
Abstract
It is important for organisms to notice signals of opportunities (i.e., chances for performance-dependent reward) and dangers (i.e., performance-dependent risks of loss). Attentional biases towards opportunity and danger signals should therefore be functionally valuable. By contrast, the functional value of attentional biases towards signals of performance-independent (i.e., uncontrollable) rewards or losses is not obvious. The present study compares attentional biases towards positive and negative stimuli, depending on whether the stimuli signal performance-dependent or performance-independent reward or loss. Specifically, we induced colour-valence associations before engaging participants in an additional-singleton task that measures attentional bias. In the valence-induction phase, one colour signalled a potential reward, and another colour signalled a potential loss; importantly, in one group, rewards and losses were performance-dependent, whereas in another group, they were performance-independent (i.e., seemingly random). In the subsequent additional-singleton task, we found increased additional-singleton effects for colours associated with performance-dependent rewards and losses (i.e., opportunities and dangers). If, however, rewards and losses were performance-independent, the singleton effect was enhanced only for reward but not loss stimuli.
There is a lively debate on the attention-capturing qualities of affectively connoted stimuli (for a review, see Anderson, 2016; Yiend, 2010). The interest in this issue stems from different backgrounds. One line of work in this area has its roots in anxiety research (e.g., Williams et al., 1988) and general evolutionary notions of threat adaptation (e.g., Öhman & Mineka, 2001). Therefore, the initial research focus was on attentional qualities of threat- or danger-related stimuli. More recently, researchers have also explored whether complementary stimuli that signal chances or opportunities have comparable attention-grabbing properties (e.g., Brosch et al., 2008; Pool et al., 2016).
A danger is characterised by the potential occurrence of an aversive event, where the negative outcome, however, might be avoided by adequate action. An opportunity is characterised by a potential gain, where again the—in this case positive—outcome depends on adequate action. Thus, the rationale for assuming an attentional bias for opportunity- or danger-signalling stimuli is directly linked to the ability to control a situation if immediate action is taken, based on a fast assessment of the situation.
Given this consideration, the question arises whether stimuli that signal positive or negative outcomes that are not under one’s control will also capture attention, as no immediate action is necessary in these cases. This is not a trivial question. On one hand, one might argue that our cognitive system is tuned to prioritise valent stimuli in general. This tuning might have evolved because in our natural ecology stimuli that signal danger or opportunity are a major subset of valent stimuli, and the costs of prioritising the remaining stimuli as well (i.e., those that signal positive or negative outcomes that are not under our control) are low (see, for example, Pratto & John, 1991, who made the argument that early attentional processes might be rather undifferentiated). On the other hand, one might argue for biases that already incorporate the control potential in the service of action preparation and motivation regulation (Brandtstädter et al., 2004; Rothermund, 2011).
In this line of research, valence-connoted materials are typically varied quasi-experimentally. For example, angry faces or images of dangerous animals are used to signal threat, happy faces or images of babies are used to signal opportunities. Using inherently valent stimuli in this manner, however, means that the difference between signals for controllable and non-controllable situations can only be imperfectly operationalised.
A second line of work that has explored the attention-capturing qualities of affectively connoted stimuli stems from learning theory. Here, the focus is dominantly on reward-related stimuli (e.g., Anderson et al., 2011b; for a review, see Anderson, 2016), although some studies have also explored whether stimuli that signal loss or punishment have comparable attention-grabbing properties (e.g., Schmidt et al., 2015; Wang et al., 2013). In this line of research, valence-connoted materials are typically varied experimentally. This means that stimuli (e.g., colours) are first associated with reward (or punishment) in an acquisition phase, before being used in a subsequent test phase, where their attentional characteristics are assessed. This approach allows for the experimental manipulation of participant control by providing reward or punishment that either is or is not contingent on participant behaviour. This is the approach used in the present experiment.
First, however, we provide a brief review of research on reward and punishment with special focus on the instrumental contingency issue (for a more general recent review, see Watson, Pearson, Wiers, & Le Pelley, 2019). There are three categories of relevant studies: first, studies exploring solely the role of reward, typically implemented as points that are directly converted into monetary gain; second, studies on punishment, typically utilising Pavlovian conditioning procedures; third, studies directly comparing reward and punishment, again typically using monetary gains and losses. Most of these studies use a two-phase design, that is, a valence-acquisition phase followed by a test phase assessing attentional bias for stimuli, which typically no longer involves rewards or punishments. Attentional bias is often measured with variants of visual search or cuing paradigms. For example, in the additional-singleton version of visual search (Theeuwes, 1992), participants search for a unique shape among several coloured stimuli (e.g., a circle among diamonds). Although colours are task-irrelevant, responses are slower if a distractor (i.e., the “additional singleton”) is presented in a reward-associated colour (indicating attentional capture; Anderson et al., 2011a, 2011b). In the cueing variant, participants categorise a target that unpredictably appears left or right of fixation; the target is immediately preceded by two cues that are presented at both possible target locations. Target responses are faster if the cue presented at the target location has a reward-associated colour, again suggesting attentional capture by reward. We hasten to add that there are alternative interpretations, most prominently explanations in terms of increased attentional dwelling on stimuli (i.e., difficulties disengaging attention). To keep this introduction succinct, we postpone discussion of this issue to the “Discussion” section. In the following overview, we use the more general term attentional bias.
Reward studies
Anderson and colleagues (Anderson et al., 2011a, 2011b; Anderson & Yantis, 2012, 2013 for a review, see Anderson, 2016) have published several experiments on attentional bias for reward signals. In their paradigm, participants complete visual search trials featuring several coloured circles in an acquisition phase. Two colours (e.g., red and green; exactly one of which was present on each trial) defined the target, which had to be categorised based on a non-colour feature (e.g., location). For one of the colours, 80% of correct responses attracted a substantial reward and 20% attracted only a small reward; for the other colour, the assignment was reversed. Reward was moderately dependent on participant behaviour because there was a response deadline of 600 ms; slower responses and erroneous responses were never rewarded. We therefore call this form of reward schedule “moderately contingent.” This paradigm produces replicable attentional bias towards reward-associated stimuli (see Marchner & Preuschhof, 2018, for a replication from another lab). Kim et al. (2021) transferred the paradigm into the auditory domain and found comparable effects.
Failing and Theeuwes (2014) used the same two-phase logic (i.e., acquisition phase followed by a reward-free test phase) but using a version of the exogenous cueing paradigm (see above). Failing and Theeuwes (2017) analysed reward effects in a version of the additional-singleton paradigm that, however, collapsed acquisition and test phases (in the test phase, a specific distractor colour indicated high or low reward for the target response). In both studies, the authors found attentional bias towards reward stimuli with a low-contingent reward schedule.
In another study, Pool et al. (2014) used Pavlovian conditioning to associate a geometric figure (CS+) with chocolate odour (US) while another geometric figure (CS−) was associated with odourless air. In an exogenous cueing task, the CS+ and CS− served as sudden onset cues, which resulted in cueing effects (i.e., faster responses to subsequent targets that were presented at the same location as the cue compared with targets presented at the opposite location). However, the cueing effect was larger for CS+ compared with CS−. In a second experiment, the authors introduced a sensor-specific satiation procedure (i.e., eating chocolate) after the conditioning task. This reward devaluation erased the attentional bias to the CS+.
In contrast to the randomised reward allocation, Le Pelley, Pearson and colleagues (Le Pelley et al., 2015; Pearson et al., 2015, 2020; Watson, Pearson, Chow, et al., 2019) implemented a more deterministic reward schedule using an oculomotor capture paradigm (Theeuwes et al., 1999). In these studies, participants were required to saccade to the shape singleton (i.e., the target) in a visual search display and fixate it. Stimuli were all grey except one distractor (for most of the trials), which was coloured in one of two colours. One designated colour signalled the availability of a large reward; the other one a low reward. Participants were told that rewards depend on how fast and accurately they respond. In fact, reward was cancelled if response times were above 600 ms and it was withheld if the first saccade went to the distractor instead of the target. This is a straightforward operationalisation of an opportunity signal, with a reward that is dependent on adequate action. Although it is counter-productive in this task to direct attention and eye gaze to the reward-associated distractor, more first saccades go to the high-reward colour than to the low-reward colour.
A recent study by Pearson and Le Pelley (2020) used this paradigm to vary instrumental contingency. They added a condition (“yoked”) in which omission of rewards was not determined by fixations on the distractor, but was determined by the omissions of a yoked participant from a second group (“omission”) of participants following the procedure in Le Pelley et al. (2015). The value-modulated attentional capture effect was found for both samples. (Indeed, it was even larger for the yoked group, indicating that the response relationship in the omission group leads to suppression of saccades towards the distractor.)
In sum, reward studies give a rather homogeneous picture: reward-related stimuli cause attentional bias and the contingency of rewards on participant behaviour does not seem to affect this process.
Punishment studies
As already noted earlier, studies investigating attentional bias towards punishment signals typically use Pavlovian conditioning procedures. Koster et al. (2004) used an aversive noise burst to condition one colour (CS+), whereas others (Anderson & Britton, 2020; Grégoire, Kim, & Anderson, 2020; Nissens et al., 2017; Notebaert, Crombez, Van Damme, et al., 2011; Schmidt et al., 2015; Wang et al., 2013, Exp. 3) have used electric shocks. The reinforcement schedules of all these studies (except Nissens et al., where shocks depended on performance) can be considered “no-contingency” schedules because presentation of the unconditioned stimulus (UCS) was not dependent on participants’ behaviour. Anderson (2017) used social feedback to associate colours with negative valence. In a training phase, one of two target colours was followed by an angry face on most trials (the “high negative” colour) and by a neutral face on the remaining trials, whereas this assignment was reversed for the second colour (the “low negative” colour). While the feedback schedule was not dependent on participant behaviour, participants were told that “the faces would ‘react to what happened on each trial’” (p. 3); thus, participants might have perceived a dependency (e.g., interpreting an angry face as an indication of a tardy response).
At first glance, all these studies found effects suggesting an attentional bias towards punishment signals. Closer inspection, however, reveals a more differentiated picture. To begin with the “non-contingent” studies, Schmidt et al. (2015) and Wang et al. (2013, Exp. 3) used the additional-singleton paradigm in the test phase and found increased response times if the CS+ colour was present (as distractor) compared with the presence of a CS− colour. Koster et al. (2004) used a version of the cueing task with only one cue (which was CS+ or CS−). The cueing effect (i.e., faster responses to the target stimulus if it appeared at the location of the cue rather than the opposite location) was significantly larger for the CS+ colour compared with the CS− colour, which can be interpreted as attentional capture by an aversive stimulus. However, as acknowledged by the authors, one detail makes this result ambiguous: participants had to categorise the location of the target. Thus, the cue location is either compatible or incompatible with the response, and cueing (i.e., attentional) effects might be conflated with response priming effects (see also Imhoff et al., 2019). Notebaert, Crombez, Van Damme, et al. (2011) used CS+ and CS− targets in a visual search task. They found that search efficiency was enhanced for aversive targets but that there was no “pop-out” (i.e., search slopes were not flat in case of an aversive target colour, a result which would have been more in line with the attentional capture hypothesis).
In his social feedback study (technically a “non-contingent” study but potentially featuring a perceived response dependency; see above), Anderson (2017) found the distractor effect of the “high negative” colour in a subsequent search task to be numerically larger than the effect of the “low negative” colour; however, the difference was not statistically significant. (Nevertheless, a second experiment yielded no distraction by neutral colours. The cross-experiments analysis indicated a significant difference between “high negative” and “neutral.”)
Nissens et al. (2017) used the oculomotor capture paradigm. Stimuli were coloured in different colours and one designated (distractor) colour signalled the possibility of an electric shock that could be prevented by a prompt saccade towards the target—a straightforward operationalisation of a danger signal, where the harm can be avoided by adequate action. The danger signal appeared on half the trials; in the remaining trials a designated “safe” colour signalled the absence of shocks. Although it is counter-productive in this task to direct attention and eye gaze to the aversive distractor (because this slows the target response), more first saccades went to the danger colour than to the safe colour (with associated slowing of the target fixation). This result was corroborated by the study of Anderson and Britton (2020)—again with the oculomotor capture paradigm—who punished fixation of the CS + colour (with 50% probability). Nevertheless, more first fixations were on the CS+ compared with a control colour (CS−).
Kim and Anderson (2021) as well as Grégoire, Kim, and Anderson (2020) used an anti-saccade paradigm, that is, participants have to saccade to the opposite side of an onset stimulus. Too slow saccades (or saccades towards the stimulus) were punished by a shock if the colour of the item was the CS+ colour. The authors found evidence for attentional capture by CS+ in a test phase. Interestingly, the dependent variable in the test phase was again saccade latency. This manoeuvre allowed to pit attentional capture against S–R learning: If the colours of the training phase trigger the avoidance behaviour that was learned during the training phase, they should produce longer saccade latencies if presented as targets (if contrasted with the neutral colour). This was indeed observed for the CS− colour, but not for the CS+ colour. The latter result can be easily explained by two opponent processes: attentional capture towards the CS+ and a response tendency away from it. Accordingly, stimulus pairs consisting of CS+ and CS− colours produced faster responses towards a CS+ target than towards a CS− target. This result indicates that in paradigms with instrumental contingency the effects of a test phase are genuine attentional capture effects and not a by-product of the behaviour needed to avoid the negative event.
Grégoire, Kim, and Anderson (2021) replicated this effect with a smaller instrumental contingency than in the Kim and Anderson study: the number of shocks was fixed at 4 (out of 10) trials per block, that is, (a) bad performance beyond the 4th bad trial was not sanctioned and (b) in the case of a lower number of bad performance trials than 4, designated CS + trials were associated with shocks non-contingent on behaviour. 1
In sum, punishment studies show attentional bias effects as well. Roughly speaking, the effects can be found irrespective of instrumental contingency. Nevertheless, evidence is sparser compared with reward studies and some of the effects of the “no contingency” studies do not unequivocally address attentional capture.
Studies directly comparing reward and punishment
Some studies have directly compared reward and punishment, using monetary gains and losses to operationalised rewards and punishments in a comparable manner. The instrumental contingency of reward and punishment on participants’ behaviour varied considerably across these studies.
The study by Bucker and Theeuwes (2016) is explicitly of the “no contingency” type, as reward or punishment was independent of participant behaviour (even error trials could attract a reward). 2 The authors tested for attentional biases towards reward and punishment signals in an exogenous cueing paradigm, utilising the well-known effect that abrupt-onset cues capture attention if the cue-target asynchrony is short (i.e., 170 ms). This early capture effect was present in Bucker and Theeuwes’ study of whether the cue was neutral, reward-signalling, or punishment-signalling. With a longer delay (i.e., 960 ms), however, the cueing effect reversed for punishment cues, but remained for reward cues (and was null for neutral cues). Thus, following initial capture, attention is maintained at the location of reward cues, whereas attention is reoriented away from the location of punishment cues. Thus, a clear asymmetry was found for reward and punishment.
Gupta et al. (2016) introduced a further variant of the learning procedure, involving a gambling task: in each trial of the acquisition phase, participants had to choose one of two (neutral) faces to maximise their outcome. Unbeknownst to participants, one face pair was associated with gains, one pair was associated with losses, and one pair was a neutral control pair where choices had no consequences. Participants learned to prefer one face of the “gain pair” because it attracted a reward 80% of the time (and no consequence 20% of the time), compared with a reward rate of 20% for the other face; they also learned to avoid choosing one face of the “loss pair” because it attracted a loss 80% of the time (compared with 20% for the other face). The procedure can therefore be classified as “moderate contingency”—participants can maximise their gains through adequate choices, although chance still plays a considerable role. In test phase trials, a face was presented centrally, surrounded by six letters that always included either an “X” or an “N” as a target letter that had to be identified. There were distractor effects on target response times for gain and loss-associated stimuli (compared with neutral ones) in a low-perceptual-load context (i.e., the other five letters were all “O”s such that the target letter popped out), but only distractor effects from gain-associated stimuli in a high-perceptual-load context (i.e., the other letters were H, K, W, M, Z, necessitating a clear search effort). This suggests that attentional bias towards reward stimuli is rather robust, whereas attentional bias towards punishment stimuli seems to be more fragile.
A strong instrumental contingency was implemented in the experiments of Wentura and colleagues (Müller et al., 2016; Wentura et al., 2014, 2018). In a simple categorisation task, a specific colour signalled that a fast (and correct) response would lead to a points gain (i.e., ultimately a monetary gain), with no consequences for a slow (or erroneous) response. A second colour signalled that there would be no consequences associated with a fast (and correct) response, but that a slow (or erroneous) response would lead to a points (i.e., monetary) loss. Most important in the present context, the criterion for receiving reward or avoiding punishment was rather strict: to obtain reward and avoid punishment, participants had to respond fast and correctly, with a moving median of recent trials serving as the criterion of a fast response. Wentura et al. (2014) found increased additional-singleton effects for gain- and loss-associated distractor stimuli compared with control colours. Wentura et al. (2018) replicated this effect. 3 Müller et al. (2016) used the same rationale of performance-contingent gains and losses in a cueing task (dot-probe task) and found attentional effects for both gain- and loss-associated stimuli. Folyi et al. (2016) and Folyi and Wentura (2019) conducted similar experiments with auditory tones instead of colours as reward/punishment signals. The amplitude of the auditory N1 was increased for valenced compared with neutral tones in a test phase, indicating enhanced attention.
A study by Wang et al. (2013) implemented rather strong instrumental contingency as well. They used a pay-off schedule of +10 points for a correct response to the reward and −10 points for a correct response to the punishment colour; errors were penalised by −5 points (i.e., +5/−15 for error responses to reward and punishment colours, respectively). Points were converted into monetary reward. Thus, a good performance (i.e., a correct response) led to the larger reward (i.e., 10 instead of 5 points gain) in the case of the rewarded colour and prevented the larger punishment (i.e., the loss is only 10 instead of 15 points) in the case of the punished colour. Thus, insofar behaviour has a deterministic consequence, we can label the schedule as a “strong contingency” one. In a subsequent visual search task, both reward and punishment colours (of distractors) slowed down target responses compared with a third colour, which was newly introduced in the test phase and is therefore a suboptimal control. For that reason, a control group completed the acquisition phase without rewards and punishments. In this group, the two colours used in the acquisition phase also produced an increase in response time in the test phase (compared with the novel colour); however, the increase was significantly reduced compared with the costs observed in the experimental group. Thus, in conclusion, this study showed symmetric attentional effects for reward and punishment.
Finally, we should mention two studies that went a somewhat different route. Becker et al. (2020) argued that in the study by Wentura et al. (2014), attention to a loss-associated distractor was actually rewarded because a fast and correct response prevented the loss. (Of course, this was the basic intention of Wentura et al., because they wanted to explore the attentional processes towards danger signals.) In a training phase, participants had to find a target colour circle in a visual search display and to categorise the line within the circle. There were three target colours: for the reward colour, correct responses were rewarded with US$0.10 in 80% of trials, but the counter was reduced by US$−0.10 in the remaining 20% of trials. For the punished colour, the payoff schedule was reversed. Responses to a third colour were not associated with payoffs. Most important, error trials and no responses within a deadline of 800 ms had no consequence in terms of payoffs. Consequently, during the blocks of the training phase, participants learned to increase the rate of correct responses for the reward colour from initial (app.) 55% to 80%. The corresponding rates for the punishment colour remained in the range of app. 53% to 57%, partly due a largely increased rate of timeout trials. That is, the variation of reward and punishment was now confounded with the type of rewarded behaviour. In the test phase (without rewards and punishments), participants have to search for a form singleton in an array of coloured items. Response times are recorded as a function of the absence or presence of one of the colours from the training phase and in the latter case the type of colour (i.e., rewarded, punished, or neutral). Interestingly, it was only the rewarded colour that prolonged responses in the test phase (i.e., showed attentional capture). With regard to the instrumental contingency issue, we can state that both reward and punishment were highly contingent on participants’ behaviour.
Finally, Grégoire, Britton, and Anderson (2020) wanted to show that attentional capture can be suppressed, given specific task requirements. In the oculomotor capture paradigm (see above), participants were instructed that rewards (i.e., money earnings) were more likely if their response (i.e., fixation of the form-singleton target) was fast and accurate and that punishments (i.e., shocks) were more likely if their response was slow. One colour-singleton distractor predicted the (infrequently realised) possibility of receiving a reward and another the (infrequently realised) possibility of an electric shock. In a test phase (without rewards and punishments); first saccades were less often directed to both reward and punishment colours compared with a neutral colour. Besides the fact that this study shed light on boundary conditions of reward/punishment-related attentional capture, we can state that instrumental contingency was high and reward and punishment signals yielded comparable effects.
In sum, studies with high instrumental contingency (Grégoire, Britton, & Anderson, 2020; Müller et al., 2016; Wang et al., 2013; Wentura et al., 2014, 2018) have found symmetric effects with one exception: Becker et al. (2020) found attentional capture for reward but no for punishment (but at the cost of a confound, see above, that we will return to in section “Discussion”). The study with no instrumental contingency (Bucker & Theeuwes, 2016) and the one with moderate instrumental contingency (Gupta et al., 2016) found asymmetries (with reward cues producing the more robust attentional effects). To the best of our knowledge, no study directly comparing rewards and punishments exists that also manipulates instrumental contingency.
It also has to be acknowledged that comparability of studies is rather low, as different valence-induction methods and different attention paradigms were used. This is unfortunate because a test of the complete 2 (valence: negative vs. positive) × 2 (control: present vs. absent) design would be of utmost interest. From a functional point of view, control might be an important moderator of attentional processes: if one’s own actions determine whether gains can be achieved and losses avoided, allocation of attention to signals of opportunity and danger seems to be highly functional. If outcomes are outside of one’s control, however, it is difficult to mount a functional argument. Of course, reward-related attentional biases are well established (Anderson, 2013, 2016) and have a clear foundation in learning theory. From a functional perspective, however, preferably directing attention to unavoidable punishments makes little sense. If we leave the narrow frame of attentional capture experiments, there is indeed evidence for the moderation of early processing of negative stimuli by control potential (e.g., Brandtstädter et al., 2004; Notebaert, Crombez, Vogt, et al., 2011).
In the present study, we used the high-contingency paradigm of Wentura et al. (2014) to test the hypothesis that attentional bias to opportunity and danger is dependent on control. To this end, we used a yoked design: the specific rewards and punishments received by each participant completing the high-contingency procedure were also provided to a yoked participant, for whom response consequences were thus seemingly random and completely independent of their behaviour.
Overview
In line with the study of Wentura et al. (2014), this study consisted of two main tasks: the valence-induction task in which colours acquired value associations, and the additional singleton task, in which the impact of these associations on attention is evaluated. The crucial manipulation in the present study concerned the valence-induction task, which was varied in a between-participants design. One sample (hereafter the high-control sample) completed the exact acquisition task of Wentura et al. (2014). That is, they had control over the outcomes in the valence-induction task: a fast and correct response in a categorisation task yielded a points gain (in the presence of the positive colour, and to a lesser degree the neutral colour) and prevented a loss (in the presence of the negative colour, and again to a lesser degree the neutral colour). A fourth colour served as a no-go signal (hereafter irrelevant colour).
By contrast, participants of the other sample (hereafter the no-control sample) received the same loss/gain sequences as the yoked high-control participants. However, no-control participants had to respond only to the irrelevant fourth colour (i.e., the no-go signal in the high-control group), with no consequences for the points tally. Thus, no-control participants simply observed trials with the critical colours; they were instructed that trials could be “good” or “bad,” and that this was randomly determined (in fact, the status was determined by the performance of a yoked high-control participant). They were told that one colour (the positive colour) signalled a points gain in a good trial and no loss in a bad trial, whereas another colour (the negative colour) signalled a loss in a bad trial and no gain in a good trial. Finally, a third colour (the neutral colour) signalled a small gain in a good trial and a small loss in a bad trial. In both groups, the final points score was converted to monetary gain at the end of the experiment.
In the test phase, we applied the additional-singleton procedure. Here both high-and low-control groups followed the same procedure from Wentura et al. (2014) with one small change. The additional singleton version used by Wentura et al. (2014, 2018) was originally suggested by Theeuwes (1992): Its target (e.g., a circle) features a vertical or horizontal line that participants classify, while distractors (e.g., diamond shapes) feature slightly tilted lines that therefore do not fall into the two response categories (see Figure 1a). The task’s primary measure is the impact of the additional singleton and its acquired connotations on target response times (i.e., the additional-singleton effect). In the alternative version used here (see, for example, Hickey et al., 2006), the distractor symbols also contain a vertical or horizontal line (see Figure 1b). As a result, the additional singleton’s line is either congruent or incongruent to the target line, and one can additionally calculate congruence effects (i.e., the orientation of the singleton line matches or mismatches the orientation of the target line), which might be a function of the processing of the singleton.
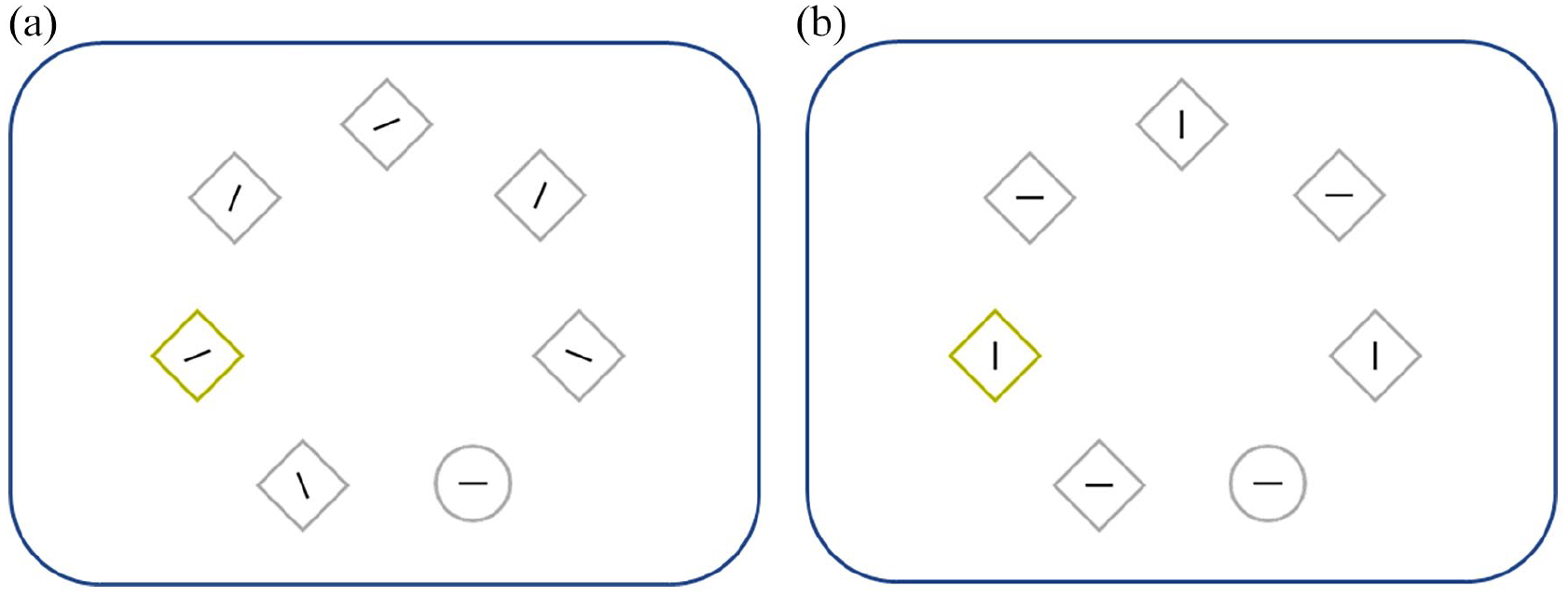
The two versions of the additional-singleton task: (a) only the target (i.e., the circle) contains a categorisable feature (as per the participants’ task; i.e., here: “horizontal”); (b) all stimuli contain a categorisable feature; depicted is an incongruent trial as the colour singleton contains a vertical line.
Finally, an evaluative priming task was administered. As in Wentura et al. (2014), it was added to show that colours had indeed acquired valence connotations during the induction phase.
Method
Participants
A sample of 96 students (48 women, 48 men) from Saarland University, Germany, participated in the experiment in exchange for M = €13.67 (range €6–€22; see the “Procedure” section). The median age was 23 years (with a range from 18 to 35 years). All subjects had normal or corrected-to-normal vision. The high-control and no-control samples initially had 48 participants each. However, the data of seven high-control participants and one no-control participant had to be discarded because of error rates >40% in the additional-singleton task.
For power calculations, we used the valence effect of dZ = .63 found in Wentura et al. (2014) as an anchor. To plan somewhat more conservatively, we proceeded from dZ = .50 (i.e., a medium-sized effect according to Cohen, 1988) for valence-related effects. That is, for the high-control sample we expected a replication of Wentura et al. (2014), that is, a significant contrast of positive/negative stimuli to neutral stimuli. 4 For the no-control sample, we aim to find possible differences between positive and negative stimuli of the same amount. Thus, both samples—that is, the high-control and the no-control sample—should be large enough to detect dZ = .50 with power 1 – β = .90 (α = .05); therefore, a sample size of N = 44 is needed. To account for some outliers (e.g., extreme error rates), we planned to recruit n = 48 for each sample. Unfortunately, size of the high-control sample reduced to n = 41 (see above); therefore, power for this sample reduced to 1 – β = .88. Sample sizes of n = 41 and n = 47 allow for the detection of between-participants effects of d = .61 (i.e., effects between medium, d = .5, and large, d = .8, according to Cohen, 1988). Power calculations were done with G*Power (Faul & Erdfelder, 2007).
Design
We employed a mixed design; the between-participants factor group (high-control vs. no-control) concerned the acquisition phase, whereas the within-participants factors singleton type (no singleton vs. irrelevant vs. neutral vs. positive vs. negative), target-distractor congruence (congruent vs. incongruent, i.e., same vs. different target and distractor orientation), and display size (five vs. seven stimuli) 5 concerned the additional-singleton task.
The assignment of colours to singleton type was counterbalanced across participants. We used the following scheme for counter-balancing and yoked assignment. The first 4 participants of each n = 8 sub-sample (i.e., participants 1–4, 9–12, and so on) were assigned to the high-control condition (1 participant per colour-balancing group). The remaining participants were assigned to the no-control sample and matched up with high-control participants, such that participant P received the same colour-assignment and the same win/loss sequence as participant P – 4 (e.g., Participant #5 was the yoked partner of Participant #1).
Materials
We used circles (diameter: 2.39° visual angle) and diamonds (diagonal length: 2.93° visual angle) as stimuli. The default colour of all items was a light grey (RGB values: 175, 175, 175). The colours for the valence manipulation were light red (255, 144, 144), light green (114, 201, 101), light blue (173, 173, 255), and yellow (180, 180, 0). Colours were matched for luminance. The background colour of the screen was white. During the valence-induction task, circles and diamonds contained either a black pentagon or a black five-pointed star. During the additional-singleton task, circles and diamonds contained either a horizontal or vertical black line.
In the evaluative-priming task, we used schematic smiley and frowny faces, which had the same size as the circles in the valence-induction and additional-singleton tasks. The targets of the evaluative-priming task were 12 affectively valenced German nouns (taken from Bermeitinger et al., 2012). Half of the nouns were positive (M = 6.69, SD = 0.10, on a scale from 1 [negative] to 7 [positive]), while the other half was negative (M = 1.89, SD = 0.28). The average word length was 5.5 letters (SD = 0.5, ranging from 5 to 6) for both positive and negative nouns.
Procedure
Upon arrival, participants received an endowment of €14 under the condition that they would invest the money as their stakes in a game. Participants were seated in an individual testing booth in front of a CRT computer screen, with a viewing distance of approximately 60 cm. All instructions were given via the screen. All participants performed three different tasks on the computer, the first one being the valence-induction task. Subsequently, the blocks of the additional-singleton task were alternated with additional blocks of the valence-induction task. The evaluative-priming task concluded the experiment.
Valence-induction task
We varied the specific nature of the valence-induction task between participants in a yoked design. The high-control group was told that they could win or lose money in a game, depending on their performance. In contrast, instructions for the no-control group emphasised that winning or losing money in the game was due to chance.
Specifically, the high-control task was identical to Wentura et al. (2014). Participants could win or lose €2 in each block of the game, depending on their final score in the block. On each trial, a coloured frame was presented at the centre of the screen; this was either a diamond or a circle. It contained a target object (either a five-pointed star or a pentagon), which subjects had to classify as quickly and accurately as possible via the “c” and “m” keys of a standard keyboard. The colour of the frame indicated the odds of winning or losing points. The “positive” colour indicated a possible gain of 20 points (in the case of a quick and correct response), with no risk of losing points in case of an incorrect or tardy response. The “negative” colour indicated a possible loss of 20 points (in the case of an incorrect or tardy response), with no opportunity for gaining points from a quick and correct response. A “neutral” colour signalled that participants could win 1 point for answering fast and correctly, and lose 1 point otherwise. Response speed was classified into “fast” or “slow” using a criterion based on the moving median of the last six trials. 6 Finally, a fourth “irrelevant” colour indicated that participants should not respond; in this case the task proceeded with the next trial after 1,500 ms. If participants did press a response key during that interval, the message “Please do not respond to items of this color” appeared; repeated responses triggered harsher reminders. Assignment of colours to conditions was counter-balanced across participants via a Latin square. Figure 2 shows an example sequence of trials.
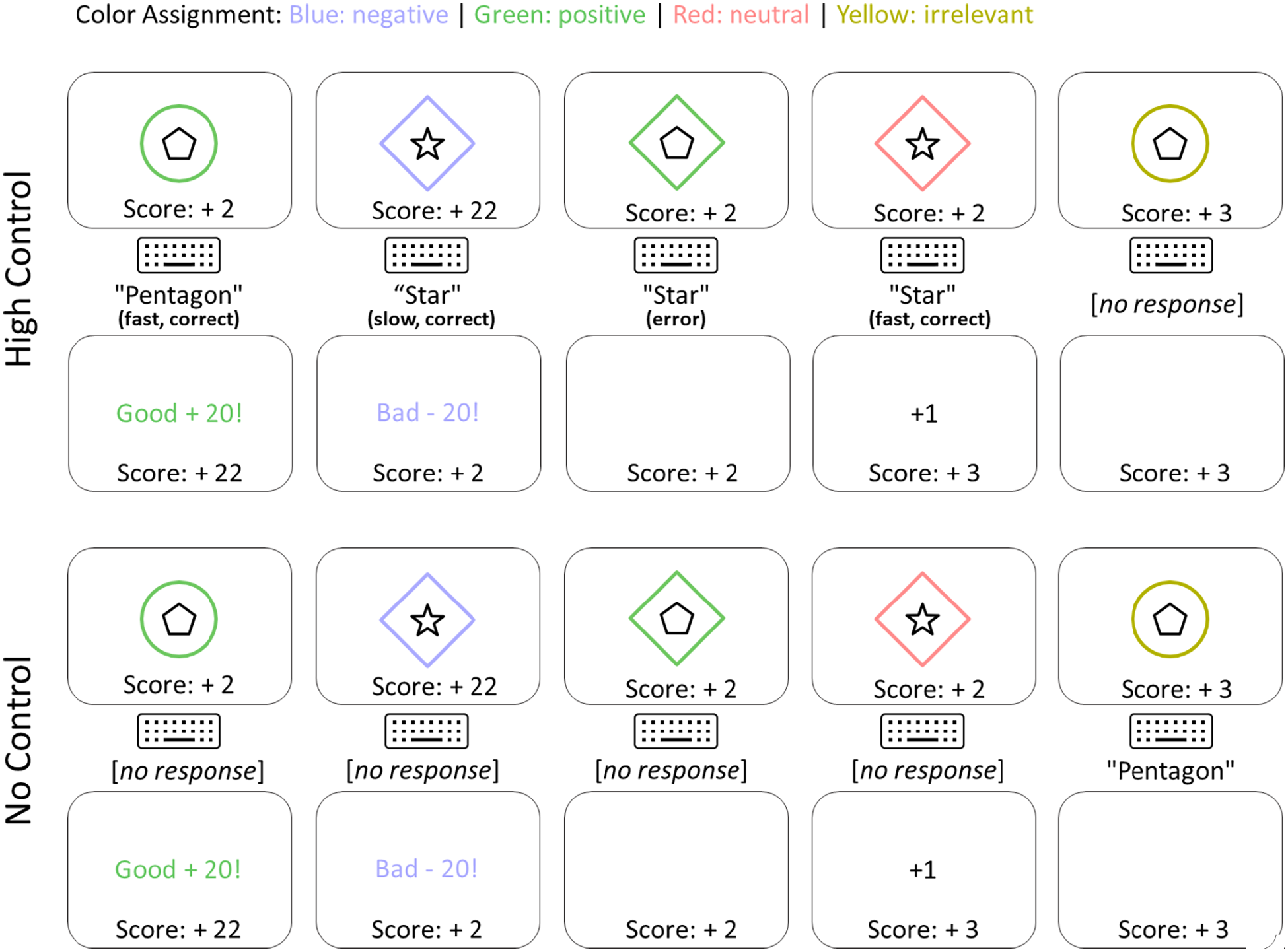
Examples trials of the training phase. Top: A sequence of stimulus screens (top), response (midst), categorisation (midst, in bold), and feedback screens (bottom) of the high-control sample. Bottom: The same sequence for the yoked participant.
At the beginning of each block, the score was set to zero. The current score was permanently shown on the bottom of the screen, and changes were accompanied by corresponding messages (i.e., “Good + 20!”; “Bad − 20!”; “+1”; “−1”; presentation time 1,000 ms). A positive score at the end of a block resulted in a €2 gain, a negative score resulted in a €2 loss. One block consisted of 48 trials (three repetitions of each combination of 4 colours × 2 shapes × 2 target categories).
Participants in the no-control group were instructed that they did not have any influence on the game score. Their task was merely to observe wins and losses in the positive, negative, and neutral colour conditions, and to classify the targets only if the frame had the fourth colour (i.e., the irrelevant colour of the high-control sample). Stimulus presentation was identical to the high-control sample. Specifically, no-control participants received the exact same visual input, including the sequence of stimuli, duration of presentation, and feedback for positive, negative, and neutral colours as their yoked partners in the high-control sample. In the case of the irrelevant colour (which was now a go signal), frame and target remained on the screen until a response was recorded. If a participant responded on a positive, negative, or neutral colour trial, the same feedback message was presented as was used in the no-go condition of the high-control sample.
The experiment began with a practice block of the valence-induction task, which did not result in monetary gains or losses. Subsequently, it was checked whether participants could remember the colour-dependent rules of winning and losing. Otherwise, the rules were repeated and understanding was checked again. Then, three blocks of the valence-induction task were administered, before participants completed alternating blocks of the additional-singleton and valence-induction tasks. In total, there were eight blocks of the valence-induction task, and six blocks of the additional-singleton task.
Additional-singleton task
In each trial of the additional-singleton task, either 5 or 7 shapes (circles, diamonds) appeared on the screen. The shapes’ midpoints were arranged with equidistant spacing on an imaginary circle centred at the middle of the screen. The diameter of this circle was 12° visual angle. Each shape contained a line that was either horizontal or vertical. On each trial, there was exactly one shape that deviated from the others (i.e., the shape singleton): either there were four/six circles and one diamond, or four/six diamonds and one circle. In addition, in half of the trials there was a colour singleton. While the default colour of the shapes was grey, colour singletons were either red, green, blue, or yellow. Colour- and shape singleton were never at the same location in a given search display. Participants were instructed to classify the orientation (horizontal, vertical) of the line in the shape singleton as fast and accurately as possible via the “c” and “m” keys, and to ignore the colour singletons. The precise sequence of events was as follows. At the beginning of a trial, a fixation cross was presented for a randomly chosen, variable time between 1,000 and 2,500 ms (varied in 250 ms steps). Subsequently, the fixation cross increased in size and remained on the screen for another 600 ms. Upon presentation of the search array, the fixation cross disappeared. The search array remained on the screen until a response was registered. The response triggered the next trial immediately.
Each additional-singleton block had 64 trials, in line with our previous work (Wentura et al., 2014). However, the design was based on 128 trials that were randomly divided into two blocks of 64. Thus, two consecutive blocks comprised a total of 128 trials, which were eight repetitions of each combination of set size (5 vs. 7), target line orientation (horizontal vs. vertical), singleton line orientation (horizontal vs. vertical), and target shape (circle vs. diamond). Four of these repetitions featured no colour singleton, the other four contained a colour singleton (one of each colour). In total, 384 trials of the additional singleton task were presented.
Evaluative-priming task
Participants were instructed to categorise target words into the valence categories “positive” (with key “m”) and “negative” (key “c”). Before each target word, a prime was briefly presented, which had to be ignored. The prime could either be a coloured shape filled with a horizontal or vertical line, or a smiley or frowny face. The exact sequence of a trial was as follows. After a 500 ms fixation cross, the prime was shown for 100 ms, followed by a 100 ms blank screen, resulting in a stimulus onset asynchrony of 200 ms. Finally, the target word appeared and stayed on the screen until a response was registered, followed by an intertrial interval of 800 ms.
Two blocks of this task were administered, containing 72 trials each—featuring all combinations of 12 target words and six primes (i.e., smiley/frowny face, and the four colours)—plus two filler trials at the start. The orientation of the line inside the prime shape was chosen randomly. There was a practice block of 12 trials. Between the blocks, participants were given the opportunity to rest.
Results
Unless otherwise noted, all effects referred to as statistically significant throughout the text are associated with p values less than .05, two-tailed.
Additional-singleton task
Mean response times (RTs) were computed using correct responses only. Across all participants, the average error rate was 7.1% (SD = 6.4%). RTs that were 1.5 interquartile ranges above the third quartile of the individual response time distribution (see Tukey, 1977) or were below 200 ms were discarded (on average 5.5% of trials).
Table 1 shows the means and error rates for the conditions of interest (see Figure 3 for the singleton effects, that is, the differences between the respective singleton condition and the no-singleton condition). For the main analysis, we collapsed across both congruence conditions (see below) to include the no-singleton distractor condition in our analysis. 7 A 2 (group: high-control vs. no-control) × 5 (singleton type: no singleton vs. irrelevant vs. neutral vs. positive vs. negative) MANOVA for repeated measures (following the approach suggested by O’Brien & Kaiser, 1985) yielded a significant main effect of singleton type, F(4,83) = 55.17, p < .001, η p ² = .727, and a significant interaction between singleton type and group, F(4,83) = 2.71, p = .035, η p ² = .116 (F < 1 for the main effect of group). 8 Therefore, we report separate analyses for the two samples.
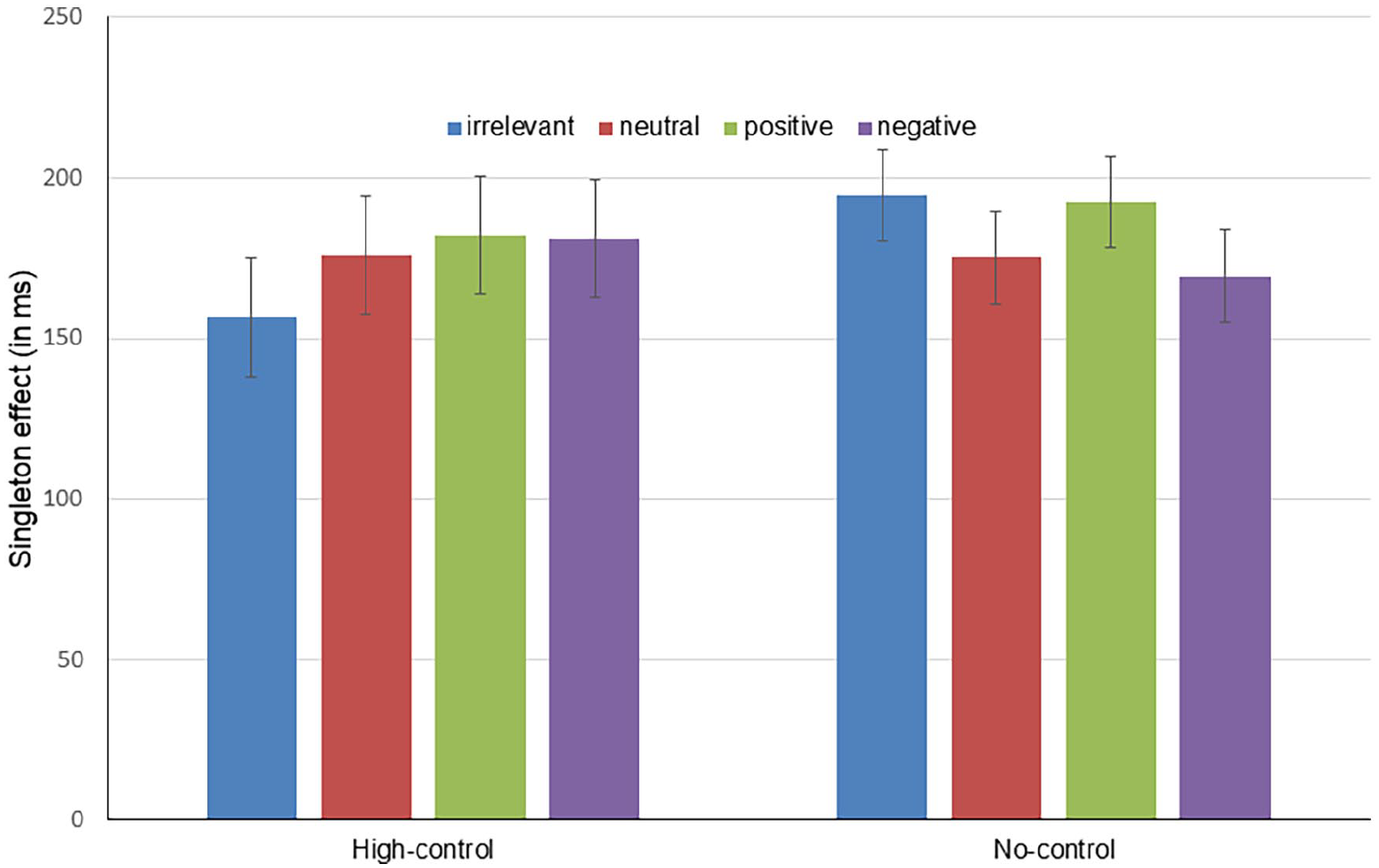
Singleton effects (i.e., RT for specified condition minus RT for the no-singleton condition) as a function of singleton type and group (high-control vs. no-control). Error bars are 95% within-subject confidence intervals (Loftus & Masson, 1994) for the main effect of singleton type in the respective group.
Mean response times (RTs, in ms) and accuracy (in %) as a function of control condition, congruence, and singleton type.
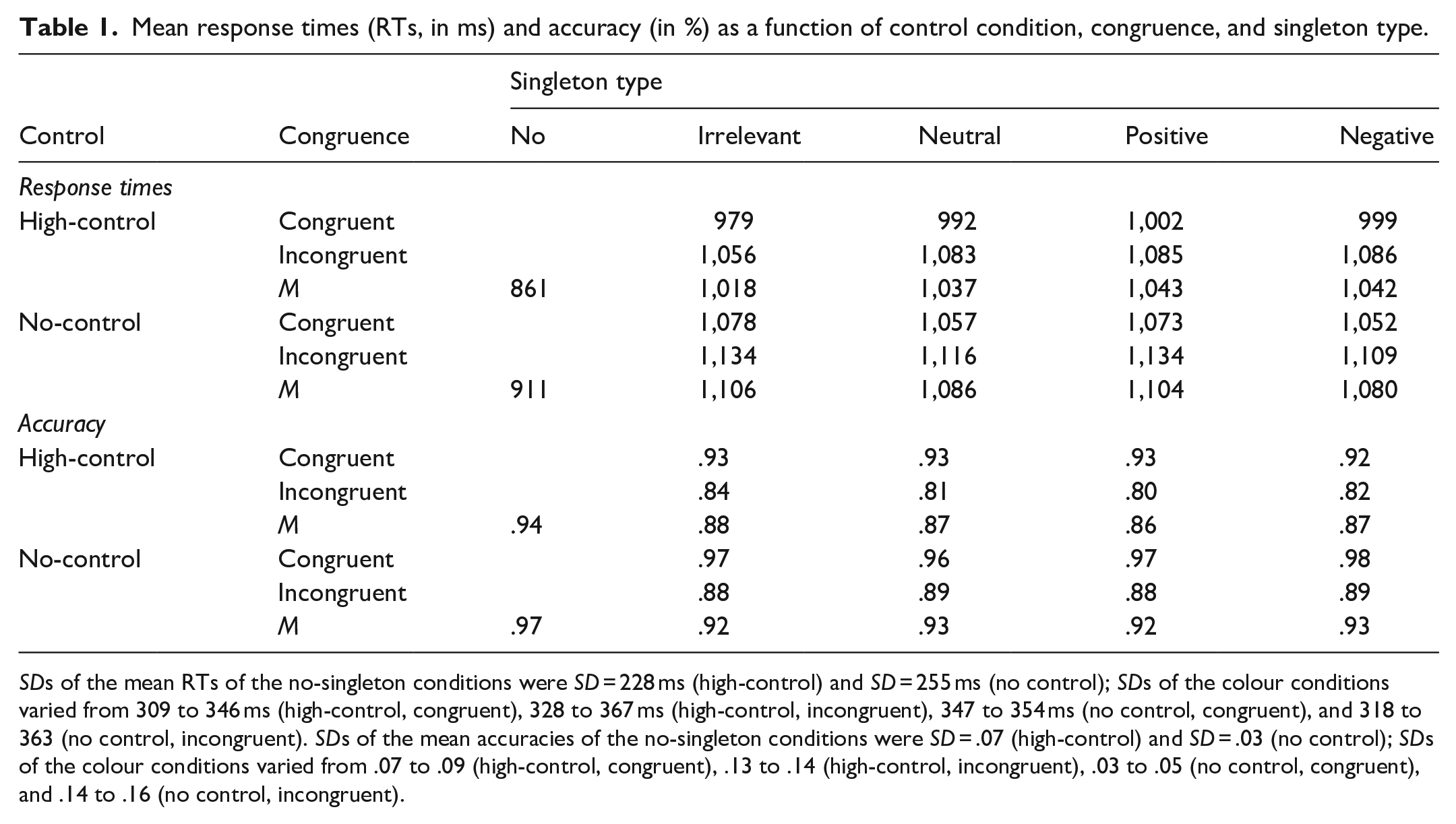
SDs of the mean RTs of the no-singleton conditions were SD = 228 ms (high-control) and SD = 255 ms (no control); SDs of the colour conditions varied from 309 to 346 ms (high-control, congruent), 328 to 367 ms (high-control, incongruent), 347 to 354 ms (no control, congruent), and 318 to 363 (no control, incongruent). SDs of the mean accuracies of the no-singleton conditions were SD = .07 (high-control) and SD = .03 (no control); SDs of the colour conditions varied from .07 to .09 (high-control, congruent), .13 to .14 (high-control, incongruent), .03 to .05 (no control, congruent), and .14 to .16 (no control, incongruent).
High-control sample
For the high-control sample, we found a significant main effect of singleton type, F(4,37) = 31.74, p < .001, η p ² = .774. The first Helmert contrast (no singleton vs. singleton conditions, i.e., the contrast testing for the basic additional-singleton effect) was significant, F(1,40) = 89.97, p < .001, η p ² = .692. The second Helmert contrast, comparing irrelevant to the remaining singleton conditions yielded a significant effect as well, F(1,40) = 4.81, p = .034, η p ² = .107, indicating that subjects reacted faster when an irrelevant (i.e., no-go) additional singleton was present compared with the relevant ones (i.e., the go singletons, incl. the valent ones; see also below). The remaining contrasts (neutral vs. negative/positive and negative vs. positive) were nonsignificant, all Fs < 1. Whereas the latter result (concerning the positive/negative contrast) is in line with Wentura et al. (2014), the former (concerning the contrast neutral vs. negative/positive) seems to be in contrast with the central result of the earlier study. Note, however, that Wentura et al. collapsed the irrelevant and the neutral condition (for reasons of conciseness and because means were almost identical). To clarify the evidence, in the online Supplementary Material of the present article we present cross-experiment analyses. To anticipate, there is substantial evidence for the positive/negative colours causing slower responses than the neutral colour.
No-control sample
In the no-control sample, a MANOVA for repeated measures yielded a significant main effect of singleton type, F(4,43) = 35.43, p < .001, η p ² = .767. Again, the first Helmert contrast (testing for the basic additional-singleton effect) was significant, F(1,46) = 129.07, p < .001, η p ² = .737. In addition, the second Helmert contrast indicated that participants had the tendency to respond slower in the presence of irrelevant colour singletons compared with neutral, negative, and positive colour singletons (combined), F(1,46) = 4.28, p = .044, η p ² = .085. The third Helmert contrast (neutral vs. negative/positive) was nonsignificant, F < 1. Finally, we found the positive colour singleton to be significantly more interfering with the search task than the negative colour singleton, F(1,46) = 8.63, p = .005, η p ² = .158.
Error rates
For error rates, a 2 (group) × 5 (singleton type) MANOVA for repeated measures yielded significant main effects of group (with more errors in the high-control sample), F(1,86) = 11.17, p = .001, η p ² = .115, and singleton type, F(4,83) = 21.94, p < .001, ηp² = .514, but no significant interaction, F(4,83) = 1.35, p = .258, η p ² = .061. The main effect of singleton type was exclusively due to the first Helmert contrast (reflecting the basic additional-singleton effect), F(1,86) = 86.02, p = .001, η p ² = .500 (Fs < 2.02, ps > .159 for the remaining contrasts.
Congruence effects
To assess the impact of the congruence between line orientations of target and singleton distractors, we conducted a 2 (group: high-control vs. no-control) × 4 (singleton type: irrelevant vs. neutral vs. negative vs. positive) × 2 (orientation congruence: congruent vs. incongruent) MANOVA for repeated measures with RTs as the dependent variable. We report only effects regarding orientation congruence to avoid redundancies. There was a significant main effect of orientation congruence, F(1,86) = 48.45, p < .001, η p ² = .360, but no significant evidence for a moderation by group, F(1,86) = 1.67, p = .199, η p ² = .019. The interaction of orientation congruence with singleton type as well as the three-way interaction was nonsignificant, for both Fs < 1 (all Fs < 1 for the corresponding Helmert contrasts). For error rates, the corresponding analysis yielded a significant orientation congruence effect, F(1,86) = 58.18, p < .001, η p ² = .404. Again, there were no significant interactions involving this factor, Fs < 1.83, ps < .148.
Action relevance effect
As the pattern of means of the irrelevant and the neutral colour (see Figure 3) suggest an effect of action relevance (i.e., whether a colour calls for a response) beyond valence-based effects, we added a post hoc explanatory analysis. We created a go-singleton variable, which was the neutral condition for the high-control sample and the irrelevant condition for the no control sample, and a no-go-singleton variable, which was based on the reversed assignment. A 2 (group: high-control vs. no-control) × 2 (singleton type: go vs. no-go) ANOVA for repeated measures with RTs as the dependent variable yielded a main effect of singleton type, F(1,86) = 4.62, p = .034, η p ² = .051, which was not moderated by group, F < 1. A corresponding analysis for error rates yielded no main effect of singleton condition, F(1,86) = 1.46, p = .230, η p ² = .017, and no interaction, F < 1.
Evaluative-priming task
Only RTs from correct responses were used in the analysis of the evaluative-priming task. The average error rate across participants was 4.0% (SD = 3.75). RTs that were 1.5 interquartile ranges above the third quartile of the individual response time distribution (see Tukey, 1977) or were below 200 ms were discarded (on average 5.2% of trials).
For each prime type, we computed the difference between positive and negative target responses, that is, a relative positivity index (see Table 2). Subsequently, we conducted separate ANOVAs for colour primes (positive vs. negative) and control primes (smiley vs. frowny face), using these difference scores as the dependent variable. For the control primes, a 2 (group) × 2 (prime valence) ANOVA yielded a main effect of prime valence, F(1,86) = 31.65, p < .001, η p ² = .269, that did not interact with group, F < 1. This reflects a typical evaluative-priming effect (i.e., the positivity index is significantly larger for smiley compared with frowny faces). The same holds for the valent colour primes, as the corresponding ANOVA yielded a significant main effect of prime valence, F(1,86) = 5.63, p = .020, η p ² = .061, that did not interact with group either, F < 1.
Mean response times (RTs, in ms) in the evaluative-priming task as a function of target and prime valence (errors in % in parentheses), and priming effects (in ms; standard errors in brackets).
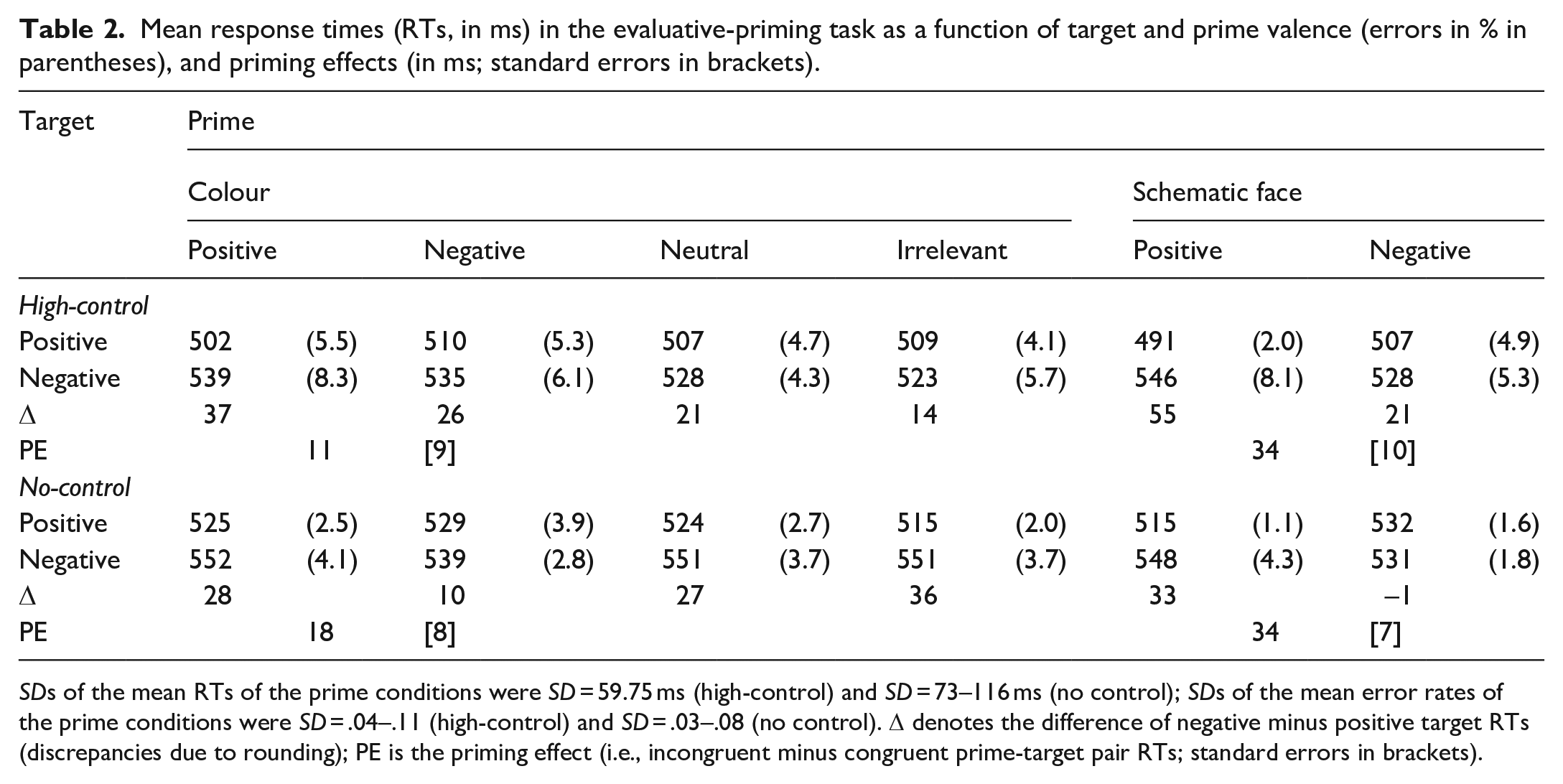
SDs of the mean RTs of the prime conditions were SD = 59.75 ms (high-control) and SD = 73–116 ms (no control); SDs of the mean error rates of the prime conditions were SD = .04–.11 (high-control) and SD = .03–.08 (no control). Δ denotes the difference of negative minus positive target RTs (discrepancies due to rounding); PE is the priming effect (i.e., incongruent minus congruent prime-target pair RTs; standard errors in brackets).
We had no a-priori hypotheses about the neutral and irrelevant colour primes. The values in Table 2 might evoke the post hoc hypothesis that the respective go category (i.e., neutral for the high-control sample; irrelevant for the no-control sample) is associated with a larger positivity index than the respective no-go category (i.e., irrelevant for the high-control sample; neutral for the no-control sample). A 2 (group: high-control vs. no-control) × 2 (prime type: go vs. no-go) ANOVA, however, yielded no significant effect of prime type, F(1,86) = 1.62, p = .206, η p ² = .019, and no interaction with group, F < 1.
Induction task
Finally, to complete the picture, we analysed the performance of high-control participants in the induction phase. We had no specific hypotheses with regard to this. Mean RTs were computed using correct responses only. Across all participants, the average error rate was 16.2% (SD = 8.1%). RTs that were 1.5 interquartile ranges above the third quartile of the individual response time distribution (see Tukey, 1977) or were below 200 ms were discarded (on average 5.7% of trials).
Table 3 shows parameters of performance in the induction phase of the high-control sample. For response times, a one-factorial analysis (type: neutral vs. positive vs. negative) yielded a significant effect, F(2,39) = 18.30, p < .001, η p ² = .484. Both Helmert contrasts were significant: F(1,40) = 16.01, p < .001, η p ² = .286 for the contrast neutral versus positive/negative collapsed and F(1,40) = 29.18, p < .001, η p ² = .422. The analysis of accuracies shows a corresponding pattern: a significant overall effect, F(2,39) = 13.49, p < .001, η p ² = .409 and significant Helmert contrasts: F(1,40) = 5.17, p = .028, η p ² = .115 for the contrast neutral vs. positive/negative collapsed and F(1,40) = 20.44, p < .001, η p ² = .338. As can be easily seen from Table 3, the positive condition is associated with a boost in performance, in RTs as well as accuracies.
Reaction times, accuracy, and success rate (SD in parentheses) in valence induction trials as a function of stimulus condition in the high-control sample and probabilities of reward and loss.
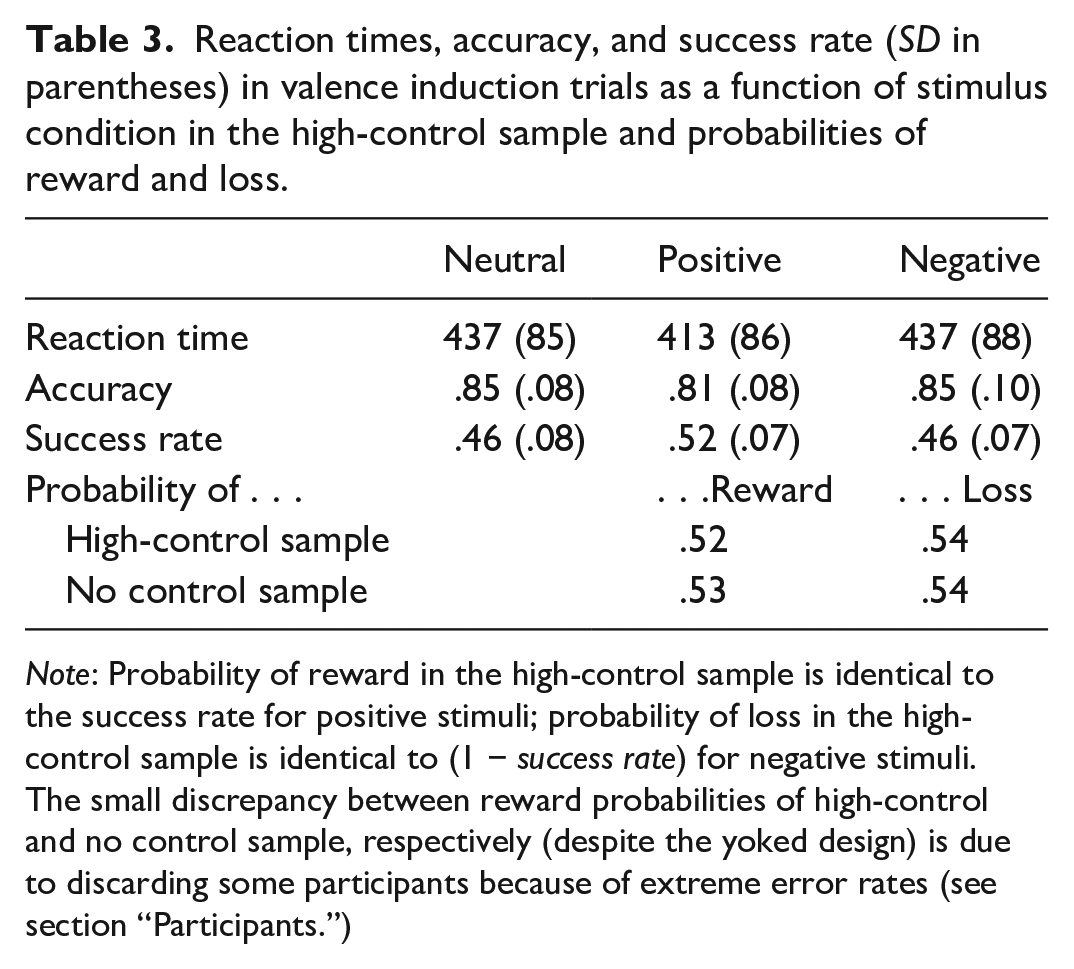
Note: Probability of reward in the high-control sample is identical to the success rate for positive stimuli; probability of loss in the high-control sample is identical to (1 − success rate) for negative stimuli. The small discrepancy between reward probabilities of high-control and no control sample, respectively (despite the yoked design) is due to discarding some participants because of extreme error rates (see section “Participants.”)
This can be additionally seen in the success rates for the different stimulus types (colours) in the valence induction blocks. To recall, “success” was defined as a correct response faster than the actual moving median (see Procedure). As can be easily seen, the moving median procedure worked properly as the average values are roughly at 50% success with rather small standard deviations. For the sake of completeness, we report the inferential statistics, although these are more-or-less redundant with regard to the results for RTs and accuracies: a significant overall effect, F(2,39) = 12.67, p < .001, η p ² = .394 and significant Helmert contrasts: F(1,40) = 9.04, p = .005, η p ² = .184 for the contrast neutral versus positive/negative collapsed and F(1,40) = 13.34, p < .001, η p ² = .250. The difference between success rates for positive and negative trials, however, means that the probabilities of rewards (= success rate for positive stimuli) and losses (= 1 − success rate for negative stimuli; see Table 3) are not different, t(40) = 1.05, p = .30, dZ = 0.16.
Discussion
We assessed the attentional bias for positive and negative connoted stimuli in two different samples. In an induction phase, the high-control sample learned that the positive stimulus signalled the opportunity to win money whereas the negative stimulus signalled the risk to lose money; winning or losing was performance-dependent. Notably, the number of winnings and losings were equal. In a yoked design, the no-control sample received the same winnings and losses, however, seemingly on a random basis, that is, completely independent of their behaviour.
In the high-control sample of the additional-singleton task, we found a pattern that closely resembled our previous findings (Müller et al., 2016; Wentura et al., 2014, 2018): conditions with positively and negatively valenced singleton distractors showed no difference in target RTs, but were associated with a numerically enhanced additional-singleton effect compared with the neutral distractor condition and a significantly enhanced additional-singleton effect compared with the irrelevant-distractor condition. Admittedly, diverging from previous findings (no RT difference between irrelevant and neutral colour conditions; Wentura et al., 2014), the neutral-distractor RTs were in-between the irrelevant- and valent-distractor RTs (and not significantly different from either). However, a cross-experiments analysis with the present data and the one from Wentura et al. (2014, see Supplement), corroborates that the high control version of the paradigm shows substantial evidence for (a) a contrast valent colours versus neutral colour and (b) the absence of a difference between positive and negative colour.
In the no-control sample, we found a different pattern, which was almost a reversal of the high-control pattern: the additional-singleton effect was enhanced for the irrelevant colour, which in this group was the only colour that required an active response during the valence-induction task (we will return to this aspect later). More important, we found a clear asymmetry for the two valent colours: the additional-singleton effect was increased for the positively connoted colour (“reward”) compared with the negatively connoted colour (“punishment”). Thus, a feature that signals a performance-independent reward attracts (or holds) attention, whereas a feature that signals an uncontrollable loss does not.
Our research was inspired by a functional point of view: signals for opportunities and dangers—that is, signals for controllable positive and negative events—should bias attention because it is of utmost importance to focus on adequate actions. In contrast, paying attention to signals of uncontrollable events (be it positive or negative ones) brings no advantage. It seems this prediction holds for signals of uncontrollable loss but not for uncontrollable reward. This asymmetry corroborates earlier research in the attentional capture field (see “Introduction”) that suggested more robust effects for uncontrollable reward versus loss.
The pattern of our results can be integrated into a broader framework of affective processing: Biases might be in service of goal pursuits and motivational regulation (Brandtstädter et al., 2004; Rothermund, 2011). Rothermund (2011) coined the term control-dependency principle: “If an individual has a sense of control over the situation, affective processing is focused on potential problems (‘negativity bias’), whereas if the individual perceives the situation to be uncontrollable, affective processing is oriented toward enhancement (‘positivity bias’). The control-dependence principle helps to prevent premature resignation as well as wasteful perseverance.” This general principle fits the present pattern, except that in the high control situation positivity and negativity biases are balanced.
How do the results fit to earlier findings in the field? First of all, the results roughly match our synopses of reward-only and reward-and-punishment studies (see “Introduction”). Of course, we have to acknowledge that our findings seem to be in contrast to some of the previous studies on aversive conditioning (low instrumental contingency) that did find increase of the additional singleton effects for CS+ over CS− (e.g., Schmidt et al., 2015). Indeed, this result did not match one aspect of our results: the complete absence of a loss-related capture effect in our no-control sample. Thus, one might speculate that an aversive stimulus like an electric shock or a sound burst might lead to a stronger attentional response to the CS+—as it might be more closely linked to a physical threat—than the possibility of a small monetary loss. However, our result has a second component: The bias effect in our no-control sample was significantly enhanced for reward-related compared with loss-related colours (whereas there was no difference in the high-contingency sample). Transferred back to the aversive conditioning experiments: we do not know whether a reward that is associated with physical pleasure and is comparable in intensity to a shock might be associated with an even larger attentional capture effect than a shock-associated stimulus. It remains that a virtue of our design is the comparability of reward and loss in a single experiment.
A further point that has to be discussed is the operationalisation of contingency. The high-control group learns the colour-reward and colour-loss associations through discriminative conditioning (stimulus–response–outcome), whereas the low-control group learns these associations via Pavlovian conditioning (stimulus–outcome). Our operationalisation of low instrumental contingency is in line with previous studies using Pavlovian conditioning (e.g., Bucker & Theeuwes, 2016; Koster et al., 2004; Notebaert, Crombez, Van Damme, et al., 2011) and allowed us to investigate to what extent a stimulus attracts attention when it has no action relevance. Another operationalisation of low instrumental contingency commonly found in the literature is to realise low instrumental contingency inside the discriminative conditioning approach by removing the association between response and outcome (Anderson et al., 2011a; Failing & Theeuwes, 2014), that is, participants are required to give a response although they know that it has no effect on the outcome. These experiments studied, however, only the effects of reward. As they found a bias effect (as we did in both reward conditions of our experiment), there is no conflict to our results. It might be a question for further research to explore whether the two forms of operationalising no-contingent losses (i.e., within the discriminative conditioning approach versus the Pavlovian way) might lead to different effects on attention.
Attentional capture or attentional dwelling?
Besides the introduction of the no-control condition, we changed one additional detail from Wentura et al.’s (2014) procedure. In the earlier experiment, the distractor stimuli in the additional-singleton task featured a non-categorisable line (i.e., the line was tilted and thus neither vertical nor horizontal). In the present experiment, the distractor stimuli had lines that either matched or mismatched the orientation of the target line, allowing for the calculation of a congruence effect. There was indeed a large congruence effect (i.e., if the line orientation in the colour singleton matches the one in the target, responses were faster than in case of a mismatch), which, however, was not moderated by the other factors (i.e., it was not different for the different colours). With some caution, this result provides support for one side of the debate regarding attentional capture versus maintenance—that is, whether the “add-on” effect of evaluative distractor connotations compared with neutral singletons is due to enhanced attentional capture or impeded attentional disengagement (Fox et al., 2002), or a mixture of both (see Wentura et al., 2014, for a discussion). The attentional-capture hypothesis proposes that the “add-on” effect is due to an increased probability of attentional capture by the specific (i.e., valent or positive) colour singletons (compared with neutral ones). The line in an attended distractor, however, will produce a larger congruence effect than the line in an unattended one. Thus, if the “add-on” is indeed caused by a larger propensity of the additional singleton to capture attention, the size of the congruence effect should covary with the size of the corresponding “add-on” part of the additional-singleton effect. This was not observed. Thus, our result is more in line with the attentional maintenance hypothesis. This assumes that the propensity of additional singletons to capture attention is constant across colours; if, however, attention is captured by a colour singleton, specific features—most important: its valence—cause additional attentional dwelling. This conclusion meshes well with our analysis in a recent article (Wentura et al., 2018) as well as the results of a further experiment, where we used the same valence-induction procedure as in this study, but employed a modified dot-probe task that allowed for an experimental separation of capture and disengagement processes (Müller et al., 2016).
Is there an effect of action relevance?
As an aside, besides the evidence for valence-based effects, we found (exploratory) evidence for an effect of action relevance in both samples: in both groups, one of the affectively neutral colours called for a response (go singleton; i.e., the neutral one in the high-control sample and the “irrelevant” one in the no-control sample) whereas the other one called for withholding a response (no-go singleton; i.e., the irrelevant one in the high-control sample and the neutral one in the no-control sample). The go singletons were associated with longer RTs (i.e., more attentional bias) than the no-go singletons. This was not the case in Wentura et al.’s (2014) experiment, which was comparable to the present high-control condition in all aspects but one. A possible reason for this might be that the distractors in the present additional-singleton task contained a categorisable line (i.e., the lines were horizontal or vertical as were the target lines). This was not the case in Wentura et al.’s (2014) experiment, where distractor lines were tilted. Thus, if attention switches to the colour singleton, the combination of processing both a response-related feature (i.e., line orientation) and a “go” signal might induce greater costs associated with suppressing a response to the wrong stimulus than either feature alone (i.e., the line in the “no-go” neutral-colour distractor in the present experiment or the “go” colour in Wentura et al., 2014). Future research should scrutinise this explanation further.
With some caution, one might discuss the study by Becker et al. (2020) in the context of this action relevance effect. To remind, the authors realised a high instrumental contingency schedule for reward and punishment, but found an asymmetric pattern of capture effects: the reward colour but not the punishment colour increased latencies in the test phase (i.e., caused attentional capture). However, during the training phase, participants learned that fast and correct responses were required to receive a reward (in reward-colour trials), while slow, withheld, or incorrect responses prevented punishment (in punishment-colour trials). That is, one can see in it a toned-down version of “go” versus “no-go.”
In conclusion, we found attentional biases for positive and negative connoted stimuli to be dependent on the control that participants had on gaining winnings or losses. Thus, signals of opportunities and dangers both cause an attentional bias, whereas an asymmetry was found if rewards and punishment were out of control of participants. In this case only rewards caused an attentional bias.
Supplemental Material
sj-docx-1-qjp-10.1177_17470218221099125 – Supplemental material for Undeserved reward but not inevitable loss biases attention: Personal control moderates evaluative attentional biases in the additional-singleton paradigm
Supplemental material, sj-docx-1-qjp-10.1177_17470218221099125 for Undeserved reward but not inevitable loss biases attention: Personal control moderates evaluative attentional biases in the additional-singleton paradigm by Philipp Müller and Dirk Wentura in Quarterly Journal of Experimental Psychology
Footnotes
Acknowledgements
The authors thank Ullrich Ecker for his helpful comments on an earlier version of this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Data accessibility statement
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.