
Abstract
Memory is a dynamic reconstruction process, emphasizing the importance of assessing not only that an event occurred but also its details (i.e., memory accuracy). Recent studies have suggested that reconstructing accurate memories requires discriminating overlapping memory traces, a process known as pattern separation (PS). PS is typically studied through tasks involving the discrimination of visual lures, highlighting the contribution of visual processes to PS. Two main types of visual processing should be considered (high spatial frequencies [HSF]; and low spatial frequencies [LSF]), and it remains unclear whether these processes contribute equally to the discrimination of visual lures. This preregistered study examined the impact of spatial frequencies on mnemonic discrimination using a mnemonic similarity task. We hypothesized that HSF, which process the fine analysis of information, would be more involved than LSF, which process the coarse analysis of information, in discriminating targets from similar lures. Forty young participants had to recognize visual stimuli under three spatial frequency conditions: LSF images, HSF images, and a no-filter (NF) condition. The results showed that the lure discrimination index was lower for the LSF images than for the NF condition. LSF images also resulted in fewer false recognitions of lures and more target misses compared to HSF images and the NF condition. These results suggest that HSF support mnemonic discrimination more than LSF but also contribute more broadly to the overall richness of memory traces. This study contributes to theoretical models of memory by highlighting the importance of rich visual information for accurate memory.
Keywords
Introduction
Confusing two similar memories is very common, such as wondering whether we told a story to one friend or another, or believing that we had seen a piece of information in a specific article when it came from a different source. These confusions illustrate the difficulty of distinguishing closely related memories, a capacity crucial for accurate memory (Stark et al., 2019; Surprenant & Neath, 2009; Versace et al., 2014). Although this mnemonic discrimination ability is well known for visual information (Mille et al., 2023a, 2023b), we have a clear lack of knowledge of the specific contribution of different components of visual information, such as fine details and the overall shape of objects, for example the small features of a cup versus its general silhouette, as assessed through spatial frequencies (e.g., Kauffmann et al., 2015; Ramanoël et al., 2015). The aim of the present study was to address this gap by investigating how different spatial frequency components contribute to mnemonic discrimination.
Memory has long been assessed in terms of the quantity of information retrieved, mainly through the number of items correctly recalled or recognized (see Koriat et al., 2000). However, this conception has a major limitation, as it tends to rely on a dichotomy between successful and failed retrieval, without sufficiently considering that retrieval itself can involve errors (e.g., Surprenant & Neath, 2009; Versace et al., 2009, 2014). These errors, such as memory distortions (e.g., false recognition of lures; Stark et al., 2019) or false memories (e.g., recall of something that never happened; Loftus & Palmer, 1974, 1996; Schacter, 2001), demonstrate that memory does not simply reproduce information faithfully. Instead, memory is a dynamic process that reconstructs memories in a flexible way (see Mille et al., 2023a). The dynamic nature of memory gives it a highly adaptive nature (Schacter, 2012) by supporting, for instance, creative behavior (Addis et al., 2016) or the ability to imagine the future (Schacter et al., 2017). This view of memory thus focuses more on the specificity of the memory, that is, its accuracy (Koriat et al., 2000; Mille et al., 2023a; Purkart et al., 2022).
Numerous studies have shown that reconstruction of accurate memories is facilitated when the memory trace is distinct from other content in the system (e.g., Hunt, 2013; Hunt & Worthen, 2006; Mille et al., 2023a). Conversely, other studies have found that the accuracy of memory reconstruction is reduced when memory traces overlap (Mille et al., 2023a, 2023b; Roediger & McDermott, 1995; Roediger et al., 2001). In this regard, many researchers from different fields have proposed modeling memory reconstruction based on the system’s ability to discriminate between memory traces, for example, in neuroscience (e.g., pattern separation models; Stark et al., 2019, for a review), in computational modeling (e.g., the scale-independent memory, perception, and learning model, Brown et al., 2007; Surprenant & Neath, 2009), and in cognitive psychology (e.g., the activation-integration model, Versace et al., 2009, 2014). The general idea underlying these proposals is that the accuracy of memory reconstruction decreases when memory traces strongly overlap, because such overlap creates interference and makes it more difficult to selectively activate the trace or traces corresponding to the target event (Mille et al., 2023b; Versace et al., 2009, 2014). Memory accuracy is thought to rely on an activity of discriminating between memory traces, as it depends on the extent to which the present situation matches specific information while excluding other information in memory (Nairne, 2006; Surprenant & Neath, 2009; Versace et al., 2009, 2014).
The ability to process the interference caused by overlapping traces has been widely described in the field based on the general principle of information coding, which is based on hippocampal functional theory (e.g., Marr, 1971; Rolls, 2013). According to this theoretical framework, reconstruction of accurate memories depends on two mechanisms: pattern separation (PS) and pattern completion (PC). PS is defined as the ability to reduce interference from similar percepts by processing non-overlapping representations, while PC allows for the reconstruction of a complete memory from an incomplete input signal by completing (activating) the missing components. At a behavioral level, although the effect of interfering items on memory precision can be assessed through multiple paradigms, it is most commonly evaluated using recognition tasks. In this context, a PS deficit can be demonstrated through a task called the Mnemonic Similarity Task (MST, Stark et al., 2019). This task generally consists of a recognition task (usually of images) in which three types of stimuli are presented during the recognition phase: targets, distractors, and lures. In the modified recognition phase, for each stimulus, the participant is asked to determine whether the presented item is “old,” “new,” or “similar.” The results showed, on the one hand, that as the similarity between lures and targets increased, false recognitions increased, and the Lure Discrimination Index (LDI), based on the probability of “similar” responses to lures minus the probability of “similar” responses to foils, decreased (Leal & Yassa, 2018; Stark et al., 2019). On the other hand, the results showed that a lesion to hippocampal structures involved in PS increased false recognitions of the lures and decreased the LDI score (Stark et al., 2019).
It is important to note that numerous studies have suggested that PS is not limited to the hippocampus (Cowell et al., 2019; Ekstrom & Yonelinas, 2020; Kent et al., 2016). For example, according to the hierarchical representational approach (Kent et al., 2016; Murray & Bussey, 1999; Saksida & Bussey, 2010), increasingly complex non-overlapping representations (PS) of visual stimuli would be processed along the ventral pathway. Basic sensory interference between individual stimulus characteristics (e.g., lines and colors) can be resolved in the sensory cortex (Kent et al., 2016). When more perceptually complex features are introduced (e.g., objects), the interference is resolved at a higher processing level (e.g., the perirhinal cortex; Barense et al., 2005, 2010, 2011, 2012). Finally, combinatorial codes (e.g., conjunctive representations of object scenes in a spatial context) are resolved in the hippocampus (for a review, Martin & Barense, 2023). Importantly, according to the hierarchical representational approach, conjunctions would not only be important for mnemonic discrimination but also for tasks requiring the discrimination of similar visual signals (see Kent et al., 2016). This suggests that mnemonic discrimination involves hierarchical processing of visual representations that partly rely on the ability to discriminate low-level characteristics of visual signals such as contrast of luminance, textures, colors, spatial frequency, dominant orientations, etc. (Bartnik & Groen, 2023).
Among these features of visual information, there is considerable evidence suggesting the importance of the spatial frequency content of visual stimuli for recognition (Kauffmann et al., 2014; Ramanoël et al., 2015; Torralba & Oliva, 2003, see also Gingras et al., 2026 for research investigating potential cultural effects on these processes). According to influential models of visual perception (Bar, 2003; Bullier, 2001; Hegdé, 2008; Kauffmann et al., 2014; Schyns & Oliva, 1994), visual analysis is initiated by the parallel extraction of various attributes across different spatial frequencies. Low spatial frequencies (LSF) allow for rapid and coarse analysis of information across the occipital cortex and high-level areas in the dorsal cortical stream, while high spatial frequencies (HSF) allow for a delayed and detailed analysis of information in the ventral cortical stream (Kauffmann et al., 2015, but see also Farhang et al., 2025, for more nuanced findings). Interestingly, it is possible to manipulate the spatial frequency content of visual stimuli by applying filters that selectively transmit either low- or high-frequency information. For example, a low-pass filter will produce a blurry image but will allow the overall shape of a tennis ball to be perceived, whereas a high-pass filter will highlight fine details, such as the surface texture and seams, at the expense of the overall shape perception (e.g., Kauffmann et al., 2015; Ramanoël et al., 2015). In the context of object perception, recent studies have highlighted the importance of spatial frequency contents during recognition processes (Ashtiani et al., 2017; Avberšek et al., 2021; Boucart et al., 2025; Roux-Sibilon et al., 2019; Wamain et al., 2023). For example, Wamain et al. (2023), using a categorization task with filtered images of living and non-living objects, reported that behavioral performance decreased for LSF pictures of furniture, consistent with a greater reliance on HSF information for these items.
This differentiated processing between LSF and HSF may also play a crucial role in memory discrimination, particularly when it involves distinguishing a target memory from a visually similar lure. More precisely, in the case of perceptual lures, which would be highly similar to the targets, finer processing would be required to detect the subtle differences that allow for their rejection. Indeed, when two stimuli share the same global configuration, only the analysis of precise details, such as slight variations in contour, texture, or element spatialization, would reliably distinguish them. Thus, coarse visual information conveyed by LSF will be sufficient to discriminate between dissimilar visual stimuli, whereas the finer processing underpinned by HSF will be necessary to separate ambiguous stimuli. If this is true, it could be expected that the LDI would rely more on HSF than on LSF. Surprisingly, although this critical step of visual discrimination appears to be particularly relevant for our understanding of memory recognition, no study has investigated this issue. More precisely, despite the fact that many studies have focused on tasks involving the rejection of perceptual lures (see Stark et al., 2019), to our knowledge, no study has sought to determine whether this rejection (and by extension, memory discrimination) could be modulated by the spatial frequency content of the stimuli.
The Present Study
The aim of this study was to investigate which spatial frequencies are the most involved in visual memory discrimination. Since the processing of visual details is mainly underpinned by HSF and memory discrimination in the MST relies on the ability to reconstruct fine visual details in memory, we hypothesize that memory discrimination should be more strongly underpinned by HSF compared to LSF. More specifically, we hypothesized that in an MST, LSF images should increase false recognitions of visual lures and decrease the LDI compared to HSF images and a no-filter (NF) condition. All hypotheses and analyses were preregistered with OSF (see https://osf.io/snm93).
Experiment
Method
Participants
We conducted a power analysis based on the data of Ramanoël et al. (2015), a study that specifically aimed to investigate the effect of spatial frequency filters on an indoor/outdoor scene categorization task. The power calculation was carried out using simulations via the SuperPower package (Caldwell et al., 2019) using R version 4.0.4 (R Core Team, 2018). Analysis indicated that 30 participants were required to achieve 95% chance to detect a significant effect for both NF/HSF and LSF/HSF contrast conditions (for more details on the parameters used, see the R script available at the following OSF link: https://osf.io/6n2ha). Although this power analysis has been based on a different dependent variable (i.e., indoor/outdoor image categorization) than the one we used, the number of participants was expected to be sufficient to detect an effect in the MST. Indeed, in this latter task, the perceptual demand was higher than in the categorization task used in Ramanoël et al. (2015). Consequently, the effect of the various filters was expected to be at least equivalent or even more pronounced.
In order to anticipate possible exclusions, data collection was stopped at 40 participants. Therefore, 40 native French speakers (Mage = 19.38 years, SDage = 1.58, rangeage = 18–25, MEducation = 12.6 years, SDEducation = 1.28, rangeEducation = 12–17, 31 female, 39 right-handed) were recruited from the general population. All the participants had normal or corrected-to-normal vision and hearing, and no history of neurological or psychiatric disorders. Two participants were excluded from the analyses: one due to a data recording error, and the other due to a reported misunderstanding of the instructions during the debriefing session. Thus, the analyses were conducted on a sample of 38 participants (Mage = 19.24 years, SDage = 1.50, rangeage = 18–25, MEducation = 12.63 years, SDEducation = 1.31, rangeEducation = 12–17, 31 female, 37 right-handed). This study was carried out in accordance with the recommendations of French Law (Loi Jardé no. 2012-300), with written informed consent being obtained from all the subjects in accordance with the Declaration of Helsinki. Ethics approval was issued for the current study by the local ethics committee of Tours University (CER-TP 2023-12-02).
Materials
Visual Stimuli
One hundred ninety-two object images were selected from the database provided by the Stark team (Stark et al., 2019; available at the following address: https://github.com/celstark/MST) to serve as the targets (96 images) and lures (96 images) for the MST. The items were selected based on the following criteria. Written text and faces were excluded from the selection due to the difficulty in achieving reliable recognition when spatial frequencies vary. Similarly, living objects were avoided. Living objects are recognized to exhibit stronger and more frequent intercorrelations between sensorial features than inanimate objects (e.g., Ventura et al., 2005) so that spatial frequencies filtering may make very difficult to distinguish one item from another. Despite this rule, some items may not have fully met this criterion and were therefore used for the practice phase. Special attention was also given to the size of the objects: those occupying a large portion of the screen were preferred to avoid small objects that may not be optimally perceived. Additionally, the items had to represent different types of semantic information (e.g., two different types of wallets to prevent them from being perceived as lures). Finally, to ensure adequate visual discriminability, each item pair (target-lure) was pretested 1 to verify that the distinction between the target and its corresponding lure remained robust across filtering conditions (LSF or HSF images). In addition, 48 supplementary object images were selected to serve as foils. Among these, 11 were taken from the same database, and 37 were taken from the Poporo image set (Kovalenko et al., 2012). Finally, 12 additional images 2 were selected for the practice session: 6 to be memorized (3 serving as targets and 3 as lures) from the Stark team’s database. The remaining three foils for the practice session were selected from the Poporo image set. In total, 252 images were selected.
Each image was first converted to grayscale (256-level grayscales) and resized to 400 × 400 pixels subtending 7.15° × 7.15° at a viewing distance of 57 cm. Then, three types of spatial filters were applied to each image to create the three PASS FILTER TYPE: NF, that is, a control condition in which spatial frequency was not manipulated, HSF, that is, images filtered with a high-pass filter, and LSF, that is, images filtered with a low-pass filter (see Figure 1, e.g., images for each type of filter). These filters were applied using MATLAB (MathWorks, Inc., Sherborn, MA, USA) based on the procedure used in Ramanoël et al. (2015). For LSF images, spatial frequencies above 1 cycle per degree (cpd) of visual angle were removed, while for HSF images, spatial frequencies below 6 cpd were removed. These values were selected from several previous studies assessing the respective role of LSF and HSF during visual perception (Faurite et al., 2025; Kauffmann et al., 2015; Ramanoël et al., 2015). Then, a luminance contrast equalization was applied to obtain a mean luminance value of 0.5 (i.e., mean luminance of 128 on a gray-level scale), setting all images to a value of 0.5, so that they would share the same average contrast amplitude in overall luminance. This was done to ensure that the effects of filtering were not simply due to differences in contrast between images. Finally, root mean square (RMS) contrast normalization was applied, with a value set to 0.3 (i.e., 78 on a gray-level scale) corresponding to the average values of RMS contrast of all images, in order to control for pixel-level variations in contrast across the different images. The stimuli are available at the following link: https://osf.io/6n2ha. As a result, a total of 756 images were generated: 720 for the main task (576 for targets and lures, and 144 for foils), and 36 for the practice session.

Example images for each pass filter type: NF condition (left), i.e., a control condition in which spatial frequency was not manipulated; LSF (center), that is, images filtered with a low-pass filter; and HSF (right), that is, images filtered with a high-pass filter.
Mnemonic Similarity Task
An MST in its explicit memory version was used (see Stark et al., 2019 for a discussion on the differences between the explicit and implicit memory versions). This choice was made to reduce the risk of floor effects on performance, as the use of filters was expected to increase task difficulty.
Procedure
Participants were seated in a dark, windowless room. The experiment was conducted using OpenSesame version 4.0.13 Melodramatic Milgram (Mathôt et al., 2012), with a screen resolution of 1,920 × 1,080 pixels. PyGame was used as the engine to develop, configure, and run the experiment. They were then informed that the experiment would begin with a practice session, followed by a main task, for a total duration of approximately 20 min.
The MST (see Figure 2 for an illustration of the procedure of the MST task) began with a practice session divided into two phases: a study phase and a test phase. Participants were informed that they would see a series of images presented one after another and were instructed to memorize as many details as possible. They were also told that, in a second phase (i.e., the test phase), they would again see images displayed sequentially and would have to indicate, for each image, whether it was exactly identical to one presented during the study phase (the “TARGET” condition, e.g., an apple and the same apple), similar but not identical (the “LURE” condition, e.g., an apple and another picture of a different apple), or entirely new (the “FOIL” condition, e.g., an apple and a printer). When a LURE was presented during the test phase, its corresponding TARGET was not shown. During the study phase of the practice session, participants were instructed to memorize six images (two per PASS FILTER TYPE: NF, HSF, and LSF), presented with a gray background. Each trial began with a fixation cross displayed for 1,000 ms, followed by an image shown for 2,000 ms, and then a 500 ms inter-trial interval (ITI). After this phase, the instructions for the test phase were repeated, and the response keys were given using the index, middle, and ring fingers of the dominant hand (“u” for “TARGET,” “i” for “LURE,” and “o” for “FOIL”). Participants were instructed to respond as quickly as possible when they believed they had the correct answer. They were asked to use their dominant hand and to keep their hand positioned on the response keys throughout the task. During the test phase, participants completed nine trials (three per PASS FILTER TYPE), with one trial for each TRIAL TYPE: “TARGET,” “LURE,” and “FOIL.” The lure was always presented with the same filter as the corresponding image from the study phase. Each trial began with a fixation cross, followed by a blank screen for 500 ms. Then the image was presented and remained on the screen until the participant responded or for a maximum of 5,000 ms. After each response, feedback indicating whether the answer was correct or incorrect was displayed for 400 ms, followed by a 500 ms ITI.
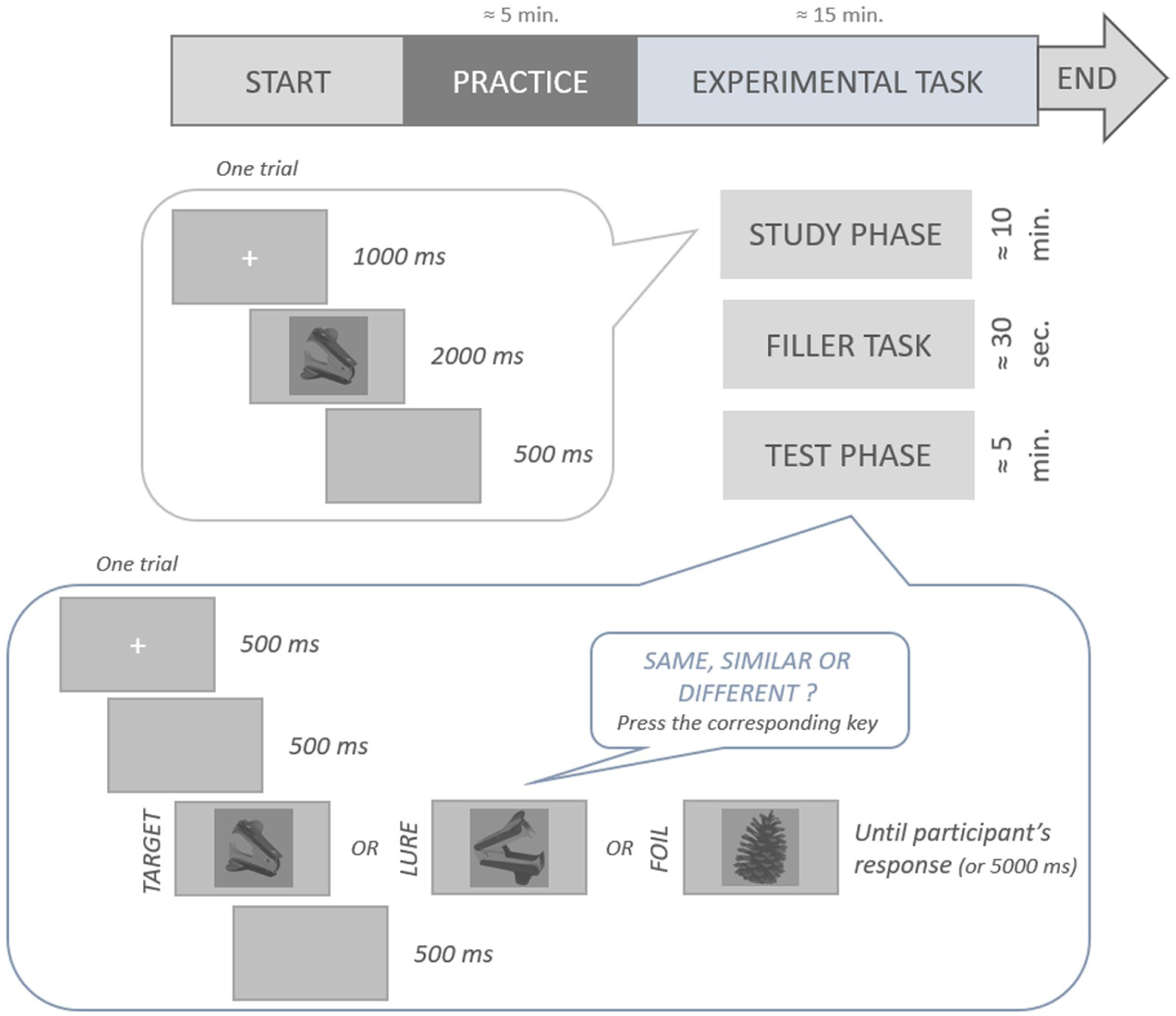
Schematic procedure of the MST adapted.
Finally, participants were informed that they would perform the main task, which would be identical to the practice session, except that the number of images to be memorized would be much larger, and no feedback would be provided. The study phase of the main task consisted of 96 images (32 for each PASS FILTER TYPE: NF, HSF, and LSF), while the test phase included 144 images (48 for each FILTER TYPE, with 16 for each trial type: TARGET, LURE, and FOIL). The filters applied to each image were counterbalanced between participants. After the study phase and prior to the test phase, participants completed a filler task, which involved counting backward from 400 by 3 for 30 s. The trials in both phases were identical to the practice session, except that no feedback was given.
Data Processing
The data were analyzed in R version 4.4.1, using RStudio software (R Core Team, 2018), in accordance with the analyses specified in the preregistration. Additional non-preregistered analyses were also conducted on response frequencies to better characterize response profiles for each trial type, particularly false recognitions for lure items. As the preregistration did not specify corrections for post hoc comparisons, we proceeded as follows: for post hoc analyses based on a priori hypotheses, a one-tailed test was used, while a two-tailed test was applied when there were no a priori hypotheses, with p-values corrected using the Holm method. The mean correct response rate (mCR), the mean response frequencies, and the mean response times for correct responses (mRT) were computed across participants for each experimental condition. Moreover, the mean LDI was calculated across all participants based on the probability of “similar” responses to lure items minus the probability of “similar” responses given to the foils (Stark et al., 2019). Response times exceeding 3,000 ms and below 300 ms were considered as late or premature responses (outliers) and were therefore excluded from the analyses (see Mille et al., 2023a for a similar procedure). The remaining data were then screened for outliers by experimental condition using the median absolute deviation (MAD; Leys et al., 2013, 2019), with a cut-off threshold set at 3. The average data exclusion rate was 3.97%, with the lowest exclusion rate being 0% for the LURE × LSF condition and the highest exclusion rate being 7.28% for the TARGET × HSF condition. As specified, participants whose mCR fell below the chance level were to be excluded from the analysis. However, no participant met this exclusion criterion. Similarly, for response accuracy, outliers were identified using the MAD, with a cut-off threshold set at 3, by experimental condition based on the mCR. Five participants were identified as outliers based on this criterion. Consequently, analyses concerning mCR, response frequency, and the LDI were conducted on 33 participants. In contrast, since response time analyses relied on a different indicator, these participants were retained for the analysis of mRT, resulting in mRT analyses being performed on all 38 participants.
Three ANOVAs were performed, one on mCR, one on mRT, and one on response frequency. Each analysis was conducted with the within-subject factors “PASS FILTER TYPE” (NF, LSF, and HSF) and “TRIAL TYPE” (TARGET, FOIL, and LURE). A fourth ANOVA was conducted on the LDI with the within-subject factor “PASS FILTER TYPE” (NF, LSF, and HSF). As with the first three ANOVAs, a one-tailed test was used for analyses based on a priori hypotheses, while a two-tailed test was applied for analyses without a priori hypotheses. The normality of the distributions of the dependent variables was assessed visually, which confirmed the absence of bimodality. As a result, no data transformation was applied. Regarding sphericity, the afex package (Singmann et al., 2012) was used for the analyses, and when the sphericity assumption was violated (Mauchly’s test), the Greenhouse–Geisser correction was automatically applied by the package. A threshold of 0.05 was used for all analyses after correction.
Results
Mean correct response rate
We first examined the effect of PASS FILTER TYPE and TRIAL TYPE on the mCR. The analyses revealed (see Figure 3) a main effect of PASS FILTER TYPE (F[1.95, 62.37] = 22.99, p < .001, η2 G = .068) on mCR. Post hoc analysis with two-tailed tests showed that the mCR was significantly lower in the LSF condition (M = 0.52; SD = 0.28; 95% CI [0.47, 0.58]) than the HSF condition (M = 0.59; SD = 0.24 [0.55, 0.64]; t[32] = 4.31, p < .001) and the NF condition (M = 0.64; SD = 0.26 [0.59, 0.68]; t[32] = −6.86, p < .001). Additionally, the mCR was significantly lower in the HSF condition than in the NF condition (t[32] = −2.51, p = .017).
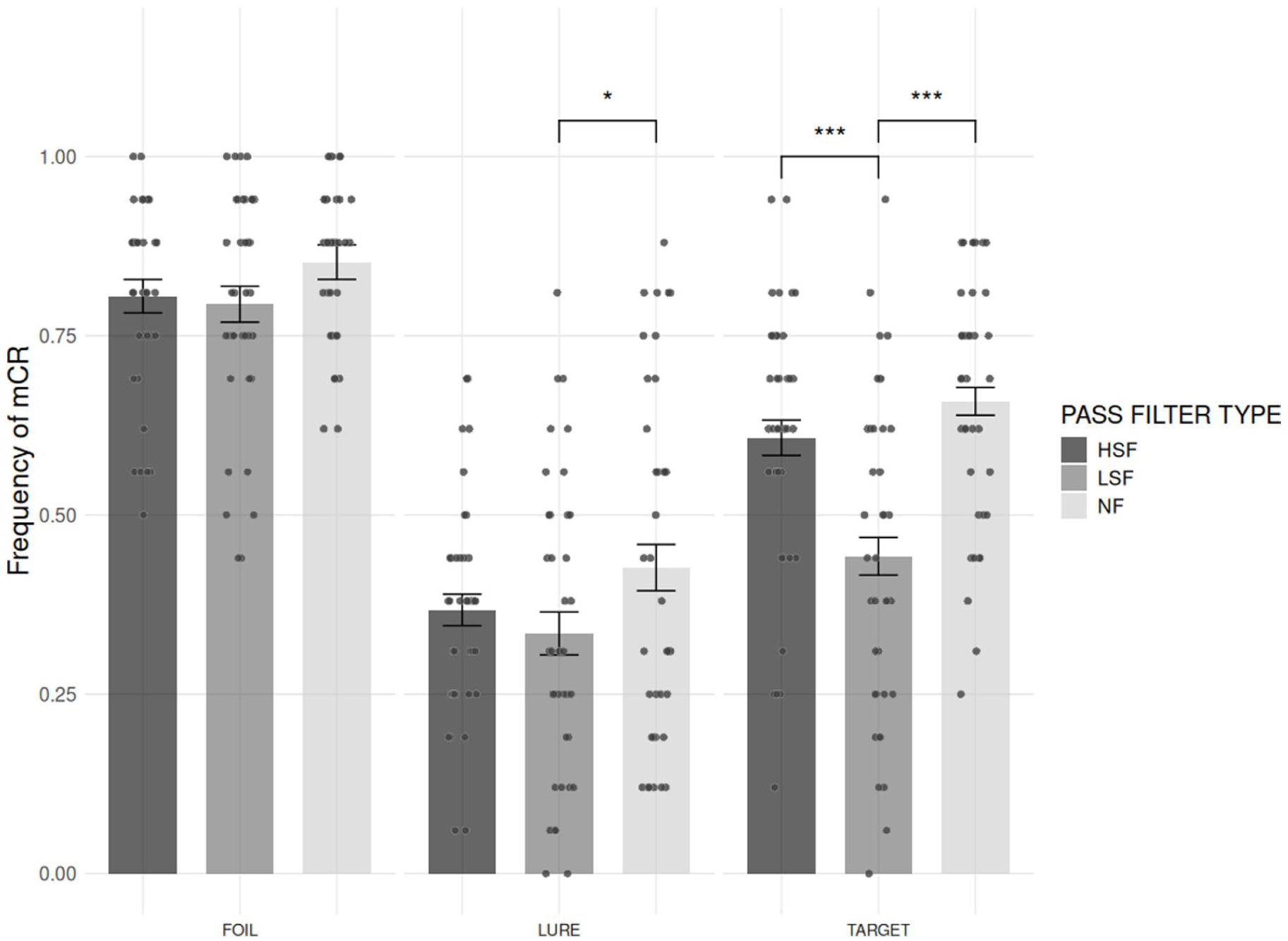
Comparison of the frequency of mCR for each TRIAL TYPE (FOILS, LURE, and TARGET) and PASS FILTER TYPE (HSF, LSF, and NF). Gray bars represent standard errors corrected for the within-participant design. Significance levels are indicated as follows: *p < .05. **p < .01. ***p < .001.
The results also showed a main effect of TRIAL TYPE (F[1.85, 59.26] = 106.98, p < .001, η2 G = .518) on mCR. Post hoc analysis with two-tailed tests showed that the mCR was significantly lower for the LURES (M = 0.38; SD = 0.21; 95% CI [0.34, 0.42]) than for the FOILS (M = 0.82; SD = 0.22 [0.77, 0.83]) trials (t[32] = 13.13, p < .001), for the TARGETS (M = 0.57; SD = 0.22 [0.53, 0.61]) than for the FOILS trials (t[32] = 7.71, p < .001), and for the LURES than for the TARGETS trials (t[32] = −7.94, p < .001).
The two-way interaction PASS FILTER TYPE × TRIAL TYPE was also significant (F[3.05, 97.51] = 5.12, p = .002, η2 G = .032). In line with our hypothesis that the PASS FILTER TYPE should only affect the mCR of LURES, the bilateral post hoc analyses revealed no effect of the PASS FILTER TYPE on the FOILS (for all comparisons, p > .1). However, in contrast to our hypothesis, these same analyses revealed a PASS FILTER TYPE effect for the TARGET, showing that the mCR was significantly higher in the NF condition and the HSF condition compared to the LSF condition (t[32] = −6.19, p < .001 and t[32] = 5.46, p < .001, respectively) while no significant difference was observed between the NF and HSF conditions (t[32] = −1.97, p = .057). Finally, regarding the effect of PASS FILTER TYPE for the LURES, the hypotheses were partially confirmed, showing that the mCR was significantly higher in the NF condition compared to the LSF condition (t[32] = −2.33, p < .039, one-tailed test), while no significant difference was observed between the HSF and the LSF conditions (t[32] = 1.16, p = .374, one-tailed test), nor between the HSF and the NF conditions (t[32] = −1.47, p = .306, two-tailed test).
Response Frequencies
In addition to analyzing mean correct response rate, we conducted an exploratory (non-preregistered) analysis of response frequencies for each TRIAL TYPE (LURE, TARGET, and FOIL) across each PASS FILTER TYPE (HSF, LSF, and NF) to provide a more detailed characterization of participants’ response patterns.
To better characterize false recognitions on LURE trials, we first examined response frequencies across PASS FILTER TYPE for this TRIAL TYPE. The analyses revealed (see Figure 4) no main effect of PASS FILTER TYPE (F[2.00, 63.93] = 0.50, p = .611, η2 G = .001) on response frequencies. However, there was a significant main effect of RESPONSE TYPE (F[1.60, 51.28] = 4.49, p = .023, η2 G = .087) on response frequencies. Post hoc analyses with two-tailed tests showed that DIFFERENT responses (M = 0.37; SD = 0.21; 95% CI [0.33, 0.41]) were given significantly more often than SAME responses (M = 0.25; SD = 0.15 [0.22, 0.28]; t[32] = 2.88, p = .014), and SIMILAR responses (M = 0.37; SD = 0.21 [0.33, 0.41]) were also given more often than SAME responses (t[32] = −3.09, p = .012). In contrast, the frequency of DIFFERENT and SIMILAR responses did not differ significantly, t(32) = −0.09, p = .924.
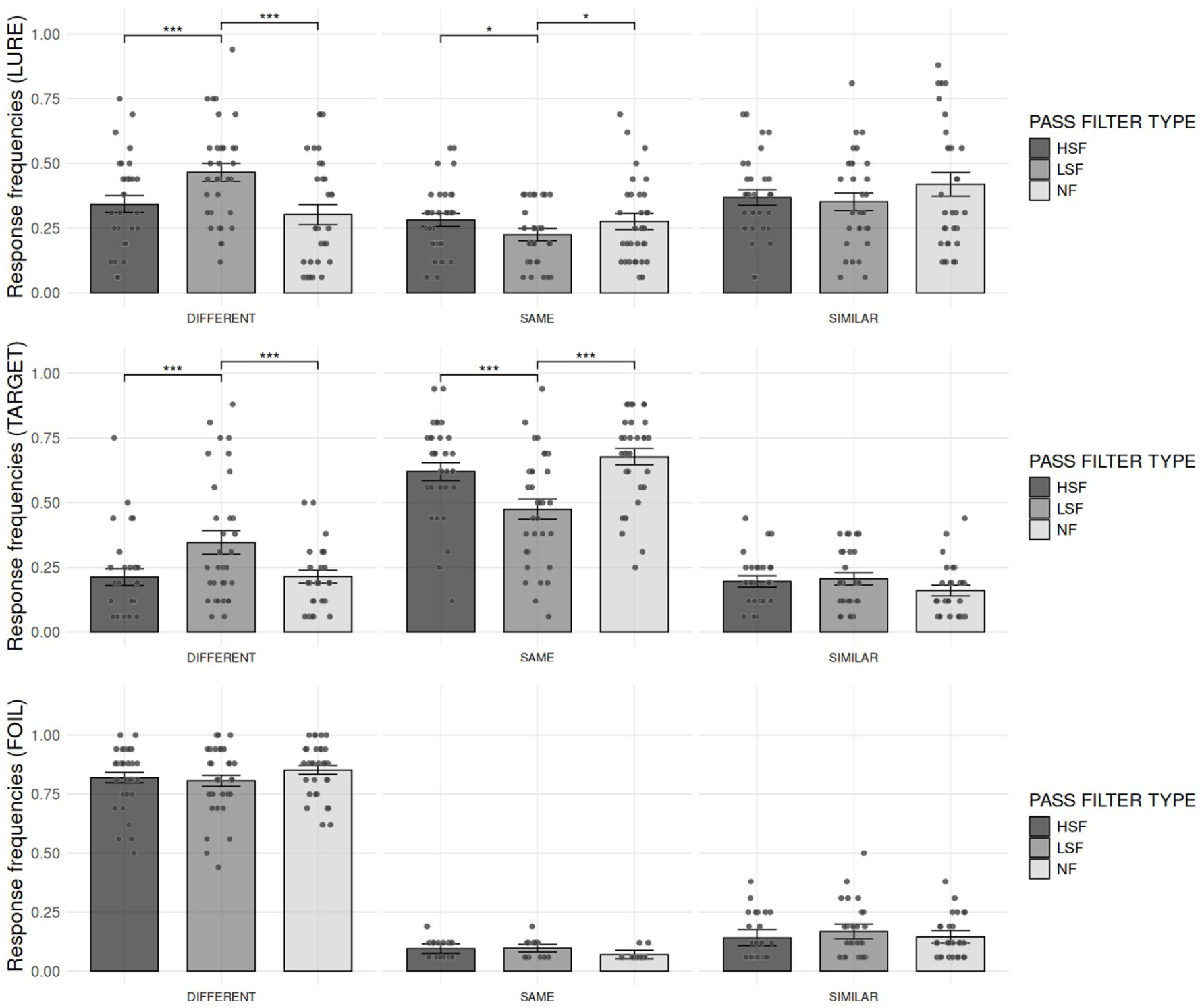
Comparison of the mean response frequencies for each PASS FILTER TYPE (HSF, LSF, and NF) across the three trial types: LURES, TARGETS, and FOILS. Bars represent the mean frequency per response type, with standard errors corrected for the within-participant design. Significance levels are indicated as follows: *p < .05. **p < .01. ***p < .001.
The two-way interaction PASS FILTER TYPE ×RESPONSE TYPE was also significant (F[3.57, 114.20] = 8.40, p < .001, η2 G = .076). Post hoc analyses revealed that DIFFERENT responses were significantly more frequent in the LSF condition than in the HSF condition (t[32] = −4.30, p < .001), as well as in the LSF condition compared to the NF condition (t[32] = 5.61, p < .001). In contrast, the comparison between the HSF and NF conditions did not reach significance (t[32] = 1.86, p = .072). The analysis also revealed that SAME responses were significantly more frequent in the HSF condition compared to the LSF condition (t[32] = 2.85, p = .023), and in the NF condition compared to the LSF condition (t[32] = −2.75, p = .023). In contrast, the comparison between the HSF and NF conditions did not reach significance (t[32] = 0.20, p = .841). For SIMILAR responses, post hoc analyses indicated that response frequencies did not differ significantly across filter types (HSF vs. LSF: t[32] = 1.16, p = .306; HSF vs. NF: t[32] = −1.47, p = .306; LSF vs. NF: t[32] = −2.33, p = .079). Taken together, contrary to our hypothesis, this pattern of results suggests on the one hand an increase in lure rejections in the LSF condition compared to both the HSF and the NF conditions, and on the other hand a reduction in false recognitions for lures in the LSF condition compared to the HSF and the NF conditions.
Then, to better characterize miss of recognitions of TARGETS, we first examined response frequencies across PASS FILTER TYPE for this TRIAL TYPE. The analyses revealed (see Figure 4) no main effect of PASS FILTER TYPE (F[1.70, 54.54] = 0.16, p = .815, η2 G = .001) on response frequencies. However, there was a significant main effect of RESPONSE TYPE (F[1.27, 40.56] = 53.51, p < .001, η2 G = .535). Post hoc analyses with two-tailed tests showed that SAME responses (M = 0.59; SD = 0.22; 95% CI [0.54, 0.63]) were given significantly more often than DIFFERENT responses (M = 0.24; SD = 0.21 [0.20, 0.28]; t[32] = −6.14, p < .001), and SAME responses were given significantly more often than SIMILAR responses (M = 0.17; SD = 0.11 [0.15, 0.20]; t[32] = 11.32, p < .001). However, the difference between SIMILAR and DIFFERENT responses (t[32] = 2.04, p = .051) did not reach significance. The two-way interaction PASS FILTER TYPE × RESPONSE TYPE was also significant (F[3.47, 110.89] = 17.92, p < .001, η² G = .148). Post hoc analyses revealed that DIFFERENT responses were significantly more frequent in the LSF condition than in the HSF condition (t[32] = −6.02, p < .001), as well as in the LSF condition compared to the NF condition (t[32] = 4.86, p < .001). In contrast, the comparison between the HSF and NF conditions did not reach significance (t[32] = 0.43, p = .673). For SAME responses, post hoc analyses indicated that they were significantly more frequent in the HSF condition compared to the LSF condition (t[32] = 5.46, p < .001), and in the NF condition compared to the LSF condition (t[32] = −6.19, p < .001). The comparison between the HSF and the NF conditions did not reach significance (t[32] = −1.97, p = .057). For SIMILAR responses, post hoc analyses showed that response frequencies did not differ significantly across filter types (HSF vs. LSF: t[32] = 0.11, p = .917; HSF vs. NF: t[32] = 1.90, p = .200; LSF vs. NF: t[32] = 1.57, p = .251). Taken together, contrary to our hypothesis, this pattern of results indicates that participants made more misses in the LSF condition than in the HSF and the NF conditions.
Finally, to better characterize false recognitions on FOIL trials, we examined response frequencies across PASS FILTER TYPE for this TRIAL TYPE. The analyses revealed (see Figure 4) no main effect of PASS FILTER TYPE (F[1.91, 61.17] = 1.81, p = .173, η² G = .001) on response frequencies, and the two-way interaction PASS FILTER TYPE × RESPONSE TYPE was also not significant (F[2.62, 83.88] = 1.80, p = .160, η² G = .021). In contrast, there was a significant main effect of RESPONSE TYPE (F[1.19, 38.10] = 661.65, p < .001, η² G = .926). Post hoc analyses with two-tailed tests showed that DIFFERENT responses (M = 0.83; SD = 0.13; 95% CI [0.80, 0.85]) were given significantly more often than SAME responses (M = 0.04; SD = 0.05 [0.03, 0.05]; t[32] = 35.79, p < .001), and DIFFERENT responses were also given significantly more often than SIMILAR responses (M = 0.13; SD = 0.11 [0.10, 0.15]; t[32] = 22.15, p < .001). Moreover, SAME responses were given significantly more often than SIMILAR responses (t[32] = −6.10, p < .001). Consistent with our hypotheses, this pattern of results indicates that PASS FILTER TYPE did not affect participants’ responses for FOILS trials.
Lure Discrimination Index
To further assess visual lure discrimination performance, we computed the LDI scores, which offer a more specific index of the trace memory discrimination process. The analyses revealed (see Figure 5) a main effect of PASS FILTER TYPE (F[1.90, 60.68] = 4.43, p < .041, η2 G = .038] on LDI. The post hoc analyses partially supported our hypotheses. The LDI was significantly lower in the LSF condition (M = 0.178; SD = 0.214; 95% CI [0.10, 0.254]) than in the NF condition (M = 0.282; SD = 0.256 [0.191, 0.373]; t[32] = −2.37, p = .035, one-tailed test), consistent with our predictions. The absence of a significant difference between the HSF (M = 0.246; SD = 0.185 [0.18, 0.31]) and NF conditions (t[32] = −0.87, p = .389, two-tailed test) was also in line with expectations. In contrast, the lack of a significant difference between the LSF and HSF conditions (t[32] = 1.91, p = .098, one-tailed test) was contrary to our hypotheses.
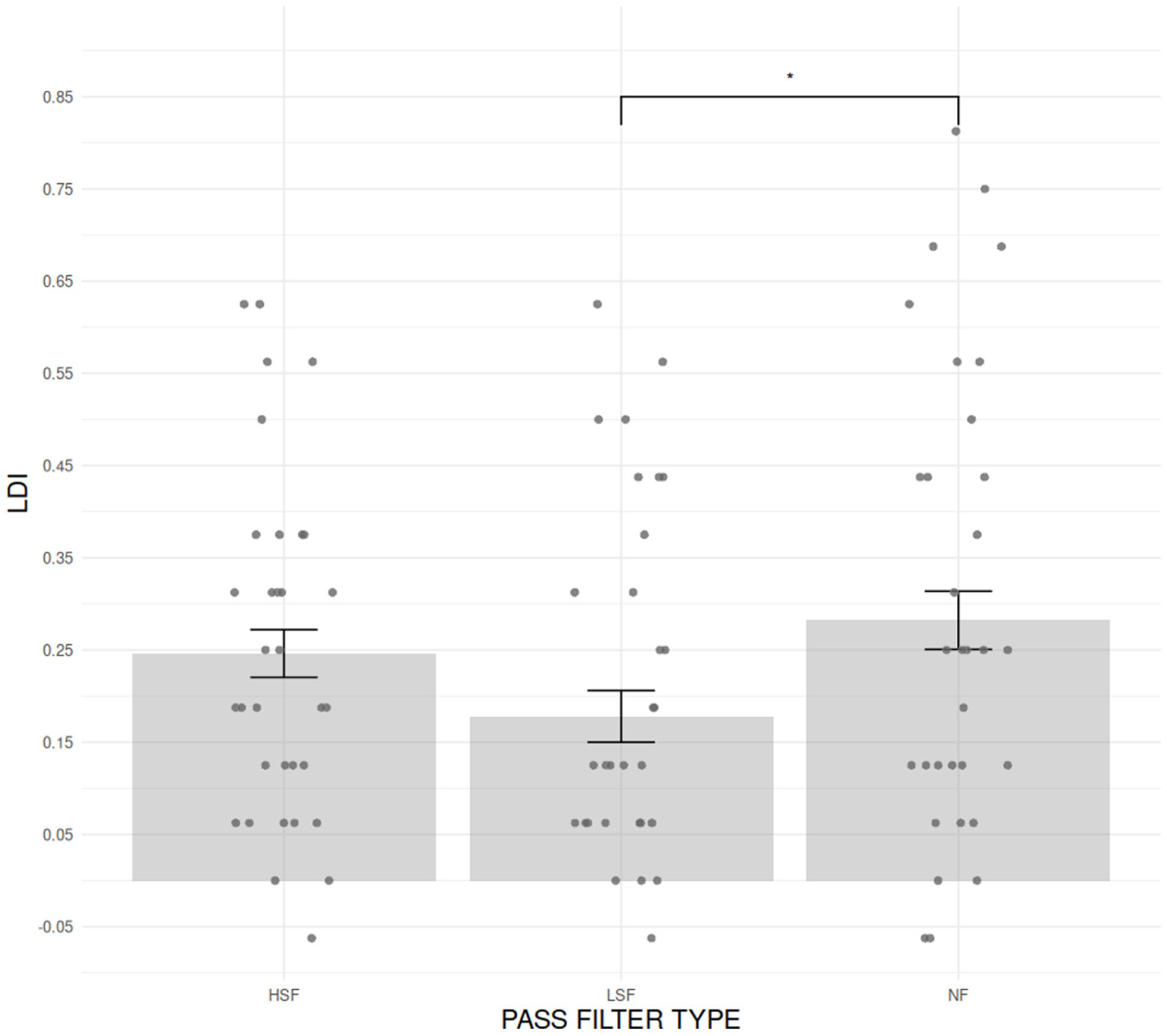
Comparison of the mean LDI for each PASS FILTER TYPE (HSF, LSF, and NF). Gray bars represent standard errors corrected for the within-participant design. Significance levels are indicated as follows: *p < .05. **p < .01. **p < .001.
Mean response times for correct responses
Finally, we examined the effect of PASS FILTER TYPE and TRIAL TYPE on the mRT. 3 The analyses revealed (see Figure 6) a main effect of PASS FILTER TYPE (F[1.91, 63.17] = 19.67, p < .001, η2 G = .070) on mRT. Post hoc analysis with two-tailed tests showed that the mRT was significantly slower in the LSF condition (M = 1,528; SD = 348; 95% CI [1,346, 1,461]) compared to the HSF condition (M = 1,404; SD = 311 [1,346, 1,461]; t[33] = −4.06, p < .001) and the NF condition (M = 1,374; SD = 320 [1,314, 1,433]; t[33] = 6.48, p < .001). The comparison between the HSF and the NF conditions did not reach significance (t[33] = 1.64, p = .109).
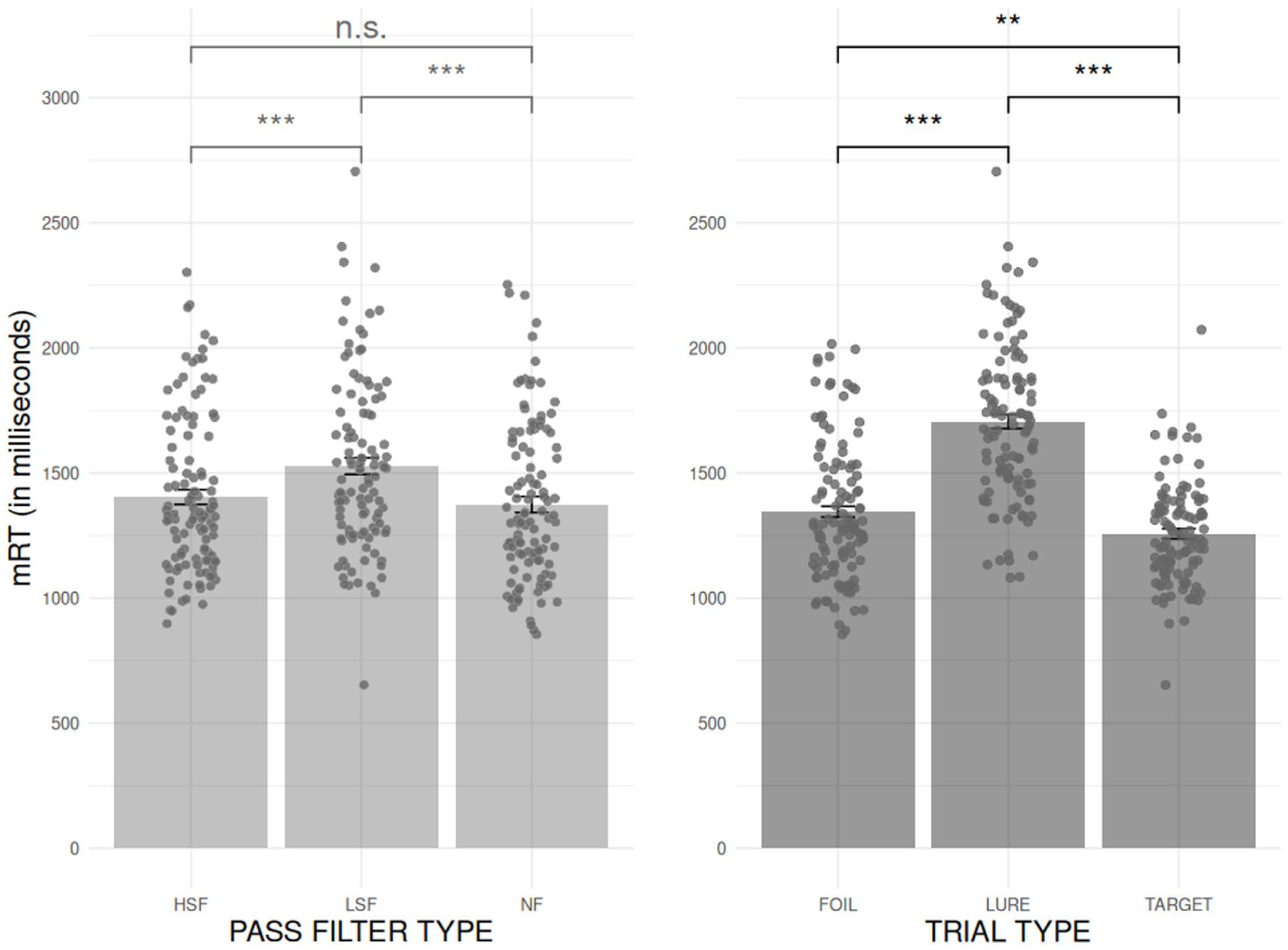
Comparison of the mRT for each PASS FILTER TYPE (HSF, LSF, and NF, graph on the left) and TRIAL TYPE (FOIL, LURE, and TARGET, graph on the right). Gray bars represent standard errors corrected for the within-participant design. Significance levels are indicated as follows: *p < .05. **p < .01. ***p < .001.
The analysis also showed a main effect of TRIAL TYPE (F[1.90, 62.72] = 79.80, p < .001, η2 G = .362) on mRT. Post hoc analysis with two-tailed tests showing that the mRT was significantly slower for the LURES (M = 1,705; SD = 319; 95% CI [1,645, 1,765]) than for the FOILS (M = 1,345; SD = 279 [1,294, 1,397]; t[33] = −8.54, p < .001), for the FOILS than for the TARGETS (M = 1,257; SD = 204 [1,219, 1,295]; t[33] = 3.35, p < .002), and for the LURES than for the TARGETS (t[33] = 11.69, p < .001). The two-way interaction PASS FILTER TYPE × TRIAL TYPE was not significant (F[3.24, 106.79] = 1.14, p = .34, η2 G = .005).
Discussion
The purpose of this study was to investigate the role of the spatial frequency content of visual stimuli in the discrimination of overlapping memory traces. Using the MST (Stark et al., 2019), which assesses the capacity to distinguish visually similar objects, participants completed the task under three different spatial frequency filtering conditions. These included a NF condition, a control in which spatial frequency was not manipulated; a LSF condition, with a low-pass filter primarily emphasizing the coarse structure of the stimuli; and a HSF condition, with a high-pass filter primarily emphasizing the fine details of the image.
First, despite the introduction of spatial frequency filters, the classic effects of trial type in the MST were replicated. Specifically, lure items resulted in significantly lower recognition performance and longer response times compared to targets and foils (e.g., Davidson et al., 2018; Stark et al., 2010, 2019). Next, the results for the main measure (i.e., the LDI) were broadly in line with our central hypothesis, which posited that LSF were less involved in the fine-grained discrimination of mnemonic traces than the HSF. Indeed, the LDI was significantly lower in the LSF condition compared to the NF condition, whereas there was no significant difference between the HSF and the NF conditions. This finding suggests that the LSF condition, by providing predominantly coarse and global visual information, may limit the reconstruction of sufficiently distinctive visual information necessary for effective mnemonic discrimination. This result is particularly noteworthy given that some stimuli could be distinguished based on simple perceptual cues (e.g., item rotation), yet the overall reduction in LDI in the LSF condition remained significant. This is consistent with the observation that LSF images did not moreover lead to an increase in false recognitions of foils. This supports the effectiveness of experimental manipulation using visuo-spatial filters.
These results suggest that the LSF may promote a global and less precise processing mode, which could lead to reduced efficiency in the PS mechanism. This interpretation aligns with the visual perception literature, where the LSF are rather associated with fast, global processing, whereas the HSF are more involved in detailed, and focused analysis (e.g., Kauffmann et al., 2015; Ramanoël et al., 2015). These results suggest that the type of visuo-spatial frequency may contribute to the nature of mnemonic traces. According to an influential theory on access to precise memories, fuzzy trace theory (Reyna & Brainerd, 1995), mnemonic precision cannot rely solely on a global (gist) encoding and requires the formation of detailed (verbatim) traces, as fine discrimination between similar stimuli is facilitated by these detailed representations (e.g., Mille et al., 2023b; Racsmány et al., 2021). Within this framework, the observed results on the LDI can be interpreted as suggesting that LSF may favor encoding more oriented toward gist, whereas HSF may support the encoding of verbatim traces. It should be noted, however, that no significant differences were observed either between the NF and the HSF conditions or between the LSF and the HSF conditions after Holm correction. These results suggest that the HSF alone are not sufficient to fully support mnemonic discrimination, reflecting the fact that they do not contain the full range of spectral information necessary. This finding is consistent with recent studies suggesting that the processing of spatial frequencies is less strictly dichotomous. Specifically, neither the LSF nor the HSF are always sufficient on their own for effective visual discrimination, and successful processing often relies on a combination of both types of frequencies (Ashtiani et al., 2017; Farhang et al., 2025).
Although this study was not designed to identify the brain regions involved, and caution is therefore warranted when interpreting the results in terms of neural substrates, our data nevertheless appear to be consistent with the hierarchical representational approach (Murray & Bussey, 1999; Saksida & Bussey, 2010). According to this approach, mnemonic discrimination relies on hierarchical visual processing, from the occipital cortex to memory-related structures like the perirhinal cortex, culminating in more complex processing in the hippocampus. In this context, HSF images would provide important visual details for effective PS, helping to avoid confusion with memory traces. In contrast, the degradation of perceptual details through LSF images could compromise these processes. These results contribute to theoretical models of memory by showing that rich sensory input, especially the HSF, is important for providing detailed visual information to memory-related structures. They also suggest that PS may not be exclusively limited to hippocampal structures but may also depend on the quality of sensory information upstream. This implies a connection between visuo-mnemonic discrimination within the ventral visual pathway and PS beyond the involvement of hippocampal structures (Amer & Davachi, 2023; Kent et al., 2016). The processing of these frequencies may rely on a network that includes the transentorhinal cortex, known to serve as an interface between perceptual regions and the hippocampus (Bastin & Delhaye, 2023). This region may play a key role in selecting and routing high-detail visual inputs to structures involved in PS.
Nevertheless, although the reduction in mCR for lures in the LSF condition may also indicate a decrease in mnemonic discrimination, the observations regarding both false recognition of lures and correct target identifications suggest that a simple reduction of mnemonic discrimination via the LSF is insufficient to account for the overall findings. Indeed, contrary to our hypotheses, the LSF condition was associated not only with a decrease in false recognitions of lures but also with an increase in misses for targets. These observations suggest a more general impoverishment of memory traces during encoding, likely due to less detailed visual information. In this regard, we could not ensure that all items were identifiable at a semantic level across the different spatial frequency conditions. Although the pre-test confirmed that each target-lure pair was visually discriminable, the absence of control over semantic identifiability may have limited the encoding of the items’ conceptual properties, in turn reducing mnemonic performance (e.g., Craik & Tulving, 1975; Pidgeon & Morcom, 2014) in the LSF condition. Future studies should aim to disentangle the respective contributions of semantic and purely visual factors to memory performance under different spatial frequency conditions. This could be achieved, for instance, through a finer manipulation of spatial frequency or the use of non-nameable stimuli, allowing a clearer distinction between perceptual and conceptual contributions to mnemonic performance.
Another, non-mutually exclusive interpretation could be that LSF images led participants to adopt a more conservative response criterion, meaning that items were identified as previously seen only when the associated memory signal was sufficiently strong. This hypothesis is consistent, on the one hand, with the decrease in false recognition of lures and the increase in target misses observed in the LSF condition, and on the other hand, with the overall slowing of response times in this condition. However, this interpretation should be considered with caution, as the use of a three-response format in the MST task does not allow for the direct computation of response criterion indices within the framework of signal detection theory (see Stark et al., 2019, for discussion), on the basis of which this hypothesis could be confirmed. Future studies should therefore employ binary response formats to test this interpretative hypothesis more directly.
At a methodological level, several limitations should be acknowledged. First, the present study used static images and applied a uniform manipulation of spatial frequencies across the entire image, which may not necessarily reflect the dynamic and localized processing of visual information in natural conditions (e.g., Roberts et al., 2022). Future studies should therefore explore whether these effects persist in more ecologically valid tasks or in other sensory modalities. The present study was also conducted only on inanimate objects. It is therefore impossible to conclude on living objects that may be processed differently under spatial frequency filtering (e.g., Farhang et al., 2025). New study should explore how spatial frequencies contributed to memory discrimination for both living and non-living objects. Another limitation is that most foil trials came from a different image set (Poporo) than the original MST stimuli set. However, this does not appear to have affected the classic trial type effects (Stark et al., 2019), demonstrating the robustness of the task despite the use of these stimuli, and the key comparisons between filter conditions remain valid because the associations between filters and stimuli were properly counterbalanced. Other limitations include the sample size, which was estimated based on data on the effects of visuo-spatial filtering from a different task (i.e., indoor/outdoor image categorization, Ramanoël et al., 2015), rather than on a pilot study using the MST itself, due to the lack of preliminary data. In addition, our sample consisted primarily of young university students with a notable gender imbalance, which may limit the generalizability of the findings. Future replications, with larger and more gender-balanced samples, using the effect size observed here, would allow a more precise assessment of the reliability, generalizability, and representativeness of the results.
Finally, our results are based solely on behavioral data, which limits the ability to identify the underlying brain structures involved in the effect of filters on mnemonic performance. Neuroimaging studies, involving functional magnetic resonance imaging or electroencephalography, would offer a clearer picture of the specific neural circuits activated during the processing of HSF and LSF in the MST. Further research is therefore needed to confirm the respective roles of spatial frequencies in the MST, as well as to clarify the underlying neural mechanisms. Based on our behavioral results, such studies could include representational similarity analyses (Kriegeskorte et al., 2008) to test specific hypotheses regarding neural similarity patterns between encoding and retrieval, for example expecting reduced similarity for stimuli presented with LSF.
Conclusion
To our knowledge, this preregistered study is the first to evaluate the role of spatial frequencies in the MST. The results suggest that the HSF play a greater role in mnemonic discrimination than LSF. However, other observations indicate that the LSF may also induce a general impoverishment of memory traces, reflected in a decrease in lure false recognitions and an increase in target misses, which would be consistent with the hypothesis of less detailed visual encoding. Further studies are needed to confirm these results and to clarify the behavioral and neural mechanisms involved.
This study offers important insights from both a fundamental and applied perspective. Theoretically, it sheds new light on the links between perception and memory, reinforcing the idea that the quality of perceptual processing conditions the success of complex mnemonic processes (Kent et al., 2016; Mille et al., 2021; Versace et al., 2009, 2014). These results could also help in better understanding the decline in mnemonic performance observed during aging, particularly in the MST, by suggesting that a deficit in HSF processing could be one of the underlying mechanisms (Leal & Yassa, 2018; Mille et al., 2021). From a clinical perspective, this approach paves the way for the development of new methods for assessing memory impairments in populations with perceptual alterations, such as patients with dementia, neurodevelopmental disorders, or older adults. The manipulation of spatial frequencies could thus serve as a relevant tool to dissociate perceptual encoding deficits from true mnemonic impairments.
Supplemental Material
sj-docx-1-qjp-10.1177_17470218261456719 – Supplemental material for Do Visual Details Drive Memory? Role of the Visuo-Spatial Frequencies in Mnemonic Discrimination
Supplemental material, sj-docx-1-qjp-10.1177_17470218261456719 for Do Visual Details Drive Memory? Role of the Visuo-Spatial Frequencies in Mnemonic Discrimination by Jordan Mille, Stephen Ramanoël and Guillaume T. Vallet in Quarterly Journal of Experimental Psychology
Footnotes
Acknowledgements
We would like to thank the students who helped with participants’ recruitment and data collection.
Ethical Considerations
This study was carried out in accordance with the recommendations of the French Law (Loi Jardé no. 2012-300) with written informed consent being obtained from all the subjects in accordance with the Declaration of Helsinki. An ethics approval was issued for the current study by the ethics committee of Tours-Poitiers University (number CER-TP 2023-12-02).
Consent to Participate
Written informed consent was obtained from all participants prior to their participation in the study, after they had been provided with information about the objectives and procedures of the research.
Author Contributions
JM participated in all the aspects of the study (design of the experiments, conduct of the experiments, analysis and interpretation of the data, and redaction of the manuscript). SR and GTV participated in the design of the experiments, interpretation of the data, and redaction of the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.