
Abstract
The parabrachial (PB) nucleus participates in neural coding for diverse sensory modalities, including nociception and taste. Recent neurophysiological studies identified a subpopulation of PB neurons that responds to both gustatory and oral nociceptive (trigeminal) stimuli. These taste-integrative neurons populate PB subnuclei that receive somatosensory input from bodily skin supplied by spinal afferents. Yet whether spinal sensory input can excite these cells was unexplored. Here, we applied electrophysiological and optogenetic techniques in anesthetized TRPV1; Ai32 mice, which support transdermal photoexcitation of cutaneous TRPV1-lineage thermo-nociceptive fibers, to study if spinal sensory stimulation could engage taste-active PB cells. Action potentials were monitored in isolated PB neurons during oral application of diverse taste and chemesthetic stimuli, oral thermal stimulation using temperature-controlled water, and pinch of the hindpaw and tail. In some cases, photostimulation was applied to pinch-sensitive receptive fields to optogenetically excite bodily TRPV1-lineage fibers. Analysis of 94 PB neurons identified 35 cells (37.2%) that responded to more than one stimulus modality, including 10 taste-excited neurons that also fired impulses to noxious oral chemesthetic (trigeminal) stimuli, oral thermal input, and hindpaw/tail pinch. Such neurons could also respond to photoexcitation of TRPV1-lineage afferents innervating body extremities and frequently displayed bitter-oriented (aversive) taste response profiles. These data suggest that spinal afferent input, including pain-related neural messages from skin, converges onto taste-active PB neurons. These neurons would be misclassified as unimodal “taste” cells under traditional methods but display multisensory response repertoires that agree with involvement of PB circuits in sensory-affective processing.
Plain language summary
The parabrachial (PB) nucleus is a brain region that receives different types of sensory information, including taste, touch, and internal body signals. Recent data show that some PB neurons that respond to taste also react to oral thermal and pain-related sensations and reside in PB locations where sensory messages from the body are received. However, whether these taste-orosensory neurons also respond to signals from other parts of the body, like pain from the limbs, was unaddressed.
To study this, we monitored electrical activity from PB neurons in anesthetized mice and tested whether pain-related signals from the body (spinal input) combine with taste and oral touch signals in these cells. We recorded neuronal activity while applying taste solutions, irritant chemicals, and hot water to the mouth, and also during pinch of the hindpaw and tail. We also used an optical-genetic technology to briefly activate pain-sensing nerve fibers in the paw to see if this stimulated PB taste cells.
We recorded responses from over 90 PB neurons and found that a little more than a third responded to more than one type of stimulus modality. Of this set, a subgroup of taste-excited neurons was found that reacted to bitter tasting chemicals and oral presence of hot water, and also to pain-related stimulation of the paw or tail.
These results show that PB neurons can integrate orosensory taste and touch with pain-related signals carried by spinal afferents that supply bodily skin. These neurons could be misclassified as “taste cells” but appear to process multiple types of sensory input. Such neurons are poised to participate in the known integrative function of the PB nucleus and may combine features of different sensations, including taste and pain, in multisensory contexts.
Introduction
Trigeminal circuits supplying craniofacial somatosensation and nociception project, in part, to the lateral parabrachial (PB) nucleus.1–5 The lateral PB nucleus is evidenced to contribute to hedonic, or affective, dimensions of behavioral states associated with diverse sensory experiences, including pain.4–7 Accordingly, lateral PB neurons can respond to nociceptive stimuli that strongly excite trigeminal afferents, such as lingual presence of noxious hot water or capsaicin.8,9 Oral sensitivity to capsaicin is a hallmark of activation of trigeminal thermo-nociceptive fibers expressing the transient receptor potential (TRP) vanilloid 1 (TRPV1) ion channel, which is associated with pain-related and heat transduction.10,11
Recent neurophysiological studies in mice have found that oral presence of capsaicin or noxious heat can excite a subpopulation of lateral PB neurons that are co-responsive to gustatory stimuli. Such neurons frequently responded to bitter tastants like quinine,8,9 which is innately avoided by mice as concentration rises in taste behavioral assays.12,13 Quinine is transduced by taste bud cells supplied by non-trigeminal nerve fibers.14,15 Accordingly, lingual presence of quinine does not excite rodent trigeminal neurons in vivo, 16 including under temperature-controlled stimulus conditions. 17 Thus, the dual sensitivity to aversive bitter taste and oral pain-related stimuli observed in PB neurons was interpreted to reflect cross-sensory convergence of upstream trigeminal and gustatory input onto these cells. This interpretation was buttressed by neural recording and optogenetic analyses that revealed evidence for neural circuit mechanisms that could drive convergence of taste and trigeminal signals in lateral PB regions.8,9
Classic 5 and modern 18 functional studies in rodents have identified that PB neurons engaged by noxious craniofacial stimulation frequently co-respond to mechano- and thermo-nociceptive stimuli applied to skin within spinal dorsal root dermatome fields. For example, pinch/squeeze and heat (e.g. 50°C) stimulation of the tongue (orosensory) and hindpaw/tail (spinal) can excite impulses in rat PB neurons, with such cells frequently located in the external lateral PB subnucleus. 5 When combined with the evidence for a confluence of trigeminal nociception with taste in lateral PB circuits,8,9 these results could imply that noxious spinal inputs may also engage taste-excited PB cells. This hypothesis may have implications for understanding the neural coding of bodily pain-related messages by multisensory central neurons.
Here, we used in vivo electrophysiological recordings and Cre-directed optogenetics in mice to examine whether stimulation of spinal afferents supplying the skin could excite taste-active neurons in lateral PB circuits. Spinal input was induced by pain-related stimuli such as pinch and transdermal photoexcitation of TRPV1-lineage fibers innervating bodily extremities like the hindpaw. TRPV1-lineage afferents are implicated in temperature sensing and pain-related neural transmission.9,19–21 PB neurons were also tested for responsiveness to oral thermal and chemesthetic stimuli associated with trigeminal nociception.
We found that while over half of the recorded gustatory-excited neurons were selective for only taste, a subgroup emerged that co-responded to oral somatosensory stimuli and stimulation of spinal sensory-supplied skin. Such “gustatory” neurons appeared to convey signals relevant to protective coding across different bodily systems, implying they support functions beyond unimodal taste.
Materials and methods
Animals
These studies involved acute neurophysiological recordings of PB neurons in anesthetized adult (mean age: 16 weeks; 95% CI: 13.2–18.7 weeks) male (n = 41; bodyweight: 26.3 (mean) ± 2.1 (SD) g) and female (n = 25; bodyweight: 22.1 ± 3.0 g) TRPV1;Ai32 mice. These mice were the F1 progeny of crossing male TRPV1-Cre mice (JAX #017769) with female Ai32 mice (JAX #024109). The latter enabled Cre-dependent expression of a channelrhodopsin-2 (ChR2)/EYFP fusion protein. TRPV1;Ai32 mice supported ChR2-assisted photoexcitation of TRPV1-lineage somatosensory fibers involved with thermal sensation and nociceptive responses.
All procedures performed on mice were approved by the University of Oklahoma IACUC and followed National Institutes of Health guidelines. Mice in this study were naive to experiments and were housed in a vivarium that maintained a 12/12 h light/dark cycle and an air temperature of ∼20°C. Food and water were available ad libitum.
Surgery
Surgical preparation for acute PB neural recordings followed our described methods.8,9 Briefly, mice were inducted by an intraperitoneal (i.p.) injection of a ketamine (10 mg/mL) and xylazine (1 mg/mL) mixture (10 mL/kg). Atropine (24 µg/kg, i.p.) was administered to minimize bronchial secretions to keep the airway clear under long-term anesthesia during the recording period. Once anesthetized, fur on the dorsal skull and ventral neck was trimmed using electric clippers. Next, mice were placed in a supine position, and a tracheostomy procedure was performed to insert a small tube into the trachea. This tube enabled unrestricted breathing when applying liquid stimulation to the whole mouth and supported maintenance of isoflurane gas anesthesia during extended recording sessions. Mice were then secured in a stereotaxic instrument with ear bars (model 930; David Kopf Instruments). The distal end of the trachea tube was placed inside the inner part of custom concentric pressure/vacuum tubing, which supplied mice with isoflurane (1.2%–1.5%, in O2) and scavenged exhaled gas. Mice respired on their own without assisted ventilation.
Once surgical-level anesthesia was achieved, a craniotomy was made at the skull location of the PB recording site. A small silk thread was passed behind the lower incisors and drawn lightly taut to deflect the mandible downward. The tongue was then extended from the mouth by a small rostroventral suture clasped with a small, lightweight bulldog clamp. Using miniature rongeurs, the lower incisors were trimmed so that they did not deflect or damage the protruding tongue. Heart rate and blood oxygen level were monitored by a pulse oximeter (MouseSTAT Jr., Kent Scientific). Body temperature was maintained at 36.5–37°C with a feedback-controlled heating pad. During recording sessions, anesthesia was monitored and confirmed by the absence of a reflex to heavy pinch applied to a hindpaw. 8
Neurophysiology
The lateral PB region where a confluence of taste and trigeminal somatosensory pathways arises was targeted using coordinates: 4.7–5.1 mm caudal of bregma, 1.1–1.4 mm lateral, and 2.2–3.0 mm below the brain surface. A tungsten micro-electrode (2–5 MΩ, FHC Inc.) was advanced towards the PB area using an electronic micro-positioner (Model 2660, David Kopf Instruments) at an angle of 2–4° in the rostral direction relative to the vertical axis. This approach helped avoid penetration of the transverse sinus while allowing the electrode tip to accurately target the PB area. Electrophysiological activity was monitored along the dorsoventral electrode tract. While approaching the expected depth of the PB region, we sought out taste-active neurons by gauging sensitivity to oral delivery of a room temperature aqueous solution of ~300 mM NaCl, delivered as below. This stimulus excites diverse groups of PB taste-excited neurons in mice, including cells responsive to electrolytic, aversive bitter, or appetitive taste stimuli. 8 Room-temperature NaCl also served as a cooling stimulus because the oral cavity was adapted to 35°C water, as below, which facilitated identification of thermal-sensitive PB neurons. When a well-isolated unit was encountered that did not respond to the NaCl search stimulus, additional tests were performed using cold water (<10°C) or pinching the hindpaw/tail to assess somatosensory responsiveness.
Electrophysiological recording sessions were usually started around 12:00, and the responses of most cells analyzed here were record in the afternoon between 14:00 to 18:00. Data were collected from a few neurons in the evening (e.g. ~22:00), in the animal’s dark (normally active) cycle. Electrophysiological responses by individual neurons were captured in early recordings using a P511 AC amplifier with high-impedance probe (Grass Technologies) and a band-pass filter (∼0.3–10 kHz). For later recordings, a Plexon OmniPlex system with a digital headstage processor was used to amplify neural responses. A template-matching algorithm sampled extracellular action potentials from well-isolated single neurons at 25 kHz (1401 interface and Spike2 software version 9, CED). Action potentials were timestamped to 0.1 ms. Data files were stored and analyzed offline after recording sessions had completed.
Oral chemical, thermal, and spinal somatosensory stimuli
Oral chemical (taste, chemesthetic) and fluid-based thermal stimuli were presented to the mouth over three separate blocks while acquiring action potentials generated by single PB neurons. Within each block, stimulus trials were randomly ordered, without replacement, for each neuron. Chemical/thermal solutions were delivered to the oral cavity using a custom computer-controlled apparatus that could provide continuous flow of a temperature-controlled water adaptation rinse and rapidly switch to a stimulus solution presented at a different temperature (Figure 2(a)), as described.8,9 This system also continuously sampled (1 kHz) fluid temperature at the moment of stimulus entry into the mouse’s mouth. Stimulus temperatures stated herein reflect oral °C based on average measurements during the plateau of the thermal ramp across trials. All fluid stimuli were preceded and followed by oral delivery of a 35°C adaptation water rinse.
Taste stimuli were aqueous (purified water) solutions of 500 mM sucrose (sweet to humans), 100 mM NaCl (salty), 10 mM citric acid (sour), 10 mM quinine-HCl (bitter), and 0.1 mM cycloheximide (bitter). Taste solutions were presented at 28°C for 5 s, with concentrations following our prior studies on taste-active PB cells.8,9 In gustatory behavioral assays, C57BL/6J mice, which are a background strain for TRPV1-Cre mice (JAX), show unconditioned preference to lick solutions of sucrose and avoid licking quinine and cycloheximide solutions compared to water in concentration-dependent manners.12,22,23 C57BL/6J mice also avoid licking 10 mM citric acid compared to water in these tests. 24
Thermal stimulation was accomplished using a flow of purified water delivered to the oral cavity at temperatures (°C) of <10°, 14°, 21°, 28°, 35°, 42°, 48°, or 52° for 5 s. These values served as the intended target oral temperatures during the thermal stimulation trials. Actual temperatures varied slightly across trials but remained close to the specified targets. The highest temperatures exceed noxious heat threshold and activate nociceptors. 25 Oral chemesthetic stimuli (Sigma-Aldrich) were aqueous solutions of 1.28 mM L-menthol and 1 mM allyl isothiocyanate (AITC; mustard oil) delivered at 35°C and presented for 20 s. One mM AITC engages the nocisensors TRP ankyrin 1 (TRPA1) and TRPV1 and causes aversion in mouse lickometry and fluid consumption tests.26,27
Spinal somatosensory stimuli included brief squeeze or pinch of the hindpaw or tail. Squeeze/pinch was achieved using standard and calibrated (20 g < force < 1000 g; Rodent Pincher algometer, Bioseb, Pinellas Park; Figure 2(a)) hand-held forceps. A keystroke or TTL pulse (for calibrated forceps) detected by the data acquisition system marked each pinch in electrophysiological records. Pinches were applied to the ipsi- or contralateral (relative to the PB recording) hind paw, but pinches to the contralateral hindpaw had a higher chance of activating PB cells. In some cases, PB neurons were also tested for sensitivity to transdermal photostimulation of TRPV1-lineage fibers supplying glabrous skin within pinch-sensitive receptive fields on the plantar surface of the hindpaw, or the tail. Here, blue light was presented in pulses (0.1–0.3 s width) using a 200 μm diameter fiber optic cable (0.39 numerical aperture; FT200EMT, ThorLabs) coupled to a 473 nm DPSS laser (OEM Laser Systems). The intensity of the blue light at fiber optic tip was 159 mW/mm² (measured using a PM100D meter with S130C photodiode power sensor, ThorLabs). The laser was controlled by programmable TTL coding from the data acquisition system. The stimulating end of the fiber optic cable was secured using a micromanipulator to reside just above the skin surface for effective light delivery.
Histological confirmation of recording sites
Electrode placement was assessed for most mice using histology. To do this, the electrode was slowly withdrawn from the brain following the last recording session of the day. The electrode tip was rinsed and then painted with 2 μL of the tissue label CM-DiI (dissolved in ethanol, 5 μg/50 μL) via pipette while viewed under a microscope. The electrode was then inserted back into the PB area under micro-positioner control using the recording coordinates and held in place for 10 min. 28 Mice were subsequently overdosed with sodium pentobarbital (270 mg/kg, i.p.) and underwent a transcardial perfusion procedure. Brains were removed and stored in a 4% paraformaldehyde/20% sucrose solution. A microtome was used to cut brains into coronal sections (40 μm), which were subsequently mounted onto slides. Roughly half of the brain sections considered were Nissl stained for histological analysis 29 while a remainder were used for fluorescence visualization of electrode placement, as marked by a yellow fluorescence tract. Not all cellular recording sites were recoverable.
Data analysis
Electrophysiological data were analyzed using custom MATLAB (The MathWorks Inc., Natick, MA) code combined with the Spike2 MATLAB SON interface (CED), which supports analysis of Spike2 data files in MATLAB. Taste and oral thermal responses by individual PB neurons were quantified by counting action potentials generated during the 5 s stimulus period. This response was corrected for baseline firing by subtracting the action potential count during the 5 s period that preceded stimulus onset. Quantified responses were expressed in Hz (spikes per second). Responses to taste or oral thermal stimulation were considered significant if the corrected responses exceeded 2 standard deviations of baseline firing rates. The gustatory tuning orientation of neurons was defined by the taste stimulus that elicited the most effective (highest) response.
PB cells were defined as oral somatosensory (trigeminal) responsive if they showed significant activity to menthol, AITC, or noxious hot water (≥48°C). 8 Responses to menthol and AITC were considered significant if spike activity in more than three consecutive 1-s bins captured after stimulus onset (for up to 115 s) exceeded the 95% confidence interval of firing in baseline period bins. Longer response windows were considered for oral chemesthetic stimuli due to their known lingering effects on PB cells.8,9 Plots of mean firing frequency were derived using standard routines offered by Spike2.
Spiking responses in PB neurons to photo and mechanical stimulation of spinal dermatomes (paw, tail) were identified by observing change in action potential frequencies that arose with, and followed, stimulus delivery over multiple trials. Because of response magnitude variance, challenges in mapping skin receptive fields, and stimulus history effects on such activity, PB neurons were classified as only receiving, or not receiving, spinal input based on the appearance of reliable change in spike responding.
Pinch responses in neurons identified to respond to both taste and spinal input were analyzed using statistical criteria. For pinch trials conducted using calibrated forceps and marked by TTL pulses, action potentials recorded during the 5 s period preceding the onset of pinch trials were defined as baseline activity and quantified in 1 s bins. For pinch trials performed using standard forceps, keystrokes marking pinch events were not precisely time-locked; therefore, pinch response windows were defined as the intervals between consecutive keystrokes, and firing rates were calculated within these periods. Baseline activity for these trials was estimated from periods without pinching and expressed as firing rates (spikes/s) across five bins. Significant neuronal responses to pinching were defined as firing rates during the response window that exceeded (excitation) or fell below (inhibition) the 95% confidence interval of baseline activity. At least five pinch trials were analyzed for each neuron.
Results
We recorded 94 PB neurons in total. All cells were tested for sensitivity to taste, oral somatosensory (trigeminal) stimulation, and for responsiveness to somatosensory stimuli delivered to hindpaw/tail skin. Assessment of histology identified that electrode placement targeted the PB area, with neurons frequently sampled from lateral PB regions (Figure 1).
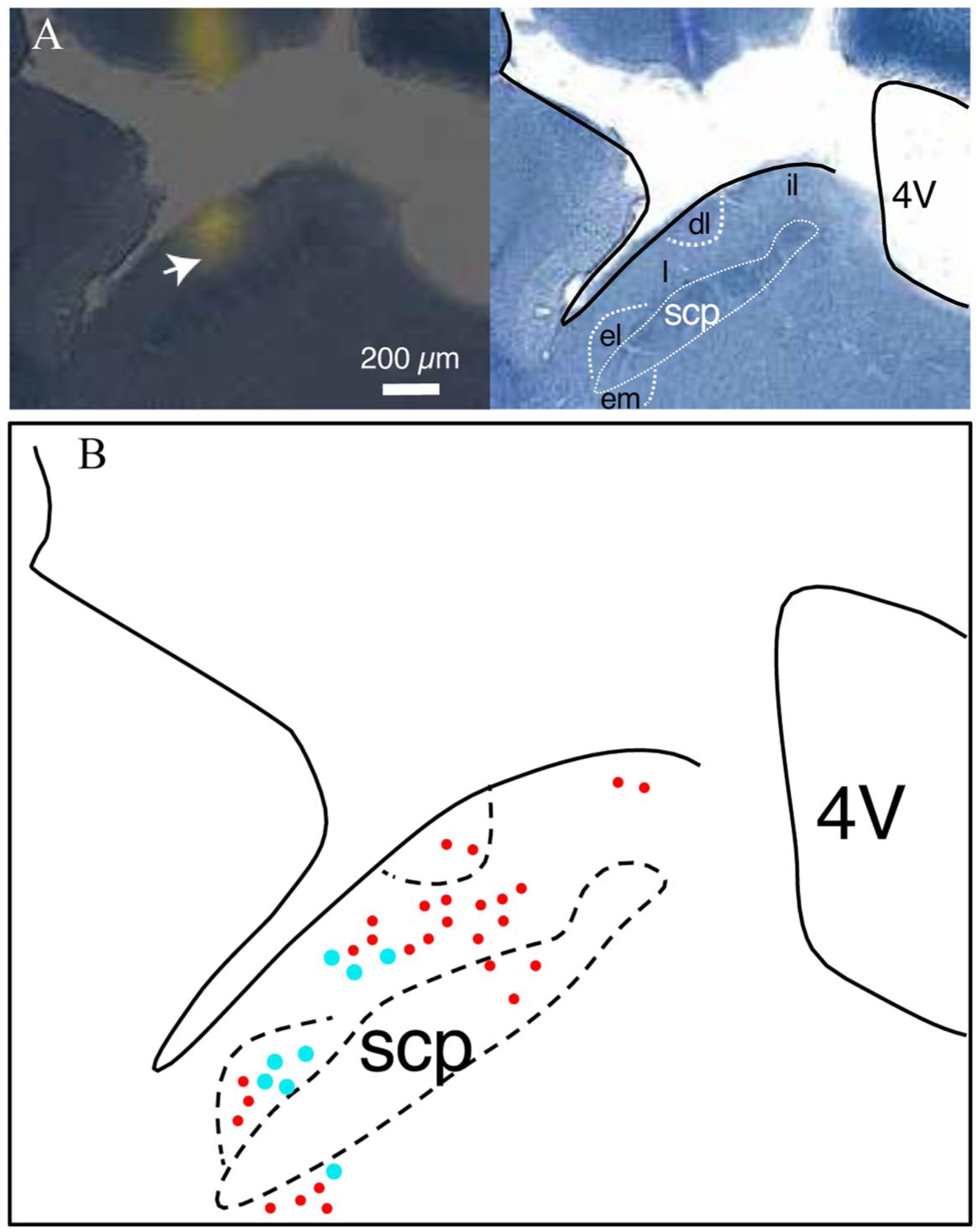
Histological analysis of recording sites in the parabrachial nucleus (PB): (a) Left photomicrograph shows a coronal section of brain tissue from one mouse depicting placement of the electrode in the lateral PB area (arrowhead, yellow fluorescence marks electrode shank/tip). Right photomicrograph shows thionine staining of an adjacent coronal section. Solid black lines demarcate the dorsal border of the pons and ventricle. Dashed white lines highlight PB structures. (b) Reconstruction of recording electrode locations in the PB area (solid circles, n = 35). Each plotted circle marks the estimated placement of an electrode tip, as in panel (a). The larger cyan circles represent taste-spinal cells (n = 8). Reconstructed recording sites are plotted on only one coronal plane of the PB for simplicity; actual marked sites extended approximately 60 µm along the rostrocaudal dimension.
Most neurons (n = 59, 62.8% of the sample) showed a change in action potential discharge rate to stimuli within a single sensory modality (frequencies of unimodal neurons: taste responsive, n = 31 (33.0%); oral somatosensory responsive, n = 12 (12.8%); spinal somatosensory responsive, n = 16 (17.0%); Figure 3(a)). Among neurons that responded to only taste, six (19.4%) exhibited quinine/cycloheximide (bitter)-dominant taste response profiles (mean firing rate to quinine = 6.7 Hz), nine (29.0%) showed NaCl-dominant taste profiles (mean response to NaCl = 9.0 Hz), eight (26.0%) responded most strongly to sucrose (mean response = 16.0 Hz), and eight (25.8%) were broadly tuned to electrolyte (NaCl, citric acid) stimuli.
On the other hand, 35 PB neurons (37.2%) responded to stimuli across more than one modality. Among these cells, 10 (10.6%) were responsive to taste, oral somatosensory stimuli, and stimulation of spinal-sensory supplied skin (Figure 3(a)). Such neurons could display low or elevated baseline firing rates and change their action potential discharge during oral presence of aversive bitter (quinine, cycloheximide) and acidic (citric acid) taste solutions, oral delivery of noxious heat and chemesthetic stimuli, and pinch or photostimulation of TRPV1-lineage afferents on bodily extremities supplied by spinal afferents, like the hindpaw (e.g. Figure 2(b) and (c)). While responses to oral stimuli were commonly excitatory (increased spikes), pinch and photostimulation directed to the hindpaw or tail could cause excitation or inhibition in PB unit spike discharge (Table 1, Figure 2(b)–(d)). One additional taste-active PB neuron was encountered that co-responded to spinal, but not oral, somatosensory stimuli (Figure 3(a)). Thus, 11 out of 94 PB neurons (11.7%) were identified as “taste-spinal” cells that received convergent spinal afferent messages and gustatory signals.
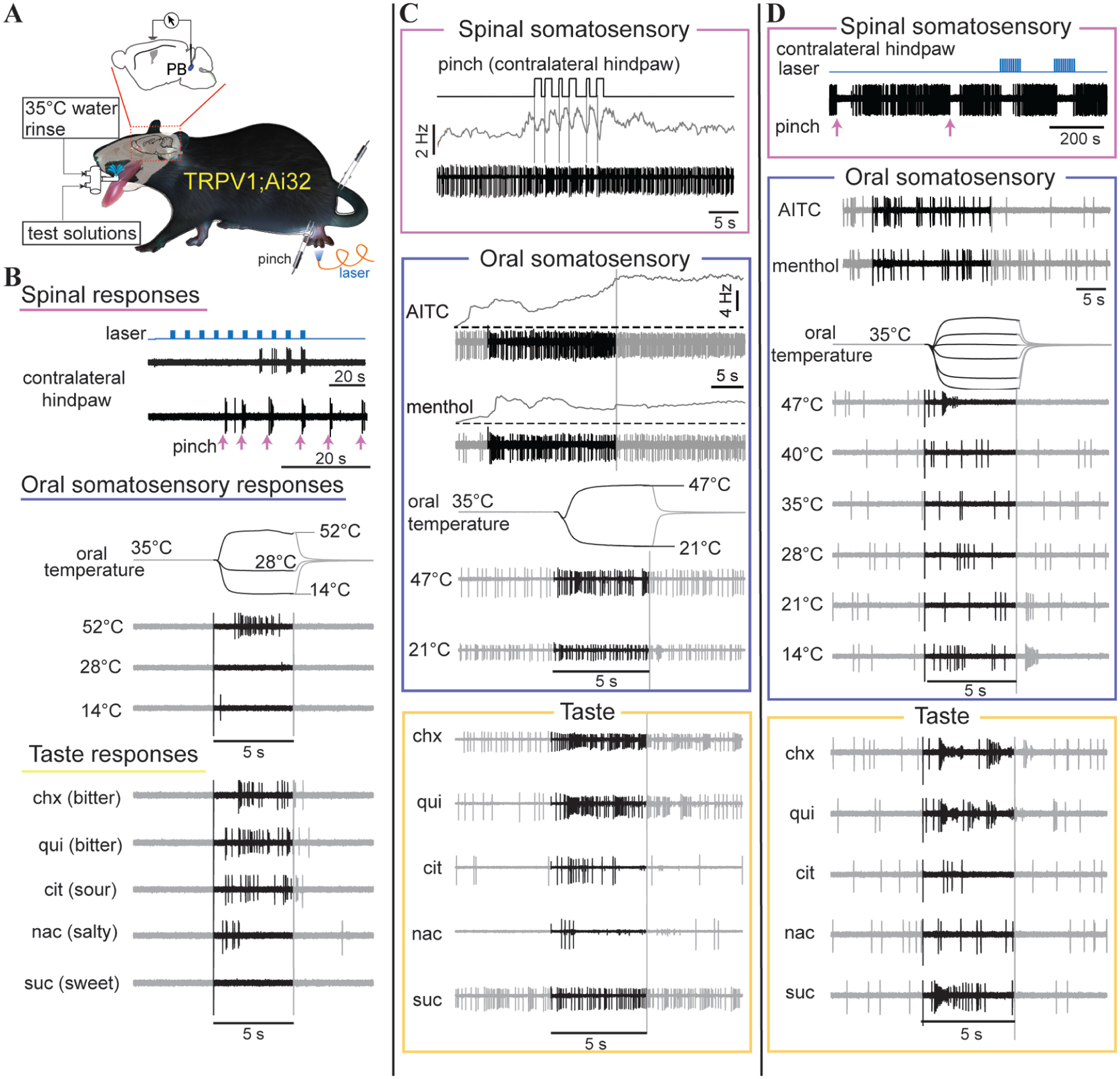
Electrophysiological responses to taste, oral somatosensory, and spinal somatosensory stimuli in individual neurons in the parabrachial nucleus (PB): (a) Illustration of experimental schematic. Extracellular action potentials were recorded from individual PB neurons in TRPV1;Ai32 mice during oral delivery of taste, thermal, and chemesthetic solutions (test solutions), pinch of the hindpaw and tail (forceps), and photoexcitation of hindpaw TRPV1-lineage fibers (blue laser). (b) Electrophysiological response traces acquired from one PB neuron excited by pinch (upward arrows) and photoexcitation of TRPV1-lineage fibers (laser pulses) on the contralateral hind paw, oral presence of noxious hot water (52°C), and bitter (quinine (qui), cycloheximide (chx)) and acidic (citric acid (cit)) taste stimuli. Black regions of electrophysiological traces for oral stimuli reflect stimulus periods (5 s). Temperature traces capture change in oral temperature over time, aligned with spike traces; final temperatures are expressed in oral °C. Other abbreviations: NaCl (nac) and sucrose (suc). Abbreviations and stimulus markers are consistent across all panels in Figures 2 and 3. (c) Electrophysiological response traces acquired from a second PB neuron excited by calibrated pinch (upward TTL deflections) of the contralateral hind paw, oral presence of the chemesthetic nociceptive agent allyl isothiocyanate (AITC) and also noxious hot water (47°C), and the bitter taste stimuli cycloheximide (chx) and quinine (qui). Traces above pinch and chemesthetic spike responses indicate mean impulse frequency. (d) Electrophysiological response traces acquired from a third PB neuron that was inhibited by TRPV1-lineage afferent photostimulation (laser) and pinch (upward arrows) on the contralateral hindpaw, but showed excitatory responses to oral presence of noxious hot water (47°C), and bitter (quinine (qui), cycloheximide (chx)) and sweet (sucrose (suc)) taste stimuli. Note that in this cell, action potentials decreased in amplitude during periods of rapid firing on trials with heat and bitter taste stimuli.
Numbers of taste-spinal neurons that were excited and/or inhibited by pinch of the hindpaw and tail.
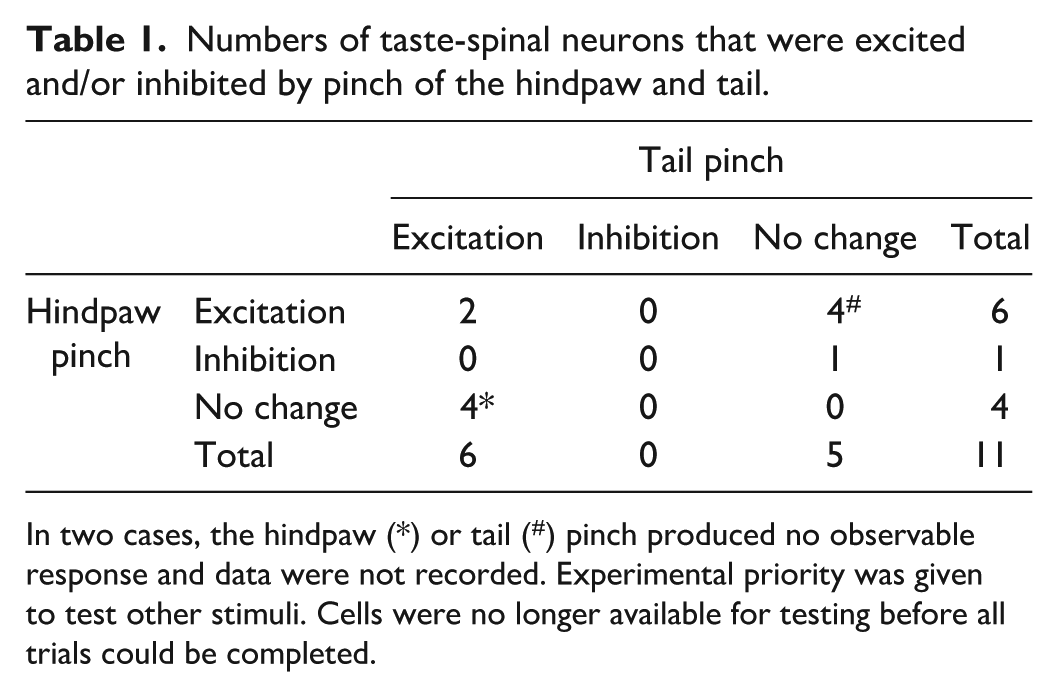
In two cases, the hindpaw (*) or tail (#) pinch produced no observable response and data were not recorded. Experimental priority was given to test other stimuli. Cells were no longer available for testing before all trials could be completed.
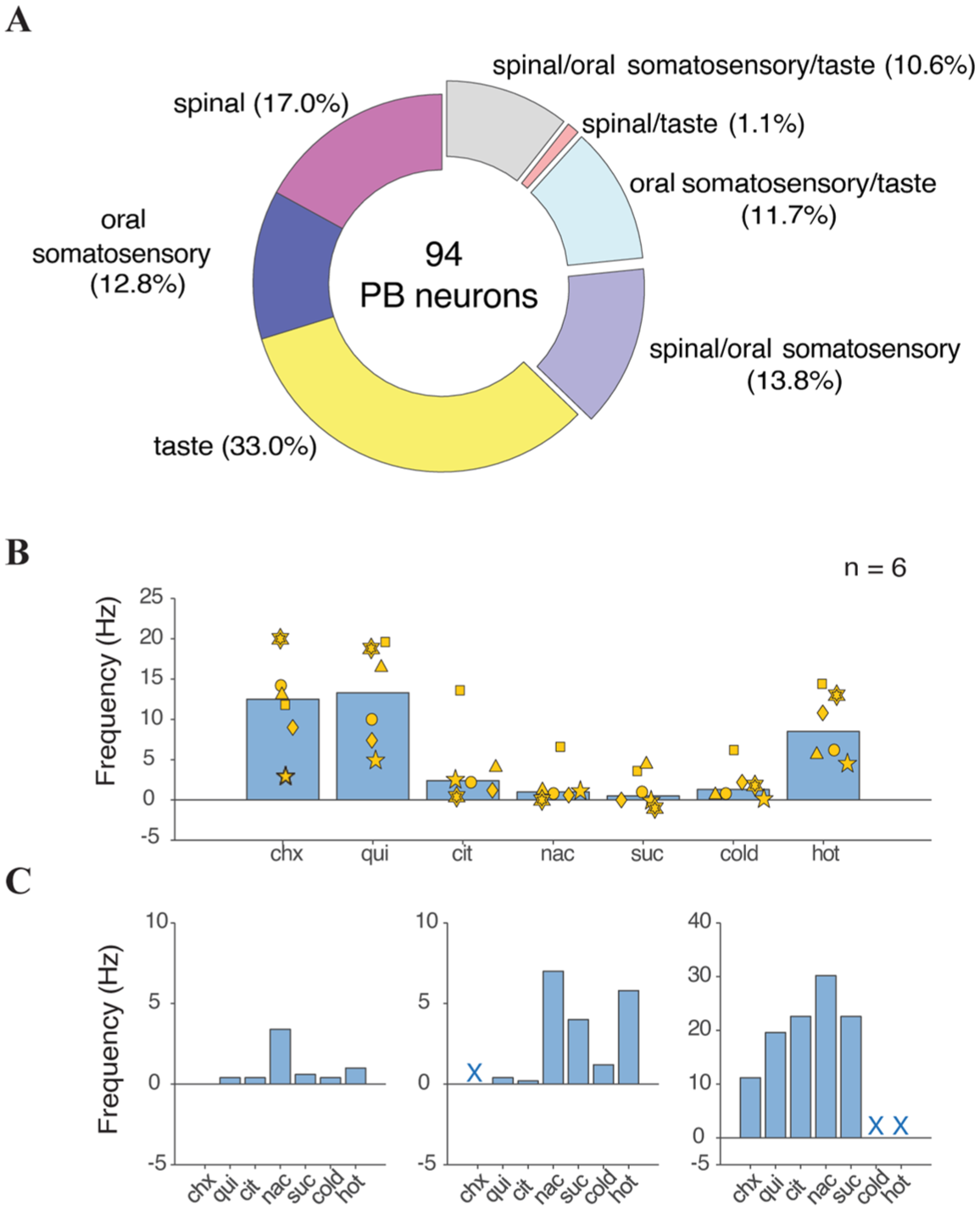
Sensory response characteristics of the sampled PB neurons: (a) Percentages of the 94 PB neurons that were excited by different unimodal and multimodal combinations of taste, oral somatosensory, and spinal afferent stimulation. (b) Mean responses by the bitter-oriented taste-spinal PB neurons (n = 6) to oral presence of taste solutions, and cold (<15°C) and noxious hot (>46°C) water. Responses by each neuron are represented by a common marker. (c) Response rates for three individual NaCl-oriented taste-spinal PB neurons to oral presence of taste solutions, and cold (<15°C) and noxious hot (>46°C) water. Missing trials are denoted by “X”.
Six of 11 taste-spinal neurons exhibited quinine/cycloheximide (bitter)-dominant taste profiles based on their evoked response magnitudes across taste stimuli (Figure 3(b)). Other taste-spinal neurons showed NaCl-dominant taste profiles (Figure 3(c)). Finally, one was classified as a sucrose/quinine-dominant cell where pinch and photoexcitation of TRPV1-lineage afferents on the contralateral hindpaw inhibited spontaneous activity (Figure 2(d)). Among eight taste-spinal neurons where location was histologically recovered, seven emerged in lateral PB regions and one was located near the external medial PB subnucleus (Figure 1(b)), which has been implicated in spinal-sensory processing. 5
Discussion
Here we identified a subpopulation of taste-excited PB neurons in mice that changed their action potential discharge rate when pain-related stimuli were applied to skin on the hindpaw or tail. These stimuli included pinch and transdermal photoexcitation of TRPV1-lineage afferents linked to thermal-nociceptive processing. Taste-spinal integrative neurons were found in neural recordings targeted to the lateral PB area. Co-sensitivity to oral presence of chemesthetic and thermal stimuli associated with trigeminal nociception was also prevalent among these cells. Our findings complement and extend data showing convergence of ascending taste and trigeminal signals onto individual PB neurons8,9 and that spinal sensory input reaches PB regions where taste-trigeminal confluence occurs.2–4
We report that electrophysiologically locating taste-spinal integrative neurons in the PB area was challenging, with this feature appearing in only 11 (about 12%) of the 94 PB neurons we recorded. This reduced cell number and the complexities associated with delineating overall excitatory, and inhibitory, spinal receptive fields in PB neurons 5 limited our analysis to a qualitative description of taste-spinal integrative phenomena. A potential caveat to our approach is that we examined how PB neurons would respond to stimulation of only focused, and limited, body regions. However, prior work suggests that rodent PB neurons that integrate cutaneous nociceptive signals from diverse bodily locations commonly display receptive fields that include hindpaw or tail skin.5,18
While extensive cellular sampling was performed, the reduced occurrence of taste-spinal neurons we encountered should be cautiously interpreted in the context of their actual proportions in PB circuits. This is because, in part, cells were recorded individually across multiple mice. Additionally, there were occurrences during the present recording sessions where taste-excited neurons were isolated while spinal stimulation engaged nearby background cells, but co-responsivity to these inputs in one neuron was not observed. Thus, taste and spinal pathways may also stimulate adjacent PB microcircuits without intersection, as gauged by the present conditions.
Six out of the 11 taste-spinal neurons we encountered responded most strongly among taste solutions to bitter quinine and cycloheximide, which evoke innate concentration-dependent reductions in fluid licking (i.e. avoidance) in rodents.12,15,23,30 This finding is consistent with prior data showing responsiveness to quinine and cycloheximide can predict co-sensitivity to trigeminal nociceptive stimuli in taste-active PB neurons.8,9 Further, these results agree with the concept that a negative hedonic (i.e. aversive) dimension partly guides sensory information convergence onto neurons in the PB area to generate a protective signal.5,18 Cell types evidenced to contribute to this signal include lateral PB cells expressing calcitonin gene-related peptide (CGRP), which were previously established to support convergence of protective sensory information from diverse modalities and body regions. 18 PB-CGRP neurons were recently implicated with roles in unconditioned gustatory behavioral avoidance of bitter quinine.23,31 PB-CGRP neurons are also evidenced to participate in neural and behavioral responses to appetitive stimuli, including sugar (sweet) taste solutions, in addition to aversive states,23,32 implying their roles may be more varied than previously found.
PB neurons, including lateral CGRP-expressing cells, are associated with promoting arousal under diverse conditions.33–37 That the present taste-spinal neurons showed brief transient changes in firing to hindpaw/tail stimulation and stimulus-selective responses to tastes and spinal input would agree with neural activity of sensory origin. Stimulus selectivity would also suggest that evoked PB activity was not attributable to vagaries in anesthesia state, which was monitored during recording sessions by lack of a reflex response to heavy pinch. Nonetheless, it is curious if the presently recorded cells normally function to stimulate arousal to external events.38,39 Understanding this would require, in part, the use of experimental techniques different from the present, including monitoring taste-spinal neurons in unanesthetized mice to examine their modulation with behavioral states.
Anesthesia was used in the present studies to support extended isolation of action potential waveforms generated by single PB neurons and for control over mechanical and optical cutaneous sensory stimulation parameters. 40 While certain features of evoked and spontaneous activity in PB neurons are reduced by anesthesia,36,37,41,42 sensory responses, including nociceptive (e.g. pinch5,18) and taste,43,44 have been consistently measured from rodent PB cells under different anesthetic conditions. Importantly, gustatory tuning profiles observed for individual PB neurons in anesthetized mice 44 resemble those measured from awake mice, 31 with similar rates of gustatory impulse discharge found for PB cells in anesthetized and unanesthetized rats. 42 Moreover, calcium imaging studies in awake mice show PB-CGRP neurons can display increased calcium during ingestion, and presumably tasting, of quinine and also following foot shock (spinal input), yet only a small fraction of these cells respond to both stimuli. 45 This would agree with the present low number of taste- and spinal-responsive neurons we encountered in PB recordings under anesthesia. Despite these similarities, it will be important in future work to assess if the pattern of taste-spinal sensory convergence across PB neurons is comparable between anesthetized and awake conditions to fully index its functional significance.
Convergence of taste-related and spinal sensory information was reported in calcium imaging studies of insular cortex in mice. In this work, a subgroup of cortical cells displayed increased calcium to oral presence of quinine and tail shock, which contributed to an interpretation that such effects reflect a cortical circuit that registers aversive and affective states. 46 The present, and other,32,45,47 data suggest that cross-sensory integration of taste and spinal pain-related signals can arise subcortically, in the brain stem – an early waypoint for ascending gustatory and somatosensory codes. Subcortical-cortical interactions can influence the integration of crossmodal sensory signals, 48 with PB circuits presently appearing to partly link oral taste and spinal pain-related messages based on their valence. Notably, the present work indicates that PB neurons supporting this function could be misclassified as “taste cells” if assessed for only gustatory responsiveness by traditional means.
Lateral PB circuits can receive bilateral input from spinal projection neurons, with evidence that select genetically identifiable spinoparabrachial circuits can engage behaviors in mice associated with affect. 49 Spinoparabrachial neurons can aggregate nociceptive messages onto broadly integrative PB neurons,4,5,18 with the present work suggesting that some spinal-integrative PB cells show co-sensitivity to taste. Additional work is needed to understand the synaptic and circuit mechanisms mediating taste-spinal convergence. It remains unclear if spinal projections reach taste-active PB neurons through mono- or polysynaptic pathways, with evidence that intermediate circuits in the PB relay spinal messages to lateral PB regions associated with nociception and sensory integration. 50 Future studies may also inform whether examining the potential for cross-modal interactions between gustatory and spinal-sensory signaling in the PB nucleus has value for studying modulation of neural transmission under pathological conditions such as chronic pain.
Footnotes
Acknowledgements
We thank Dr. Chris Brooks for his thoughtful comments and feedback on an early draft of this paper.
Author note
Portions of these data were presented in abstract form at the 2025 meeting of the Association for Chemoreception Sciences (AChemS) and the 2025 meeting of the Society for Neuroscience.
Author contributions
C.H.L. conceived research project. J.L. designed and performed experiments. J.L. and C.H.L. analyzed data, made figures, and wrote the paper.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by NIH grant DC011579 to C.H.L.