
Abstract
Inflammatory pain is mediated by complex interactions between immune signaling and sensory neurons, with prostaglandin E2-dependent pathways playing a central role in nociceptive sensitization. Metformin, a first-line antidiabetic drug, has emerged as a potential non-opioid analgesic due to its pleiotropic anti-inflammatory and neuroprotective properties. In this study, we investigated the analgesic effects of metformin using both an in vitro neuronal sensitization model and an in vivo model of inflammatory pain. In SH-SY5Y neuronal cells, PGE2 exposure significantly reduced Na+/K+-ATPase activity and increased intracellular cAMP levels. Post-treatment with metformin restored Na+/K+-ATPase activity and suppressed cAMP elevation in a concentration-dependent manner. In vivo, metformin produced dose-dependent analgesia in mice with complete Freund’s adjuvant-induced inflammatory pain, with 0.1 g/kg/day significantly increasing hind paw withdrawal thresholds. Molecular analyses revealed significant downregulation of pain-associated proteins TRPV1 and NaV1.7 in metformin-treated groups, supported by both Western blot and immunohistochemical findings. Histological examination further demonstrated reduced inflammatory cell infiltration and improved tissue recovery. Collectively, these findings indicate that metformin alleviates inflammatory pain by modulating neuronal excitability and inflammatory signaling pathways, highlighting its potential as a non-opioid therapeutic candidate for inflammatory pain management.
Introduction
Inflammatory pain is a common clinical condition arising from tissue injury, infection, or immune dysregulation, and is characterized by peripheral and central sensitization mediated by pro-inflammatory cytokines, chemokines, and oxidative stress.1,2 Despite the availability of nonsteroidal anti-inflammatory drugs, corticosteroids, and opioids, current therapies often provide incomplete relief and may lead to significant adverse effects.3–5 Therefore, investigating the potential of drugs originally developed for other indications, yet possessing anti-inflammatory, antioxidative or neuroprotective properties, may open new therapeutic avenues for inflammatory pain management.
Metformin, a first-line oral hypoglycemic agent for type 2 diabetes, has recently gained attention as a potential analgesic due to its pleiotropic, AMPK-mediated effects beyond glucose regulation. Activation of AMPK by metformin suppresses pro-inflammatory cytokines and modulates neuronal excitability, producing antinociceptive and anti-inflammatory actions. 6 In models of diabetic neuropathy, chronic constriction, and chemotherapy-induced pain, metformin restored p-AMPK activity, inhibited NF-κB and STAT3 signaling, reduced glial activation, and lowered cytokine levels via endogenous opioidergic pathways.7–9 Beyond neuropathic pain, metformin has shown efficacy in inflammatory pain models, where early administration reduced pain behaviors and tissue edema. 10 Acting through regulation of synaptic plasticity, inflammatory signaling, and neuronal survival, metformin represents a potential non-opioid therapeutic approach for chronic pain with limited treatment efficacy. 11
Inflammatory pain is mediated by complex interactions between immune signaling molecules and peripheral sensory neurons, with prostaglandin E2 (PGE2) playing a pivotal role. PGE2 activates EP2 and EP4 receptors on nociceptors, leading to increased intracellular cAMP levels and propagation of pro inflammatory signaling pathways that enhance neuronal excitability and pain perception. 12 To model this phenomenon in vitro, the SH-SY5Y human neuroblastoma cell line was selected due to its expression of neuronal markers and pain related receptors. SH-SY5Y cells have been validated in studies investigating neuroinflammation and nociceptive signaling, including experiments utilizing PGE2 to induce cAMP dependent stress responses. 13 In this study, PGE2 served as a pro inflammatory stimulus to induce neuronal sensitization. PGE2 exposure disrupts neuronal homeostasis by increasing intracellular cAMP levels and impairing membrane-associated enzyme activities, including Na+/K+-ATPase, thereby establishing a controlled in vitro model for evaluating neuroprotective and analgesic interventions. Accumulating evidence indicates that reduced Na+/K+-ATPase activity contributes to nociceptive sensitization, whereas preservation or restoration of its function is associated with analgesic effects. 14
Metformin was investigated as the primary therapeutic agent. Prednisone was included as a positive control due to its glucocorticoid mediated suppression of cyclooxygenase 2 (COX-2) expression and PGE2 synthesis,15,16 serving as a benchmark for evaluating the anti-inflammatory and analgesic effects of metformin.
For in vivo modeling, complete Freund’s adjuvant (CFA) was injected into the plantar region of the hind paw to induce localized inflammation and pain, as previously described. 17 This approach is widely applied to mimic peripheral inflammatory conditions and to assess the efficacy of anti-inflammatory and analgesic agents.
The research consists of two phases: (1) an in vitro study using SH-SY5Y cells stimulated with PGE2 to assess modulation of pain-related pathways and (2) an in vivo study using CFA-induced inflammatory pain in mice. Behavioral assessment by the Von Frey test and molecular analysis of TRPV1 and NaV1.7 expression were conducted to elucidate the mechanisms underlying the analgesic and anti-inflammatory effects of metformin.18–21
Materials and methods
In vitro experimental design
Compounds
PGE2 was used as an in vitro inducer of inflammation and pain. Metformin served as the primary compound under investigation. Prednisone, a glucocorticoid, was included as a positive control. All compounds were purchased from Sigma-Aldrich (USA). PGE2 100 mg/ml and prednisone 100 mg/ml stock solutions were dissolved in ethanol, whereas metformin 300 mg/ml was dissolved in PBS. The final concentration of ethanol did not exceed 0.1% (v/v), and equivalent vehicle controls were included.
Cell line
The SH-SY5Y human neuroblastoma cell line (ATCC® CRL-2266™, Manassas, VA, USA) was obtained from the American Type Culture Collection. The cells were maintained in a 1:1 mixture of Ham’s F-12 and DMEM, supplemented with 10% fetal bovine serum, 0.25% HEPES, 0.25% sodium bicarbonate, and 1% penicillin–streptomycin (Gibco™, Waltham, MA, USA). To ensure consistency across all subsequent experiments, cells were maintained at a uniform density of 5 × 104 cells/cm2 under standard culture conditions 37°C in a humidified environment containing 5% CO2.
Determination of IC50 of Metformin for SH-SY5Y cells
The CCK-8 assay was used to determine the IC50 of metformin and to identify non-cytotoxic concentrations for subsequent experiments.
Previous studies report that the IC50 of metformin in various cell types typically ranges from 20 to 30 mM and remains below 100 mM.22,23 To determine IC50, seed cells at six concentrations prepared by 2-fold serial dilution starting from density of 5 × 104 cells/cm2 in a 96-well plate, with each concentration plated in triplicate and including three blank wells containing only culture medium. After 24 h of incubation under standard conditions, perform the CCK-8 assay to obtain OD450 readings and construct a standard curve relating OD450 to viable cell numbers. For metformin treatment, expose cells to 0, 0.1, 1, 3, 10, 30, and 100 mM for 48 h using the same triplicate setup and blank wells. Use the standard curve to calculate % inhibited cells and determine IC50 using GraphPad Prism 10. Three metformin concentrations concentrations <1/2 IC50 will be selected for the analgesic experiment, prednisone 1.5 µM was selected as the positive control.15,16
Evaluation of the pain-promoting effects of PGE2 in vitro
Based on previous studies, SH-SY5Y cells were treated with PGE2 at concentrations of 0, 1.25, 2.5, 5, 10, and 15 µM for 30 min. 13 2 × 106 cells were then collected to assess Na+/K+-ATPase activity (Creative Biomart, USA). The PGE2 concentration that caused a significant reduction in Na+/K+-ATPase activity was selected for establishing the in vitro pain model.
Determine the analgesic effect of metformin on Na+/K+-ATPase activity and intracellular cAMP concentration in a medium containing PGE2
SH-SY5Y cells were seeded at a density of 5 × 104 cells/cm2 in 10 cm culture dishes and incubated for 24 h. To induce a nociceptive-like response, cells were exposed to PGE2 at the predetermined concentration for 30 min. Following PGE2 stimulation, the medium was replaced with fresh medium containing metformin or prednisone, and cells were further incubated for 24 h. This treatment duration was selected to allow sufficient modulation of intracellular signaling pathways and the design mimics a therapeutic rather than preventive intervention. At the end of the treatment period, cells were collected for downstream analyses, including Na+/K+-ATPase activity and intracellular cAMP quantification (GenScript, USA).
All experiments were independently repeated at least three times.
In vivo experimental design
Animals
A pilot study using 24 C57BL/6 mice (LESCO Biotechnology, Taipei, Taiwan) was conducted to estimate sample size. Based on Von Frey test data, the main study required 40 mice (including 4 backups), as determined using G*Power 3.1.9.7 (Heinrich-Heine-Universität Düsseldorf, Germany). All procedures were approved by the Institutional Animal Care and Use Committee of Fu Jen Catholic University (IACUC No. FJU-A11232, 2023/09/30) and complied with ICLAS Ethical Guidelines. Prior to the experiments, mice were housed under standard conditions and acclimated for 2 weeks.
Model of inflammatory pain
Inflammatory pain was induced in C57BL/6 mice by plantar injection of CFA (Sigma-Aldrich, USA) into the hind paw. Based on previous studies, a reference volume of 10 µl CFA was selected, and two additional volumes (5 and 15 µl) were included to evaluate dose-dependent effects and determine the optimal condition for the model. Mice were randomly assigned to experimental groups receiving 5, 10, or 15 µl CFA, while control mice did not receive any injection (naive control group). Each group consisted of n = 6 mice, and results were expressed as mean ± SEM. Dose-dependent effects of CFA were evaluated based on behavioral observations and hind paw withdrawal thresholds. The volume that produced the most robust and consistent inflammatory pain phenotype was selected as the optimal condition for subsequent experiments. Following CFA administration, mice were divided into four experimental groups and treated via intraperitoneal injection as follows: C (control group): Saline, M1: metformin 0.05 g/kg/day, M2: metformin 0.1 g/kg/day, P: Prednisone 0.015 g/kg/day. The doses of metformin and prednisone used were about ≤1/10 of the LD50 values provided by the manufacturer (metformin LD50: 1 g/kg, prednisone LD50 mouse: 135 mg/kg). To determine the duration in analgesic effect of metformin, the Von Frey filament test was performed at multiple time points following a single compounds administration (2, 4, 6, and 24 h). Mechanical sensitivity of C57BL/6 mice was evaluated using the manual Von Frey test. Mice were placed in small, dark acrylic chambers with a wire-mesh floor, sized to limit excessive movement, and habituated for 15–30 min. Calibrated Von Frey filaments (0.008–4 g) were applied perpendicularly to the plantar surface of the hind paw until the filament slightly bent about 2–3 s). A positive response was defined as brisk paw withdrawal, licking, or shaking. The 50% hindpaw withdrawal threshold was determined using the up–down method. 24 Testing was performed by a blinded experimenter. Based on the results, the optimal time point for assessing analgesic activity was selected. A 21-day treatment followed, with Von Frey measurements on days 1, 4, 7, 14, and 21. Hind paw tissues were collected at study end: fixed in 4% formaldehyde for IHC and H&E staining, or frozen at −80°C for Western blot analysis.
Animal anesthesia and euthanasia: No anesthesia was used during the experimental procedures, as no invasive interventions were performed on the animals. At the end of the experiment, mice were euthanized using carbon dioxide (CO2) inhalation in accordance with the AVMA Guidelines for the Euthanasia of Animals. 25 The procedure involved administering 100% CO2 into a chamber at a controlled flow rate until the animals lost consciousness and respiration ceased, ensuring a humane and ethical endpoint.
Western Blot analysis
Total protein was extracted from tissue using a solution from iNtRON Biotechnology (Serdang, Malaysia), and concentrations were determined by BCA assay. 30 μg of total protein were separated on 12% SDS-PAGE and transferred to PVDF membranes (Merck Millipore, USA). After blocking with 5% non-fat milk for 1 h, membranes were incubated overnight at 4°C with mouse anti-: phospho-TRPV1 GTX54762 (1:200), NaV1.7 GTX54800 (1:200), and β-actin GTX109539 (1:10,000) (GeneTex, USA). Following five PBST washes, membranes were incubated 1 h at room temperature with HRP-conjugated goat anti-mouse IgG (1:10,000) (Thermo Fisher, USA). Experiments were performed in triplicate. Signals were detected with UVP ChemiDoc-It (UVP, USA) and analyzed using ImageJ (LOCI, University of Wisconsin).
Immunohistochemical analysis
Formalin-fixed, paraffin-embedded tissue blocks were sectioned at 4 μm for immunohistochemistry using the Leica BOND-III automated system (Nussloch, Germany). Sections were mounted on silanized slides, dried overnight at 37°C, deparaffinized, and rehydrated. Endogenous peroxidase was blocked with 3% hydrogen peroxide for 5 min. After antigen retrieval (30 min, pH6), slides were incubated with TRPV1 primary antibody (1:100, GeneTex, Inc., USA) for 20 min, washed, and incubated with secondary antibodies (Leica Biosystems, Nussloch, Germany). Immunoreactivity was visualized with DAB, followed by hematoxylin counterstaining, dehydration, and coverslipping.
Histopathological analysis
H&E staining was performed to assess and compare inflammation across groups. Tissue specimens were paraffin-embedded, sectioned at 3–5 μm, and stained with Hematoxylin and Eosin. Whole-slide scanning at 200× (3DHISTECH, Budapest, Hungary) and computer-assisted quantification were used to ensure accurate evaluation. Tissue recovery was assessed according to the criteria in Table 1.
Grading criteria for assessing tissue recovery following inflammation.
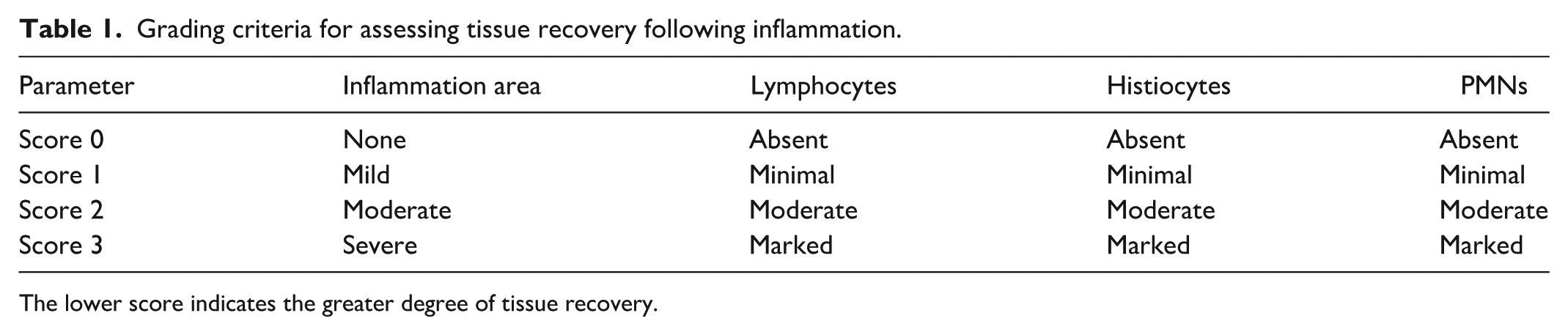
The lower score indicates the greater degree of tissue recovery.
Data analysis
Data were analyzed using SPSS version 22 (IBM Corp., USA) and are presented as mean ± SEM (n = number of replicates). Differences among groups were assessed using one-way ANOVA followed by Scheffé’s post hoc test. A value of p < 0.05 was considered statistically significant.
The IC50 value of metformin in the SH-SY5Y cell toxicity assay was calculated using a four-parameter nonlinear regression model in GraphPad Prism version 10.
Results
Establishment of a PGE2-induced in vitro pain model
Determination of metformin IC50 in SH-SY5Y cells
Each drug has a beneficial concentration range, while higher doses may cause toxicity or cell death. The IC50 of metformin was determined using CCK-8 assays and calculated with GraphPad Prism 10 (Figure 1). Three concentrations ≤1/2 IC50 were selected for subsequent experiments including 10, 8, 6 mM.
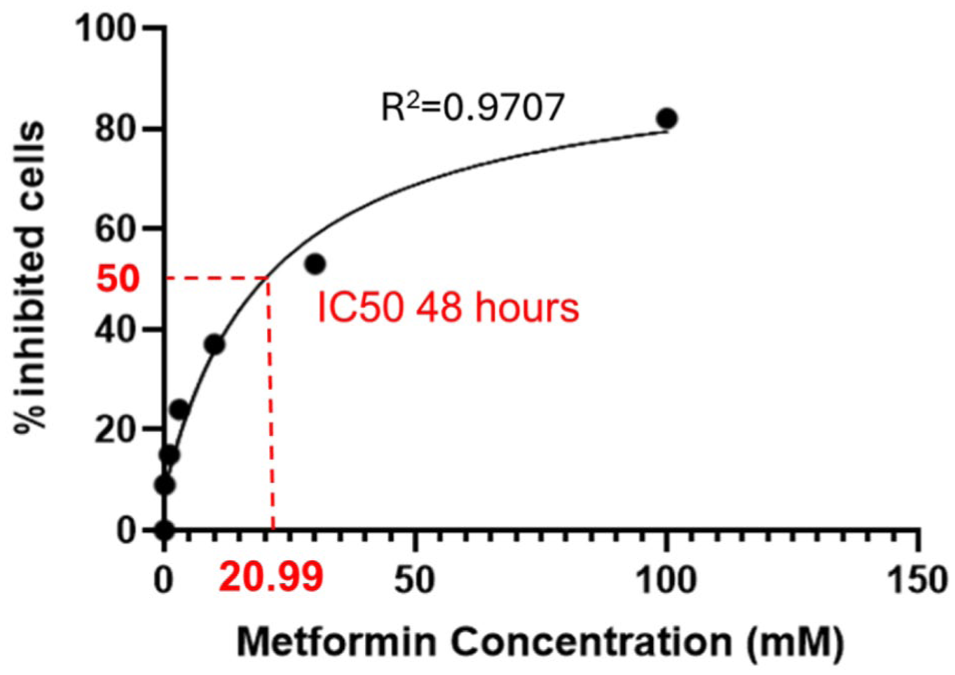
Determination of metformin IC50 in SH-SY5Y cells.
SH-SY5Y cells were treated with increasing concentrations of metformin for 48 h, and cell viability was assessed to determine cytotoxicity. The IC50 value of metformin was calculated as 20.99 mM, which was used to guide the selection of non-toxic concentrations for subsequent in vitro experiments.
PGE2 induces pain-like cellular responses in SH-SY5Y cells
Na+/K+-ATPase activity was measured in SH-SY5Y cells following 30 min exposure to increasing concentrations of PGE2 (1.25–15 µM). Data presented in Figure 2 indicate that PGE2 treatment resulted in a significant, concentration-dependent reduction in enzyme activity, decreasing to 0.86 ± 0.005 and 0.66 ± 0.0032 fold of the untreated control at 10 and 15 µM, respectively, compared with the untreated control (−). These findings demonstrate the functional impact of PGE2 on cellular ion regulation and support its role in inflammatory pain signaling. Based on the observed dose–response pattern, 15 µM PGE2 was selected for establishing the in vitro inflammatory pain model.
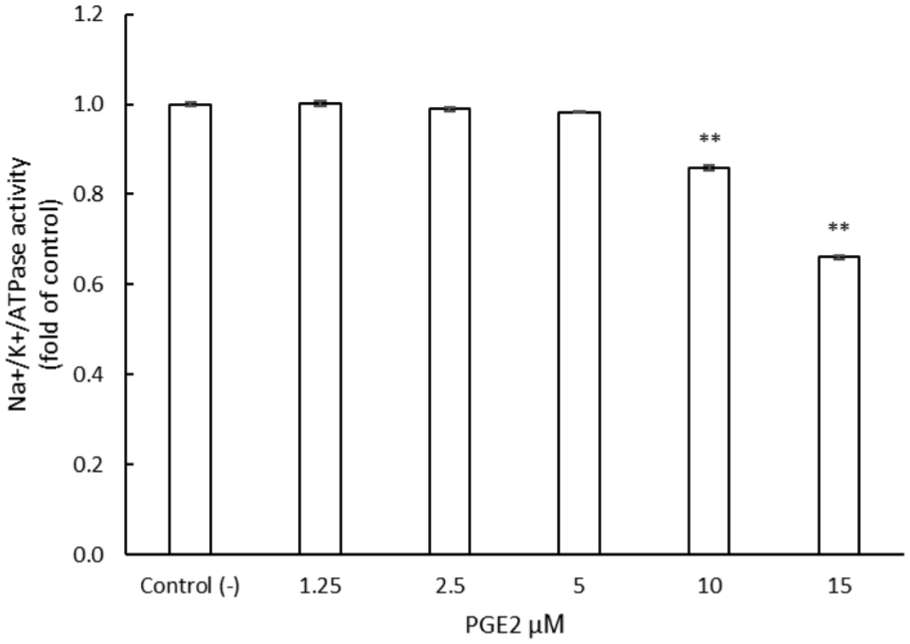
Effect of PGE2 on Na+/K+-ATPase activity in SH-SY5Y cells.
SH-SY5Y cells were exposed to increasing concentrations of PGE2, and Na+/K+-ATPase activity was measured. PGE2 significantly reduced enzyme activity at concentrations of 10 and 15 µM, indicating inhibition of Na+/K+-ATPase under inflammatory stimulation (data were presented as mean ± SEM, **p < 0.01 vs control).
Metformin reverses PGE2-induced nociceptive signaling in vitro
Metformin restores Na+/K+-ATPase activity
As shown in Figure 3, metformin restores Na+/K+-ATPase activity in a concentration-dependent manner. At 6 mM, metformin produced a modest increase to 0.855 ± 0.007-fold of control, while 8 and 10 mM increased activity to 0.93 ± 0.0145 and 1.01 ± 0.0116-fold, respectively, significantly reversing the inhibitory effect of PGE2 (0.66 ± 0.0024) and restoring enzyme activity to nearly control levels. Prednisone 1.5 µM, used as a positive control, similarly reversed PGE2-induced inhibition, increased activity to 1.09 ± 0.00496-fold of control. All values are expressed as fold of control. These results indicate that metformin effectively counteracts the suppression of Na+/K+-ATPase activity mediated by PGE2.
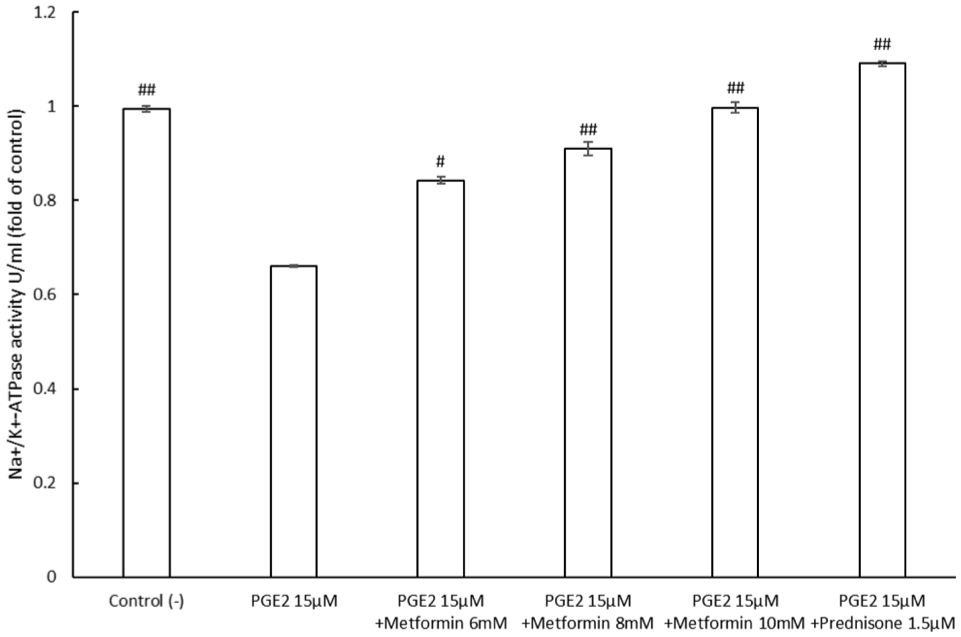
Metformin reverses PGE2-induced inhibition of Na+/K+-ATPase activity.
Metformin reduces intracellular cAMP levels
As illustrated in Figure 4, PGE2 markedly increased intracellular cAMP levels compared with the control, rising from 0.368 ± 0.012 to 1 ± 0.01. Treatment with metformin or prednisone following PGE2 exposure significantly reduced cAMP levels relative to the PGE2 group. Specifically, cAMP levels decreased to 0.807 ± 0.009 with 6 mM metformin, 0.502 ± 0.005 with 8 mM metformin, and 0.451 ± 0.009 with 10 mM metformin. Treatment with 1.5 µM prednisone further reduced cAMP levels to 0.364 ± 0.015. All values are expressed as fold relative to the PGE2 group. These results show that metformin reduces intracellular cAMP concentration in SH-SY5Y cells, counteracting the inhibitory effects of PGE2.
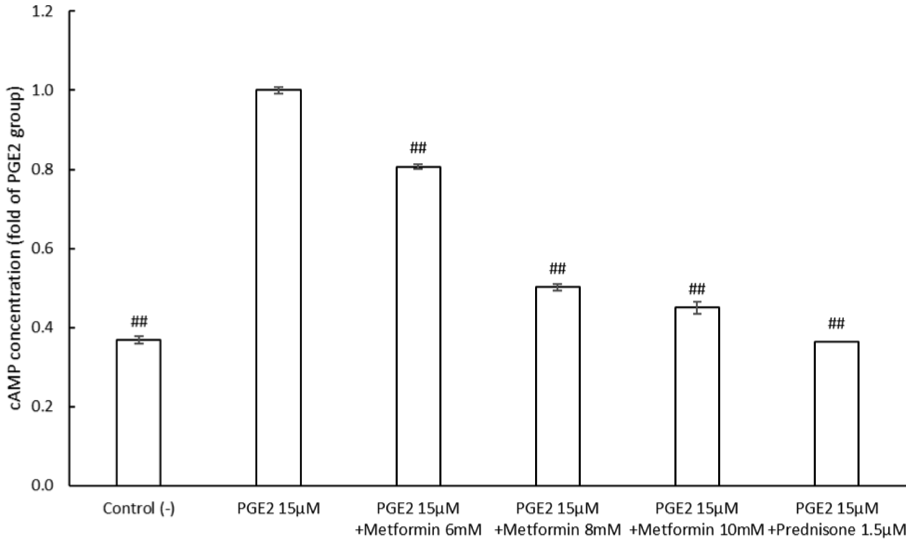
Metformin suppresses intracellular cAMP in SH-SY5Y cells exposed to PGE2.
SH-SY5Y cells were treated with PGE2 alone or co-treated with metformin or prednisone. Na+/K+-ATPase activity was measured and normalized to untreated control cells. PGE2 markedly suppressed enzyme activity, whereas metformin dose-dependently restored Na+/K+-ATPase function. Prednisone served as a positive anti-inflammatory control. (All values are expressed as fold of control, data were presented as mean ± SEM #p < 0.05, ##p < 0.01 vs PGE2 group).
Intracellular cAMP levels were measured in SH-SY5Y cells treated with PGE2 in the presence or absence of metformin or prednisone. PGE2 significantly increased cAMP levels, while both metformin and prednisone significantly reduced cAMP accumulation, suggesting inhibition of PGE2-mediated signaling (data were presented as mean ± SEM, **p < 0.01 vs PGE2).
Establishment of a CFA-induced in vivo pain model
Figure 5(a) showed that plantar injection of 5 µl CFA induced a mild inflammatory response, at the higher doses (10 and 15 µl), mice exhibited prominent signs of inflammation, including swelling, redness, and edema; in some cases, toe curling or extension was observed, although gross differences between these two groups were difficult to discern visually. Following CFA administration, mice displayed altered gait patterns, characterized by reduced weight-bearing on the affected hind paw. Mild alopecia was occasionally observed, but no changes in appetite, anxiety-related behaviors, or significant body weight loss occurred during the first week. By approximately day 10 post-injection, the inflamed region began to resolve, and by week 3, no observable differences remained compared with control animals.
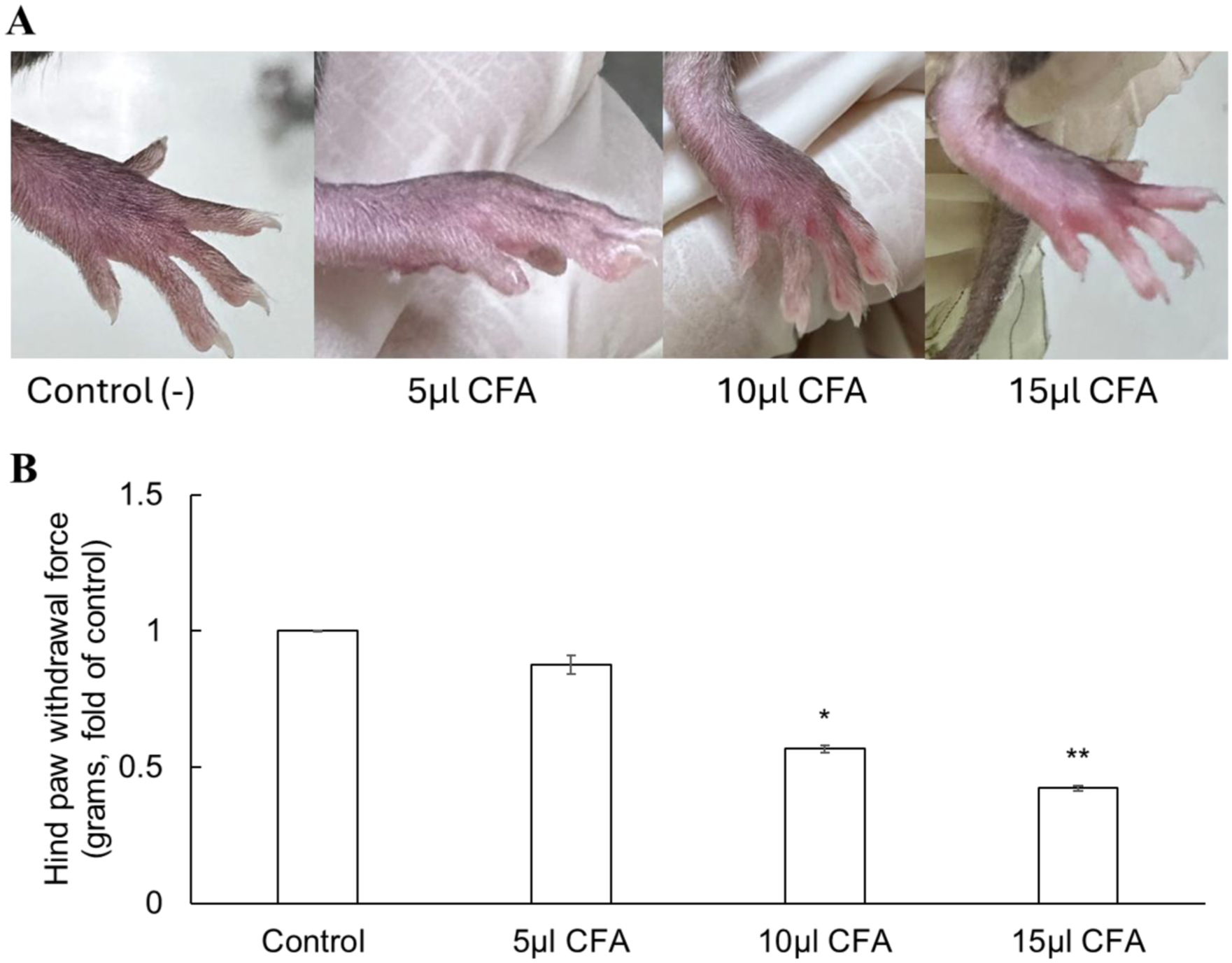
Mechanical hypersensitivity in mice 3 days after CFA injection. (a) Hindpaw of C57BL/6 mice 3 days after intraplantar CFA injection. Observed changes include redness, swelling (edema), localized inflammation, and curling or extension of the toes. (b) Hind paw withdrawal thresholds decreased in a dose-dependent manner (5, 10, and 15 µl CFA). Data are presented as mean ± SEM. *p < 0.05, **p < 0.01 versus control.
Mechanical hypersensitivity was assessed in C57BL/6 mice 3 days after plantar injection of CFA at different volumes (5, 10, and 15 µl) using the von Frey test (Figure 5(b)). Hind paw withdrawal thresholds decreased in a dose-dependent manner, with values of 0.875 ± 0.034, 0.567 ± 0.014, and 0.423 ± 0.009 fold of control for 5, 10, and 15 µl CFA, respectively (n = 6 mice/group). Statistically significant reductions were observed at 10 µl (p < 0.05) and 15 µl (p < 0.01) compared with control mice.
These results indicate that 15 µl CFA produces the most robust mechanical hypersensitivity, and this volume was therefore selected for subsequent experiments as the standard dose for establishing the inflammatory pain model.
Metformin exerts analgesic effects in vivo
Peak analgesic effect at 4 h post-injection
Analysis of Figure 6 reveals that the administration of metformin at doses of 0.05 and 0.1 g/kg/day, and prednisone at 0.015 g/kg/day, significantly increased the hind paw withdrawal threshold (grams) compared with the control, by 1.66 (p < 0.05), 1.67 (p < 0.05), and 2.5-fold (p < 0.01), respectively, with the peak effect observed at 4 h. Consequently, subsequent experiments measured responses at 4 h post-administration, including assessments on days 4, 7, 14, and 21 to track analgesic effects over time. This result confirms that metformin exerts a measurable analgesic effect, with the most pronounced response observed at 4 h post-administration.
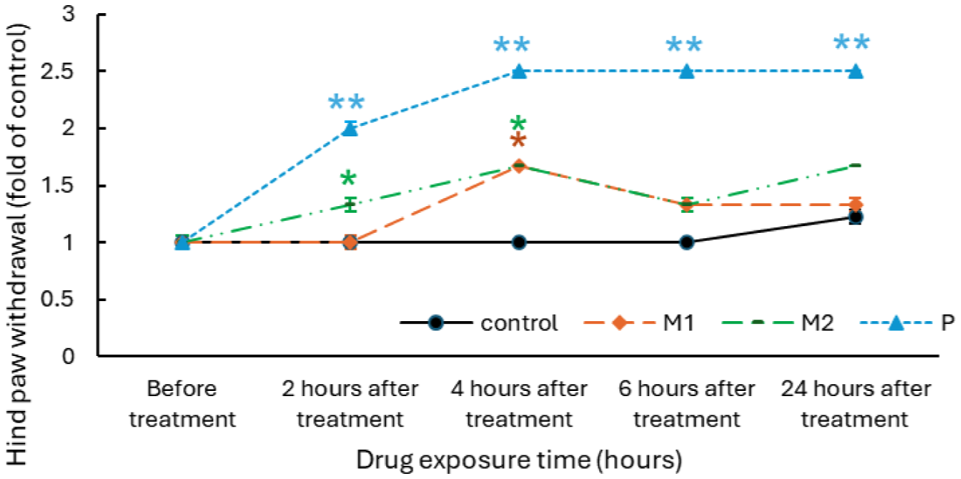
Time-course analysis of the analgesic effect of Metformin in C57BL/6 mice. Mechanical pain responses were evaluated in C57BL/6 mice following administration of metformin (M1, M2) or prednisone. Both doses of metformin and prednisone produced significant analgesic effects, with maximal pain relief observed 4 h post-injection (data were presented as mean ± SEM, *p < 0.05, **p < 0.01 vs control at the same time point).
Metformin increases hind paw withdrawal threshold in mice
In the Von Frey test, metformin significantly increased hind paw withdrawal thresholds over 21 days compared to saline controls, indicating improved mechanical pain tolerance. According to Figure 7, On day 1, all treated groups showed modest increases, with prednisone exhibiting the strongest early effect, approximately 2.5 times control. From day 4, the 0.1 g/kg/day metformin group (M2) consistently exceeded the 0.05 g/kg/day group (M1), peaking on day 7 at 1.5 and 1.28 times control (p < 0.05), respectively, while prednisone maintained the greatest effect about 2.16 times control (p < 0.01). After day 7, thresholds gradually rose across all groups, reflecting natural recovery, and by day 21, values converged about 7–8 times baseline with no significant differences. These results demonstrate a dose-dependent analgesic effect of metformin, most pronounced within the first 7–14 days before merging with spontaneous recovery.
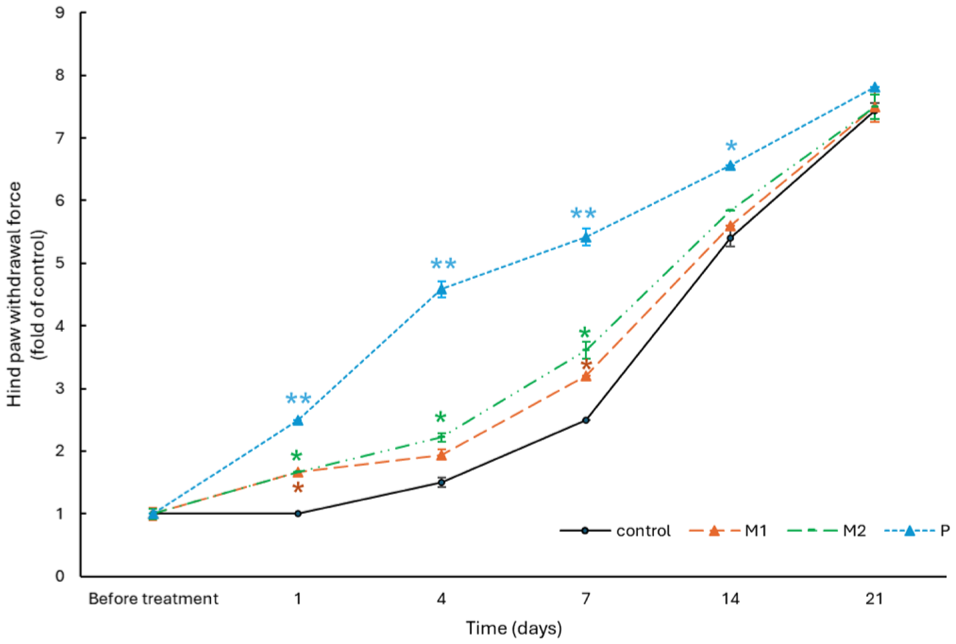
Metformin enhances mechanical pain threshold in mice, with higher doses producing greater analgesic effects during the early phase of treatment. Time course of hind paw withdrawal force in C57BL/6 mice over 21 days after daily treatment with metformin (M1, M2), prednisone (P), or Control (C). Data are shown as fold change from control baseline (mean ± SEM). Significant differences occurred on days 4 and 7, notably in M2 and P groups (*p < 0.05, **p < 0.01 vs control).
Metformin reduces TRPV1 and NaV1.7 expression in vivo
Western Blot analysis
To elucidate the molecular basis of the behavioral analgesic effects, Western blot analysis was performed on left hind paw tissues collected on day 7, when differences in withdrawal thresholds were most pronounced. As shown in Figure 8, quantitative analysis of NaV1.7 expression (normalized to β-actin) revealed a dose-dependent decrease: Control (1) > M1 (0.783 ± 0.048) > M2 (0.677 ± 0.099) > prednisone (0.526 ± 0.135), with M2 and prednisone groups showing significantly lower levels than control, p < 0.05. Similarly, TRPV1 expression declined in a dose-dependent manner, reaching its lowest level in the prednisone group (0.445 ± 0.08, p < 0.01), followed by M2 (0.66 ± 0.107, p < 0.05) relative to control. These quantitative results confirm significant reductions in TRPV1 in the M2 and prednisone groups, indicating attenuated peripheral nociceptive sensitization.
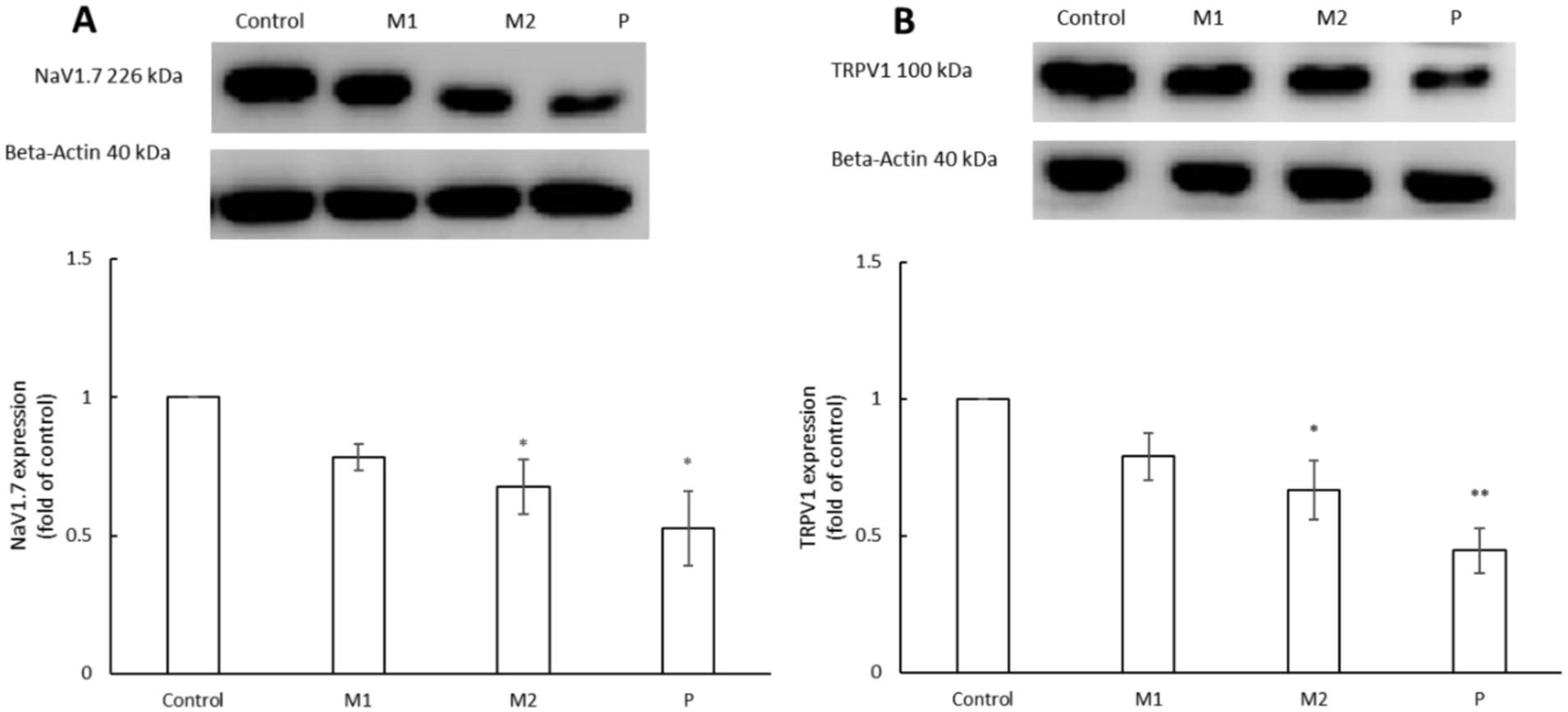
Metformin regulates the expression of pain-associated markers NaV1.7 and TRPV1 in mouse paw tissue. Protein expression levels of NaV1.7 (a) and TRPV1 (b) in mouse paw tissue were analyzed by Western blotting following treatment with metformin (M1, M2), prednisone (P), or control (C). Representative immunoblots and corresponding densitometric quantification normalized to the loading control are shown. metformin treatment, particularly at the higher dose (M2), significantly reduced NaV1.7 and TRPV1 protein expression compared with control. Prednisone served as a positive control (data were presented as mean ± SEM, *p < 0.05, **p < 0.01 vs control).
Immunohistochemical analysis
As illustrated in Figure 9, IHC staining revealed reduced NaV1.7 and TRPV1 expression in metformin- and prednisone-treated tissues compared with saline controls. The consistent downregulation of NaV1.7 across treatment groups supports a biologically relevant effect, consistent with Western blot findings. TRPV1 showed a clear dose-dependent decrease, with the most pronounced reduction in the high-dose metformin group (M2, 0.1 g/kg/day), where TRPV1-positive areas declined from 25% in controls to 7%. Both proteins were mainly localized in the central region corresponding to the CFA injection site.
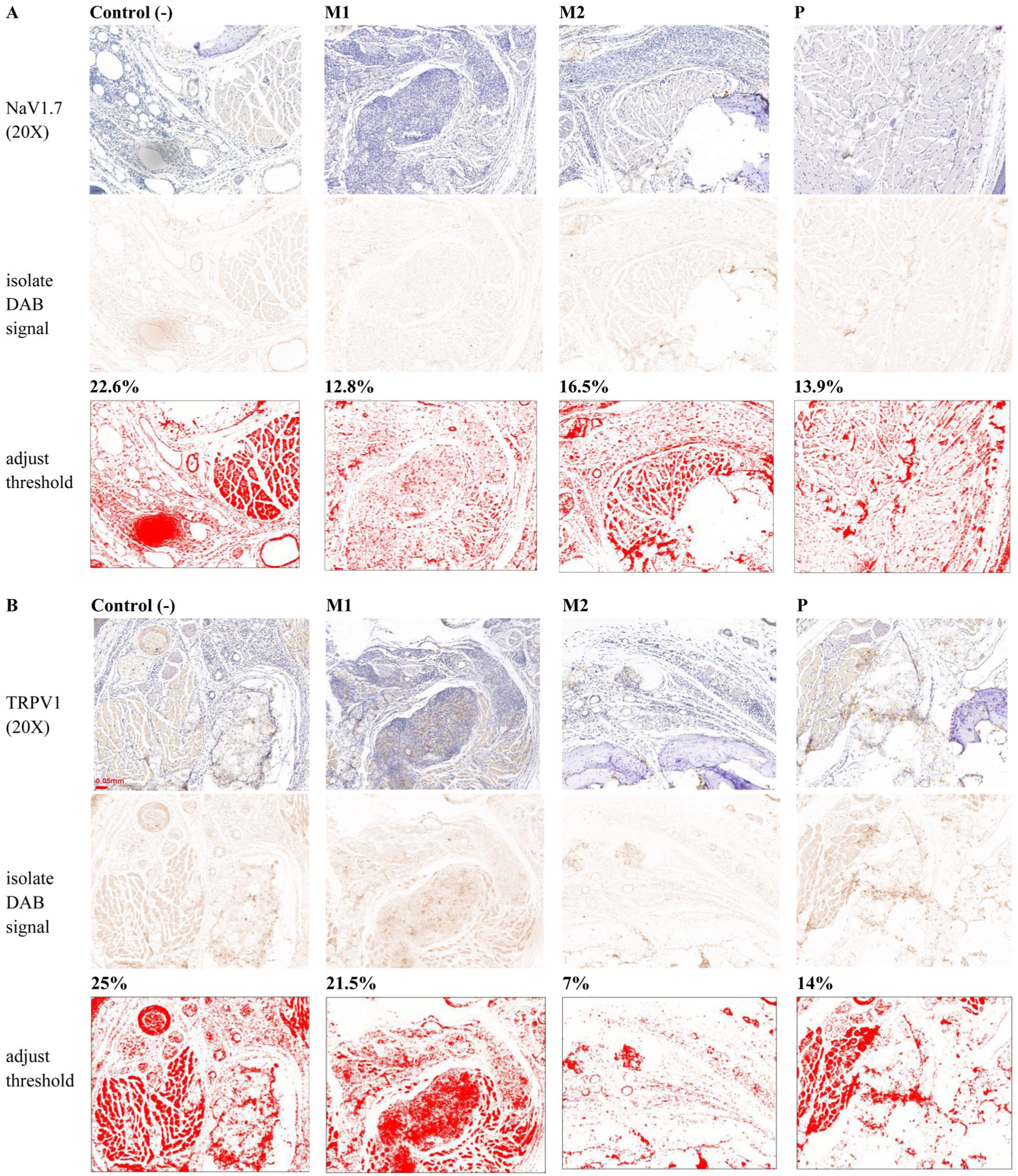
Metformin suppresses NaV1.7 and TRPV1 expression in tissue immunohistochemical staining was performed to visualize the tissue distribution of NaV1.7 and TRPV1 in mouse paw sections from control (C), metformin-treated (M1, M2), and prednisone-treated (P) groups. Representative images at low (10×) and high (100×) magnification are shown. Compared with control tissue, reduced staining intensity of NaV1.7 and TRPV1 is observed in metformin-treated groups, particularly at the higher dose (M2), consistent with the Western blot results shown in Figure 8.
Metformin promotes inflamed tissue recovery, supporting its analgesic effect
Histological analysis revealed varying chronic inflammation across groups (Figure 10). The control group (C) showed severe inflammation beneath the dermis, with dense infiltration of lymphocytes, histiocytes, and PMNs. M1 displayed moderate inflammation with a smaller inflammatory area, marked lymphocytes, minimal histiocytes, and moderate PMNs. M2 exhibited the greatest anti-inflammatory effect, showing only a small inflammatory area, moderate lymphocytes, and minimal histiocytes and PMNs. The positive control (P) showed negligible inflammation, confirming effective suppression. The inflammatory scores are summarized in Table 2.
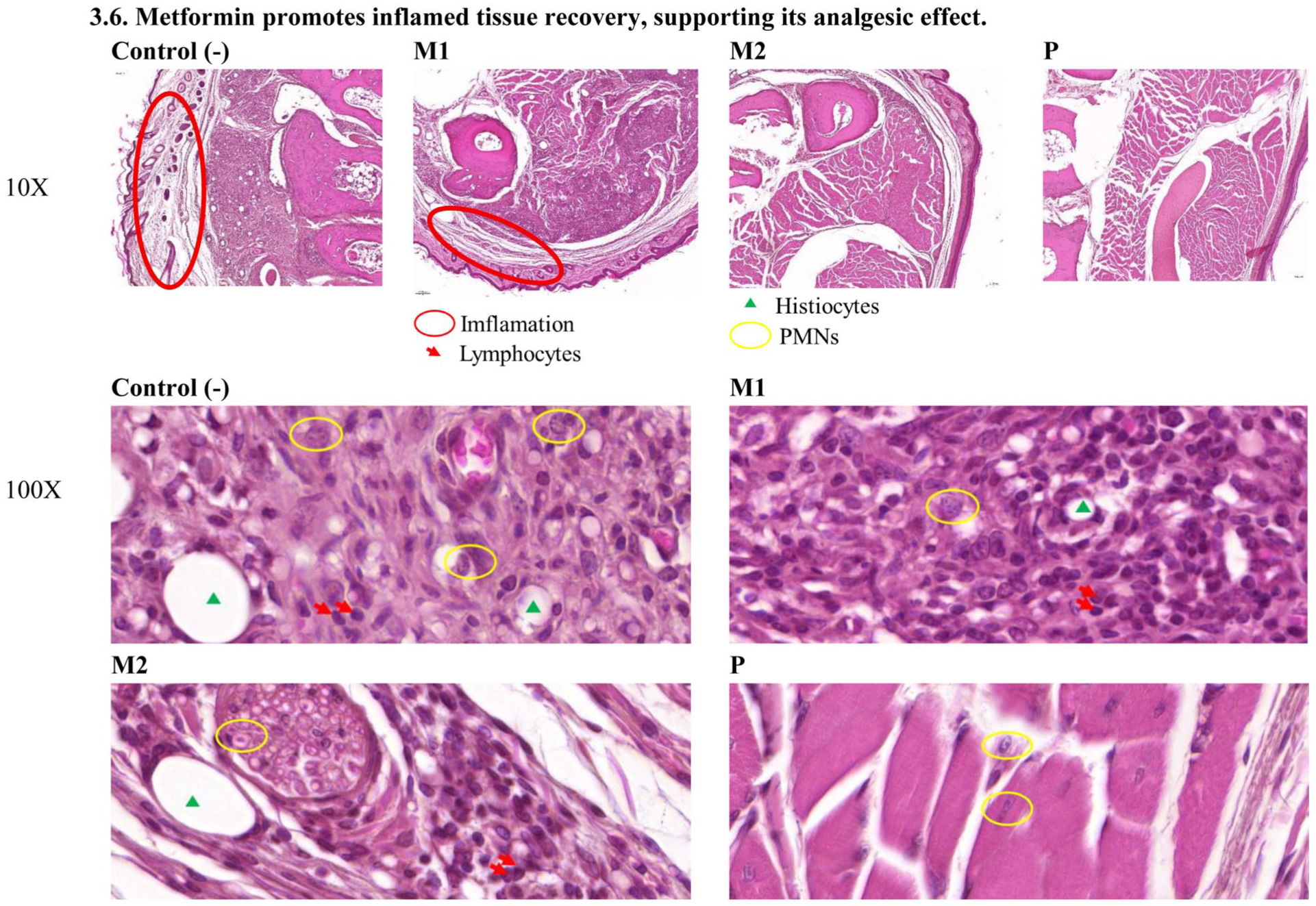
Analgesic action of Metformin is accompanied by improved inflammatory tissue recovery. Hematoxylin and eosin (H&E)–stained sections of mouse paw tissue were examined to assess inflammatory changes following treatment with metformin (M1, M2), prednisone (P), or control (C). The control group shows marked inflammatory cell infiltration and tissue edema. In contrast, metformin-treated groups display reduced inflammatory infiltration, with a more pronounced improvement observed in the M2 group. Minimal inflammatory features are evident in the prednisone-treated groupInflammation is most severe in control group and gradually decreases across M1 and M2 groups, with minimal presence in the P group (positive group).
Inflammatory assessment results.
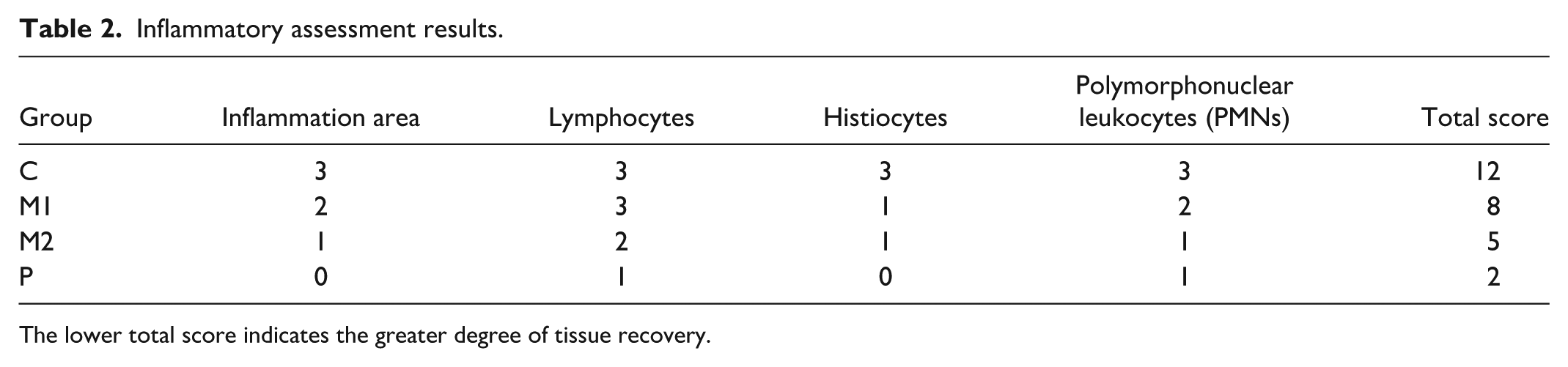
The lower total score indicates the greater degree of tissue recovery.
Discussion
Validation of the PGE2-induced in vitro pain model
Determination of Metformin IC50 in SH-SY5Y cells
Based on the experimentally determined IC50 of metformin in SH-SY5Y cells (20.99 mM), three sub-toxic concentrations (6, 8, and 10 mM; ≤1/2 IC50) were selected for subsequent experiments. This concentration range was chosen to preserve cellular viability and minimize cytotoxicity, thereby enabling reliable assessment of metformin regulatory and signaling effects in viable neurons without confounding influences from apoptosis or necrosis. Although these in vitro concentrations exceed clinically achievable plasma levels, their selection below the IC50 ensures that the observed restoration of Na+/K+-ATPase activity and suppression of cAMP signaling reflect genuine mechanistic actions of metformin rather than nonspecific cell damage.
PGE2 induces pain-like cellular responses in SH-SY5Y cells
The functional assay assessing Na+/K+-ATPase activity revealed that PGE2 induces a clear, concentration-dependent inhibition of the enzyme in SH-SY5Y cells. As outlined in the Introduction, PGE2 is known to modulate neuronal excitability and disturb ionic homeostasis under inflammatory conditions,12–14 and our findings are consistent with this established role. A marked reduction in pump activity was observed at 15 µM PGE2, indicating its effectiveness as a stimulus for modeling inflammatory conditions in vitro. Collectively, these results provide a rational basis for the dosing strategy and confirm the suitability of the experimental conditions for investigating the cellular mechanisms underlying the effects of metformin in PGE2-induced dysfunction.
Mechanisms underlying metformin-mediated modulation of nociceptive signaling
In this study, metformin effectively restored Na+/K+-ATPase pump function in SH-SY5Y cells exposed to PGE2 in a concentration-dependent manner. While 6 mM induced a modest increase, 8 and 10 mM significantly reversed PGE2-mediated inhibition, restoring enzyme activity to near control levels, similar to the effect observed with 1.5 µM prednisone. These findings indicate that both agents can counteract PGE2-induced disruption of ionic homeostasis, supporting a potential analgesic effect via normalization of enzyme activity under inflammatory conditions.
Consistent with previous reports, PGE2 markedly increased intracellular cAMP levels by activating EP2/EP4 receptors, which contributes to nociceptive sensitization.26,27 Treatment with metformin or prednisone significantly reduced PGE2-induced cAMP in a concentration-dependent manner, with metformin most effective at 10 mM, comparable to prednisone. Elevated cAMP enhances ion channel activity and promotes the expression of nociceptive genes,26,28 and its reduction by metformin may help decrease neuronal excitability and inflammatory pain.
PGE2 can amplify its own production via a positive feedback loop by upregulating COX-2 and mPGES-1 expression, thereby increasing endogenous PGE2 synthesis and sustaining inflammatory and pain responses.29,30 Previous studies have shown that both prednisone and metformin suppress endogenous PGE2 production through inhibition of COX-2 expression or activity.15,16,31,32 However, no evidence currently indicates that these agents directly inhibit PGE2 itself. Thus, it is likely that they interfere with the PGE2 positive feedback pathway through suppression of COX-2. Accordingly, the observed enhancement of Na+/K+-ATPase activity and reduction in intracellular cAMP following treatment with prednisone or metformin may be partly attributed to their inhibitory effects on endogenous PGE2 production.
Together, these results suggest that metformin mitigates the biochemical consequences of PGE2 exposure through both restoration of Na+/K+-ATPase activity and suppression of cAMP signaling, providing a mechanistic basis for its anti-inflammatory and analgesic properties.
Relevance of the CFA-induced in vivo pain model
Mechanical sensitivity testing using the von Frey assay demonstrated a dose-dependent reduction in paw withdrawal thresholds following CFA administration. Notably, mice receiving 10 and 15 µl CFA showed statistically significant decreases compared with control animals, indicating the development of mechanical hypersensitivity. Among the tested conditions, the 15 µl CFA group exhibited the most pronounced reduction in withdrawal threshold, suggesting a stronger and more consistent induction of inflammatory pain.
Importantly, despite the robust local inflammatory response, mice did not exhibit significant changes in body weight, feeding behavior, or anxiety-related behaviors during the early phase, indicating that the model induces localized and controlled inflammatory pain without causing severe systemic distress. Furthermore, the gradual resolution of inflammation by approximately day 10 and the absence of observable differences by week 3 suggest that this model captures a transient yet sustained inflammatory pain state, which is suitable for evaluating analgesic interventions.
Taken together, these findings confirm that plantar CFA injection produces a reliable and reproducible model of inflammatory pain in mice. Based on the observed dose-dependent effects and the robust mechanical hypersensitivity induced, 15 µl CFA was selected as the optimal condition for subsequent experiments.
Analgesic effects of metformin in vivo
Peak analgesic effect at 4 h post-injection
The analgesic profile of metformin indicates a clear time-dependent pattern, with maximal efficacy observed at 4 h post-injection. The gradual decline in analgesia after 4 h likely reflects metabolic clearance or feedback regulation of inflammatory mediators. Nevertheless, pain thresholds remained elevated compared with control, implying a sustained modulatory influence on peripheral nociceptive signaling. These findings highlight 4 h as the optimal window for analgesic effect of metformin in the acute phase of inflammatory pain.
Metformin increases hind paw withdrawal threshold in mice
The results show that metformin increases mechanical pain tolerance in mice in a dose-dependent manner, as measured by the Von Frey test. Analgesic effects became noticeable from day 4, with the 0.1 g/kg/day dose producing a stronger response than 0.05 g/kg/day, and were most prominent during days 7–14. After day 14, withdrawal thresholds gradually converged between metformin-treated and control groups, likely reflecting natural resolution of inflammation.
These findings align with previous studies demonstrating the analgesic effects of metformin in various pain models. A 200 mg/kg dose reversed mechanical allodynia in a spinal nerve injury model, with sustained effects 33 and peri-injury administration reduced postoperative mechanical hypersensitivity. 34 Similar improvements in paw withdrawal thresholds were reported in complex regional pain syndrome and chemotherapy-induced neuropathy models.35,36
Compared to these studies, which used higher doses and often more severe pain models, our data indicate that even lower doses of metformin can produce significant analgesia during the early phase of inflammation, supporting its potential for managing mechanical pain in acute and subacute stages.
Regulation of TRPV1 and NaV1.7 expression by metformin
In this study, metformin produced dose-dependent analgesic effects under inflammatory conditions, reflected by improved behavioral responses and reduced expression of key nociceptive proteins. At 0.1 g/kg/day (M2), metformin significantly decreased TRPV1 expression, as confirmed by Western blot and immunohistochemistry. This aligns with previous findings showing metformin inhibits TRPV1 via the TGFβ1–TGFβRI pathway in bone cancer pain, 37 and through AMPK-mediated suppression in inflammatory pain. 11 In our study, TRPV1-positive areas decreased from 25% in controls to 7% in M2.
NaV1.7, essential for pain signal initiation, was also downregulated in metformin-treated groups, paralleling the improved paw withdrawal thresholds. These changes are consistent with reports that metformin reduces NaV1.7 membrane localization via NEDD4-2–mediated ubiquitination. 38 and may be mediated by AMPK activation, which can inhibit NF-κB signaling and enhance mitochondrial and antioxidant functions. 39 The observed downregulation of TRPV1 and NaV1.7 of metformin may also reflect adaptive neuronal responses, reducing nociceptor activation and remodeling neuronal excitability during the transition from acute inflammation to tissue repair.
Histopathological improvements associated with metformin treatment
Mechanistically, metformin acts as an anti-inflammatory and tissue-protective agent. It reduces inflammatory infiltration and fibrosis after ischemia/reperfusion injury, 40 inhibits M1 macrophage polarization and pro-inflammatory cytokine production in osteoarthritis, 41 and suppresses TNF-α, IL-6, NF-κB, and HMGB1 through AMPK activation. 42 In inflammatory pain models, doses of 250–1000 mg/kg alleviate allodynia and edema via ATP-sensitive K+ channel activation and TNF-α inhibition. 10 Consistently, our study showed that metformin reduced lymphocyte and PMN infiltration and limited inflammatory zones, especially at higher doses (M2). At 0.1 g/kg/day, it produced histological improvements comparable to prednisone but acted more selectively by modulating macrophage polarization and cytokine release, potentially reducing systemic side effects. Overall, metformin promoted dose-dependent tissue recovery and exerted dual effects on nociceptive and inflammatory pathways, supporting its role as a safe, effective analgesic candidate.
Limitations of the study
Model considerations: SH-SY5Y cells and the CFA-induced hind paw model were used to investigate cellular mechanisms and acute inflammatory pain. Extension to DRG neurons, higher CFA doses, or alternative chronic pain models could improve translational relevance; however, such experiments would constitute a separate, more comprehensive study.
Mechanistic analysis: Direct assessment of AMPK activation and more detailed neuronal-specific analyses were not performed in this study and remain to be conducted in future investigations to clarify the underlying mechanisms.
Overall: Despite these limitations, meaningful early insights were obtained regarding analgesic effects of metformin and its impact on TRPV1 and NaV1.7, providing a foundation for future, more detailed mechanistic investigations.
Conclusion
Metformin showed good cellular viability in SH-SY5Y cells, restored Na+/K+-ATPase activity, and attenuated PGE2-induced cAMP elevation, suggesting potential modulation of neuronal sensitization and inflammatory signaling. In a mouse inflammatory pain model, metformin produced dose-dependent analgesia, improved hind paw withdrawal thresholds, suppressed TRPV1 and NaV1.7 expression, and promoted histological recovery with reduced immune infiltration. Compared with prednisone, it achieved analgesic and anti-inflammatory effects that appeared comparable, potentially through an AMPK-mediated mechanism. Collectively, these results identify metformin as a promising non-opioid candidate for inflammatory pain with both symptomatic and potential to modulate both nociceptive and inflammatory processes, warranting further validation in chronic models.
Footnotes
Author contributions
Thi-Chau-Loan Phan: Writing—original draft, Methodology, Experimental work, Data analysis, Conceptualization. Jui-Ting Chang: Methodology, Chih-Hsiang Ma: Methodology, Yao-Jen Liang: Writing—review & editing, Supervision, Methodology, Conceptualization.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Ethical statement
This study was conducted and reported in accordance with the ARRIVE guidelines to ensure transparent and reproducible animal research. All experimental procedures were reviewed and approved by the Ethics Committee of the Fu Jen Laboratory Animal Center, Fu Jen Catholic University (IACUC No. FJU-A11232).
Data availability statement
The data supporting the findings of this study are available upon request. Please contact the corresponding author for access to the data.