
Abstract
Working memory consolidation (WMC) is known to consume central attentional resources, impairing performance on subsequent attention-demanding tasks. However, the specific nature of this impairment, particularly its effect on the initial stage of visual selection, remains unclear. The current study tested three competing hypotheses: No-suppression, Pure-temporal-suppression, and Spatial-suppression hypotheses. We used a sequential target-matching task where the inter-stimulus interval (ISI) between a target item and a subsequent probe item was varied to modulate the state of consolidation. Experiment 1 revealed that a spatial cue before the probe item failed to guide attention during consolidation (10-ms ISI) but did so effectively after consolidation was complete (1,000-ms ISI), thus ruling out the No-suppression hypothesis. In contrast, Experiment 2 showed that when a larger target item encompassed the cue’s location, the cueing effect was restored during consolidation (0 and 150-ms ISIs), a finding that argues against the Pure-temporal-suppression hypothesis. Taken together, these findings support the Spatial-suppression hypothesis, revealing that visual selection during WMC is governed by an attentional window whose size is flexibly tied to the consolidated representation. This work clarifies the nature of the attentional bottleneck in WMC, reframing it as a spatial phenomenon.
Introduction
Working memory is a cognitive system that provides a “mental scratchpad” for temporarily storing and processing information. This system is crucial for higher-order cognitive tasks such as learning, decision-making, and problem-solving. Working memory involves four core processes: encoding (forming representations), consolidation (stabilizing representations), maintenance (maintaining representations), and retrieval (accessing representations). Among these, working memory consolidation (WMC) is a cognitive process of converting transient sensory inputs into stable working memory representations (Ricker & Hardman, 2017). WMC plays a critical role in the memory processes, as it directly affects the performance of subsequent memory retention (Bayliss et al., 2015; Nieuwenstein & Wyble, 2014).
Previous studies have shown that WMC is a process that requires attention (Ricker et al., 2018). During this process, other cognitive activities that also require attention may be impaired (Jolicœur & Dell’Acqua, 1998). A typical phenomenon reflecting this impairment is the attentional blink (AB), as many studies consider WMC as the cause of AB (Chun & Potter, 1995; Ricker & Hardman, 2017; Wyble et al., 2009). In a typical AB paradigm, researchers present two target stimuli (e.g., two digits) within a rapid serial presentation of visual stimuli (e.g., letters). Participants are asked to report the identity of these two targets in the order in which they were presented. The results typically show that participants can accurately report the first target (T1). However, they have difficulty correctly reporting the identity of the second target (T2) if the two targets are separated by 200 to 500 ms (Broadbent & Broadbent, 1987; Chun & Potter, 1995; Raymond et al., 1992).
However, it is still unclear which stage of T2 processing is disrupted by T1 consolidation during the AB. As an attentional-demanding task, T2 processing can be divided into two stages (Cunningham & Wolfe, 2014). The first is selection, in which a subset of the visual input is prioritized and attention is deployed. The second is post-selection processing of the selected information, including identification and comparison with a target template. Accordingly, deficits in T2 reporting during T1 consolidation could arise from two mechanisms: (a) impaired visual selection of T2, or (b) intact selection with impairment at the post-selection stage. These alternatives warrant further examination because, in many AB paradigms, the spatial location of T2 varies across trials (e.g., Ghorashi et al., 2009; Karabay et al., 2022; Zivony & Lamy, 2016). Consequently, successful T2 reporting necessarily entails selecting its spatial location before identification.
Some studies suggest that visual selection may be unaffected during the AB. For example, in Experiment 3 of Zivony and Lamy (2016), participants had to identify T1 and T2 in two rapid serial visual presentation (RSVP) streams. These streams were presented simultaneously on the left and right sides, each 3° away from the center of the screen. The T1 was a pair of simultaneously presented digits of the same color. The T2 was a letter with the same color as T1. Participants first reported the identity of T2 and then judged whether the two T1 digits were the same or different (this judgment needed to be consolidated and maintained in working memory). Before T2 onset, a square cue with the same or a different color as T1/T2 would randomly appear in the stream on the same side as T2 or on the opposite side. The results showed that same-colored cues produced a spatial cueing effect, where T2 accuracy was significantly higher when the cue and target were on the same side. Critically, the cueing effect did not differ between the within-blink (short T1–T2 interval) and post-blink conditions (long T1–T2 interval). Ghorashi et al. (2009) found similar results. In their study, participants memorized two letters that appeared sequentially. T1 and T2 appeared randomly in two of four peripheral locations, each 2° from central fixation. A spatial cue could appear at the location where T2 would appear 94 ms before T2 onset. The results showed that this cue led to a performance benefit, and this benefit was the same during and after the AB. These findings all suggest that visual selection and the consolidation process causing the AB might be two independent, non-interfering processes.
However, this conclusion is challenged on both theoretical and experimental grounds. First, the conclusion seems inconsistent with the predictions of some mainstream theories in the field of spatial attention. For example, guided search divides information processing in visual search into three stages: visual selection, target identification, and comparison with a target template (Cunningham & Wolfe, 2014; Wolfe, 2021). This theory posits that visual selection is a serial process that depends on attentional control. Logically, such a serial process would occupy attentional resources and therefore should be impaired by the consolidation process, which also requires attention. The threaded cognition theory (Taatgen et al., 2009) offers a similar view. This theory conceptualizes the AB not as a passive resource bottleneck but as an overexertion of control, implemented through a production rule that actively blocks target detection to protect the ongoing consolidation process, thereby blocking the visual selection of subsequent stimuli.
There is also experimental evidence inconsistent with the conclusion that visual selection and consolidation do not interfere. In a study by Du and Abrams (2009), participants had to identify two sequentially presented letters. T1 appeared in a central RSVP stream, while T2 appeared 9° to the left or right of the screen’s center. A spatial cue with 50% validity appeared 100 ms before T2 onset. The results showed that the cueing effect completely disappeared at short T1 to T2 intervals. However, the cueing effect was robust at long T1 to T2 intervals. This result seems inconsistent with the findings of Ghorashi et al. (2009) and Zivony and Lamy (2016). It suggests that WMC impairs the visual selection of subsequent items.
Taken together, the interaction mechanism between WMC and visual selection remains controversial. The present study aims to further investigate this issue by testing three possible theoretical hypotheses. The first is the No-suppression hypothesis. This hypothesis posits that WMC and visual selection are two independent cognitive processes that do not mutually interfere (e.g., Ghorashi et al., 2009; Zivony & Lamy, 2016). According to this hypothesis, the visual selection mechanism can still operate normally even during WMC. Potential targets that appear at new locations can still be selected.
The second hypothesis is the pure-temporal-suppression hypothesis. This hypothesis posits that WMC interferes with visual selection within a specific temporal window (e.g., Du & Abrams, 2009). Specifically, consolidation suppresses the spatial deployment and shifting of attention either via active cognitive control (e.g., Taatgen et al., 2009; Wyble et al., 2009) or a resource bottleneck (e.g., Wolfe, 2021). As a result, visual selection is impaired and spatial cueing effects should no longer be present. In fact, the Pure-temporal-suppression hypothesis itself is one of the more classic explanations of the AB phenomenon (Taatgen et al., 2009).
Notably, both hypotheses described above are essentially temporal in nature. That is, they focus on what does or does not occur during the time course of WMC. However, there remains another possibility: time may not be the sole determining factor. We therefore propose a third hypothesis: the spatial-suppression hypothesis. According to this hypothesis, WMC affects visual selection in the spatial domain. Specifically, the attentional window might be restricted to a relatively small size during consolidation (e.g., Jefferies et al., 2023; Müller et al., 2005). This restriction affects the range of visual selection (see also Belopolsky & Theeuwes, 2010; Belopolsky et al., 2007; Tamber-Rosenau et al., 2025; Theeuwes, 1994, 2023a, 2025). Therefore, when the cue and T2 are spatially close to T1, they are more likely to fall within this window and induce a spatial cueing effect. This hypothesis can explain the different findings in previous studies. In studies reporting that visual selection is unaffected by WMC (Ghorashi et al., 2009; Zivony & Lamy, 2016), the spatial distance between T1 and the spatial cue was relatively small (<4°). The cue and T2 were likely to be within the window, so a significant cueing effect was observed. In contrast, in the study reporting that visual selection is impaired by WMC (Du & Abrams, 2009), the T1-cue distance was relatively large (9°). The cue and T2 were more likely to be outside the window, so no cueing effect was observed. Importantly, the Spatial-suppression hypothesis does not deny the possible existence of temporal suppression mechanisms. It merely posits that time is not the only factor, and that spatial and temporal influences may jointly operate during WMC. Within this framework, the pure-temporal-suppression and spatial-suppression hypotheses make mutually exclusive claims: the former posits that suppression is entirely time-based, whereas the latter claims that suppression cannot be explained by temporal factors alone.
In the present study, we conducted two experiments to test the three competing hypotheses. Participants were required to perform a sequential target-matching task. In this task, a target item (which needed to be consolidated into working memory) was presented, followed by a probe item. A valid or invalid spatial cue was presented before the probe item. Experiment 1 tested the hypothesis that WMC does not affect visual selection. The results were similar to those of Du and Abrams (2009). Specifically, the spatial cueing effect disappeared during WMC. This finding ruled out the No-suppression hypothesis. Experiment 2 further tested the Pure-temporal-suppression and Spatial-suppression hypotheses. Building on Experiment 1, we used a target item with a larger size to enlarge the size of the attentional window during consolidation (e.g., Belopolsky et al., 2007; Lee & Jeong, 2020). Under this condition, we found that the consolidation process did not affect the cueing effect. This result further ruled out the Pure-temporal-suppression hypothesis and supported the Spatial-suppression hypothesis.
Experiment 1
Experiment 1 aimed to test whether WMC has an effect on visual selection (i.e., the No-suppression hypothesis). Observers performed a sequential target-matching task. We manipulated the inter-stimulus interval (ISI) between the target item and the spatial cue (10 ms vs. 1,000 ms) before the probe item. To ensure that participants could successfully encode the target and initiate consolidation, 1 we used a staircase procedure to determine the minimal exposure duration required for the target item (details below). Moreover, prior research has shown that consolidation is not completed within 10 ms after mask onset (Nieuwenstein & Wyble, 2014). Therefore, this manipulation created two conditions: one with consolidation ongoing (10-ms ISI) and one with consolidation completed (1,000-ms ISI). Within each ISI condition, we also manipulated the spatial distance between the cue and the target item (1.5°, 3°, or 4.5°). If the ISI duration modulated the magnitude of the cueing effect at any spatial distance (i.e., an interaction between ISI and cue validity), it would suggest that WMC affects visual selection. Conversely, if WMC does not affect visual selection, then no interaction between ISI and cue validity should be observed at any distance. Additionally, if an effect of WMC on visual selection was present, the different target-cue distances would allow us to explore the spatial boundary of this effect.
Method
Observers
We employed a Bayesian sequential design which allowed for flexible data collection with the option to terminate at any stage based on specified statistical criteria (Berger & Wolpert, 1988; Rouder, 2014). Our research protocol specified a minimum sample size of 24 observers, with data collection continuing until the primary analysis (ISI × validity interaction) achieved a Bayes-factor threshold (BF10 >5 or <0.2; Schönbrodt et al., 2017) or until reaching a maximum sample size of 50 observers. The final sample comprised 50 students (18 female) recruited from Zhejiang University, who participated in exchange for course credit or compensation. They ranged in age from 18 to 34 years (M = 22.0, SD = 4.3) and all reported normal or corrected-to-normal vision.
Task and Stimuli
Observers performed a sequential target-matching task (Figure 1a), in which a target digit (visual size 1° × 1°) was first shown at the center of the screen, followed by a mask. They were asked to judge whether a probe digit matched this target digit. The probe digit (visual size 1° × 1°) was shown at a random position on an invisible circle centered around the target digit. Before the probe digit appeared, a square cue (visual size 1.2° × 1.2°) was shown on the screen. Its position could either overlap with the probe digit (valid cue) or be symmetrically opposite to the probe digit relative to the screen center (invalid cue). The validity of the cue in both experiments was 50%. Two independent variables were manipulated: the ISI between the target digit and spatial cue (10 or 1,000 ms), and the distance between the target digit and the probe digit (1.5°, 3°, or 4.5°; Figure 1b). A frame rate of 100 Hz was used. The viewing distance was fixed at 60 cm.
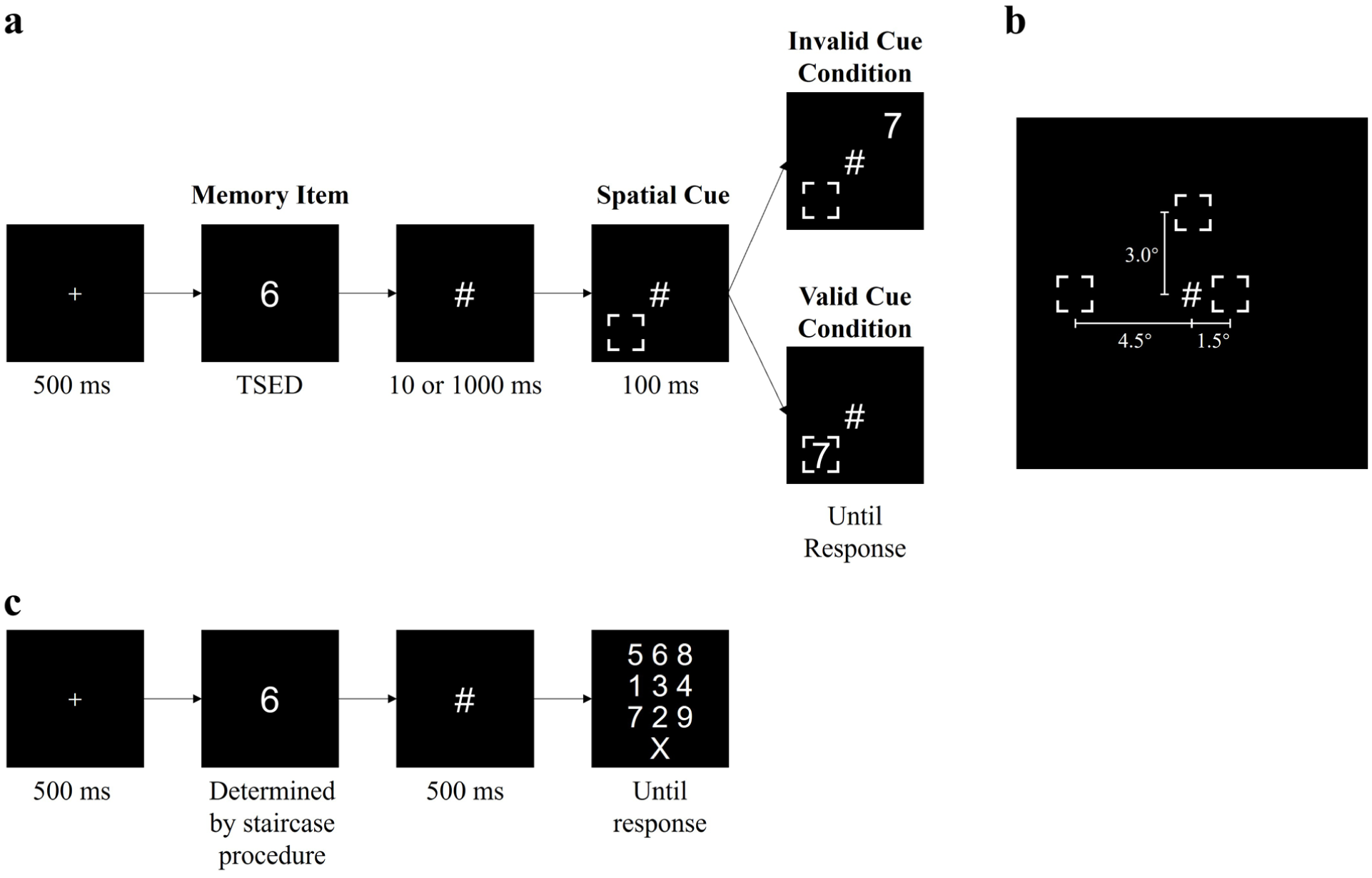
The experimental paradigm, stimuli, and procedure used in Experiment 1. (a) Stimuli and procedure of the sequential target-matching task. (b) A schematic diagram of the three target item-spatial cue distance conditions. (c) Stimuli and procedure of the task for determining TSEDs.
Design and Procedure
Each trial began with a 500 ms fixation point, followed by the presentation of the target digit. The exposure duration of the target digit was determined by a staircase procedure (details below). The digit was masked by a “#” symbol. The mask then remained on the screen for an ISI of 10 or 1,000 ms, after which the spatial cue was displayed on an invisible circle centered around the target digit. Then, the probe digit appeared 100 ms 2 after the cue onset. The probe item was either inside the cue or at a position symmetric to the cue relative to the screen center. Observers pressed the right arrow key if the probe digit matched the target digit and the left arrow key if it did not. They were instructed to respond as quickly as possible while maintaining a high response accuracy. No response time limit was imposed. The experiment adopted a within-subject block design, consisting of a total of 576 trials divided into 6 blocks (2 ISIs × 3 spatial distances), with the order of blocks randomized for each observer. Each block was preceded by a practice phase: each practice session included four trials, and if the accuracy was less than 100%, the practice was repeated until a session with 100% accuracy rate was achieved. The experiment lasted about 40 min for each observer.
Staircase Procedure
We first determined the threshold stimulus exposure duration (TSED; Li et al., 2018, 2019) of the target digit (i.e., the minimum stimulus exposure time required for identifying the target digit; (Figure 1c). The purpose of measuring TSED was to minimize the exposure time of the target digit while still ensuring successful memory encoding. If the exposure time was too short, consolidation could not begin due to encoding failure. If the exposure time were too long, consolidation might have already begun or even completed before mask onset. If so, even the shortest ISI (10 ms) might no longer reflect an ongoing consolidation state. The TSED measurement procedure was as follows: At the beginning of each trial, a fixation point was displayed at the center of the screen for 500 ms, followed by the presentation of the digit, whose duration was determined by the following staircase procedure. The digit was then masked by a “#” symbol for 500 ms, after which a 3 × 3 array of randomly arranged digits was displayed. Observers were asked to select the previously presented digit using the mouse, or choose the “X” symbol if uncertain.
To obtain the TSED, a 4-down-1-up staircase procedure was employed: that is, the initial exposure time of the digit was 60 ms, which decreased by 10 ms after four consecutive correct responses and increased by 10 ms after a wrong response or an UNCERTAIN response. A turning point was defined when the direction of two consecutive time changes reversed. The staircase procedure terminated after 10 turning points, and the TSED value was calculated as the average of the last 4 turning points. If the exposure time decreased to 10 ms and the observer achieved nine consecutive correct responses, the procedure also stopped, and the TSED was considered to be 10 ms.
Data Analysis
In this experiment, we used response time to access the cueing effect. In the data analysis, we planned to exclude observers with an accuracy rate below 75% in the sequential target-matching task (but no observers were excluded based on this criterion) and only retained trials with correct responses for further data analysis. Additionally, trials with response times shorter than 100 ms or longer than 1,500 ms were excluded. Following these steps, a total of 9.45% of the trials were excluded. 3
All data preprocessing was performed using R 4.1.1 (R Core Team, 2021). Statistical inference was conducted using JASP 0.19.1 (JASP Team, 2024). To strengthen the methodological rigor, we reported both Bayesian and frequentist analysis results.
Results
The mean exposure time of the target digit, measured using the staircase procedure, was 39.7 ms (SD = 10.5 ms), and the overall accuracy rate of the sequential target-matching task was 93.3% (SD = 4.5%). Figure 2 shows the mean response times under different conditions in the sequential target-matching task. To examine the influence of WMC on spatial cueing effect, we conducted a three-way ANOVA with cue validity × ISI × cue distance. For each participant and condition, we computed mean response times and entered these means into the repeated-measures ANOVA. The results show moderate evidence supporting the interaction between ISI and cue validity, BFinclusion = 5.41, F(1, 49) = 9.16, p = .004, η2 p = .158, strong evidence against the interaction between cue distance and cue validity, BFinclusion = 0.08, F(2, 98) = 0.958, p = .387, η2 p = .019, and ambiguous evidence for the three-way interaction, BFinclusion = 1.11, F(2, 98) = 2.43, p = .093, η2 p = .047. These results indicate that the spatial cueing effect was not affected by the distance between the target digit and the spatial cue. However, there were notable differences in the spatial cueing effect between the short-ISI condition and long-ISI condition, suggesting that WMC can affect visual selection.
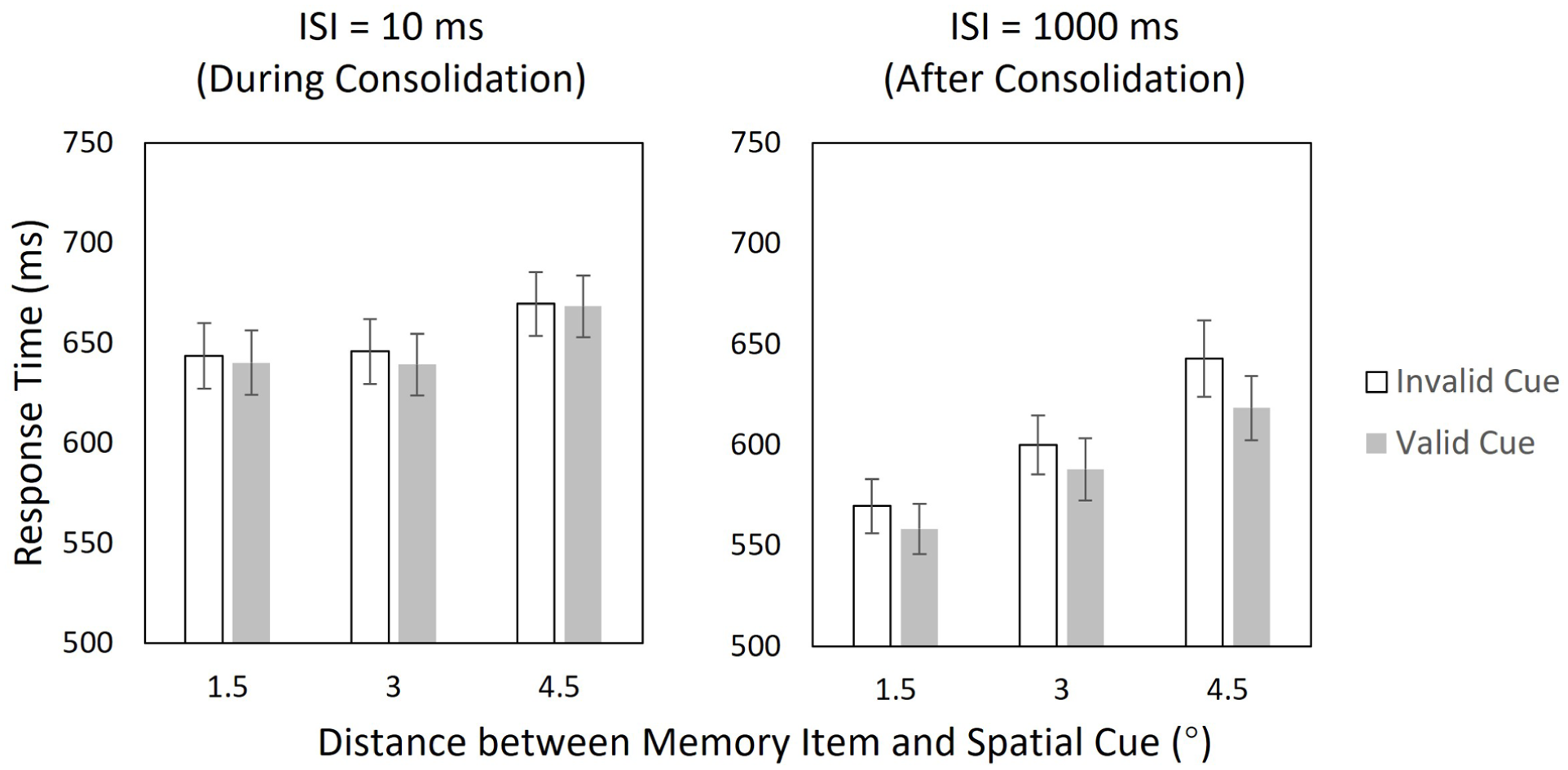
Results of Experiment 1. Response times for valid and invalid cues across different distances during (left) and after (right) consolidation. Error bars represent the 95% within-subjects confidence interval (Morey, 2008).
Based on the above results, we further conducted two-way ANOVAs (cue validity × cue distance) for the short- and long-ISI conditions respectively. The results revealed that, during consolidation (i.e., ISI = 10 ms), there was moderate evidence against the main effect of cue validity, BFinclusion = 0.31, F(1, 49) = 1.14, p = .291, η2 p = .023, moderate evidence supporting the main effect of cue distance, BFinclusion = 7.20, F(2, 98) = 5.42, p = .006, η2 p = .100, and strong evidence against the interaction between cue validity and cue distance, BFinclusion = 0.100, F(2, 98) = 0.474, p = .624, η2 p = .010. In contrast, after the completion of consolidation (i.e., ISI = 1,000 ms), there was strong evidence supporting the main effects of cue validity, BFinclusion = 5.96 × 10³, F(1, 49) = 34.62, p < .001, η2 p = .414, strong evidence supporting the main effects of cue distance, BFinclusion = 7.18 × 107, F(2, 98) = 29.13, p < .001, η2 p = .373, and weak evidence against the interaction between cue validity and cue distance, BFinclusion = 0.79, F(2, 98) = 2.81, p = .065, η2 p = .054. These results show that the spatial cueing effect was absent during consolidation but present after consolidation.
Discussion
The results of Experiment 1 indicate that the effect of a spatial cue on the response time (RT) of sequential digit matching is strongly affected by the status of WMC. While a clear cueing effect emerged once consolidation was completed, it was absent when the consolidation process was ongoing, regardless of the target-cue distance. The results challenge the No-suppression hypothesis, which predicts that the cueing effect should appear across all the tested ISIs and target-cue distances.
Experiment 2
The results of Experiment 1 demonstrated that visual selection can be impaired by WMC, but it is unclear whether this impairment is spatial or temporal in nature. Because the target digit subtended only 1° × 1° of visual angle, the ensuing attentional window was likely comparably small, placing all tested cue locations outside its boundaries. To adjudicate between spatial- and pure-temporal-suppression hypotheses, we employed a refined experimental design in Experiment 2. While maintaining the same memory encoding and consolidation requirements as Experiment 1, we increased the physical size of the central target item so that its spatial extent would encompass the peripheral cue locations, thereby inducing a large attentional window (e.g., Castiello & Umiltà, 1990; Gmeindl et al., 2020; Lee & Jeong, 2020; Jefferies et al., 2023; van Moorselaar et al., 2025). This design generates different predictions from the two hypotheses. If the suppression of visual selection during WMC is temporal and irrelevant to spatial location, the spatial cueing effects should remain absent under short-ISI conditions. Conversely, if the suppression is spatially constrained, the cues will now fall within the boundary of the attentional window, and thus the spatial cueing effects should appear under short-ISI conditions.
Method
Observers
In Experiment 2, we employed the same sample plan as used in Experiment 1. We recruited a new set of 24 observers (17 female) from Zhejiang University. They were aged from 18 to 25 years (M = 21.8, SD = 1.70) and had normal or corrected-to-normal visual acuity.
Task and Stimuli
The task and stimuli were similar to those used in Experiment 1, with three differences. First, both the target digit and its corresponding mask were larger (subtended a visual angle of 5.56° × 5.56°). Second, the target digit-to-spatial cue distance was fixed at 2° of visual angle. This spatial arrangement ensured that the cue always appeared within the spatial boundaries defined by the large central memory item and its mask (see Figure 3). 4 Third, three ISIs between target digit and spatial cue (0, 150, or 500 ms) were used. We selected these ISIs to systematically test the cueing effects. The 0-ms ISI was the earliest possible time point. At this moment, consolidation has just begun after mask onset. The 150-ms ISI also falls within the consolidation stage (Jolicœur & Dell’Acqua, 1998; Ricker & Hardman, 2017). At 500-ms ISI, consolidation was expected to be complete. This is consistent with Jolicœur & Dell’Acqua (1998, Experiment 6). Their experiment used a similar single-memory-item paradigm, and the consolidation time was about 350 ms.

Spatial configuration of stimuli in Experiment 2. The memory digit and its mask subtended a visual angle of 5.56° × 5.56°, whereas the probe digit was 1° × 1° and was shown 2° away from the screen center.
Design and Procedure
Experiment 2 employed a 3 (ISI: 0, 150, and 500 ms) × 2 (cue validity: valid, invalid) within-subject design, with all testing conditions randomly mixed within the experiment. The experiment consisted a total of 300 trials. The sequential target-matching task procedure and the staircase procedure remained identical to those used in Experiment 1.
Results and Discussion
The data exclusion and analysis criteria were the same as in Experiment 1. 5 The mean exposure time of the target digit, determined by the staircase procedure, was 32.5 ms (SD = 10.1 ms). The accuracy rate in the sequential target-matching task was 88.4% in average (SD = 5.4%). Following the same exclusion criteria as in Experiment 1, 15.5% of trials were excluded from analysis. 6 Mean response times for the sequential target-matching task across conditions are shown in Figure 4. A two-way ANOVA (3 ISIs × 2 cue validities) revealed moderate evidence for the main effect of cue validity, BFinclusion = 6.91, F(1, 23) = 17.89, p < .001, η2 p = .437, strong evidence for the main effect of ISI, BFinclusion = 4.49 × 1015, F(2, 46) = 112.55, p < .001, η2 p = .830, and moderate evidence against the ISI × cue validity interaction, BFinclusion = 0.13, F(2, 46) = 0.128, p = .880, η2 p = .006. These results demonstrated faster responses for valid versus invalid cues across all ISI conditions, indicating robust spatial cueing effects in all testing conditions. Obviously, these findings are compatible with the spatial-suppression hypothesis: that is, when the physical size of memory items was enlarged to encompass the cue locations, the attentional window extended correspondingly, enabling spatial attention to reliably select the cue during WMC.
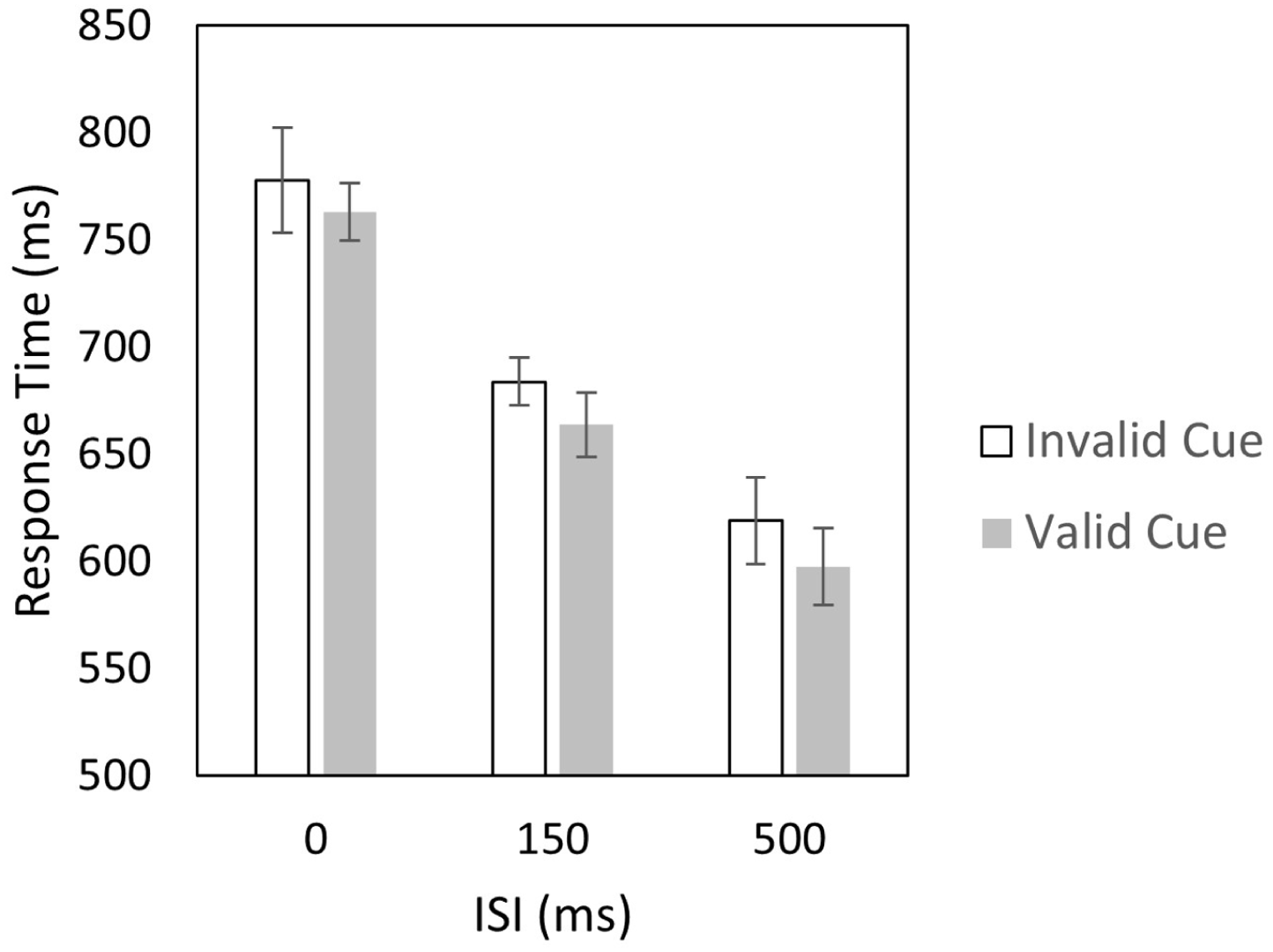
Results of Experiment 2. Response times for valid and invalid cues across different ISIs between the target digit and the spatial cue. Error bars represent the 95% within-subjects confidence interval (Morey, 2008).
General Discussion
The present study was designed to investigate whether WMC impairs visual selection (the initial stage of information processing) for a concurrent task. To investigate this question, we proposed three theoretical hypotheses: the No-suppression, Pure-temporal-suppression, and Spatial-suppression hypotheses. We tested these hypotheses using a sequential target-matching paradigm and examining the spatial cueing effects under different conditions. In Experiment 1, we used a physically small target item (1° × 1°). The spatial cue was presented outside the area covered by this item (target-cue distance: 1.5°, 3°, or 4.5°). The results showed that no spatial cueing effect was found during consolidation under any cue distance condition. This finding argued against the No-suppression hypothesis. In Experiment 2, we used a physically large target item (5.56° × 5.56°). The spatial cue was presented within the area covered by this item (target-cue distance: 2°). The results showed a spatial cueing effect under all three ISI conditions, which argued against the Pure-temporal-suppression hypothesis. Furthermore, the results of Experiment 2 also suggested that new stimuli could still be selected by spatial attention as long as they appeared within the attentional window. The results of both experiments jointly support the Spatial-suppression hypothesis, which suggests that the suppression of visual selection by WMC is spatial. During WMC, visual selection can only occur within the attentional window.
Prior studies have typically used two paradigms to examine attentional selection during WMC. One is the RSVP paradigm (Du & Abrams, 2009; Zivony & Lamy, 2016), in which a stream of distractors appears at the same location as the target items. The other is the dwell time paradigm (Ghorashi et al., 2009), originally developed by Duncan et al. (1994). In this paradigm, targets appear sequentially at different locations, and each target is followed by a mask. Our focus was not on the processing or filtering of distractor streams. Instead, we aimed to isolate WMC, and a simpler stimulus sequence better serves this goal. We therefore adopted a task that is closer to the dwell time paradigm. Notably, the dwell time and RSVP paradigms typically require participants to report both targets. In contrast, our task requires a matching judgment on two sequentially presented stimuli. This change makes consolidation of the first target a core requirement of the task. Participants cannot ignore the first target and must consolidate the first target into working memory to perform the judgment correctly.
Despite these differences, our results replicate key findings from both approaches. Experiment 1 replicated the absence of a cueing effect during WMC reported by Du and Abrams (2009). Experiment 2 replicated the presence of a cueing effect during WMC reported by Ghorashi et al. (2009) and Zivony and Lamy (2016). These findings suggest that the paradigm itself does not determine whether a cueing effect emerges.
These findings also provide new insights into the mechanism of attention allocation during WMC. Previous research has established that WMC constitutes a central processing bottleneck, impairing performance on subsequent tasks (e.g., Jolicœur & Dell’Acqua, 1998; Ricker & Hardman, 2017; but see Nieuwenstein & Wyble, 2014). This bottleneck has often been attributed to a non-spatial-suppression or resource occupation mechanism. In this view, attentional resources are occupied or suppressed during the consolidation process and thus cannot be deployed to new external tasks. However, this explanation is incomplete as it ignores the influence of spatial factors, which may be the source of contradictory findings in previous literature. Some studies found that even when T2 processing was impaired by the ongoing T1 processing, a spatial cueing benefit still persists. Crucially, the magnitude of this benefit was equivalent to conditions without such impairment (Ghorashi et al., 2009; Zivony & Lamy, 2016). This finding may be because the spatial cue fell within the participant’s attentional window, which is consistent with Experiment 2 of the present study. In contrast, other studies found that a spatial cue preceding T2 provided no benefit when attentional resources were occupied by T1 (Du & Abrams, 2009). The result may stem from the cue lying beyond the attentional window, consistent with Experiment 1. Together, our findings reconcile prior discrepancies by showing that attention during WMC is spatially dependent and constrained by an attentional window. Notably, even for stimuli within the attentional window, the present study still found longer response times in the short-ISI conditions. 7 This suggests that while performance on a concurrent task is generally impaired during WMC, the nature of this impairment is spatially dependent. For stimuli within the attentional window, the impairment occurs at the post-selection stage, leaving visual selection unaffected. In contrast, for stimuli outside the attentional window, the impairment begins at the initial stage of information processing (i.e., visual selection), and also affects the post-selection stage. In other words, WMC appears to affect attention allocation through a dual mechanism: it spatially gates whether attention can be deployed to select new stimuli, and it temporally delays the post-selection processing of the selected stimuli.
In previous studies, the attentional window account was used to explain the absence of attentional capture effect in some conditions (Belopolsky & Theeuwes, 2010; Belopolsky et al., 2007; Theeuwes, 2023a, 2023b). A long-standing finding in attention research is that physically salient stimuli can capture attention (Schreij et al., 2008, 2010; Theeuwes, 1994; Theeuwes et al., 2003; Yantis & Hillstrom, 1994), even if they are task-irrelevant. However, some studies have also found that salient stimuli do not capture attention under some conditions (e.g., Belopolsky et al., 2007; Gaspelin et al., 2015). The attentional window account was proposed to reconcile these seemingly contradictory findings by positing a spatial boundary for attentional capture (Theeuwes, 2023a). According to this account, only salient stimuli falling inside the attentional window can capture attention, while salient stimuli outside the window will be ignored. This claim has received empirical support from a series of studies (i.e.,Belopolsky & Theeuwes, 2010; Belopolsky et al., 2007).
The results of the present study are consistent with the classical attentional window account. However, it is noteworthy that our paradigm employed a task-relevant and informative spatial cue. After cue onset, observers know exactly that it will occur either inside the cue or at the spatially opposite position on the circle (instead of at any location on the circle). Therefore, observers were motivated to utilize this cue. This results in a mixture of top-down and bottom-up processing in our task, unlike previous studies about attentional windows that often aim to test purely bottom-up mechanisms. Our spatial-suppression account therefore provides an important extension to the attentional window account, suggesting its applicability extends beyond attentional capture in visual search. The window also appears to govern spatial cueing effects that involve both bottom-up and top-down components within a working memory paradigm. In Experiment 1, even though we used an informative and abrupt onset cue, which was considered to possess greater attentional capture advantage (see Zhang et al., 2025 for a review), cues outside the attentional window still failed to be selected by attention. In contrast, Experiment 2 showed that the cues inside the attentional window can be selected by attention.
It could be argued that participants might have voluntarily expanded their attentional window to better exploit the predictive cue. However, the exposure duration of the memory item was very brief. This required efficient processing of the target digit, and a smaller attentional window is known to support higher processing efficiency (Castiello & Umiltà, 1990). More importantly, if such a strategy were effective, a cueing effect during consolidation should have also been observed in Experiment 1. Yet it was not. This suggests that participants did not strategically enlarge their attentional window.
The present results also align with previous studies showing that when attention is highly focused, observers are less able to detect salient targets in the periphery. Joseph et al. (1997) demonstrated that even feature searches for targets with unique orientations—usually considered to operate independently of focused attention—are impaired when observers must simultaneously detect a target in a centrally presented RSVP stream. This suggests that directing attention to the center creates a narrowly focused attentional window, which limits the ability to detect peripheral targets.
Conclusion
In conclusion, the present findings suggest that visual selection is spatially constrained within an attentional window surrounding the memory item during WMC. Consequently, a spatial cue can only be effective when it falls within the boundaries of this window.
Footnotes
Ethical Considerations
The experimental protocol received prior approval from the local research ethics committee.
Consent to Participate
Written informed consent was obtained from all observers before they began the experiments.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a grant (#32071043) from the National Natural Science Foundation of China and by Zhejiang Key Laboratory of Neurocognitive Development and Mental Health (Grant No. 2025E10037).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.