
Abstract
Chronic pain is highly prevalent among active-duty service members (ADSMs) and contributes to substantial physical, psychological, and social burden. Genetic factors are recognized as central to pain susceptibility and treatment response. While few studies have examined the genes involved in cortisol synthesis, changes in the coding of cytochrome P450 (CYP) enzymes responsible for cortisol production have been implicated in chronic pain. We evaluated whether exonic variants in CYP11B1 and CYP17A1 are associated with salivary cortisol and pain-related outcomes in ADSMs with chronic pain using baseline genomic, salivary cortisol, and web survey data from 118 participants in a pragmatic clinical trial. We identified 40 variants across CYP11B1 and CYP17A1 exons and examined both common and rare variant burden scores. Linear regression models estimated associations with 11 outcomes, including pain intensity, sleep-related impairment, depression, and physical and social functioning. Mediation by cortisol was also assessed. Variants in both genes showed consistent associations with pain outcomes. Rare CYP11B1 variants (minor allele frequency <5%) demonstrated the strongest relationships with pain intensity, pain impact, depression, fatigue, and sleep-related impairment. CYP17A1 variants were strongly associated with sleep-related impairment, depression, fatigue, anxiety, and pain impact. Rare CYP11B1 variants were associated with salivary cortisol; however, mediation analyses indicated that cortisol accounted for only a modest proportion (4%–18%) of the gene–pain associations. These exploratory findings suggest that functional variation in cortisol-related CYP genes may contribute to pain-related outcomes among ADSMs with chronic pain, warranting replication in larger cohorts. The research protocol was registered in ClinicalTrials.gov (NCT03297905).
Introduction
Chronic pain is a major health concern among the adult population worldwide. Compared to the general population, military personnel are at heightened risk, with one-third of active-duty service members (ADSMs) diagnosed with chronic pain. 1 Chronic pain has substantial and varied consequences, 2 which impact work and physical function as well as mental health and social wellness.3,4
The multifactorial nature of chronic pain makes evaluating the effectiveness of therapeutic interventions challenging. Several risk factors have been described for chronic pain, including high physical exertion, age, sex, chronic medical conditions, and neurological issues. 5 More recently, studies examining the genetics of pain have revealed loci associated with chronic pain, 6 the pharmacogenetics of pain therapies, 7 and allelic differences linked to pain sensitivity. 8
Cytochrome P450 (CYP) genes are of particular interest in pain research due to their key roles in the metabolism of drugs and hormones. 9 Of particular note, CYP genes code for the enzymes that synthesize cortisol, known as a biomarker of stress and chronic pain. 10 Two CYP genes in particular—CYP11B1 and CYP17A1—are linked to cortisol production.11–14 Specifically, CYP17A1 is essential for the conversion of progesterone to the cortisol precursor, while CYP11B1 is essential to the final step of cortisol synthesis.
Evidence from previous studies suggests that chronic pain severity may be negatively associated with cortisol levels, 15 while other work has reported lower cortisol concentrations and a blunted diurnal slope among individuals with chronic pain. 16 Although the relationship between cortisol regulation and chronic pain remains inconclusive, the close interplay between physiological stress and pain responses provides a biologically plausible rationale for examining related genetic pathways. 17
To date, variants within CYP11B1 and CYP17A1 have been linked to hypertension, 18 cardiometabolic traits,19–21 and alterations in cortisol synthesis. Accordingly, those genetic variation in CYP17A1 and CYP11B1 may also underlie pain-related outcomes. However, there has been little investigation into direct associations between genes responsible for cortisol synthesis and pain outcomes. 22
We hypothesized that variants in genes essential to cortisol synthesis would be associated with measures of chronic pain. Our study examined whether exonic variants in CYP11B1 and CYP17A1 are associated with patient-reported pain outcomes, including pain, depression, anxiety, and other psychosomatic and physical functioning measures, in a sample of ADSMs with chronic pain. We also sought to identify whether exonic variants in CYP11B1 and CYP17A1 are associated with levels of salivary cortisol, and whether salivary cortisol mediates associations between genetic variants and pain outcomes.
Methods
Study design and setting
This was a secondary analysis using pre-intervention (baseline) data from a pragmatic clinical trial, 23 the details of which have been published previously. 24 The parent study was conducted at the Interdisciplinary Pain Management Center of the Madigan Army Medical Center in Tacoma, Washington. Participant enrollment occurred between May 2021 and October 2022.
Sample
Study participants were ADSMs referred to the Interdisciplinary Pain Management Center, the majority being members of the United States Army. Participants were aged 18–58 years, had pain that had lasted 3 or more months, and reported a 7-day average pain intensity of 3 or greater on an 11-point scale 25 or scored at least 1 standard deviation worse than the mean on the Patient-Reported Outcomes Measurement Information System (PROMIS) Pain Interference or Physical Function score.26,27 Individuals who had undergone or were planning to undergo surgery within the next 6 months and individuals with psychosis, at high risk of suicide, or meeting Diagnostic and Statistical Manual of Mental Disorders, fifth edition (DSM-5) criteria for severe substance use disorder were not eligible to participate in the study. A total of 254 participants provided informed consent for genomic and salivary data collection and completed web-based surveys from the Pain Assessment Screening Tool and Outcomes Registry (PASTOR).
Outcome measures
Pain-related outcomes
Each ADSM who participated in the study completed PASTOR assessments at baseline. For this analysis, we considered 11 separate self-reported pain outcome measures from PASTOR: the Defense and Veterans Pain Rating Scale, which assesses average pain intensity over the past 7 days 25 ; the Pain Catastrophizing Scale 28 ; eight PROMIS measures: physical function, 27 pain interference, 26 fatigue, 29 sleep-related impairment, 30 satisfaction with social roles and activities (social function), 31 depression, anxiety, and anger 32 ; and a pain impact score calculated by compositing pain intensity, pain interference, and physical functioning scores. 33 Each PROMIS domain is reported as a T-score, which is a standardized value calculated from participants’ responses. These T-scores are referenced to the U.S. general population, where the mean is set at 50 and the standard deviation at 10. Higher scores correspond to worse levels of impairment or distress for all measures except for physical functioning and social functioning, for which lower scores indicate worse functioning.
Salivary cortisol
Salivary cortisol samples were also collected at baseline. Samples were collected during weekdays in participants who work during the daytime. Participants were asked to provide three saliva samples prior to treatment for chronic pain: at awakening, 30 min after awakening, and at bedtime. This collection schedule enabled us to analyze both the cortisol awakening response and diurnal variability.34,35 Participants were asked to collect samples on the day prior to the first treatment day and to turn samples in the following day. Because treatment occurs during the Monday–Friday workweek, cortisol samples may have been collected on any day between Sunday and Thursday. Participants were encouraged to schedule treatment during periods when they were not working night shifts, but shiftwork was not an exclusion criterion. Participants were asked not to consume caffeinated or sugary drinks and/or food, nor engage in physical exercise, before taking their morning samples, to avoid the impacts of these activities on the cortisol awakening response. Participants were instructed to refrigerate samples immediately after collection and to submit the samples to the clinic on the first day of their treatment program. Saliva samples were collected using Salivette Cortisol collection tubes (SARSTEDT, Inc., Newton, NC, USA) following the manufacturer’s instructions. After sample collection, the collection tubes were centrifuged at 1000 relative centrifugal force for 2 min. The saliva samples were then aliquoted and stored at −80 °C for long-term storage. Salivary cortisol was measured using an enzyme-linked immunosorbent assay (ELISA) from R&D Systems (Minneapolis, MN, USA), following the manufacturer’s protocol. Cortisol measurements were used to calculate the area under the curve with respect to ground (AUCg) as previously described by Pruessner et al. 36
Exonic CYP variants
We collected baseline CYP samples via buccal swab collection and genomic DNA (gDNA) isolation, using the Qiagen Gentra Puregene Buccal Cell Kit (Germantown, MD, USA) per the manufacturer’s instructions. Participants were advised not to eat or drink for at least 1 h prior to scraping the insides of their cheeks with the buccal brush, 10 times on each side of the mouth. Buccal swabs were temporarily stored at 4 °C until gDNA was extracted and purified to remove proteins and RNAs. Purified gDNA samples were then stored at −20 °C until ready for exome sequencing. Exome sequencing was conducted by the Northwest Genomics Center at the University of Washington. Samples failed initial quality control if the total amount, concentration, or volume was too low, a 63-SNP OpenArray fingerprint assay derived from a custom exome produced poor genotype data or integrity of DNA, or sex-typing was inconsistent with the sample manifest. Base calls were generated on an Illumina NovaSeq6000 instrument (RTA 3.1.5) and a combined suite of Illumina and other industry standard software packages (i.e. Genome Analysis ToolKit (GATK), Picard, BWA, SAMTools, and in-house custom scripts) were used for alignment, local realignment, duplicate removal, quality recalibration, data merging, variant detection, genotyping, and annotation. Exome completion was defined as having >90% of the target at >10× coverage and >80% of the exome target at >20× coverage. Quality control included sample mean coverage of >10×, contamination of <3%, fingerprint concordance >99%, and ensuring that <7% of SNPs were novel variants relative to dbSNP. As an exploratory study using buccal swab collection, the QC standards used were less than current standards.
Of the 254 buccal swab samples provided, 202 passed the initial quality control for library preparation. Of these, quality libraries were generated and sequenced from 160 samples. After sequencing, samples from 118 individuals passed the initial sequencing quality control and were used for genetic analysis. Genomic Variant Call Format (gVCF) files were compressed using CombineGVCFs 37 and aligned to a reference FASTA file from HG38. Phred-scaled genotypes were used to infer allele frequencies at each varying site within CYP11B1 and CYP17A1. Genotypes were filtered to include exonic regions of the two genes. There were 40 variants within the two genes that varied across the sample and had inferred genotypes among at least 50 individuals in the sample (see Supplemental Table S1).
Table 1 provides basic information about the four common (minor allele frequency (MAF) >10%) exonic variants in the sample. All four variants (two in CYP11B1 and two in CYP17A1) had MAF >0.35, did not show evidence of being out of Hardy–Weinberg equilibrium (p > 0.05, based on chi-squared goodness-of-fit test using estimated MAF), and are annotated as synonymous coding variants. 38 In addition to these variants, data were available on an additional 25 variants in CYP11B1 with MAF <10%, 21 of which had MAF <5%. Similarly, an additional 11 variants in CYP17A1 had available genotypic data, all of which had MAF <5%. All single nucleotide polymorphisms (SNPs) were coded such that the major allele was coded as 0 and the minor allele was coded as 1, so that each individual received a score of 0, 1, or 2 for each variant, representing the number of minor alleles.
Commonly occurring exonic variants in CYP11B1 and CYP17A1.

MAF: minor allele frequency; SNP: single nucleotide polymorphism.
We created “burden” style measures of genetic variation separately within each gene by summing the total minor allele count within a gene, representing an additive disease risk model. We created three burden scores for CYP11B1 (MAF <5%: mean 0.4 (SD = 0.8); MAF <10%: mean 0.9 (SD = 1.3); all SNPs: mean 2.4 (SD = 1.6)) and two burden scores for CYP17A1 (MAF <5%: mean 0.3 (SD = 0.6); all SNPs: mean 1.8 (SD = 1.5)).
The UCSC Genome Browser 39 provides integrated information about GRCh38/hg38 including each genes start-stop positions, exon start–stop locations and data on which SNPs from dbSNP 38 have MAF >1%. These resources indicate a total of seven variants (one missense; six synonymous) with MAF >1% in the exonic regions of CYP11B1 and two variants (both synonymous) in the exonic regions of CYP17A1. All of these variants were observed in the sample with observed MAF close to global population allele frequencies (Table 1 and Supplemental Table S1b).
Statistical analysis
Participant characteristics at baseline were analyzed using descriptive statistics. Frequencies with percentages were computed for categorical data, and means with standard deviations were computed for continuous data. Separate linear regression models were fit, predicting baseline PASTOR measures by commonly occurring (MAF >10%) variants in CYP11B1 and, separately, CYP17A1 or by using quantitative measures of genetic rare variant “burden” (described above).
Additional models adjusted for participants’ baseline demographics (age, sex, race, ethnicity, smoking, income, education, and marriage/partnership). We followed a similar approach to predict log-transformed salivary cortisol levels. Finally, we added salivary cortisol to models predicting pain-related outcomes (PASTOR measures) and assessed the percentage change in genetic variant coefficients as a measure of salivary cortisol’s potential mediational impact on CYP genetic variants and pain. Since this was exploratory research we used 90% confidence intervals, considered p < 0.05 to be a statistically significant and p value’s between 0.05 and 0.10 identified as notable trends.
Results
Sample characteristics
A total of 118 participants were included in the study. Table 2 provides an overview of the participant characteristics. The sample was primarily male (79%), between the ages of 25 and 44 years (85%), and White (66%). More than two-thirds (70%) of the sample had attended at least some college, and nearly two-thirds (63%) had a spouse or partner.
Participant characteristics (N = 118).

AA: associate of arts; BA: bachelor of arts; BS: bachelor of science; GED: general equivalency diploma; MA: master of arts; MD: doctor of medicine; PhD: doctor of philosophy; PROMIS: Patient-Reported Outcomes Measurement Information System.
CYP variants and pain-related outcomes
We examined each of the 11 pain outcome measures to determine whether there was evidence of association with variants in the CYP genes. Five of the 11 outcomes showed statistically significant evidence of association (p < 0.05) in the adjusted models (Tables 3–7); while all 11 showed at least a notable trend (p < 0.10). Specifically, pain intensity and pain impact both showed associations with variants in the CYP11B1 gene, with the strongest evidence for the burden test including all SNPs with MAF <5%. Fatigue, depression, and sleep-related impairment also showed associations with variants in CYP11B1, particularly for the results of the burden test. The more common variants in CYP17A1 showed associations with pain impact, fatigue, depression, and sleep-related impairment. In a follow-up analysis, we added rs5283 to the pain intensity model with the MAF <5% burden score, and both rs5283 and the burden score remained statistically significant with similar coefficients. Results from other outcomes are shown in Supplemental Table S2.
Pain intensity (N = 118).
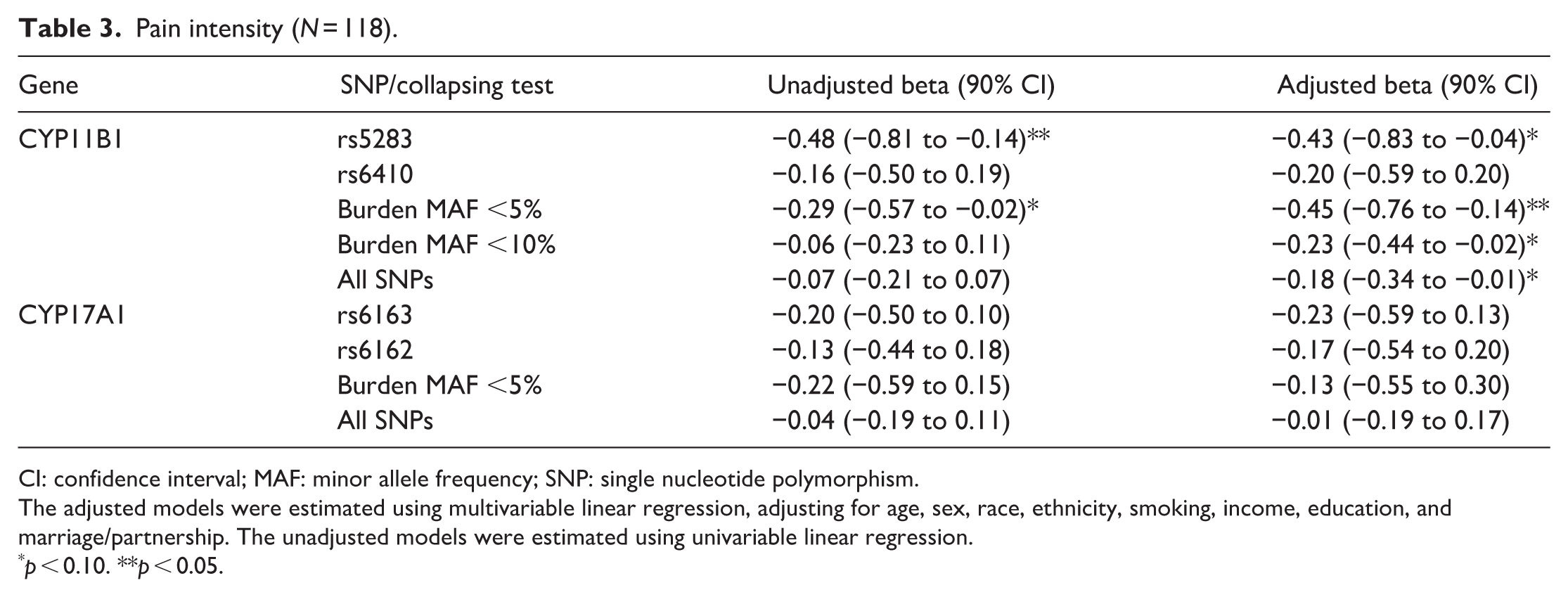
CI: confidence interval; MAF: minor allele frequency; SNP: single nucleotide polymorphism.
The adjusted models were estimated using multivariable linear regression, adjusting for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership. The unadjusted models were estimated using univariable linear regression.
p < 0.10. **p < 0.05.
Pain impact score (N = 118).

CI: confidence interval; MAF: minor allele frequency; SNP: single nucleotide polymorphism.
The adjusted models were estimated using multivariable linear regression, adjusting for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership. The unadjusted models were estimated using univariable linear regression.
p < 0.10. **p < 0.05.
PROMIS fatigue (N = 118).
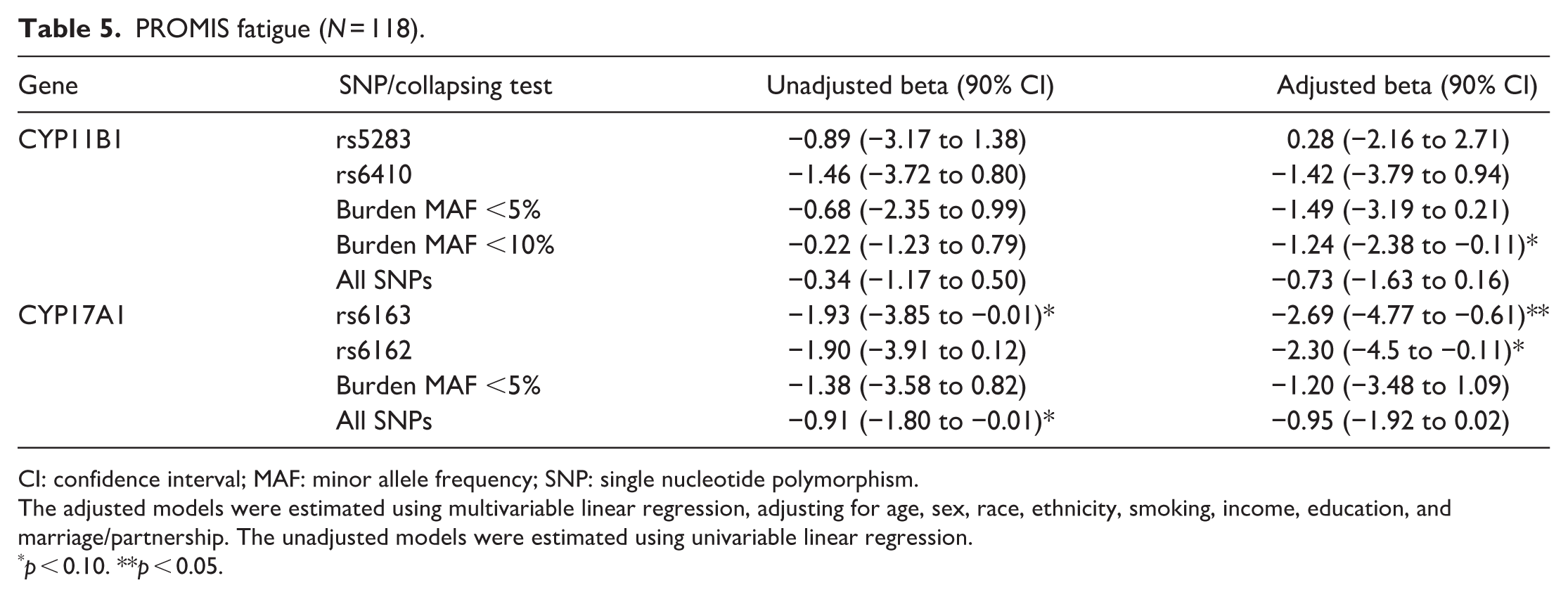
CI: confidence interval; MAF: minor allele frequency; SNP: single nucleotide polymorphism.
The adjusted models were estimated using multivariable linear regression, adjusting for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership. The unadjusted models were estimated using univariable linear regression.
p < 0.10. **p < 0.05.
PROMIS depression (N = 118).
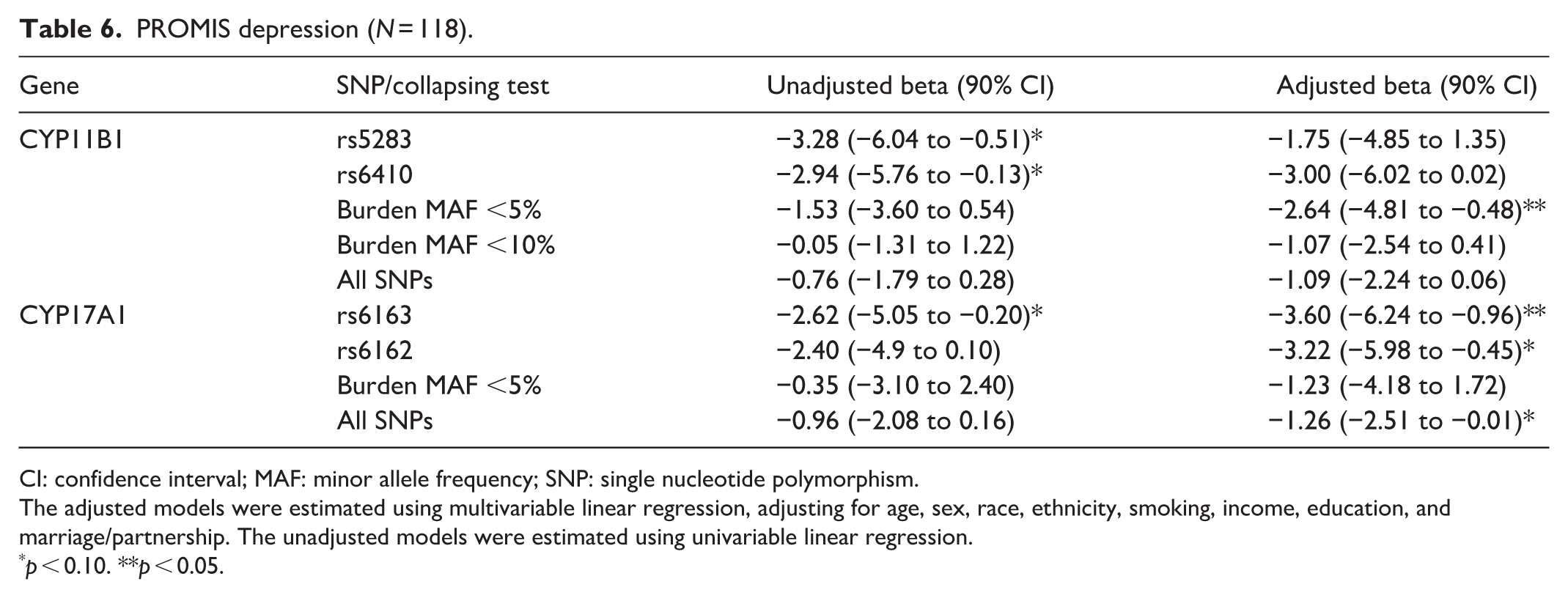
CI: confidence interval; MAF: minor allele frequency; SNP: single nucleotide polymorphism.
The adjusted models were estimated using multivariable linear regression, adjusting for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership. The unadjusted models were estimated using univariable linear regression.
p < 0.10. **p < 0.05.
PROMIS sleep-related impairment (N = 118).

CI: confidence interval; MAF: minor allele frequency; SNP: single nucleotide polymorphism.
Adjusted: multivariable linear regression adjusting for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership; unadjusted: univariable linear regression.
p < 0.10. **p < 0.05. ***p < 0.01.
CYP variants and salivary cortisol
Salivary cortisol levels showed a statistically significant (p < 0.05) association with less frequent variants in CYP11B1 in an adjusted model (Table 8), and a notable trend (p < 0.10) in the unadjusted model. Common variants in both CYP11B1 and CYP17A1 showed similar levels of association with salivary cortisol yet did not attain statistical significance.
Association of salivary cortisol levels and SNPs in two CYP genes (N = 118).

CI: confidence interval; CYP: cytochrome P450; MAF: minor allele frequency; SNP: single nucleotide polymorphism.
Adjusted: multivariable linear regression adjusting for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership; unadjusted: univariable linear regression.
p < 0.10. **p < 0.05.
Mediation by salivary cortisol
Table 9 gives the results of a mediation analysis, which evaluated the extent to which the associations between the CYP genetic variants and patient-reported pain outcomes were mediated by salivary cortisol levels. The focus of this analysis was on CYP11B1 rare variants (MAF <5%), which had shown the strongest overall evidence of association with pain outcomes. None of the statistically significant associations between CYP11B1 genetic variants and pain-related outcomes became nonsignificant after accounting for cortisol levels. Furthermore, the percent mediation estimates ranged from 4.5% to 18.1%, representing only modest mediation.
Mediation of CYP11B1 rare variant burden test relationships with pain outcomes, by salivary cortisol (N = 118).
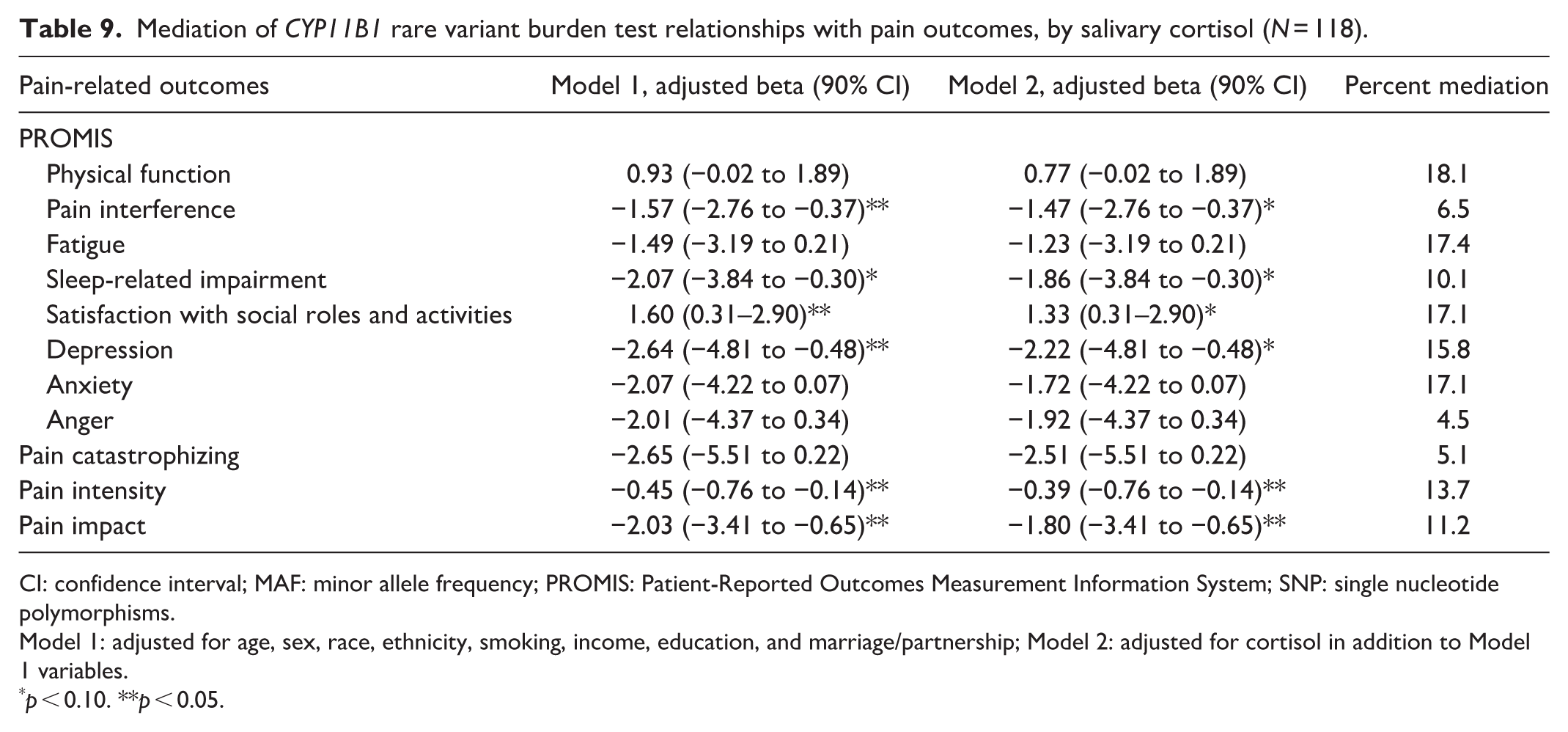
CI: confidence interval; MAF: minor allele frequency; PROMIS: Patient-Reported Outcomes Measurement Information System; SNP: single nucleotide polymorphisms.
Model 1: adjusted for age, sex, race, ethnicity, smoking, income, education, and marriage/partnership; Model 2: adjusted for cortisol in addition to Model 1 variables.
p < 0.10. **p < 0.05.
Discussion
We identified a clear and consistent signal between certain CYP genetic variants and a variety of patient-reported pain outcomes in our study sample of individuals with chronic pain. In our analysis, we found that SNPs in both CYP11B1 and CYP17A1 were associated with pain-related outcomes. Interestingly, these associations spanned all 11 patient-reported outcomes we measured, were found with both CYP genes we investigated, and included both more common and less common genetic variants.
CYP11B1 and pain-related outcomes
Of the 11 patient-reported outcomes we assessed, four directly relate to pain (pain intensity, impact, catastrophizing, and interference). Three of these showed a statistically significant association with at least one variant or for the burden score in CYP11B1, and the fourth showed a notable trend. In all cases, individuals with more variants in the gene had less pain intensity, impact, catastrophizing, or interference. In fact, nearly all SNPs and burden scores in CYP11B1 showed inverse associations with these four pain measures, even the SNPs and burden scores that did not attain statistical significance or show a notable trend. The two strongest associations were between the MAF <5% burden score and pain intensity and the MAF <5% burden score and pain impact. Given that the two common CYP11B1 variants (rs5283 and rs6410) had associations in a similar direction, albeit weaker, and that both variants are synonymous, it is plausible that any direct relationships between genetic variants in CYP11B1 are due to missense rare variants that are correlated with the common variants. However, our follow-up analysis, which modeled both the common rs5283 and the MAF <5% burden score simultaneously, yielded similar, independent, estimated effects. Bioinformatic analysis of the CYP loci with larger samples to better understand haplotype structure and recombination may provide more insights into possible mechanisms of action.
Variants in CYP11B1 also showed inverse associations with depression, fatigue, sleep-related impairment, and social functioning, with the strongest relationships found for sleep-related impairment. While several common variants were associated with sleep-related impairment, only the rare CYP11B1 variants showed measurable associations with salivary cortisol. This pattern may support the possibility that CYP11B1 is related to sleep via neuroendocrine pathways beyond cortisol concentration alone. 40 Sleep-related impairment is particularly sensitive to subtle alterations in signaling related to the hypothalamic–pituitary–adrenal axis and broader steroidogenic processes.41,42 This sensitivity may allow even modest genetic effects to be detected more easily, whether they arise from rare functional variants or from common variants that tag nearby regulatory loci. In addition, sleep-related impairment may be a more sensitive and reliably measured phenotype in the study population of ASDMs, which would potentially explain why the genetic associations appeared strongest for this domain. Previous studies have shown that methylation changes in the CYP11B1 promoter region correlate with overproduction of cortisol, 43 but evidence is lacking about direct relationships with pain. Therefore, these interpretations are speculative and require validation in larger studies designed to evaluate mechanistic pathways with deeper genomic coverage.
CYP17A1 and pain-related outcomes
Variants in CYP17A1 also showed evidence of associations with pain-related outcomes, which were in line with emerging findings about SNPs in CYP17A1 and pain in people with cancer.22,44,45 The strongest inverse relationships were observed with sleep-related impairment, followed by depression, fatigue, anxiety, and pain impact. In contrast to CYP11B1 which is specific to the end stages of cortisol and corticosterone synthesis, CYP17A1 is involved in the generation of neuroactive steroids such as progesterone and dehydroepiandrosterone (DHEA). 12 It is possible that our observed associations relate to these other steroids, in particular DHEA which regulate sleep and stress responses by modulating the activity of gamma-aminobutyric-acid-type-A (GABA(A)) receptors46,47; a class of ionotropic receptors involved in pain sensation. 48 Understanding these associations is important because it strengthens emerging evidence that genetic differences in CYP17A1 may influence how people experience multiple pain-related symptoms. Pain symptoms often cluster, yet their biological underpinnings remain poorly understood. Identifying a shared genetic contributor like CYP17A1 could help clarify potential mechanistic pathways, support the development of more targeted symptom management strategies, and move the field closer to approaches that tailor interventions to an individual’s genetic profile.
CYP variants and salivary cortisol
While we expected to find strong and consistent associations between CYP11B1 and CYP17A1 genetic variants and salivary cortisol, our results were limited to an inverse association between salivary cortisol and the rarer variants in CYP11B1 (MAF <5% burden score). This finding aligns with the dearth of prior research about genetic variants in these two CYP genes and their associations with cortisol levels in the body. 49 It is possible that compensatory mechanisms offset the impact of genetic variations on cortisol production. For example, biochemical responses may increase cortisol synthesis and/or inhibit the reduction of cortisol to its inactive metabolites. However, the inverse association between rarer variants in CYP11B1 and salivary cortisol suggests that some less abundant variants in CYP11B1 may play a larger role in directly impacting cortisol levels.
Epigenetic mechanisms
Epigenetic mechanisms may explain how SNPs, even if synonymous, may regulate cortisol levels. For example, Maharaj et al. observed that synonymous SNPs alter mRNA splicing leading to a nonfunctional CYP11A1 protein. 50 Although the impact of synonymous SNPs on CYP11B1 transcription remains unknown, synonymous polymorphisms in CYP17A1 are associated with differences in hormone levels in vivo and shown to disrupt genes transcription in vitro. 18 Given the potential impact on CYP transcription, synonymous variants have been associated with adverse health, such as coronary heart disease and skeletal growth disorders.20,51 However, in many of these studies, it remains unclear whether epigenetic mechanisms increase or decrease cortisol levels and if systemic changes in cortisol promote harmful or protective signaling responses.
Immune and inflammatory response
The inverse relationship with pain and cortisol may be explained by related immune and inflammation responses. Cortisol is a potent immunomodulator reported to instigate both pro- and anti-inflammatory responses. 52 It is possible that in subjects with chronic pain, higher levels of cortisol levels reflect anti-inflammatory responses. This possibility is supported by the anti-inflammatory and immunosuppressive effects of synthetic cortisol variants such as dexamethasone, that are commonly used for pain management. 53 Given the central role of inflammation in chronic pain, depression and poor sleep, immune response to cortisol may relate to our findings. Future analysis of inflammatory cytokines in conjunction with cortisol are needed to clarify this potential relationship.
Mediation role of salivary cortisol
This study’s hypothesis was based on the reasoning that cortisol would underlie the mechanisms by which associations between genetic variants and pain-related outcomes operate. In other words, that genetic variation in CYP genes would impact cortisol levels, which would in turn impact patient-reported pain measures. However, in this study’s sample of ADSMs, estimates of mediation were low (<15%–20%), indicating otherwise. A separate manuscript reporting data from this sample has also reported little evidence of associations between salivary cortisol levels and PASTOR measures, further limiting the potential mediational explanation. 54 Alternative explanations, which are not possible to explore in this dataset, include that SNPs in CYP genes influence pain through mechanisms independent of systemic cortisol (e.g. processing other steroid precursors), or that cortisol does mediate pain but due to local cortisol actions or impacts of genetic variation on cortisol function, but not on cortisol levels.
Strengths and limitations
Our study offers a novel and necessary look at potential genetic mechanisms of pain in a sample of individuals largely free of confounding health complications such as hypertension and cardiovascular disease and that has a comprehensive collection of paired salivary cortisol and genetic analyses. Our experimental design and analysis of salivary cortisol provide several advantages by normalizing diurnal fluctuations that can impact cortisol concentrations. Another major advantage of analyzing salivary cortisol is it reflects free (unbound) cortisol levels. 55 Within the blood the majority of cortisol is bound to albumin or cortisol binding protein (CBP) and unavailable for signaling. In contrast, salivary cortisol levels reflect physiological concentrations that are available for signaling. Nevertheless, heterogeneity in sample days, times or participant work/sleep patterns could contribute to variation in cortisol levels. This study has several important limitations. First, the modest sample size (N = 118) limits statistical power, particularly for detecting small effect sizes and rare variant associations. Second, we interpreted p < 0.10 as notable trends, and did not adjust for multiple testing, given the exploratory nature of the analyses and the limited sample size. While this approach may enhance sensitivity to potential associations, it also increases the risk of type I error. Third, the study population was consisted of ADSMs with relatively homogeneous demographics, predominantly male and White. This demographic homogeneity limits the external validity of the findings. Consequently, the generalizability of the finding to women, diverse racial/ethnic groups, or civilian populations remains uncertain. Accordingly, the reported results should be interpreted as largely preliminary rather than statistically supported associations. Replication in a demographically diverse population with larger sample sizes is necessary to evaluate the robustness and reproducibility of these findings.
Conclusions
Our exploratory findings suggest that functional variation in cortisol-related CYP genes may be associated with pain-related and psychosocial outcomes among ADSMs with chronic pain. Future work should explore additional mechanistic pathways by which the putative associations we have identified may exist (e.g. chronic conditions, post-transcriptional CYP protein regulation).
Supplemental Material
sj-docx-1-mpx-10.1177_17448069261454823 – Supplemental material for CYP gene variants show preliminary evidence of association with patient-reported outcomes in active-duty service members with chronic pain
Supplemental material, sj-docx-1-mpx-10.1177_17448069261454823 for CYP gene variants show preliminary evidence of association with patient-reported outcomes in active-duty service members with chronic pain by Nathan L Tintle, Nathan A Ryder, Jason Westra, Dahee Wi, Sotaro Shimada, Nicholas Ieronimakis, Honor M McQuinn, Tyler J Snow, Ardith Z Doorenbos and Diane M Flynn in Molecular Pain
Footnotes
Acknowledgements
We acknowledge the editorial support of Kyra Freestar and Rica Asuncion-Reed, who contributed to making the manuscript clearer.
Consent to participate
All participants in this study provided signed consent for their participation in the study.
Consent for publication
Not applicable.
Data availability statement
Any request for original data sets will be honored upon approval of a data-sharing agreement.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Disclaimer
The views expressed in this research article are those of the author and do not necessarily reflect the official policy or position of the Defense Health Agency, Department of War, nor the U.S. Government. The study protocol was approved by the Madigan Army Medical Center Institutional Review Board in compliance with all applicable Federal regulations governing the protection of human subjects.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The U.S. Army Medical Research Acquisition Activity (820 Chandler Street, Fort Detrick, MD 21702-5014) is the awarding and administering acquisition office. This work was supported by the Assistant Secretary of War for Health Affairs and Department of War through the Neuromusculoskeletal Injuries Rehabilitation Research Award (award no. W81XWH-18-2-0023) and National Institutes of Health/National Institute of Neurological Disorders and Stroke (award no. K24 AT011995).
Ethical considerations
The research protocol of the present study was approved by the Madigan Army Medical Center’s ethical review committee (approval number 221011) and registered in ClinicalTrials.gov (NCT03297905) prospectively.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.