences in evidence maturity rather than inconsistency: preclinical studies consistently demonstrate metabolic reprogramming and tumor-selective radiosensitization, whereas clinical studies are still limited by small cohort sizes, variable protocols, and mixed endpoints. Accordingly, current clinical findings should be interpreted as preliminary, highlighting translational potential while underscoring the need for well-controlled, radiotherapy-specific trials.
3.3.2.2. Tumor-Selective Radiosensitization Through Metabolic Restriction
A major achievement of ORA research is the demonstration of tumor-selective radiosensitization via metabolic stress. Short-term starvation has been shown to enhance radiation-induced DNA damage and clonogenic cell death in metastatic cancer cells while sparing non-malignant cells, providing direct experimental evidence for differential stress resistance. 114 This principle has been reinforced by fasting and FMD studies, which consistently show that nutrient deprivation suppresses growth signaling and antioxidant defenses in tumor cells that are unable to appropriately downregulate oncogenic pathways. Targeted amino acid restriction represents another important advance. Restriction of serine and glycine impairs nucleotide synthesis, redox homeostasis, and central carbon metabolism, thereby amplifying radiation-induced oxidative stress and DNA damage across multiple tumor models.115,116 Similarly, methionine restriction disrupts one-carbon metabolism, enhances sensitivity to RT and chemotherapy in resistant tumors, and induces systemic metabolic changes in humans that mirror preclinical findings. 117 These studies provide mechanistic proof that selective nutrient limitation can be leveraged to overcome tumor radioresistance.
3.3.2.3. Protection of Normal Tissues and Mitigation of Treatment-Related Toxicity
A second major achievement of ORA research lies in its contribution to normal tissue protection. Preclinical models demonstrate that dietary fiber accelerates intestinal crypt regeneration, preserves epithelial barrier integrity, and prevents bacterial translocation and chronic inflammation following irradiation. 118 Although clinical evidence for fiber modification during pelvic RT remains inconsistent and of low certainty,119,120 these mechanistic insights highlight the therapeutic relevance of diet–microbiota interactions in radiation injury. Strongest clinical support within ORA is observed for interventions targeting oral and mucosal toxicity. Meta-analyses show that zinc supplementation and amino acid–rich elemental diets significantly reduce the incidence and severity of oral mucositis, particularly in GI and esophageal cancers.121,122 Probiotic supplementation further reduces severe mucositis without increasing GI adverse events, supporting both efficacy and safety. 123 Glutamine supplementation consistently improves mucosal repair and symptom control across multiple anatomical sites. 124 Together, these findings establish ORA as a viable strategy for mitigating dose-limiting toxicities that compromise treatment continuity and quality of life.
3.3.2.4. Immune Modulation and Microbiome-Mediated Effects
ORA studies have substantially advanced understanding of immune and microbiome-mediated modulation of RT response. Metabolic dysfunction and nutrient deficiencies are increasingly recognized as contributors to immune suppression and radiation resistance, particularly in vulnerable patient populations, highlighting metabolic correction as a potential strategy to improve therapeutic index. 108 At the tumor level, intratumoral bacteria such as Fusobacterium nucleatum have been identified as drivers of radioresistance through suppression of oxidative stress and inhibition of radiation-induced cell death pathways. Notably, dietary leucine restriction reduced bacterial burden and restored radiosensitivity in vivo, providing a compelling example of diet-driven modulation of tumor–microbe–radiation interactions. 111 In parallel, intermittent fasting and other metabolically restrictive strategies have been shown to enhance antitumor immune responses while reducing systemic toxicity.109,113
3.3.2.5. Context-Dependent Effects of Dietary Bioactives and Lipid Metabolism
Calorie Restriction (CR) and Responses to Ionizing Radiation (IR)
3.3.3. Main Conclusions
Other Restriction Approaches (ORA) and Responses to Ionizing Radiation (IR)
On the normal-tissue side, evidence supports roles for fiber, probiotics, elemental diets, glutamine, dietary nitrate, and micronutrients in mitigating GI toxicity, mucositis, xerostomia, and skin injury, with zinc and probiotics showing the most consistent mucositis benefits to date.118-123,132,133 Emerging data also emphasize immune and microbiome regulation—including intratumoral bacterial signaling—as integral mediators of dietary effects on RT response.111,134,135
However, limitations include heterogeneous clinical evidence, small cohorts, protocol variability, and adherence challenges during intensive therapy, favoring individualized, supervised implementation over uniform prescriptions.112,130 In addition, unsupervised supplementation—particularly antioxidants—may risk attenuating therapeutic efficacy, reinforcing the need for evidence-based guidance. 136 Future progress will require adequately powered trials incorporating mechanistic biomarkers, stratification by metabolic/immune status, and clinically meaningful endpoints.110,128,131
3.4. Mechanistic Insights of Dietary Interventions (DI) and IR
DI, encompassing CRD and ORA, modulate biological responses to IR through interconnected effects on metabolism, redox homeostasis, DNA damage responses, immune regulation, and tissue microenvironments. A unifying mechanistic principle emerging from preclinical and translational studies is that DI amplify intrinsic metabolic and stress-response differences between tumor and normal cells, thereby enhancing the therapeutic index of RT while attenuating radiation-induced normal tissue injury.
3.4.1. Metabolic Reprogramming and Redox Regulation
A central mechanism by which both CRD and ORA influence radiation responses is metabolic reprogramming. CRD reduces systemic energy availability and suppresses insulin/IGF-1–PI3K–Akt–mTOR signaling, leading to decreased cellular proliferation, enhanced mitochondrial efficiency, and improved oxidative stress control. These changes lower basal mitochondrial ROS production, enhance antioxidant defenses, and promote mitophagy, collectively reducing radiation-induced oxidative damage in normal tissues.95,100 ORA strategies, including fasting, FMD, ketogenic diets (KD), and targeted macronutrient or amino acid restriction, further exploit tumor metabolic inflexibility. By limiting glucose and specific amino acids (e.g., serine, glycine, methionine, leucine), these interventions disrupt tumor redox balance, nucleotide synthesis, and mitochondrial function, thereby increasing oxidative stress and radiosensitivity in cancer cells.115-117 In contrast, normal cells enter a conserved stress-resistant state characterized by reduced anabolic signaling and enhanced cellular maintenance, conferring radioprotection. 110
3.4.2. DNA Damage Responses and Genomic Stability
DI influence radiation-induced DNA damage and repair at multiple levels. CRD enhances DNA damage surveillance and repair capacity, accelerating clearance of γ-H2AX foci and supporting non-homologous end joining and base excision repair pathways. These effects are linked to AMPK activation, mTOR suppression, increased NAD+ availability, and improved mitochondrial quality control, which collectively preserve genomic integrity under radiation-induced stress.101,102
At the genomic level, CRD preferentially suppresses proliferation-associated genomic instability, including copy-neutral LOH and replication-associated rearrangements, while exerting weaker effects on mutation-centric pathways driven by dominant-negative point mutations or complex structural alterations. This mechanistic distinction explains the strong protective effects of CRD against late-occurring solid tumors and myeloid malignancies, contrasted with its limited efficacy against early-onset, mutation-driven lymphoid tumors such as TL.94,97 ORA strategies similarly modulate DNA damage outcomes. Short-term starvation selectively increases radiation-induced DNA damage and clonogenic death in tumor cells while sparing normal cells, reflecting differential activation of cell-cycle checkpoints and stress-response pathways. 114 Amino acid restriction further impairs tumor DNA repair capacity by limiting substrates for nucleotide synthesis and one-carbon metabolism, thereby amplifying IR-induced cytotoxicity.
3.4.3. Immune Modulation and Inflammatory Control
Immune regulation represents a key interface between DI and radiation response. CRD suppresses chronic inflammation by inhibiting NF-κB signaling and pro-inflammatory cytokine production, mitigating late radiation injury and carcinogenesis. In RT settings, CRD remodels antitumor immunity by preventing radiation-induced expansion of regulatory T cells, increasing CD8+ T-cell activation, and improving CD8+: Treg ratios, effects associated with enhanced tumor control. 86
ORA interventions exert complementary immunomodulatory effects. Intermittent fasting, FMD, and KD enhance antitumor immune responses through systemic metabolic reprogramming, increased T-cell fitness, and reduced immunosuppressive signaling.109,113 Conversely, metabolic dysregulation in obesity, aging, or nutrient deficiency is associated with immune suppression and radiation resistance, highlighting the importance of metabolic context in shaping immune-mediated radiation outcomes.52,108
3.4.4. Tissue Microenvironment, Microbiome, and Normal Tissue Protection
Beyond tumor-intrinsic mechanisms, DI influence radiation responses through effects on tissue microenvironments and host–microbiome interactions. CRD ameliorates radiation-induced metabolic derangements such as hepatic steatosis and NASH-like inflammation by suppressing PPARγ–Cd36–mediated lipid uptake, thereby delaying HCC development after early-life irradiation. 96
ORA strategies targeting gut health, such as dietary fiber, probiotics, elemental diets, glutamine, and dietary nitrate, preserve mucosal integrity, reduce inflammation, and support epithelial regeneration following irradiation. These effects are mediated through modulation of microbial composition, barrier function, immune signaling, and apoptosis pathways, contributing to reduced GI toxicity, oral mucositis, xerostomia, and skin injury.118,123,132 Emerging evidence further implicates intratumoral bacteria in radioresistance. Fusobacterium nucleatum promotes tumor survival by suppressing oxidative stress and inhibiting PANoptosis via amino acid–dependent mTORC1 signaling; dietary leucine restriction restores radiosensitivity, underscoring diet–microbiome–tumor interactions as actionable modulators of radiation response. 111
3.4.5. Integrated Mechanistic Framework
Mechanistic Insights Into Caloric Restriction (CR) and Responses to Ionizing Radiation (IR)
Mechanistic Insights Into Other Restriction Approaches (ORA) and Responses to Ionizing Radiation (IR)
4. Integrative Summary and Emerging Paradigms
4.1. Key Findings and Shared Mechanisms
DI encompassing CRD and ORA, including fasting, FMD, ketogenic strategies, and targeted nutrient modulation, consistently emerge as powerful modifiers of biological responses to IR. Across preclinical, translational, and early clinical studies, these interventions influence both tumor and normal tissue responses, shaping radiation sensitivity, therapeutic efficacy, toxicity profiles, and long-term carcinogenic risk.
At the phenotypic level, a robust and reproducible observation is that DR improves tolerance to IR while, in many contexts, enhancing tumor control. In normal tissues, CRD and ORA attenuate acute and chronic radiation injury, particularly in metabolically active and rapidly renewing organs such as the GI tract, hematopoietic system, skin, and salivary glands. These protective effects manifest as reduced oxidative damage, preserved tissue architecture, improved regenerative capacity, and diminished inflammatory sequelae, translating into improved functional outcomes and quality of life in clinical settings. In parallel, multiple tumor models demonstrate that DR can sensitize cancer cells to radiation, suppress metastatic progression, and enhance RT efficacy, particularly in aggressive solid tumors.
Mechanistically, these divergent effects on tumor versus normal tissues are unified by the concept of differential stress resistance. CRD and ORA induce systemic metabolic reprogramming characterized by reduced circulating glucose, insulin, and IGF-1 signaling, activation of AMPK, suppression of mTOR, and shifts in mitochondrial substrate utilization. Normal cells respond to these changes by entering a protected, stress-resistant state marked by reduced proliferation, enhanced mitochondrial quality control, improved redox homeostasis, and more efficient DNA damage surveillance and repair. In contrast, cancer cells, constrained by oncogenic signaling, high biosynthetic demand, and limited metabolic flexibility, fail to fully engage these adaptive programs, resulting in heightened oxidative stress, impaired redox buffering, disrupted nucleotide and lipid metabolism, and increased susceptibility to radiation-induced DNA damage and cell death. A central mechanistic axis linking DR to radiation response is the modulation of oxidative stress and mitochondrial function. CRD and related interventions lower basal mitochondrial ROS production, enhance antioxidant defenses, and promote mitophagy in normal tissues, thereby limiting fixation of radiation-induced damage. In tumors, however, restriction of glucose, amino acids, or specific metabolic pathways (e.g., serine–glycine–one-carbon metabolism, methionine metabolism, fatty acid oxidation) compromises redox balance and biosynthetic capacity, amplifying radiation-induced oxidative and genotoxic stress. Mutation-spectrum analyses and reporter assays further demonstrate that DR selectively suppresses oxidative stress–linked mutations and proliferation-associated genomic instability, providing a mechanistic basis for reduced long-term carcinogenic risk.
DR also exerts profound effects on DNA damage responses and genome stability. CRD enhances DNA repair capacity and checkpoint control in normal tissues, facilitating resolution of double-strand breaks and limiting delayed genomic instability. At the organismal level, multistage carcinogenesis modeling indicates that CRD primarily acts by increasing the number of rate-limiting steps required for tumor development, thereby lowering age-specific cancer mortality independent of radiation exposure, whereas IR accelerates transitions between steps. This conceptual distinction explains why CRD robustly suppresses many late-occurring solid tumors while exhibiting limited efficacy against early-onset, mutation-driven lymphoid malignancies that rely less on proliferative or metabolic contexts.
Beyond cell-intrinsic effects, DR reshapes the immune and inflammatory landscape that critically determines radiation outcomes. CRD and ORA suppress chronic inflammation, reduce NF-κB signaling, and modulate cytokine profiles, mitigating late radiation injury. Simultaneously, these interventions can enhance antitumor immunity by increasing CD8+ T-cell activity, limiting regulatory T-cell expansion, and improving immune–metabolic fitness within the tumor microenvironment. Emerging evidence further implicates the gut and intratumoral microbiome as intermediaries of dietary effects on radiation response, with dietary fiber, fasting, and targeted nutrient restriction influencing microbial composition, immune signaling, and even tumor radioresistance mechanisms. Importantly, these benefits are context-dependent and mechanistically constrained. Tissue type, age at exposure, timing and intensity of restriction, genetic background, and dominant oncogenic pathways all influence outcomes. Early occurring lymphoid malignancies driven by point mutations or complex structural alterations show relative resistance to CRD-mediated protection, underscoring the need for complementary strategies targeting mutation-centric pathways. In clinical settings, adherence, tolerability, and the risk of malnutrition necessitate careful patient selection and medical supervision.
In conclusion, CRD and ORA define a coherent class of dietary interventions that modulate responses to IR through integrated effects on metabolism, redox biology, DNA repair, immune regulation, and tissue homeostasis. By simultaneously protecting normal tissues and, in many contexts, sensitizing tumors, these interventions have the potential to enhance the therapeutic index of RT and reduce long-term radiation-associated cancer risk. Future progress will depend on rigorously designed clinical trials, incorporation of mechanistic biomarkers, and stratification by metabolic, immune, and microbiome profiles to translate these biologically compelling phenomena into safe, personalized, and evidence-based clinical applications.
4.2. Integrative Themes and Emerging Paradigms
Across decades of investigation, DI, spanning classical CRD and more recent ORA, have revealed a set of convergent biological principles governing responses to IR. Early studies, grounded primarily in lifespan analysis, tumor incidence, and gross physiological outcomes, established the empirical observation that reduced nutrient availability profoundly alters radiation sensitivity, carcinogenesis, and tissue resilience. Contemporary work now provides mechanistic resolution to these phenomena, reframing dietary modulation as a systems-level regulator of radiation response rather than a simple modifier of energy balance.
A central unifying theme is differential stress adaptation between normal and malignant cells. Classical CRD studies demonstrated reduced radiation-induced tumor burden and delayed carcinogenesis, long before the molecular basis of these effects was understood. Modern data now show that CRD, fasting, ketogenic strategies, and targeted nutrient restriction converge on conserved nutrient-sensing pathways, AMPK, mTOR, insulin/IGF-1, and NAD+-dependent networks, that promote cellular maintenance, genomic stability, and metabolic efficiency in normal tissues, while simultaneously exposing the inflexibility of tumor metabolism. This framework reconciles early observations of tumor suppression and lifespan extension with current evidence for selective tumor radiosensitization and normal-tissue protection. A second integrative insight is the shift from mutation-centric to host-context–centric models of radiation carcinogenesis. Historically, radiation-induced cancer risk was viewed largely through the lens of DNA damage and fixed mutations. CRD studies challenged this paradigm by demonstrating profound reductions in radiation-associated cancer incidence without eliminating initial mutational events. Mechanistic analyses now clarify that DR primarily suppresses promotion and progression by limiting proliferative drive, oxidative stress, inflammatory signaling, and stem-cell cycling, processes required for clonal expansion of initiated cells. This host-level modulation explains why CRD is highly effective against late-occurring solid tumors but less so against early, mutation-driven lymphoid malignancies, and highlights the importance of tissue context and timing. Third, both old and new studies converge on metabolic regulation as a master determinant of radiation response. Early CR experiments implicitly altered mitochondrial function and redox balance; contemporary work explicitly demonstrates that dietary interventions remodel mitochondrial quality control, ROS handling, lipid and amino acid metabolism, and one-carbon flux. These metabolic shifts influence DNA repair capacity, epigenetic regulation, immune cell function, and microbial ecology, integrating tumor-intrinsic and host-mediated mechanisms. Importantly, newer ORA strategies, such as FMD, amino acid restriction, and ketogenic regimens, can reproduce many CRD-associated benefits with greater temporal precision and potential clinical feasibility. A fourth emerging paradigm is the integration of immunity and the microbiome into radiation–diet interactions. Earlier CRD studies noted changes in immune competence without mechanistic clarity. Recent evidence demonstrates that dietary modulation reshapes antitumor immunity, alters regulatory versus effector T-cell balance, and influences radiation-induced inflammation. In parallel, gut and intratumoral microbiota have emerged as diet-sensitive regulators of radioresistance, normal tissue injury, and immune tone. These insights extend classical concepts of “radioprotection” into a broader ecological framework involving host–microbe–immune crosstalk.
Collectively, these themes underscore a conceptual evolution: dietary interventions are no longer viewed as nonspecific modifiers of radiation damage, but as precision tools capable of reprogramming systemic and cellular states that determine radiation outcomes. By integrating historical phenomenology with modern molecular, metabolic, and immunological insights, the field now recognizes that effective modulation of IR responses depends on when, how, and in whom DR is applied. This integrative perspective provides a foundation for rational design of next-generation dietary strategies that complement RT, reduce long-term radiation risk, and advance personalized radiation medicine.
5. Perspectives and Future Directions
5.1. Outstanding Questions and Conceptual Gaps
Despite substantial progress over several decades, critical gaps remain in our understanding of how DI, including CRD and ORA, interact with IR across biological scales. One major unresolved issue concerns context specificity: the magnitude and direction of DI effects vary markedly by tissue type, developmental stage, radiation quality and dose, and underlying genetic background. While CR robustly suppresses many late-onset solid tumors, its limited efficacy against mutation-driven lymphoid malignancies underscores the need to delineate mechanism-dependent boundaries of protection, particularly where carcinogenesis is dominated by early, irreversible genomic lesions.
Another key gap lies in defining the temporal dimension of dietary modulation. Evidence indicates that the timing of restriction relative to radiation exposure, pre-exposure, peri-exposure, or post-exposure, critically influences outcomes by differentially affecting initiation, promotion, and progression phases of carcinogenesis. However, systematic frameworks integrating timing, duration, and intensity of restriction with radiation biology are lacking. Relatedly, the long-term consequences of intermittent versus sustained restriction, especially under chronic or fractionated radiation exposure, remain insufficiently characterized. At the mechanistic level, important questions persist regarding how DI reshape DNA damage processing, mutational spectra, and clonal selection following IR. While CR consistently suppresses proliferation-associated genomic instability and oxidative base damage, alternative mutation-driven pathways can persist or even be selected under restricted conditions. How dietary modulation influences clonal competition, stem-cell hierarchy, and tissue repopulation after radiation injury is only beginning to be understood. In parallel, the roles of the microbiome, intratumoral bacteria, immune-metabolic crosstalk, and cancer stem cell niches introduce additional layers of complexity that demand integrative, systems-level investigation.
Finally, translational uncertainty remains substantial. Most clinical data derive from small or early-phase studies, with heterogeneous protocols and limited biomarker integration. Identifying predictive markers of responsiveness, defining safe boundaries to avoid malnutrition or impaired treatment efficacy, and distinguishing beneficial metabolic stress from harmful catabolism are essential prerequisites for broader clinical adoption.
5.2. Translational Horizons Across Radiation-Exposed Settings
The expanding mechanistic foundation of dietary modulation of IR responses opens multiple translational avenues spanning medicine, occupational health, and extreme environments. In radiation oncology, DI hold promises as adjunct strategies to improve the therapeutic index of RT by simultaneously enhancing tumor radiosensitivity and protecting normal tissues. Carefully designed, time-limited interventions, such as short-term fasting, FMD, or metabolically targeted restriction, may offer clinically feasible alternatives to chronic CR, particularly when guided by metabolic and immune biomarkers.
In radioprotection, especially for occupational or accidental exposures, dietary modulation represents a non-pharmacological approach to reduce long-term cancer risk and late tissue injury. The ability of CR to suppress radiation-associated leukemias and solid tumors even when initiated after exposure suggests potential relevance for exposed populations where prophylactic interventions are no longer possible. Integrating dietary strategies with conventional countermeasures could enhance resilience without the toxicities associated with many radioprotective drugs.
The implications extend further to space biology, where astronauts experience chronic low-dose and high-linear energy transfer radiation in combination with metabolic stressors such as microgravity and circadian disruption. Dietary interventions that stabilize mitochondrial function, limit oxidative stress, and preserve immune competence may represent practical countermeasures for long-duration missions, provided their effects on muscle mass, bone density, and overall performance are carefully balanced.
From a public health perspective, the intersection of nutrition, metabolism, and radiation risk is particularly relevant for populations exposed early in life, individuals with metabolic syndrome or obesity, and aging societies with heightened vulnerability to both cancer and radiation injury. Tailored dietary guidance, informed by mechanistic insights rather than generalized restriction, could contribute to risk mitigation strategies following medical, environmental, or occupational radiation exposure.
Looking forward, progress will depend on integrative clinical trial designs, incorporation of mechanistic biomarkers (metabolic, genomic, immune, and microbiome-based), and cross-disciplinary collaboration among radiation biologists, nutrition scientists, oncologists, and epidemiologists. Rather than a universal prescription, dietary interventions should be viewed as precision modifiers of radiation response, whose optimal application will vary by context, mechanism, and individual susceptibility.
5.3. Personalized Dietary Interventions: Toward Precision Translation
DI, including CR, fasting regimens, time-restricted feeding, LHF, and nutrient-directed strategies such as dietary inversion, which strategically supplements or increases intake of selected amino acids, vitamins, minerals, and antioxidant-rich foods, are increasingly recognized as adaptable adjuncts to cancer therapy because of their capacity to reprogram systemic and cellular metabolism. Conventional modalities, including chemotherapy, RT, and immunotherapy, remain limited by variable efficacy and substantial normal-tissue toxicity. RT, in particular, frequently compromises nutritional status, especially in GI and head-and-neck malignancies, where anorexia, mucosal injury, and GI dysfunction contribute to malnutrition, treatment interruptions, and reduced tolerance. 137 Within this clinical landscape, appropriately designed DI offer an integrated strategy to enhance therapeutic effectiveness while supporting patient resilience across radiation-exposed settings.
Mechanistically, CR, intermittent fasting, and pharmacological CR mimetics enhance tumor sensitivity to RT and chemotherapy by suppressing glycolytic dependence, improving insulin sensitivity, inducing autophagy, and attenuating inflammatory and angiogenic signaling. In parallel, nutrient-enriching approaches, such as dietary inversion, may mitigate radiation-induced normal-tissue injury, restore metabolic homeostasis, and facilitate recovery following treatment. 138 Together, these complementary strategies illustrate how DI can simultaneously preserve normal-tissue integrity and exploit tumor-specific metabolic vulnerabilities, thereby improving the therapeutic index of radiation-based treatments.
The biological rationale for personalized DI is reinforced by the observation that aggressive and treatment-resistant tumors often display heightened glycolysis and metabolic inflexibility, features that overlap substantially with pathways targeted by CR and carbohydrate restriction. Systemic metabolic dysfunction, such as obesity, insulin resistance, and diabetes, is frequently associated with poorer oncologic outcomes and radioresistance, underscoring the importance of host metabolic state in shaping radiation response. Preclinical and early clinical studies suggest that integrating CR or CR mimetics with anticancer therapies can suppress tumor growth, enhance cancer-cell death, and potentially permit dose de-escalation of cytotoxic treatments, thereby reducing toxicity. Notably, cyclic FMD have demonstrated clinical feasibility, and early reductions in intratumoral glycolysis have emerged as candidate biomarkers predicting response to nutrient-based interventions.
Despite these advances, successful translation of DI requires careful personalization. Patient-specific factors, including age, baseline nutritional status, comorbidities, treatment intensity, and radiation field, critically influence safety, adherence, and efficacy. Overly restrictive or poorly timed interventions risk exacerbating frailty, impairing immune competence, or undermining treatment tolerance. Moreover, heterogeneity in dietary protocols, tumor biology, metabolic phenotypes, and combination treatment strategies contributes to variability in clinical outcomes, limiting the generalizability of existing evidence.
5.4. Clinical Caution and Malnutrition Risk Management
Implementation of restrictive dietary interventions during radiotherapy demands strict nutritional surveillance and multidisciplinary supervision, particularly in patients with head-and-neck or gastrointestinal malignancies who are highly vulnerable to treatment-associated malnutrition. Recent American Society for Parenteral and Enteral Nutrition (ASPEN) clinical recommendations emphasize that all oncology patients must undergo regular malnutrition risk screening before and throughout radiation treatment using validated tools such as the Malnutrition Screening Tool (MST), Malnutrition Universal Screening Tool (MUST), Nutritional Risk Screening 2002 (NRS-2002), or Patient-Generated Subjective Global Assessment (PG-SGA).
139
Particular caution is warranted in patients receiving concurrent chemoradiotherapy, where treatment-related mucositis, dysphagia, and gastrointestinal toxicity can rapidly accelerate metabolic deterioration.
140
To ensure patient safety and prevent severe cachexia during radiotherapy, clinicians should establish clear thresholds for modifying or immediately discontinuing restrictive regimens (such as Fasting-Mimicking Diets). Based on consensus clinical guidelines for oncology nutrition, it is recommended to discontinue restrictive DI if any of the following parameters are met: • Unintentional Weight Loss: A cumulative weight loss exceeding 5% within 1 month, or more than 10% over 6 months.
141
• Body Mass Index (BMI): A drop below 18.5 kg/m2 for adults under 65 years, or below 20.0 kg/m2 for patients aged 65 and older, signaling severe thinness.
141
• Oral Intake: A progressive decline in daily oral food intake where the patient consumes less than 60–70% of their estimated energy requirements for more than 1–2 weeks.
142
• Clinical Presentation: Manifestation of severe sarcopenia, physical frailty, or severe (Grade 3+) radiation-induced toxicities (e.g., severe mucositis or enteritis) that limit baseline hydration and recovery.
143
Incorporating these practical thresholds into clinical oncology workflows allows for individualized surveillance, ensuring that the metabolic stress induced by restrictive DI remains within a safe therapeutic window without driving the patient into harmful catabolism.
Accordingly, future research must prioritize precision nutrition frameworks that tailor dietary strategies to individual metabolic, immunologic, and tumor-specific characteristics. Well-designed clinical trials that incorporate mechanistic biomarkers, spanning metabolic flux, immune profiling, microbiome composition, and circadian regulation, are essential for defining optimal nutrient composition, timing, duration, and safety parameters. Aligning DI with patient-specific vulnerabilities and therapeutic contexts will be critical for realizing their translational potential across oncology, radioprotection, and other radiation-exposed environments.
6. Conclusion
In conclusion, DR—encompassing caloric, amino acid, and fasting-based regimens—has transitioned from a classical longevity intervention into a powerful systems-level metabolic strategy capable of modulating biological responses to ionizing radiation (IR). Governed by the principle of differential stress adaptation, DR selectively reprograms normal cells away from anabolic growth toward cellular maintenance, reinforcing redox homeostasis, DNA damage repair, and immune-inflammatory control to mitigate radiation-induced toxicities. Conversely, metabolically rigid tumor cells, constrained by oncogenic signaling, fail to mount equivalent adaptations under nutrient deprivation, rendering them more vulnerable to radiation-induced oxidative and genotoxic stress. Preclinical and emerging clinical findings, including innovative nutrient-directed strategies like dietary inversion, strongly support the potential of DIs to simultaneously protect healthy tissues and enhance the therapeutic index of radiotherapy.
However, translating these biological phenomena into clinical oncology and radioprotective frameworks remains heavily context-dependent and presents substantial translational challenges. The efficacy of DR is strictly constrained by tissue specificity, the timing and duration of the protocol relative to IR exposure, radiation quality, and individual tumor genotypes. Furthermore, unsupervised or overly restrictive regimens risk compromising the nutritional status and therapeutic tolerance of vulnerable patients, reinforcing the absolute necessity for rigorous clinical monitoring. Future progress must therefore prioritize well-designed, radiotherapy-specific clinical trials that incorporate mechanistic biomarkers. Shifting the paradigm from uniform dietary restrictions toward personalized, biomarker-guided precision nutrition will be essential to safely and effectively translate these interdisciplinary insights into actionable strategies for precision oncology and targeted public health countermeasures.
Footnotes
Acknowledgments
The authors gratefully acknowledge Ms. Hiromi Arai for her exceptional technical assistance and administrative support.
Author Contributions
Conceptualization, J.K., Y.S. and B.W.; methodology, J.K., Y.S., T.K., D.N. and B.W.; data curation, Y.S., B.W. and J.K.; writing—original draft preparation, Y.S., B.W. and J.K.; writing—review and editing, Y.S., T.K., D.N., T.M., S.K., B.W. and J.K.; visualization, B.W. and J.K.; supervision, J.K.; funding acquisition, Y.S., T.K., T.M., S.K. and B.W. All authors have read and approved the final version of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI: Grant-in-Aid for Scientific Research (A) (Grant Numbers JP21H04932 and JP24H00764) and the Faculty of Associated Medical Sciences, Chiang Mai University, Thailand: Grant-in-Aid for Scientific Research (C) (Grant Numbers JP19K12335, JP22K12387, JP23K11431, JP24K15298 and JP25K15450).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data discussed in this review were obtained from previously published studies; no new data were generated.