
Abstract
Recent research on the relation between learning and cognitive control has assumed that conflict modulates learning, either by increasing arousal and hence improving learning in high-conflict situations, or by inducing control, and hence inhibiting the processing of distracters and their eventual association with the imperative responses. We analyse whether the amount of conflict, manipulated through the proportion of congruency in a set of Stroop inducer trials, affects learning of contingencies established on diagnostic trials composed by neutral words associated with colour responses. The results reproduced the list-wide proportion of congruency effect on the inducer trials, and showed evidence of contingency learning on the diagnostic trials, but provided no indication that this learning was modulated by the level of conflict. Specific analyses conducted to control for the impact of episodic effects on the expression of learning indicated that contingency effects were not driven by the incremental processes that could be expected by associative learning, but rather they were due to the impact of the most recent trial involving the same distracter. Accordingly, these effects disappeared when tested selectively on trials that required a non-matching response with respect to the previous occurrence of the distracter. We interpret this result in the context of the debate on how learning and memory interact with the processes of cognitive control.
Intelligent behaviour involves a complex balance between two hardly compatible abilities: it requires us to keep focused on our long-term goals while resisting interference arising from more habitual and compelling response tendencies (Cohen, 2017), and it also requires us to remain flexible, considering detours and updating our plans whenever we encounter contexts from which we can take advantage to improve our overall outcomes (Braem & Egner, 2018). The mechanisms of cognitive control and their adaptive functions are jointly responsible for finding the right balance between these two opposing pressures, by adopting strategies that satisfy our goals and by updating them continuously in the most efficient way.
Research on the mechanisms of adaptive control has been conducted using variants of three main interference paradigms, designed by Stroop (1935), Simon and Wolf (1963), and Eriksen and Eriksen (1974). In these paradigms, stimuli are composed of two main features, one of which is irrelevant, but strongly associated with the response dimension, while the other is relevant, but less directly linked to the imperative response. In the Stroop task, for instance, the meaning of words referring to colour names (e.g., RED) plays the role of the irrelevant dimension, and participants are instructed to report on the colour of these words, regardless of their semantic content. The results indicate that participants strive to ignore the irrelevant word meanings, but their responses get slower and less accurate when this semantic dimension is incongruent with the relevant one. Moreover, even though this congruency effect appears to be automatic, in that, it cannot be intentionally avoided, it gets modulated by past experience, as shown by its reduction after an incongruent trial (Gratton et al., 1992), on trial lists containing mainly incongruent trials (Logan & Zbrodoff, 1979), and for contexts (Crump et al., 2006) or even single items (Jacoby et al., 2003) associated with a high proportion of incongruent trials.
The modulation of congruency effects in terms of past experience has been interpreted as showing that the amount of control exerted by the cognitive system becomes automatically adapted to past conflict. This mechanism was modelled by the influential model of conflict monitoring (Botvinick et al., 2001), but since then, a number of alternative accounts have been proposed, emphasising the roles of learning and memory over the modulation of control functions (see Egner, 2017, for a comprehensive review). These alternatives suggest that the modulation of congruency effects may arise from memory effects if both relevant and irrelevant features from an episode are bound together and become encoded in memory as a single “event file” (Hommel et al., 2004), that would allow for a more efficient response if the same episode is repeated on the following trial, as compared with cases in which the following trial contains no feature repetition, or includes the repetition of only one of the members of the ensemble (Mayr et al., 2003). Other authors suggested that targets and distracters may become associated with practice, producing faster responses for those pairs that reoccur more often (Schmidt & Besner, 2008). Further proposals suggested that learning may indeed interact with cognitive control functions, so that a control state can become associated with a specific context (Egner, 2014), or that the amount of control required in a trial can modulate the efficiency with which associative learning works, hence affecting the way in which control will be exerted on the following trials, as a consequence of the strengthening of these associative bindings (Verguts & Notebaert, 2008, 2009).
The purpose of the present study is to further analyse the learning processes that might be playing a role in determining the dynamics of cognitive control in interference tasks (Abrahamse et al., 2016). As a case in point, Whitehead et al. (2018) reported evidence showing that contingency learning is affected by the amount of conflict encountered in such tasks. Specifically, they reported that contingency learning was larger in a group exposed to non-conflicting trials compared with a group that performed exclusively on incongruent trials. The authors tested four conditions of a Stroop task, using either neutral words or incongruent words as distracters, and arranging two levels of word–colour contingencies. Participants showed evidence of learning in all groups, but they showed larger effects in those exposed to higher contingencies, and in those trained with neutral words. Moreover, the authors reported an interaction between conflict and contingency levels, which was interpreted as showing that the presence of conflict did effectively reduce the impact of contingency learning.
Whitehead et al. (2018) discussed their results in contrast to episodic views (Schmidt, 2013) that predicted equivalent effects regardless of the amount of conflict, and in favour of models that assume that the presence of conflict should modulate the amount of associative learning induced by the repeated target-distracter pairing. Indeed, the conflict monitoring hypothesis (Botvinick et al., 2001) predicted that, in conditions of high proportion of conflict, participants would inhibit the processing of distracters, while paying more attention to the targets. This would produce not only a decrease in the effect of congruency, but also a reduction in any enduring association produced between repeated targets and distracters (Blais et al., 2007). In contrast, however, the model of “adaptation by binding” proposed by Verguts and Notebaert (2008, 2009) posited an arousal–learning interaction, according to which conflict would trigger a neuromodulatory response that would improve the efficiency of Hebbian learning processes, thus strengthening the connections between all representations active at this moment, even if they are irrelevant to the task.
The pattern of results reported by Whitehead et al. (2018) reinforces the conclusion that word–colour associations may decrease in conditions of high conflict. However, a possible confound of this design comes from the fact that learning was assessed in trials that differed not only in the overall amount of conflict created by the surrounding trials, but also with respect to the semantic relation established in that particular trial between distracters and targets: In the conflict conditions, the word–colour associations involved words already associated with a different member of the response category (e.g., GREEN-> blue), whereas in the neutral conditions, they corresponded to words not previously associated with any of the colours (e.g., GRIP-> blue). As pointed out by Schmidt (2019, see also Schmidt, De Houwer, & Moors, 2020), this might have biased the results due to the higher difficulty of learning associations contradicting previously over-learned ones. Moreover, to ascertain whether cognitive control does really affect learning in a continuous and reactive way, rather than provoking a strategic decision to minimise attention to the distracters, it would be important to reveal whether similar differences in contingency learning arise in terms of the overall proportion of congruency, rather than in terms of the absolute presence versus absence of conflict over the full procedure.
To analyse whether the overall level of conflict in an interference task really affects the efficiency of associative learning, we set out to extend the effects reported by Whitehead et al. (2018) to conditions differing on the overall proportion of conflict trials, and in which the impact of the overall level of conflict was measured on the learning of comparable contingencies established between neutral words and target colours. This procedure allowed us to separate inducers from diagnostic trials, a strategy proposed as an optimal procedure to measure the effects of adaptive control on interference tasks (Braem et al., 2019). We used colour words to induce a different list-level proportion of congruency, and included neutral words contingently associated with the target colours as the diagnostic trials on which to assess the effect of conflict on associative learning. The contingency values for the neutral words as predictors of the colour responses was fixed at ΔP = .5. 1 The proportion of conflict was manipulated between groups, including colour words in about half of the trials, and arranging a different proportion of congruent and incongruent trials in each group. We compared two groups of high versus low conflict that experienced conflict, respectively, in 80% and 20% of their inducer trials.
Finally, since some recent studies (Giesen et al., 2020; Schmidt et al., 2020) have raised the concern that contingency learning in these settings could be reduced to episodic effects, with a special impact coming from the most recent occurrence of the distracter rather than to a gradual accrual of associative strength, we sought to explore whether recency effects might be responsible for these effects, and could be unduly taken as evidence of contingency learning. Indeed, the higher frequency of high contingency trials implies that the most recent occurrence of any given distracter would tend to be another high contingency trial, one that would match the response required by the subsequent high contingency trial, but that would prime a non-matching response in the case of a subsequent low contingency trial. In other words, the different contribution of recency effects over high and low contingency trials could be passing as evidence of associative learning. To distinguish between this recency factor and the potential impact of more gradual and associative learning processes, we assessed whether recency effects were present in this paradigm, and reassessed the effects of proportion of congruency and contingency learning selectively on trials in which the influence of recency matching was not present. We did so by excluding all trials in which the most recent appearance of the current distracter had been paired with the same response as in the present trial, thus measuring the effect exclusively on trials that required a response different from that primed by the most recent previous experience with that distracter (see Figure 1).
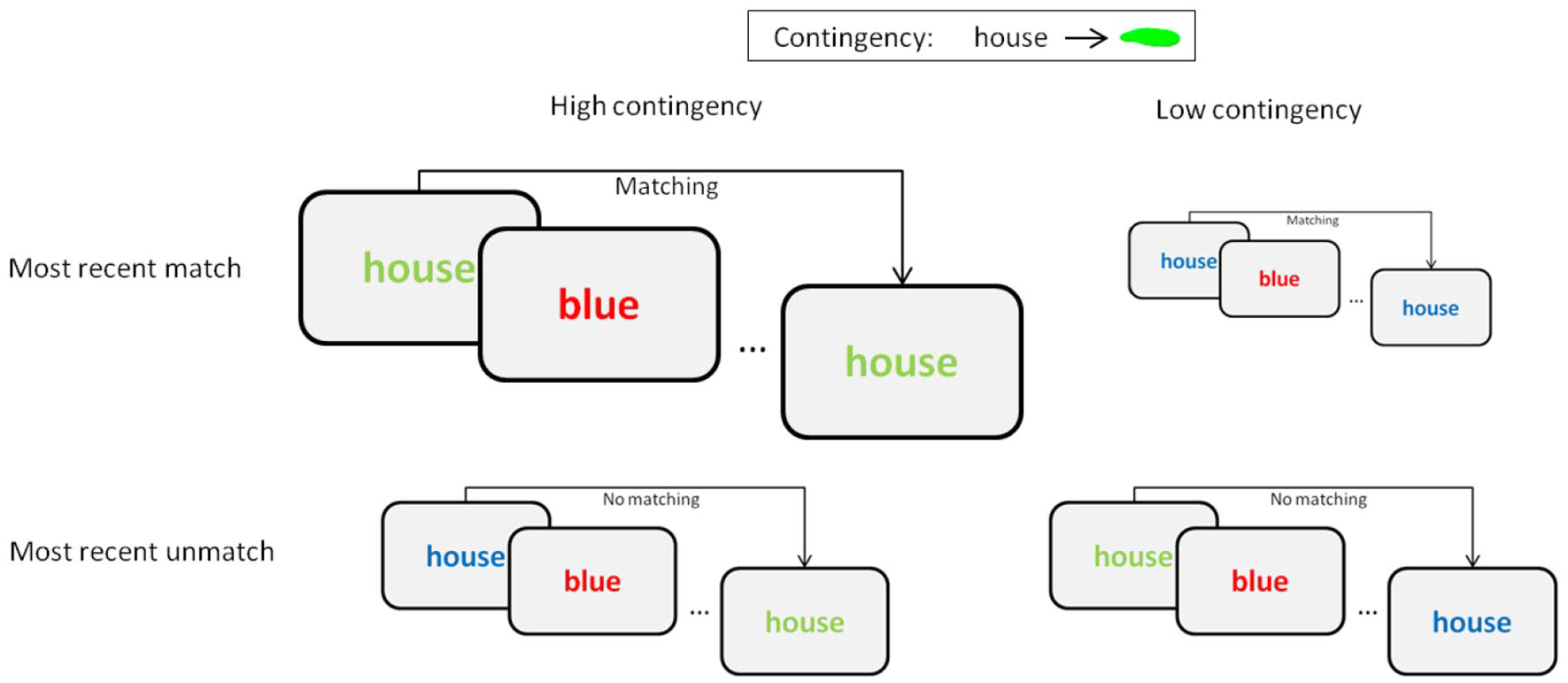
Illustration of the relationship between word–colour contingency (represented in columns) and recent matching effects (represented in rows), for a hypothetical relation between the word “house” and colour green. If this particular combination was highly likely, the most recent occurrence of the word “house” prior to another episode of the same word would often correspond to the ensemble house-in-green. This amounts to a matching trial for high contingency trials (depicted on the upper left quadrant), but to a non-matching trial for the low contingency condition (on the lower right quadrant). The relative size of each of these diagrams represents the frequency of each of these possible cases, showing how recent matching effects could affect the measures of contingency learning. To control for this impact, we assessed contingency learning effects exclusively on non-matching trials (i.e., on the type of trials depicted on the bottom row).
In sum, the purpose of the experiment was to assess the impact of the overall proportion of congruent trials on learning associations between neutral distracters and the colour responses associated to them in the context of a four-choice manual Stroop task. As a way of distinguishing between inducers and diagnostic trials, about half of the trials were composed of colour words which could be congruent or incongruent with the target colours. The other half of trials included four neutral words, probabilistically associated with each of the four colours. This arrangement allowed us to ascertain whether participants were sensitive to the proportion of congruency, whether they were able to learn about the word–colour contingencies established on neutral trials, and whether that learning was affected by the frequency with which participants faced conflict trials.
To foreshadow the results, we found that participants were sensitive to the overall proportion of congruency, as they showed higher congruency effects on the inducer colour trials in the condition of low conflict. They were also sensitive to the manipulation of contingency learning, as they responded faster to those diagnostic trials in which the target was highly associated with the neutral distracter. However, the results showed no evidence of differences in contingency learning depending on the conflict conditions. Moreover, when learning was reassessed exclusively on trials in which the most recent previous occurrence of the distracter was paired with a different target, to remove the episodic impact derived from the most recent matching, the effect of contingency learning disappeared.
Methods
Participants
The experiment was conducted in accordance with Spanish regulations, and complied with the ethical standards of the 1964 Declaration of Helsinki. We used G*Power 3.1.9.2 to compute the sample size necessary to have at least a power of .80 to obtain results comparable with those found in the work of Whitehead et al. (2018). These authors reported an eta partial squared of .068 for the condition × congruency interaction, amounting to an effect size f = .27. Assuming a correlation among repeated measures of .5, a sample of 30 participants would be enough to get that power, but we almost duplicated the sample to obtain a power of .975 of capturing a comparable effect size. Fifty-six volunteers (45 women) aged 18–43, (M = 23, SD = 3.9) were recruited by word of mouth, and signed informed consents to participate in the experiment. Most of them were students from the University of Santiago de Compostela, and they were all native Spanish speakers. Half of them were randomly assigned to the low- or high-conflict conditions.
Apparatus and stimuli
The experiment was designed and controlled using INQUISIT 4 (Millisecond_Software, 2015) software, running on personal computers connected to 22 inch monitors with resolutions of 1,920 × 1,080. Participants viewed the monitors from an unrestricted distance of approximately 60 cm, and responded on a standard QWERTY keyboard placed in front of them. On each trial, participants saw a word corresponding either to a set of four Spanish words referring to the colours red (“rojo”), blue (“azul”), green (“verde”), and yellow (“amarillo”), or to the following set of arbitrary neutral words: car (“coche”), house (“casa”), plant (“planta”), and zone (“zona”). All the words were in Arial, lower case, 32-point font, coloured either in red, blue, green, or yellow, and were presented against a grey background. Participants were asked to respond to the colour in which the word was written using the keys corresponding to the letters “z,” “x.” “n,” and “m” to respond to the colours red, green, blue, and yellow, respectively. The response keys were marked with appropriate coloured stickers.
Procedure
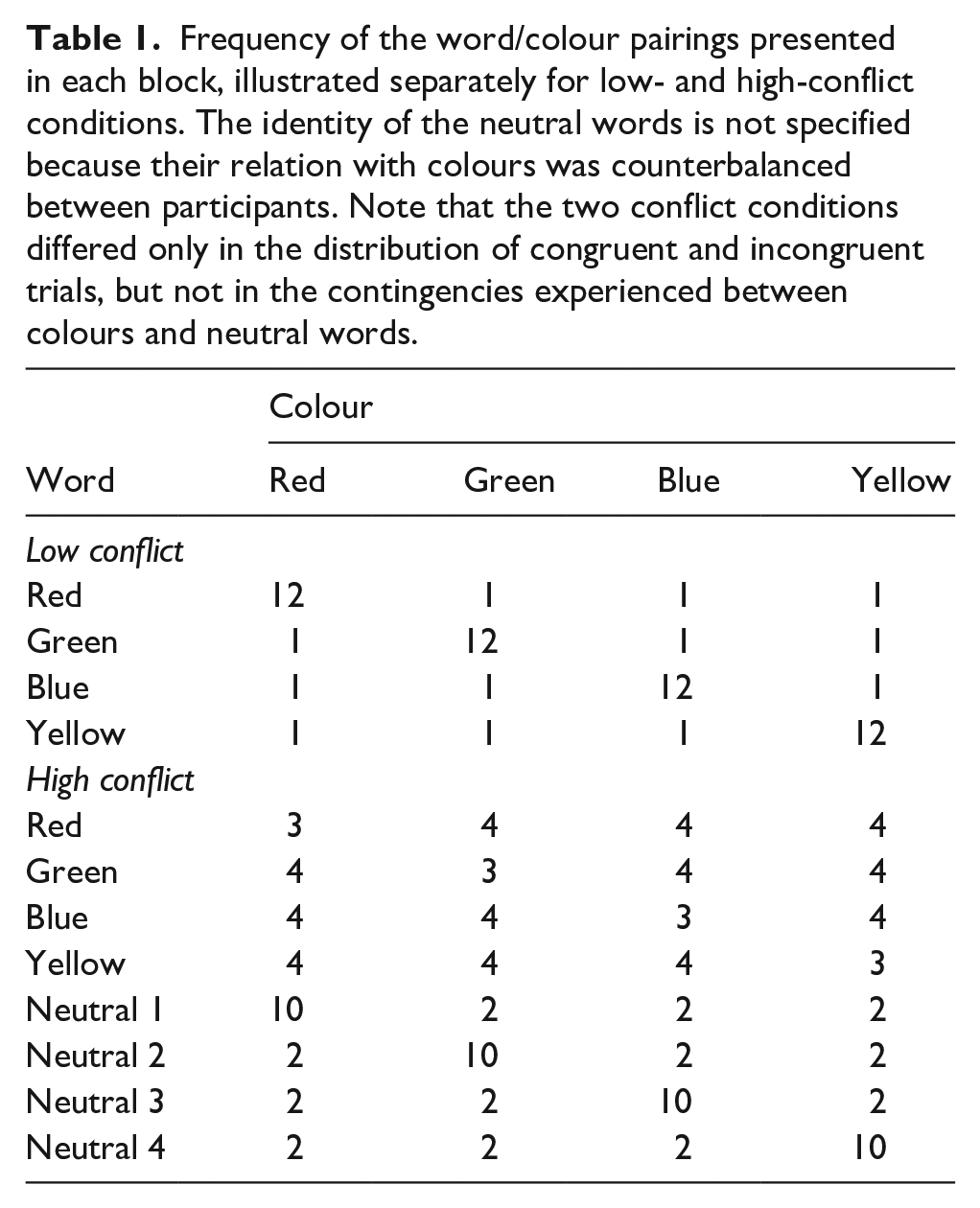
Participants were randomly assigned to either low- or high-conflict conditions, and began the task with a pre-training block composed of 50 trials, in which they were able to familiarise themselves with the key-colour mapping, using only the neutral words, which were randomly chosen and paired with any of the four possible target colours. During this initial block, the mapping between colours and response locations was explicitly marked at the bottom of the screen, using four word labels displayed at the relative position corresponding to the location of its response key. After this block, the word labels were removed, and participants were warned that they should keep responding to the colour in which the words were written, but that now, they could see trials containing either one of the previously presented words or new words corresponding to the names of these four target colours. The experimental task was performed over five blocks of 124 trials, 60 of these corresponding to colour words and 64 corresponding to neutral words. The specific association between neutral words and colour responses was counterbalanced between participants, so that each neutral word was most frequently paired with a different colour for each participant, producing four possible arrangements. The frequency of each word/colour pairing on each block is shown in Table 1.
Frequency of the word/colour pairings presented in each block, illustrated separately for low- and high-conflict conditions. The identity of the neutral words is not specified because their relation with colours was counterbalanced between participants. Note that the two conflict conditions differed only in the distribution of congruent and incongruent trials, but not in the contingencies experienced between colours and neutral words.
Each trial began with a fixation cross presented at the centre of the screen, which was replaced by a blank screen after 750 ms. This was followed by the coloured word after 250 ms, and it remained on the screen until response. Correct responses were immediately followed by the next trial. To discourage errors, these were marked by a tone, and followed by a blank screen that delayed the progress of the trials for 2,000 ms. After an experimental block, participants were allowed to take a short rest. During this period, they were informed about their average reaction times and the proportion of correct responses produced over the past block, and were reminded that their goal was to respond as fast as possible, while maintaining the proportion of correct responses above 90%. At the end of the five experimental blocks, participants were presented with a short survey in which, for each distracter word, they could choose with which colour was it more frequently paired. If they did not believe that a particular word was more frequently paired with any of the four targets, they were also allowed to respond that the word was evenly paired with the four colours.
Design
The experiment included the conditions of high versus low conflict as a between-participants variable, and two within-participants factors: congruency (congruent vs incongruent trials) and contingency (high- vs low-frequency pairings). As a preliminary test of the efficacy of the conflict and contingency manipulations, we analysed participants’ responses to the final survey, as a way to ascertain whether participants in each group noticed the different proportion of congruent trials, as well as the contingencies introduced between the neutral words and each colour response. Moreover, we analysed the effect of the overall proportion of congruency on their responses to colour words, as a way to confirm that the congruency effect was larger for the low-conflict condition than for the high-conflict condition. Then, we assessed the effect of contingency on the neutral words, and tested for the possible influence of the level of conflict on the magnitude of this contingency learning. Finally, as a means of discerning between a gradual accrual of associative learning and the episodic impact of the most recent episode involving the same distracter, we analysed the recency effects on the overall performance on any given distracter, depending on whether or not the most recent presentation of the distracter matched the same response, regardless of the number of trials spanning between the two events. If this effect was significant, we planned to analyse the effects of list-level proportion of congruency and contingency, relying exclusively on the trials in which the relevant response did not match that required on the most recent presentation of the same distracter. 2
With the exception of the results coming from the post-experimental questionnaire, which were analysed through chi-square tests to ascertain whether the proportion of responses of each type differed between groups, for all the other analyses, we used response time (RT) and proportion of correct responses as dependent variables, conducting repeated-measures analyses of variance (ANOVAs) to test the main hypotheses. RT is usually most sensitive to these effects, and thus we focused mainly on the measures of latency, and we also analysed errors for completeness, and to confirm that any effect observed in RT could not be attributed to a speed–accuracy tradeoff. From the measures of RT, we removed the first trial of each block, and RT from incorrect responses and from the trial that immediately followed an error. From the remaining trials, we computed the mean and standard deviation for each block and participant, and we also removed outliers, defined as those trials with RT beyond three standard deviations from the mean. As for the proportion of correct responses, we removed the first trial from each block and the trial that immediately followed an error, because we were interested in measuring errors attributable to the conditions presented on any given trial, not those that might have been motivated by the error committed on the preceding trial.
Results
Postexperimental survey
Responses to the question of what colour was more frequently paired with each colour word was coded in three different categories, in terms of whether it corresponded to the congruent colour, to any one of the three remaining colours, or to the residual category: “all evenly paired.” On average, participants in low-conflict condition selected the congruent colour in 61.6% of their choices, whereas they chose one of the incongruent colours in 24.1%, and the residual category in the remaining 14.3%. In contrast, participants in high-conflict condition chose the congruent colour in 15.2% of their choices, whereas they chose an incongruent colour in 56.3%, and reported that all were evenly paired in 28.6% of them. Chi-square tests conducted separately for each colour word indicated that the pattern of choices differed between groups in response to the four colour items: χ2 RED (2, N = 56) = 13.38; p < .001; χ2 GREEN (2, N = 56) = 11.52; p = .003; χ2 BLUE (2, N = 56) = 18.05; p < .001; and χ2 YELLOW (2, N = 56) = 15.46; p < .001. In contrast, similar analyses conducted for the neutral words to ascertain whether participants in each group were able to discriminate on the colour that was contingently associated with those words indicated that responses were similar between groups: χ2 neutral 1 (2, N = 56) = 1.15; p = .56; χ2 neutral 2 (2, N = 56) = 3.49; p = .18; χ2 neutral 3 (2, N = 56) = 4.33; p = .12; and χ2 neutral 4 (2, N = 56) = 2.80; p = .25. On average, participants chose the correct, contingently associated colour only in 25.9% and 15.2% of their choices, respectively, in high- and low-conflict groups; they chose one of the non-contingent colours in 50% and 58% of the cases, and selected the residual category in 24.1% and 26.8%, respectively, for each group. Thus, participants exposed to a high proportion of congruent trials were able to notice it, but they were not more able than those exposed to a low proportion of congruent trials to detect the contingency established between the colours and neutral words.
Proportion of congruency effect
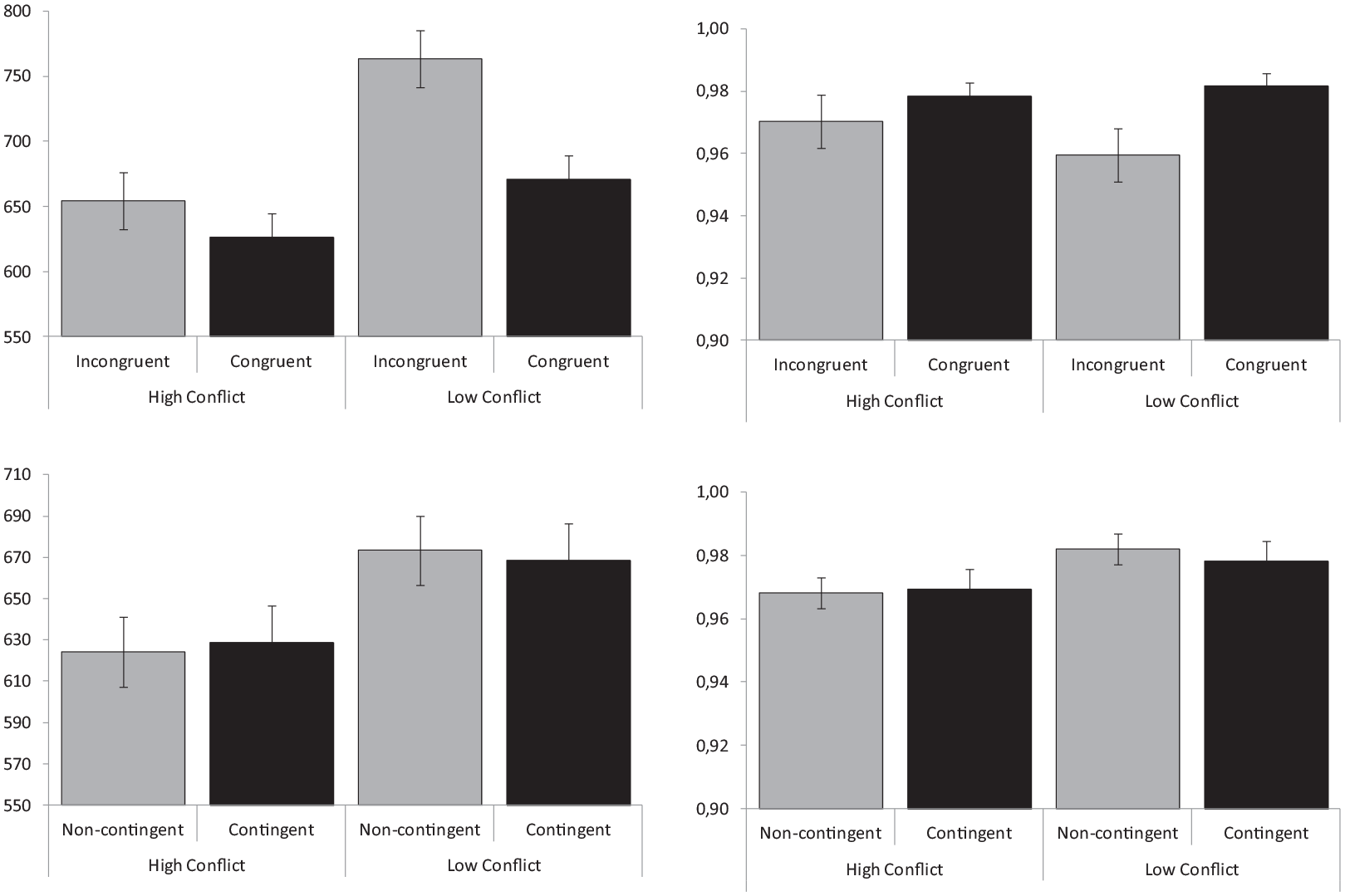
Average RT and proportion of correct responses obtained in response to congruent and incongruent trials are represented in Figure 2, separately for participants trained in conditions of high versus low conflict. The repeated-measures ANOVA conducted on RT with congruency as a within-participants factor and with conflict level as a between-participants variable showed, somewhat surprisingly, that participants assigned to high-conflict conditions responded faster than those trained with low-conflict conditions (M = 636 vs 706 ms), F(1,54) = 6.55, p = .01; ηp2 = .11. The main effect of congruency was also significant, F(1,54) = 102.13, p < .001; ηp2 = .65, showing faster responses to congruent than to incongruent trials (M = 633 vs 708 ms), and so was the congruency × conflict-level interaction, F(1,54) = 38.43, p = .001; ηp2 = .42, which showed larger effects of congruency for the low-conflict (M = 645 vs 766 ms) as compared with the high-conflict condition (622 vs 650 ms). The analysis conducted on the proportion of correct responses only showed a significant effect of congruency, F(1,54) = 6.56, p < .013; ηp2 = .11, but not an effect of conflict level, F(1,54) = 0.05, p = .83; ηp2 = .001, nor a significant interaction between them, F(1,54) = 3.14, p = .08; ηp2 = .06.
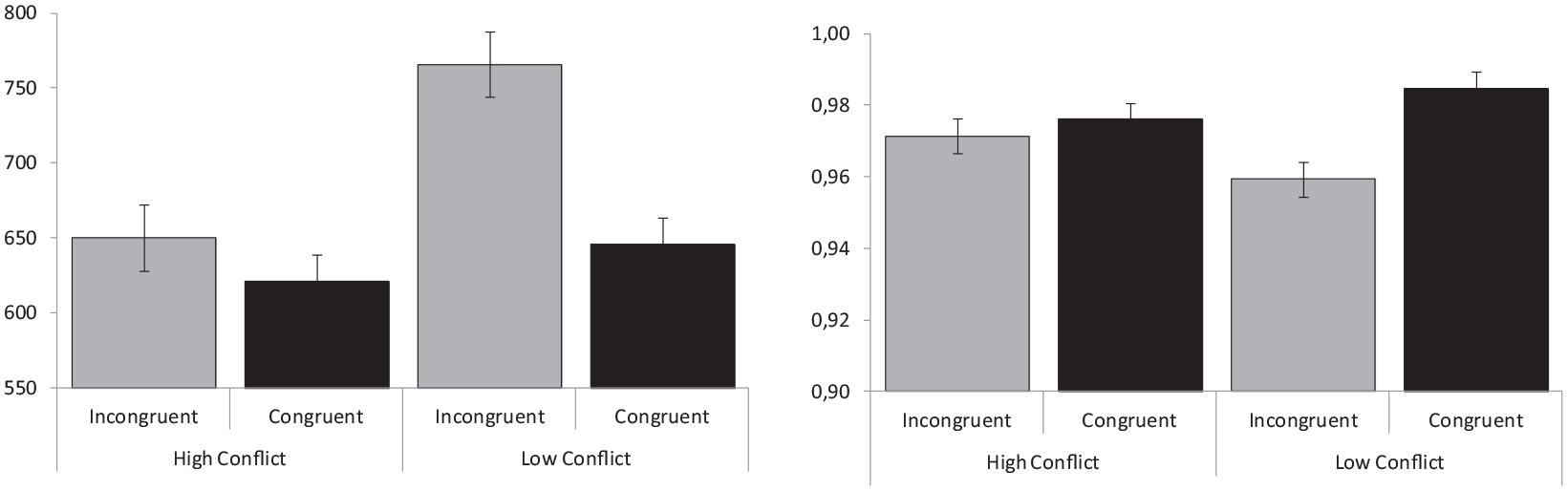
Congruency effects. Mean reaction times (left panel) and proportion of correct responses (right panel) represented separately for congruent and incongruent trials in conditions of low versus high conflict.
Contingency learning effect
RT and proportion of correct responses to neutral trials involving either contingent or non-contingent word–colour pairs are represented in Figure 3, separately for participants trained under high- or low-conflict conditions. The ANOVA conducted on RT with contingency (contingent vs non-contingent trials) as a within-participants factor and with conflict level as a between-participants variable showed a non-significant trend to produce slower responses in the low-conflict condition (M = 617 vs 662 ms), F (1,54) = 3.82, p = .06; ηp2 = .07, and a significant effect of contingency (M = 635 vs 644 ms), F(1,54) = 7.80, p = .007; ηp2 = .13, but not a contingency × conflict-level interaction, F(1,54) = 2.22, p = .14; ηp2 = .04. The analysis conducted on the proportion of correct responses showed no significant main effects of contingency, F(1,54) = 0.05, p = .94; ηp2 < .001, or conflict level, F(1,54) = 2.97, p = .09; ηp2 = .05, and no evidence of an interaction between them, F(1,54) = 1.11, p = 30; ηp2 = .02.
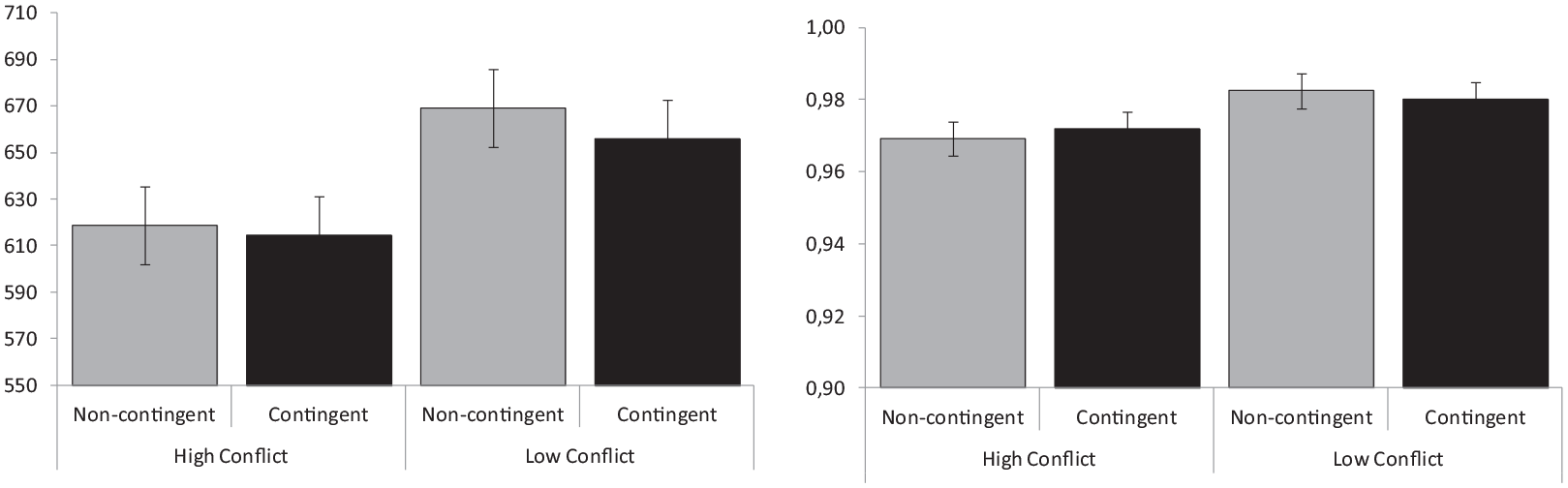
Contingency effects. Mean reaction times (left panel) and proportion of correct responses (right panel) represented separately for contingent and non-contingent trials in conditions of high versus low conflict.
Recency effects
As discussed above, recency effects could be present in this paradigm and can improve performance in response to the most frequently repeated distracter–target ensembles, including high contingency trials and the congruent trials most repeated in low-conflict conditions (see Giesen et al., 2020; Schmidt et al., 2020 for discussion). To explore the extent to which contingency effects could actually depend on such recency effects, and whether they could also be partially responsible for the observed list-wide proportion of congruency effects (see Bugg, 2014), we compared responses to all the trials in terms of whether the response required in that trial matched or not with the response required in the most recent previous episode involving the same distracter. As a way to estimate the impact of the distance with respect to that previous episode, we also considered this factor, including only distances from one to six previous episodes, so as allow for a minimum number of trials contributing to each cell (the average number of trials within each of these cells was 28, ranging from 7 to 72).
Two ANOVAs conducted on RT and proportion of correct responses, with conflict level (high vs low) as a grouping factor, and using distance to the previous presentation of the same distracter (1–6) and matching versus not matching response as repeated measures, showed significant main effects of matching F(1,54) = 160.20, p < .001; ηp2 = .75 and distance F(5,270) = 29.69, p < .001; ηp2 = .36 on RT, but not on the proportion of correct responses, F(1,54) = 3.13; p = .08; ηp2 = .06, and F(5,270) = 2.04, p = .09; ηp2 = .04, respectively. These analyses showed also a significant matching × distance interaction in both RT, F(5,270) = 92.44, p < .001; ηp2 = .63 and proportion of correct responses, F(5,270) = 4.45, p = .001; ηp2 = .08. As can be seen in Figure 4, this pattern indicated that responses were generally faster and more accurate when they matched the response required in the most recent previous presentation of the distracter, and that the impact of that matching was stronger when the previous appearance occurred right on the immediately previous trial. Bonferroni-corrected comparisons showed that the advantage in RT in favour of matching trials was significant not only when the distracter appeared on the immediately past trial (193 ms, p < .001), but also when it appeared in trial n–2 (18 ms, p = .02), n–5 (17 ms, p = .04), and n–6 (16 ms, p = .03). As for the effect observed in percentage of correct responses, this was also significant when the matching trial appears in n–1 (.025, p < .001) and n–3 (.011, p = .02). The main effect of conflict level was not significant neither for RT, F(1,54) = 3.32, p = .07; ηp2 = .06, nor for the proportion of correct responses, F(1,54) = 2.20, p = .14; ηp2 = .04, but there was a significant conflict level × matching interaction in the measure of RT, F(1,54) = 8.07, p = .006; ηp2 = .13.
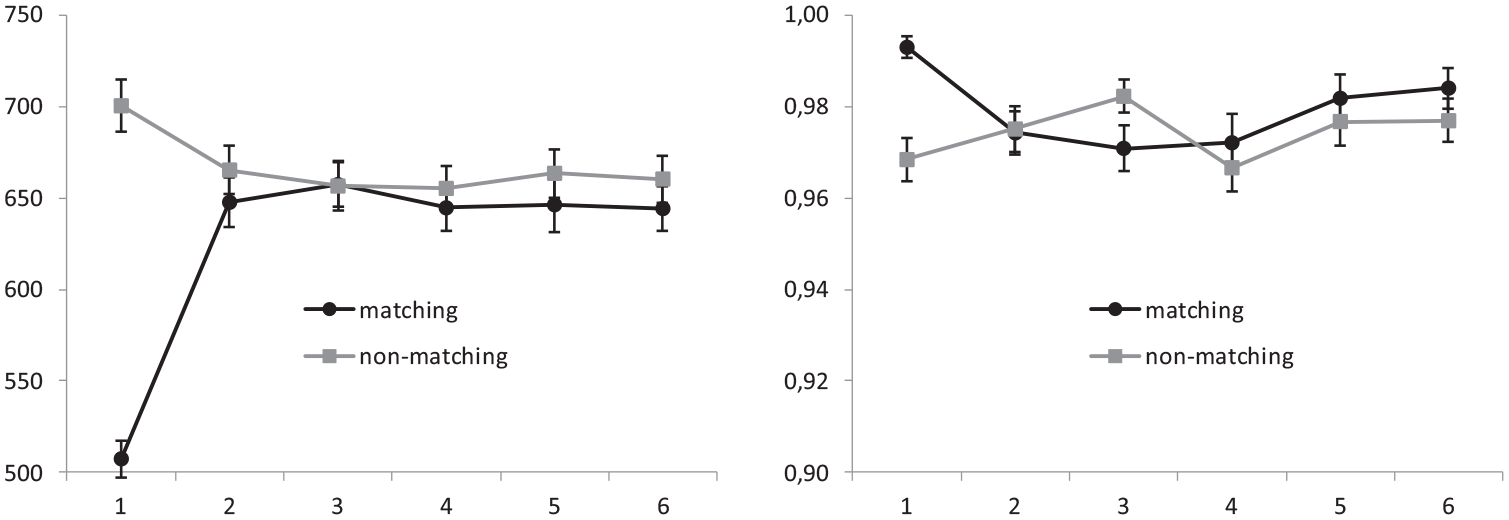
Proportion of congruency and contingency learning effects without the nearest recency effect
Given the strong effects provoked by the most recent matching between a given trial and the last previous occurrence of the same distracter, and given that such matching trials would tend to occur more often (1) in the congruent trials arranged in the low-conflict condition and (2) in the contingent distracter–target pairs arranged in both groups, we deemed it important to reassess those effects exclusively on those trials which were not differentially affected by the recency factor. Thus, we recomputed both list-level proportion of congruency and contingency effects exclusively on trials requiring a different response from that demanded by the most recent occurrence of the distracter (see Figure 1).
The average RT and the proportion of correct responses obtained in this subset of trials are represented in Figure 5, separately for congruent and incongruent trials, and for high-versus low-conflict conditions. As can be seen in the figure, the removal of the recency effects did not affect the pattern of results involving the list-wide proportion congruency effect, in that the effect of congruency was again significantly stronger in the low-conflict group as compared with the high-conflict group. This interaction was again significant in the analysis of RT, F(1,54) = 17.93, p < .001; ηp2 = .25, but not in the analysis of the proportion of correct responses F(1,54) = 1.38, p = .25; ηp2 = .03. Importantly, the same analysis conducted on the effects of contingency learning showed that the removal of the impact of recent matching did completely wipe out the effect of contingency from both the measures of RT and proportion of correct responses. Neither the main effects of contingency, F(1,54) = 0.01, p = .93; ηp2 < .001 and F(1,54) = 0.18, p = .68; ηp2 = .003, for RT and proportion of correct responses, respectively, nor the contingency × conflict-level interactions (F(1,54) = 1.62, p = .21; ηp2 = .03 for RT and F(1,54) = 0.60, p = .44; ηp2 = .01, for proportion of correct responses) were even close to significance in these analyses.
Proportion of congruency and contingency effects without the nearest recency effects. Mean reaction times (left panel) and proportion of correct responses (right panel) computed selectively on trials that required a response different from that required on the previous occurrence of the distracter, represented separately for congruent and incongruent trials (upper panels) and for contingent and non-contingent trials (lower panels), in conditions of high versus low conflict.
Given the lack of contingency learning effects after removing the impact of the most recent appearance of the distracter, we were especially interested in estimating the likelihood of the null hypotheses. Therefore, we conducted a series of Bayesian repeated-measures ANOVAs, using JASP Version 0.14 (Computer Software; JASP, 2018, see Wagenmakers et al., 2018). The Bayesian mixed ANOVA conducted on the measures of RT, using conflict level as a between-subjects factor and congruency effects without the nearest recency factor as a repeated measure, indicated that the data were best represented by a model including both main factors plus the conflict level × congruency interaction. The Bayes factor (BF10) corresponding to that model was 1.40 × 109, indicating decisive evidence in favour of this model, compared with the null model. The BF10 in favour of the relevance of the interaction effect, on top of the two main effects, also amounted to 183.29. An analogous ANOVA conducted on the proportion of correct responses indicated that the best model in that case was the model including exclusively the main effect of Congruency, which produced a BF10 = 3.97, indicating moderate evidence in favour of this effect.
As for the effect of contingency, the analysis conducted on the measure of RT showed that the data were best modelled by taking into account exclusively the effect of conflict level. The Bayes factor for this effect was slightly better than the null model BF10 = 1.26, and thus showed anecdotal evidence in favour of this effect. Importantly, the BF10 in favour of the relevance of the contingency effect was 0.11, compared with the null model, thus showing moderate to strong evidence against that effect. The analysis conducted with the measure of proportion of correct responses showed that the best model to account for these results was the null model, and that the BF10 in favour of the relevance of contingency was 0.21, showing moderate evidence against the existence of this effect.
Discussion
The goal of this study was to assess whether conflict interacts with contingency learning in the context of a Stroop task. We manipulated conflict as the proportion of congruency in a set of inducer trials, and estimated its impact on the learning of contingencies established on independent diagnostic trials, in which a neutral word was most frequently paired with each one of the four possible colour responses. The results clearly showed that the amount of conflict manipulated between groups produced a significant difference in congruency effects, consistent with the list-wide proportion congruency effect, even in conditions in which the conflict trials were interspersed with other neutral trials. However, they showed no evidence consistent with the claim that the overall amount of conflict experienced over such interference task modulated the strength of the associative learning produced between the neutral words and the colour responses associated with them. Indeed, even though the analysis of contingency learning appeared to show a significant main effect of contingency, the removal of the effects attributable to the episodic retrieval of the most recent response given to that distracter did completely remove the effect of contingency.
In what follows, we will discuss on what we consider the three main issues underlying our goal, namely (1) the extent to which the procedure described in this study was appropriate to manipulate the overall level of conflict, and therefore to analyse its impact on associative learning; (2) the extent to which the present results, that provide no evidence of associative learning between a set of neutral words and the colour responses associated with them, can be made compatible with other recent studies dealing with the same issue; and (3) the ways in which the present results contribute to the ongoing debate on how learning and memory processes may affect the dynamics of cognitive control.
Proportion of congruency as a manipulation of conflict
The most relevant precedent of the present study (Whitehead et al., 2018) manipulated conflict in a more drastic and specific way, comparing learning between groups that responded exclusively to conflict or to non-conflicting, neutral trials. This had the obvious advantage of allowing a stronger manipulation of conflict, but it came with a number of complementary drawbacks, as it only allowed the authors to assess learning on the very same trials on which conflict was being manipulated, and it compared learning of two different types of associations, ones involving the target colours and a set of neutral words, while the other related the same colours with a set of closely related words, which were already associated with an incompatible colour (cf. Schmidt, 2019; Schmidt et al., 2020). By contrast, separating the trials that induce a given amount of conflict from those in which learning was assessed allowed us to test learning of the same associations in both cases, under conditions differing exclusively in terms of the overall amount of conflict experienced in the surrounding trials. One might claim that the conflict conditions included in the present manipulations might have been just too weak to produce a significant impact on the amount of learning, and that stronger effects could be obtained by comparing more extreme conditions, for instance, using groups exposed either to 100% congruent or 100% incongruent ensembles over the inducer trials. We believe that this manipulation might constitute a valuable extension of the present study, but it would not allow us to assess the congruency effects produced over the inducer trials, as a way to make sure that the conflict manipulation was efficient to modulate the amount of cognitive control exerted by the agents. By contrast, arranging two unbalanced proportions of congruency, such as those used in the present experiment allowed us to confirm that the manipulation of conflict level was effective to produce the expected modulation of list-level congruency effects (Logan & Zbrodoff, 1979), but that still this difference in the amount of control did not affect the learning of the contingencies established between the neutral words and their assigned colours and responses.
Another important issue in this regard concerns whether the list-wide proportion congruency effect observed in this experiment really reflects the impact of the level of conflict on the up-regulation of control, as suggested by the model of conflict monitoring (Botvinick et al., 2001), or whether it might reflect the impact of episodic factors analogous to those posited to account for the effects of contingency learning. Indeed, just as most neutral word–colour pairings would tend to reproduce the ensemble presented on the prior occurrence of the distractor, episodic repetition would also tend to occur for the congruent trials in the conditions of high proportion of congruency, thus improving responses to such congruent trials, and hindering responses to the incongruent trials presented in low-conflict conditions (cf. Giesen et al., 2020). To assess whether the list-wide proportion congruency effect would still be observed after this episodic factor was filtered out, we conducted a reanalysis of congruency effects over the inducer trials, considering only trials in which the previous occurrence of the distractor had been paired with a non-matching colour, and confirmed that the manipulation of high versus low conflict was still effective in those trials to produce the expected difference in congruency effects depending on the overall level of conflict.
Contingency learning or episodic effects
The analyses of the diagnostic trials indicated that responses were significantly faster for high-frequency pairs, thus apparently showing a contingency learning effect. However, this effect was smaller than that found in the study by Whitehead et al. (2018), and subsequent analyses conducted to distinguish between the incremental effect of associative learning and the impact of the most recent previous episode involving the same distracter indicated that no effect of learning survived the removal of such episodic factor. This result highlighted the importance of episodic factors in experience-based modulation of congruency effects (Bugg et al., 2015), which can account for the repeated finding that so called “learning” effects arise early in practice, and are already present over the first block of training (Schmidt et al., 2007).
Alternatively, one might adduce that gradual and associative learning effects might arise later in training, but that episodic effects might be larger at the beginning of practice, thus blurring any difference between groups when both effects are collapsed into a single measure. To assess whether the effect of associative learning, and perhaps a possible interaction with conflict level, may arise more strongly over the final blocks, we reproduced the analyses of contingency learning over the final blocks, following Whitehead et al.’s (2018) lead, who analysed participants’ performance after their second training block. However, the analyses restricted to Blocks 3–5 produced results analogous to those yielded in the analyses conducted over the full training period, thus reinforcing the conclusions that contingency learning between words and colours was not affected by the level of conflict, and that most, if not all, of these effects were driven by the impact of the most recent episode involving the same distracter.
One must notice, however, that in the present procedure, we were not able to show any consistent evidence of associative learning after five blocks of training, in conditions in which participants responded to a 62.5% of high contingent trials from the diagnostic stimuli, a percentage that was exactly in between the two conditions of 50% and 75% arranged in the experiments by Whitehead et al. (2018). Interestingly, they reported their contingency effects on a block by block basis, and showed that these effects were already obtained over the first block, and grew progressively with training, especially in the groups exposed to neutral words and to larger proportion of contingent trials. It is possible, therefore, that mixing neutral words with colour-related words within the same procedure could have made the neutral word–colour associations more difficult to observe in the present conditions, as compared with those arranged by Whitehead et al., in which larger learning effects were obtained even with lower levels of contingency. This could resemble a version of the “fan effect” (Anderson & Reder, 1999) often found in associative memory, indicating that the learning of a particular association may depend on the presence of competing associations involving the same target.
The interaction between conflict and memory and/or learning
The absence of contingency learning observed in this study, together with the persistent effects of the most recent previous episode involving a distracter, can be seen as informative on the current debate on how learning and memory modulate conflict effects, and vice versa. It is important to note that the argument about the role of conflict on both learning and memory was raised to justify both observed improvements in these processes as a result of conflict (Krebs et al., 2015; Ramm et al., 2021; Rosner et al., 2015; Verguts & Notebaert, 2008) and a decrease in learning in conditions of higher conflict (Whitehead et al., 2018). Other studies have failed to substantiate either of these two opposing claims (Ortiz-Tudela et al., 2017, 2018), even under conditions in which the trained items were repeatedly presented in either congruent or incongruent contexts (Jiménez et al., 2020).
In Jiménez et al. (2020, Experiment 2), for instance, participants responded to the gender of faces which were repeatedly presented with an overlaid word that could be either congruent or incongruent with the required response (i.e., the words male or female). As in the present experiments, the authors found clear evidence indicating that the repetition of distracter–target pairs speeded up performance, producing larger congruency effects for those faces which were most frequently paired with a congruent distracter, relative to those faces which were mostly associated with an incongruent distracter. However, the authors found no differences in memory performance between both types of faces, thus suggesting that responding to an episode prepares participants to respond more efficiently to the reoccurrence of the same episode, but that it does not lead to a better encoding of the item.
Similarly, the present results show that responding to a distracter–target ensemble in a word–colour Stroop task improves participants’ responses to an identical episode the next time it arises, but it does not produce an enduring association between distracter and response that could survive the appearance of a single extinction trial. The observed pattern of results could be better explained in terms of a temporary binding between distracters and targets or responses, rather than as the effect of a gradual and cumulative learning process that associates distracters with their most frequently associated responses. This analysis is similar to a recent proposal made by Giesen et al. (2020), who emphasised the impact of previous bindings between distracters and responses. However, in contrast to their analysis of the extent to which habit acquisition could be taken as the result of an accumulation of previous bindings, our purpose was to distinguish between the episodic effect of the single, most recent, distracter–response binding, and the gradual effects that might be derived from a weighted aggregation of such bindings. In common with the present results, Giesen et al. also reported evidence from a similar word–colour Stroop task, suggesting that the effect of contingency learning did completely vanish after taking into account the episodic effect derived from the last prior occurrence of the distracter (but see Schmidt et al., 2020, for a report of conditions in which previous occurrences seem to exert additional effects).
Even though our results highlight the importance of previous experience on the adaptive regulation of cognitive control, they point to the relevance of episodic factors that go beyond the impact of the immediate previous trial (i.e., beyond earlier accounts based only on repetition priming, such as those put forward by Hommel et al., 2004 and Mayr et al., 2003), to encompass longer term modulations of cognitive control accounted for in terms of the reinstatement of conditions experienced in more distant trials, rather than in terms of the overall statistics aggregated over a series of previous trials (see Aben et al., 2017; Dey & Bugg, 2021 for discussions on the number of previous trials which are taken into account in computing such statistics). We believe that this emphasis on the impact of the last episode resonates well with the three-stage view of memory consolidation proposed by Fiebig and Lansner (2014), where the authors incorporate working memory to the traditional two-stage complementary learning systems proposed, among others, by McClelland et al. (1995). Fiebig and Lansner assume that, in addition to the slow learning mechanisms implemented through neocortical structures, and the index and offline reactivation roles played by the hippocampus and surrounding medial temporal structures, prefrontal cortex structures provide a faster short-term indexing role, that reinstates past episodes, either upon reflection or repetition, as a necessary preliminary step to establish the longer lasting and sparse representations held in the medial temporal lobe, that would later allow long-term consolidation of learning via offline reinstatement over sleep and rest periods. According to this view, neocortical learning would be adaptively very slow, but hippocampal coding, rather than being conceived as one-shot learning, would depend on the accumulation of several repetitions of short-term episodes, and thus would be mediated by the function of the prefrontal cortex. Moreover, given that the prefrontal structures work basically as an index that improves the reinstatement of neocortical representations of previous episodes, this framework would predict strong interference coming from the most recent episodes, and that arbitrary ensembles would require more practice than more semantically related associations to produce a longer lasting code that could be transferred and maintained in the medial temporal lobe. This could explain why contingency learning effects independent from the episodic factor were observed exclusively for the inducer items, but not for arbitrary pairings between colours and neutral words.
In sum, the present study has shown that, even though contingency learning effects are ultimately equivalent to the accumulated and weighted effects of a collection of previous episodes (cf. Giesen et al., 2020), the sensitivity of the control dynamics to the manipulation of episodic factors, such as the last prior occurrence of a distracter (Schmidt et al., 2010), may be a useful tool to distinguish between the overall effect of aggregated learning processes versus the punctual impact of recent memory episodes.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.