
Abstract
Normal sensitivity of the Drosophila larval nociceptor requires the activity of the Wg/WNT signaling pathway. When the nociceptor expression of Wg/WNT co-receptor Arrow, Drosophila ortholog of LRP5/6, is impaired by RNA interference, the larva becomes significantly hyposensitive to noxious thermal and mechanical stimulation. The reduced sensitivity was not associated with significant changes in dendritic architecture.
Introduction
Pain is essential for protecting the body from harm. It reduces injury and promotes healing by alerting us to tissue damage. 1 However, abnormal pain states including chronic pain affect approximately 30% of people worldwide and 20% of Americans, 2 costing roughly $600 billion annually in treatment and lost productivity. 3 Opioids are a primary treatment method, 4 but addiction liability has fueled a global overuse and overdose crisis. Of the 63,623 overdose deaths in 2016 in the United States, 66.4% involved opioids. 5 More information on alternative analgesic drug targets is sorely needed. Many of the complex biological pathways and processes, including many components of neurogenesis, are highly conserved between fruit flies and humans, 6 suggesting that manipulation of the orthologs of targets identified in fruit flies may be effective at reducing pain in humans.
The Class 4 dendritic arborization (C4da) neurons of the Drosophila larva serve as mechanical and thermal primary nociceptors. In response to noxious stimuli, these sensory neurons trigger an escape response consisting of a lateral rolling about the larva’s longitudinal axis. 7 Various cellular signaling pathways have been shown to influence the sensitivity of Drosophila nociceptors. It has been shown that Hedgehog signaling is a regulator of injury-induced allodynia, 8 and downstream Bone Morphogenetic Protein (BMP) signaling controls nociceptive sensitivity via Decapentaplegic. 9 Other BMP components, including Glass Bottom Boat, Wishful Thinking, Dally, and Brinker, also influence nociceptive sensitization.10–13 The Wingless/Wingless Integration Site (Wg/WNT) pathway has been linked to chronic and neuropathic pain in humans14,15 and also explored in flies. Overexpression of a central Wg/WNT component, Armadillo, the Drosophila homolog of β-catenin16,17 in the fly nociceptor has been shown to increase nociceptive sensitivity, while its suppression reduces sensitivity, 18 indicating the Wg/WNT pathway’s ability to affect nociception.
Arrow, which, with Frizzled, is a receptor for Wingless, helps to activate the pathway and increase Armadillo’s availability.19,20 Mutations in Arrow’s mammalian ortholog, LRP5/6 21 are linked to diseases such as osteoporosis and cancer. 22 This study tests the hypothesis that Wg/WNT receptor Arrow is a mediator of the Wg/WNT pathway’s influence on nociceptive sensitivity in Drosophila. Flies with reduced Arrow expression were measured for altered responses to mechanical and thermal stimuli. Morphometric approaches were employed to assess whether altered sensitivity correlated with alterations to morphology of larval nociceptors.
Methods
Fly Husbandry and Genetics: Flies were obtained from the Bloomington Drosophila Stock Center (BDSC) and maintained in 6oz stock bottles containing sucrose-cornmeal-yeast medium in Invictus Next Gen incubators set to 25°C and 55% humidity. The incubators operated on a 12-h light and dark cycle (9am–9pm). Experimental genotypes were prepared using the Gal4/UAS system23,24 with Gal4 driver line featuring the pickpocket promoter: ppk1.9-GAL425–27 (in w1118). ppk1.9-Gal4, UAS-mCD8-GFP (in yw) was used for neuromorphometric and imunohistofluorescent analysis. Transgenic lines included: UAS-Arr IR1 (in y1v1) (BDSC_53342) and UAS-Arr IR2 (BDSC_82978).28,29 Wild-type control lines for the above Transgenic RNAi Project lines used were w1118 (BDSC 3605) and y1v1 (BDSC 36303). Each Gal4/UAS genotype used in the thermal and mechanical nociception assays was compared with two normal controls: one with the genetic background (w1118) of the Gal4 driver crossed with the UAS transgenic line (no Gal4 control) and one with the Gal4 driver line crossed to the genetic background (y1v1) of the UAS transgenic line (no UAS control).
Thermal Nociception Assays: Foraging third instar larvae were assayed using previously described methods.7,8 A thermal probe (ProDev Engineering, Missouri City, TX) set to a predetermined noxious temperature of 45°C 30 was placed on the dorsal side of selected larvae between segments A5 and A7. The probe was removed once a larva displayed the typical nocifensive escape behavior of a 360° roll or 20 s elapsed. Data were recorded as either the latency of response or no response.
Mechanical Nociception Assays: Drosophila larvae also exhibit the rolling escape response to mechanical stimulation. 31 A von Frey filament calibrated to 3951 kPA of pressure was applied to the posterior dorsal side, segment A8 of a third instar larva for up to 2 s until the probe bent. The probe was then removed, and a positive response was recorded if the larva rolled 360° within 10 s. If there was no roll within 10 s, the larva was recorded as a nonresponder.
Immunohistofluorescent analysis: Third instar larvae expressing GFP in their nociceptors (ppk1.9-Gal4, UAS-mCD8-GFP) were filleted using previously described methods. 9 Fillets of normal and Arrow-underexpressing larvae were incubated together in (1:150) guinea pig anti-Arrow. 32 No primary control consisted of fillets of normal larvae incubated in PBTB only. All fillets were washed and then incubated with goat anti guinea pig secondary antibody, diluted to 1:500 in PBTB +5% NGS. Fillets were mounted using Vectashield Antifade Mounting Medium with DAPI (H-1200, Vector Laboratories).
Imaging and CTCF analysis: Single nociceptors from third instar larvae fillets were imaged using a Leica Stellaris laser scanning confocal microscope, with a 40×/1.30 oil immersion objective. Z-stacks were obtained with a system optimized step size, and a scan format of 2048 × 2048. Using previously described methods,18,33 settings were obtained uniformly across experiments and control fillets including smart gain, laser power, zoom, frame averaging, and pinhole. Using Fiji 34 five z-slices from each nociceptor were sum projected and cropped to remove all but the soma. A macro 18 was used to establish a consistent approach to making masks and eliminate selection bias. Soma ROIs were used to measure area and integrated density in Fiji 34 for fluorescence within the cropped projections. CTCF analysis was used to measure protein expression levels of Arrow in both control and experimental samples. 35 Using previously described methods. 18 CTCF and integrated density for each sample was measured, averaged and a student’s t test with Welch’s correction was applied.
Neuromorphometry: To evaluate whether behavioral changes corresponded to neuronal morphological alterations, dendritic length and branching were measured in vivo. Third instar larvae were anesthetized with ether for up to 4 min. Larvae measuring between 4.0 and 4.5 mm in length were selected for imaging. Larvae were placed on microscope slides in a halocarbon-ether mixture (2:1) with a 22 × 55 mm coverslip for live imaging. A Leica Stellaris laser scanning confocal microscope at 20×/0.04 objective with zoom set to 0.7 was used to image single GFP (Alexa 488) tagged neurons. A resolution of 1024 × 1024 was used and z-stacks incorporated the very beginning to ending of neuron visualization. Images were converted to max projected tifs, traced and measured for dendritic length and branching with Fiji SNT. 36 Images were measured using Fiji AnalyzeSkeleton (2D/3D) plugin. 34
Statistical analysis: Thermal nociception assays were analyzed using Log-rank analysis in R statistical coding software with the survival analysis package. 37 Benjamini and Hochberg procedure was applied for multiple testing. Mechanical nociception assays were evaluated using chi square analysis. Calculation of dendritic length and branching morphology was performed using Student’s t-test with Welch’s correction for unequal variances. CTCF and Integrated Density analysis was analyzed using Student’s t test with Welch’s correction. All other statistical tests and plots utilized Microsoft Excel and Real Statistics Resource Pack software package (Release 7.6).
Results
To determine if Arrow assists in regulating nociceptive sensitivity, we used the Gal4/UAS system in conjunction with RNAi to underexpress Arrow. The Gal4 driver used was ppk1.9-Gal4, which targets Class IV dendritic arborization neurons (Figure 1), along with inverted repeat lines targeting the Arrow mRNA. We used immunohistofluorescence (IHF) to quantify and compare Arrow protein levels in underexpressing and normal control animals. Results confirmed the presence of Arrow protein in the nociceptor as well as other cells of the larva (Figure 1(a) and (b)). Corrected Total Cell Fluorescence (CTCF) analysis using background subtraction from no primary controls (Figure 1(a)) indicated that Arrow fluorescence was significantly reduced in nociceptors of underexpresser animals (13545 a.u. Figure 1(c) and (d)), compared with those of normal control animals (31192 a.u., Figure 1(b) and (d)). It was observed that Arrow was also significantly reduced in non-nociceptor cells of underexpresser animals (257187 a.u., Figure 1(c) and (e)), as compared with those of normal control animals (698050 a.u., Figure 1(b) and (e)).
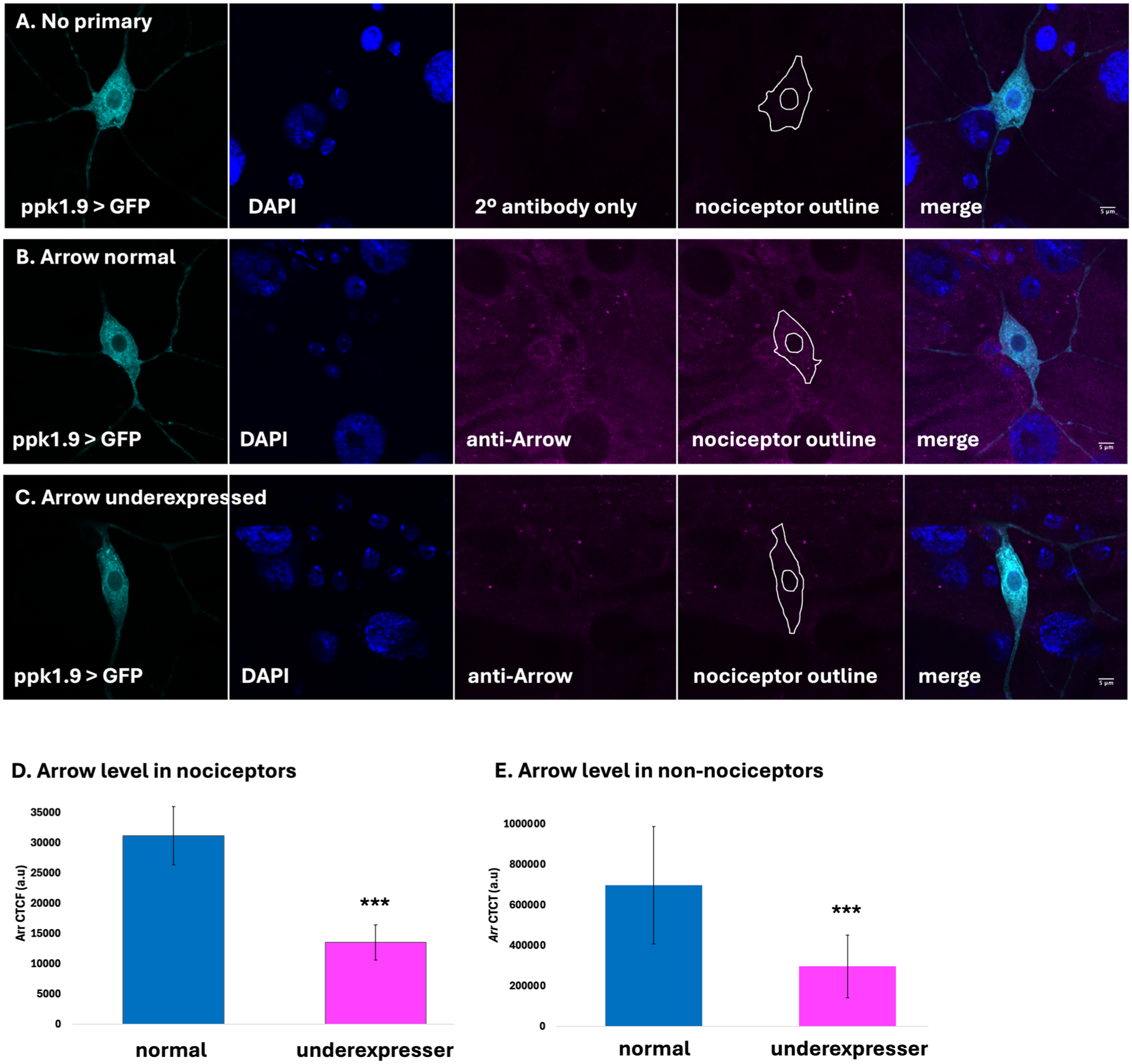
Immunohistofluorescent analysis demonstrates Arrow underexpression in experimental genotype. (a) Ppk1.9-Gal4, UAS-mCD8-GFP over y1v1, incubated with the secondary antibody but no primary antibody results in a very low amount of background staining (magenta). (b) Ppk1.9-Gal4, UAS-mCD8-GFP over y1v1, incubated with primary anti-Arrow antibody and secondary antibody. Arrow (magenta) is observed in the nociceptor (outlined) as well as other cells. (c) Ppk1.9-Gal4 driving both UAS-mCD8-GFP and UAS-Arr-IR2 with primary anti-Arrow antibody and secondary antibody. (d, e) Results of CTCF analysis, with background fluorescence corrected using the signal measured in the no-primary sample, shows that Arrow is significantly underexpressed in nociceptor cells (d) and non-nociceptor cells (e). *** indicates p < 0.001.
To determine if underexpression of Arrow influenced larval response to a noxious thermal temperature, third instar larvae were stimulated using a thermal probe set to 45°C. 30 Arrow underexpression via Arr IR 1 produced a statistically significant reduction in nocifensive response frequency compared to normal control animals (Figure 2(a)). A second non-overlapping Arrow IR line (Arr IR 2 ) was tested, with similar results (Figure 2(b)). A mechanical nociception assay 31 was conducted to determine whether Arrow mediates response to noxious mechanical stimulation. When stimulated with a pressure of 3951 kPa, animals underexpressing Arrow showed a significant (p < 0.001, chi-square) decrease in sensitivity, compared with normal controls (Figure 2(c)).
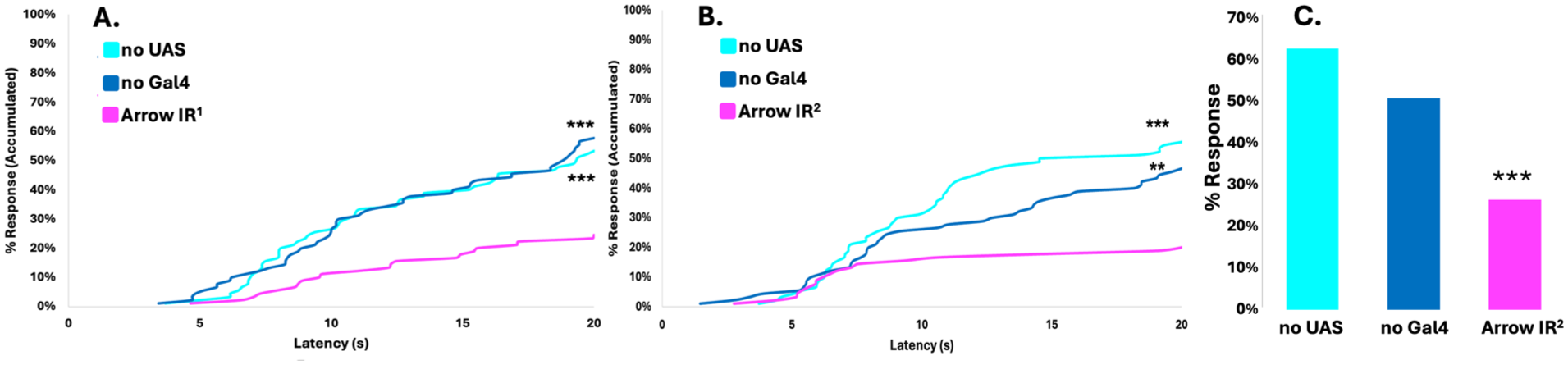
Arrow underexpression results in reduced nociceptive sensitivity. (a) Percent response plotted against response latency for thermal assays at 45°C for experimental animals ppk1.9-Gal4 > UAS-Arr-IR1, shown in magenta, compared with normal controls ppk1.9-Gal4 > y1v1 (no UAS), shown in cyan and w1118 > UAS-Arr-IR1 (no Gal4), shown in blue. After 20 s, a significantly lower proportion, 24%, of the experimental group had responded, compared to the normal controls no UAS (53%), and no Gal4 (58%). (b) The percent response was plotted against response latency for thermal assays at 45°C for a second experimental genotype, ppk1.9-Gal4 > UAS-Arr-IR2, shown in magenta, compared with normal controls ppk1.9-Gal4 > y1v1, (no UAS), shown in cyan, and w1118 > UAS-Arr-IR2 (no Gal4), shown in blue. After 20 s, a significantly lower proportion, 20%, of the experimental group had responded, compared to the normal controls no UAS (56%) and no Gal4 (47%). Every group had n = 90, results were statistically analyzed using log rank analysis. (c) Animals of experimental genotype ppk1.9-Gal4 > UAS-Arr-IR2 responded to noxious mechanical stimulation (3951 kPa) significantly less frequently (26%) than normal controls ppk1.9-Gal4 > y1v1, (no UAS), (63%), and w1118 > UAS-Arr-IR2 (no Gal4), (51%). Each group (N > 90), distributions analyzed using chi-square. ** indicates p < 0.01, *** indicates p < 0.001.
To assess whether Arrow underexpression affected nociceptor dendritic development, morphometric analysis of the dendritic arbor was performed by measurement of dendritic length and branching pattern. To visualize nociceptor morphology, nociceptor neurons were made to express GFP via ppk1.9-Gal4, allowing for comparison between the nociceptor dendritic arbor of animals with normal Arrow expression and those in which Arrow was underexpressed (Figure 3(a)). Statistical analysis using a two-tailed t-test with Welch’s correction revealed no significant differences in total dendritic length (Figure 3(b)) or dendritic branching (Figure 3(c)) between the two groups.
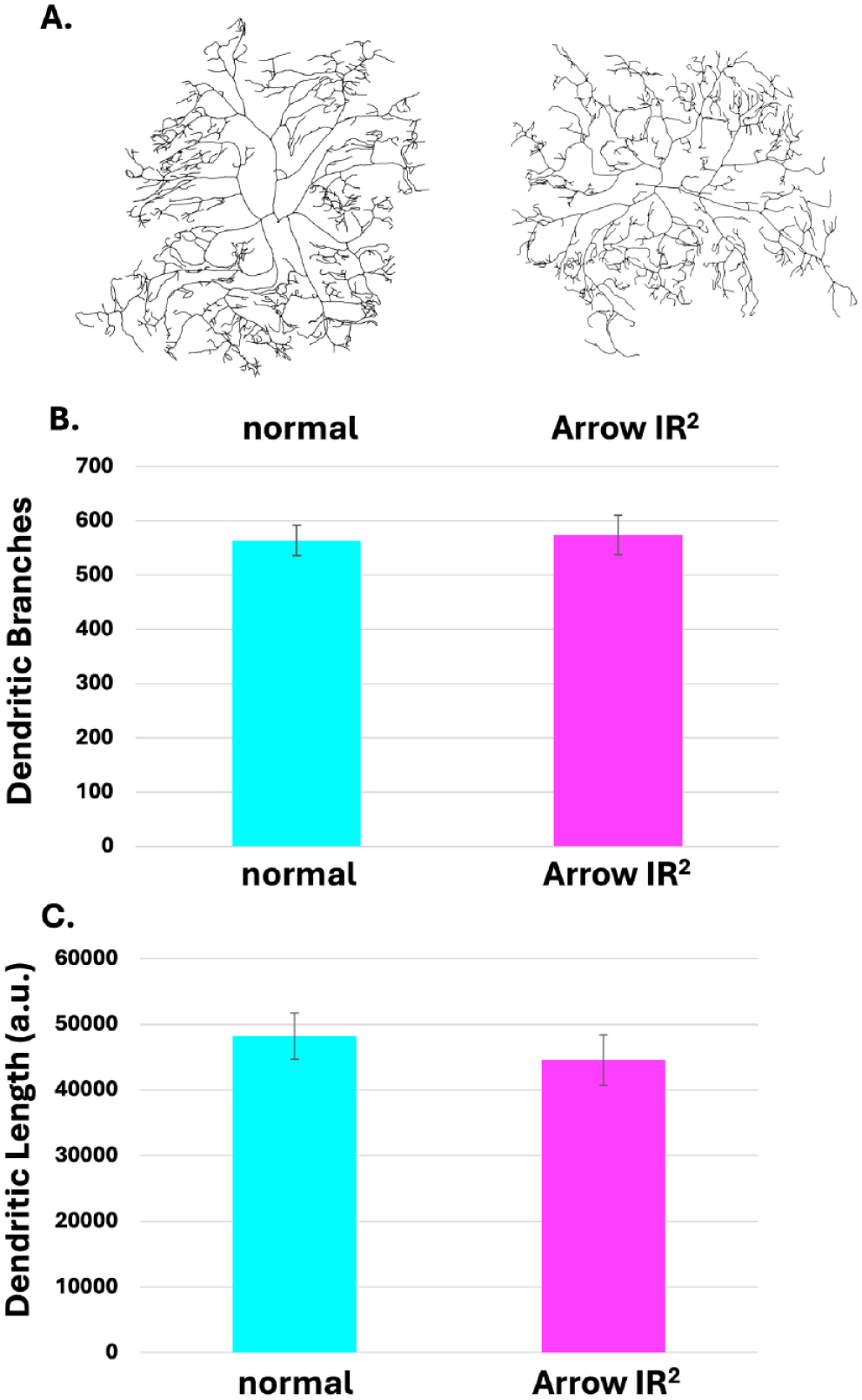
Arrow underexpression does not affect dendritic length or branching. Morphometric analysis shows no significant difference in the nociceptor dendritic arbor of animals in which Arrow was underexpressed, and those of normal controls. (a) Skeletonized trace (using ImageJ) of the dendrites of the normal control (ppk1.9-Gal4; UAS-mCD8-GFP > y1v1) (Left) and skeletonized trace of the dendrites of Arrow-underexpressing larva (ppk1.9-gal4; UAS-mCD8-GFP > UAS-Arr-IR2) (Right). (b) No significant difference in the dendritic branching is observed when comparing normal animals to the Arrow underexpressing animal (Student’s t-test with Welch correction, p = 0.155). (c) No significant difference in the dendritic length is observed when comparing normal animals (Left) to the Arrow-underexpressing animals (Right; Student’s t-test with Welch correction, p = 0.614). N = 8 for control, N = 8 for Arrow underexpresser.
Discussion
The Wg/WNT signaling pathway has been well studied for its role in development. Results presented in this study support the notion that the pathway affects the sensitivity of the Drosophila primary nociceptor. Arrow and Frizzled, acting as membrane-bound co-receptors for the extracellular Wg/WNT ligand, contributes to the inactivation of the destruction complex that reduces Armadillo levels.38–42 When the ligand is present, Arrow is activated and Armadillo levels increase, leading to the transcription of the Wg/WNT responsive genes. Evidence is presented that experimental underexpression of Arrow (Figure 1) leads to significant hyposensitivity to noxious thermal and mechanical stimulation (Figure 2). This supports the hypothesis that Arrow normally serves a pronociceptive function, presumably by increasing the expression of downstream Wg/WNT effector proteins that promote sensitivity. This result is consistent with prior studies showing that Armadillo underexpression also produces hyposensitivity. 18 If Arrow is underexpressed, it stands to reason that the destruction complex activity would be increased, in turn reducing Armadillo levels.
The Gal4/UAS-RNAi system is extensively used in Drosophila to reduce translation of target genes in a cell-specific manner. 23 In these experiments, the tools used to reduce Arrow expression in the Class IV dendritic arborization (C4da) neuron also reduced its expression outside this neuron (Figure 1). The Gal4 tool employed, ppk1.9-Gal4, is regarded as quite specific to the C4da cell, as seen in immunohistofluorescent (Figure 1) and neuromorphometry experiments (Figure 3). However, the presence of low but effective levels of Gal4 expression in other cells cannot be excluded, which could explain the reduction of Arrow expression via the UAS-driven inverted repeat element employed. The lack of nociceptor specificity of Arrow underexpression observed in the immunohistofluorescence experiments (Figure 1) could also be due to the UAS responder element, such that an effective amount of inverted repeat RNA is produced in non-C4da cells, even in the presumed absence of Gal4 transcriptional activation, leading to a reduction in Arrow levels in those cells as well. In any case, due to the observed lack of specificity of underexpression, it is not possible to determine which cell type requires Arrow to promote nociceptive sensitivity in this experiment. However, given the known function of the C4da neuron as a thermal and mechanical nociceptor,7,26 it seems logical that Arrow’s pronociceptive function is fulfilled in this cell.
The Gal4 tool used to select the nociceptor in this experiment, ppk1.9-Gal4, employs the promoter of Pickpocket, which encodes a mechanotransducing ion channel. 26 Pickpocket expression has been detected in the peripheral nervous system as early as embryonic stage 17, 25 several days before the age at which the larvae in these experiments were studied. At that time point, the C4da neuron may have already been specified. However, Wg/WNT activity is either not required, or manipulation of the Wg/WNT pathway in these experiments apparently does not occur early enough, to affect the development of the dendritic arbor of this cell (Figure 3). The results reported here suggest that whether or not Wg/WNT plays an organizational role in the development of this primary nociceptor’s dendritic arbor, it may affect nociceptive sensitivity in other ways and may thus serve an activational role once development is completed.
Overall, the results presented here support prior observations suggesting that the Wg/WNT signaling pathway plays a role, although apparently not through influence over dendritic morphogenesis, in determining the thermal and mechanical nociceptive sensitivity of the Drosophila larva.
Footnotes
Author contributions
TF, JKM, and GKG designed the research. TF, MC, TM and JKM performed the experiments and analyzed the results. TF, JKM, and GKG wrote the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIH award 1R15NS131952 to Geoffrey Ganter, and the Histology and Imaging Core, funded by P30GM145497 to Ian Meng. Stocks obtained from the Bloomington Drosophila Stock Center (NIH P40OD018537), including those generated by the TRiP at Harvard Medical School (NIH/NIGMS R01-GM084947), were used in this study. The ppk1.9-Gal4 and ppk1.9-Gal4, UAS-mCD8-GFP lines were a generous gift from M. Galko. Custom Python algorithms and scripts used in morphometric analysis were a generous gift from D. Cox.