
Abstract
In general, non-sprinters (team sport athletes) exhibit lower sprint performance than sprinters. The differences in spatiotemporal and ground reaction force (GRF) variables between non-sprinters and sprinters have been reported previously. The differences may not simply reflect sprint ability but rather sport-specific characteristics, implying variation in joint kinetics. Therefore, this study aimed to examine the differences in lower limb joint kinetics during the stance phase of maximal speed sprinting between non-sprinters and sprinters. Twenty-six male non-sprinters (13 soccer and 13 baseball players) and 12 male sprinters participated. Sprint data around the maximal speed phase were measured using a motion capture system and force plates. Kinetic variables of the lower limb joints and GRFs during the stance phase were compared between the two groups. As a result, maximal sprint speed was lower in non-sprinters (8.53 ± 0.38 m/s) than in sprinters (9.62 ± 0.25 m/s). The vertical and anteroposterior GRFs throughout the stance phase were smaller in non-sprinters than in sprinters. Moreover, compared with sprinters, non-sprinters exhibited smaller ankle plantarflexion torques but larger knee and hip extension torques. The results indicate that knee and hip extension torques compensate for insufficient ankle plantarflexion torque in generating vertical and anteroposterior GRFs in non-sprinters. The differences in sprint mechanics between non-sprinters and sprinters might be influenced by differences in sports specificities. Therefore, increasing ankle plantarflexion torque may improve maximal-speed sprint performance in non-sprinters; however, the direct transfer of this suggested modification to short sprints involving frequent changes of direction requires careful consideration.
Introduction
Sprint performance is essential to scoring points and goals1,2 for non-sprinters (team sport athletes) such as soccer, basketball, and baseball players. In particular, soccer players frequently sprint over a 20-m distance (45% per total sprints), 3 and baseball players sprint over 27.431 m (the distance between bases) after hits. Thus, improving long-sprint ability, along with increasing maximal sprint speed, not only short-distance sprint ability, is crucial for non-sprinters. In general, non-sprinters exhibit lower maximal sprint speeds and inferior acceleration ability compared with sprinters. Thus, identifying the factors contributing to the inferior sprint performance of non-sprinters and the mechanism by which these factors differ between sprinters and non-sprinters would be crucial for developing effective sprint training programs.
Previous studies identified the differences in spatiotemporal and ground reaction force (GRF) variables between non-sprinters (mainly soccer players) and sprinters.4–6 At the maximal speed phase, non-sprinters exhibited longer contact times and smaller vertical and anteroposterior GRFs than sprinters,4,6 indicating that non-sprinters had a lower ability to generate larger GRFs within a short time, which is a key feature of superior sprint performance.7,8 These differences were identified at the same sprint speed (8.0, 8.5, and 9.0 m/s): non-sprinters exhibited smaller net anteroposterior force, as well as smaller braking and propulsive forces, and smaller vertical force than sprinters.5,6 Non-sprinters also had an inferior force application technique, with a smaller ratio of force 5 (the ratio of anteroposterior force to the resultant force 9 ) than sprinters. Moreover, a previous study reported that non-sprinters exhibited significantly different lower limb joint kinematics during the stance and swing phases compared with sprinters even at the same maximal sprint speed (non-sprinters, 9.31 ± 0.19 m/s; sprinters, 9.34 ± 0.21 m/s). 10 In their study, non-sprinters exhibited a larger range of leg extension motion from contact to toe-off during the stance phase than the sprinters, indicating the “ground-based” strategy for non-sprinters in contrast to the “front-side” kinematic strategy for sprinters. 10 These observed differences imply that the differences in joint kinematics and GRF variables may represent not only the lower sprint ability but also the different sprint mechanics.
Given the relationship between larger GRFs and higher sprint performance,7,11,12 larger lower limb joint torques contribute to higher sprint performance. The vertical and anteroposterior GRFs during the stance phase of sprinting were mainly generated by the lower limb extensors.13–16 The ankle plantar flexors primarily contributed to the generation of vertical and propulsive GRFs, followed by the knee and hip extensors.13–16 In addition, Nagahara and Murata (2024) 17 found that a larger plantarflexion torque was associated with a higher sprint speed, but no significant correlation was observed in knee and hip extension torques. Thus, a larger ankle plantarflexion torque may be a key determinant of a higher sprint performance, but not knee and hip extension torques. The knee extensors contributed to the generation of vertical GRF but also predominantly contributed to the generation of braking GRF.13–16 Although larger vertical GRF would be essential for sprinting, a larger braking force may be associated with a smaller ratio of force, which could limit sprint performance.5,9 Therefore, the ankle plantarflexion torque would be a key determinant of sprint performance, but knee and hip extension torques would be a limiting factor by increasing the braking force and decreasing the ratio of force.
With regard to sports specificity, whether training programs based on sprinters’ sprint mechanics can be applied to non-sprinters remains to be considered. Non-sprinters habitually engage in various sports motions, such as changes of directions and landing, including large lower limb flexion. These specificities may affect the lower limb joint kinetics during sprinting as well as joint kinematics. 10 Thus, understanding the differences in sprint mechanics between non-sprinters and sprinters may provide insights into the development of non-sprinter-specific sprint training programs. Clark et al. (2025) 10 showed larger leg extension motions from contact to toe-off in non-sprinters than in sprinters, implying that non-sprinters relied more on hip extension torque. Consequently, non-sprinters may exhibit a smaller ankle plantarflexion torque than sprinters but a larger or same knee and hip extension torque. Therefore, to test this hypothesis, the differences in lower limb joint kinetics between non-sprinters and sprinters were examined in this study during the stance phase of maximal speed sprinting.
Methods
Participants
A total of 26 non-sprinters, including 13 soccer and 13 baseball players (age, 19.9 ± 1.1 years; height, 176.2 ± 5.7 cm; body mass, 73.3 ± 7.7 kg), and 12 sprinters (age, 20.0 ± 0.9 years; height, 175.1 ± 5.3 cm; body mass, 69.2 ± 4.3 kg) participated in this study. All participants reported no pain during maximal-effort athletic motions, such as jumping and sprinting, and they were free from injury. A written informed consent was obtained from all participants prior to the experiment. This study was approved by the Human Subjects Committee of the University of Tsukuba (reference number: 020–9).
Experiments and data collection
Experiments were conducted outdoors on an athletic surface. The participants wore well-fitted clothes and their own running shoes: spiked shoes for sprinters and non-spiked shoes for non-sprinters. A total of 47 retroreflective markers were placed on the trunk and limbs, in accordance with our previous studies.18,19 Each participant completed an individual warm-up, which included dynamic stretching, jumping, running at different speeds, and maximal-effort sprinting. After the warm-up, the participants performed more than two and up to four maximal-effort sprint trials. Sprinters initiated sprints from starting blocks, whereas non-sprinters began from a self-selected standing position. Adequate recovery time was provided between trials to minimize the effects of fatigue.
A motion capture area was placed between the 35–45 m marks from the starting line for non-sprinters and the 40–50 m marks for sprinters to capture sprint motion near the maximal sprint speed. This area covered two full sprint cycles. In this study, the kinematics and kinetics of the stance leg were analyzed. Only one stance leg was analyzed because of the limited length of the force plates. Following the capture area, a deceleration area of approximately 20 m was provided. A three-dimensional motion capture system (26 cameras; VICON-MX, Vicon Motion Systems Ltd, UK), sampling at 250 Hz, was used to record the three-dimensional marker trajectories. Three ground-embedded force plates (one 9287 B and two 9281 A, Kistler, Switzerland), sampling at 1000 Hz, were used to record GRFs during the stance phase of sprinting. The marker trajectories and GRF data were synchronized and simultaneously recorded by using the motion capture system.
Data analysis
Following data analysis performed using MATLAB software (MATLAB 2023b, MathWorks Inc., USA). Three-dimensional marker trajectories were low-pass filtered using a fourth-order zero-lag Butterworth digital filter with a cut-off frequency of 18 Hz, as determined via residual analysis, 20 in accordance with our previous study. 18 Similarly, the GRF data were low-pass filtered at a cut-off frequency of 18 Hz. 21 The kinetics of the lower limb joints on the stance leg side, either right or left, were analyzed. The stance phase was defined as the period from foot strikes to toe-off. The instances of foot strike and toe-off were detected using a vertical GRF threshold of 20 N. The stance phases of the right or left leg in more than two sprint trials were selected for subsequent analysis.
In calculating kinetic and kinematic data, the whole body was modeled as a rigid-linked skeletal model comprising 15 body segments and 14 joints. The segments included the head–neck, upper trunk, pelvis, upper arm, forearm, hand, thigh, shank, and foot. The joints included the neck, lumbosacral, shoulder, elbow, wrist, hip, knee, and ankle joints. The lumbosacral and hip joint centers were determined following the methodology of Sado et al. (2024). 22 The elbow, wrist, ankle, and knee joint centers were defined as the midpoints between the lateral and medial markers attached to each segment. The shoulder and neck joint centers were defined in accordance with the method described by Reed et al. (1999). 23 Body segment inertial parameters were defined based on previous studies.24,25 The kinematics of the whole-body center of mass (CoM) was estimated based on the above-described modelling method. Sprint velocity was calculated as the average horizontal velocity of the whole-body CoM during the analysis phase.
Joint angles at the hip, knee, and ankle were calculated from the proximal and distal segment coordinate systems as Cardan angles (x–y–z sequence). Inverse dynamics problem-solving was employed to calculate joint torques at the hip, knee, and ankle during the stance phase. 20 Joint powers were calculated as the dot product of joint torque and angular velocity. These joint torques and powers were normalized by body mass. The joint angles, angular velocities, torques, and powers of the lower limb joints during the stance phase were normalized to 101 points for comparison between non-sprinters and sprinters. In addition, discrete variables of joint kinetics were extracted. The mean values of joint torque and power (negative and positive) were calculated. Furthermore, anterior–posterior, vertical, and resultant GRFs were normalized to 101 points for comparison between non-sprinters and sprinters. Spatiotemporal and GRF variables during the stance phase were also calculated as follows: contact time, braking time, propulsive time, mean values of vertical and anteroposterior (net, braking, and propulsive components) forces, vertical and anteroposterior (net, braking, and propulsive components) impulses, and the mean ratio of force. The mean ratio of force was calculated as the ratio of the anteroposterior force to the resultant force at each data point, 9 and then averaged over the entire stance phase.
Statistics
Outcome variables were defined as the mean of more than two stance phases. Descriptive data are represented as mean ± one standard deviation. Statistical significance was set at 0.05 for all tests. Parametric statistical analysis with independent comparisons in the SPM-1D code 26 (version M.0.4.9, https://spm1d.org/) was performed to compare waveform variables between non-sprinters and sprinters. These waveform variables included GRFs (anterior–posterior, vertical, and resultant components), joint angle, angular velocity, torque, and power during the stance phase of maximal speed sprinting. Moreover, an independent t-test was performed using the Statistics and Machine Learning Toolbox in MATLAB to compare discrete variables, such as spatiotemporal and kinetic variables, between non-sprinters and sprinters. Effect sizes were calculated using Cohen's d 27 and categorized as trivial (<0.2), small (0.2 to <0.6), moderate (0.6–1.2), and large (>1.2). 28
Results
The average sprint velocity was 8.53 ± 0.38 m/s (range 7.81–9.37 m/s) and 9.62 ± 0.25 m/s (range 9.26–10.14 m/s) for non-sprinters and sprinters, respectively. Non-sprinters exhibited significantly longer contact, braking, and propulsive times than sprinters, with large effect sizes (Table 1). The mean values of anteroposterior (net, braking, and propulsive), vertical, and resultant GRFs were significantly smaller in non-sprinters than in sprinters, with moderate and large effect sizes (Table 1). The impulses of these forces did not differ between the two groups (p > 0.05; Table 1). The mean ratio of force in non-sprinters was significantly smaller than that in sprinters, with a moderate effect size (Table 1). Waveform statistical analysis (Figure 1) revealed that non-sprinters produced smaller anteroposterior, vertical, and resultant GRFs throughout the stance phase than sprinters.
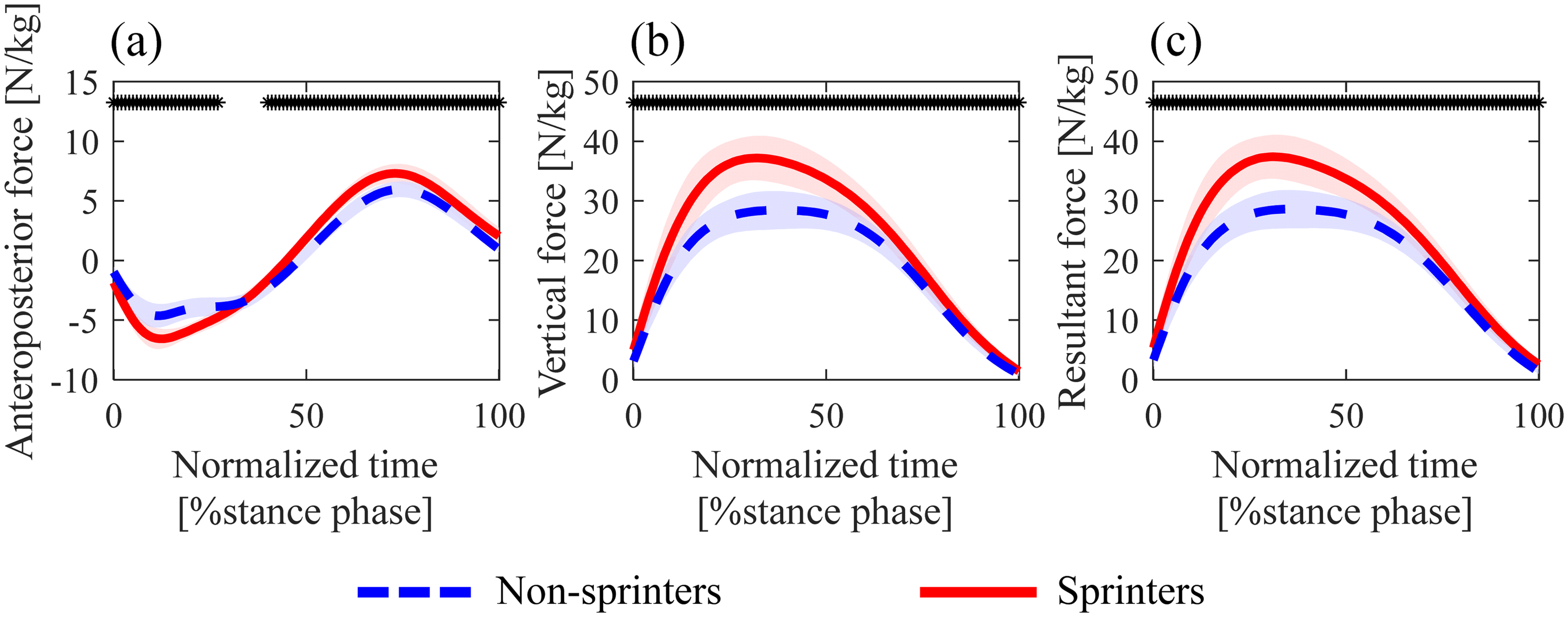
Mean of anteroposterior (a), vertical (b), and resultant (c) GRFs during the stance phase in the maximal speed phase. Each shadow area represents one standard deviation. Each GRF data (a–c) was normalized by participants’ body mass. The black asterisk (*) at the top of the graph represents the time when significant differences were found between non-sprinters and sprinters.
Differences in spatiotemporal and GRF variables during the stance phase in maximal speed phase between non-sprinters and sprinters.
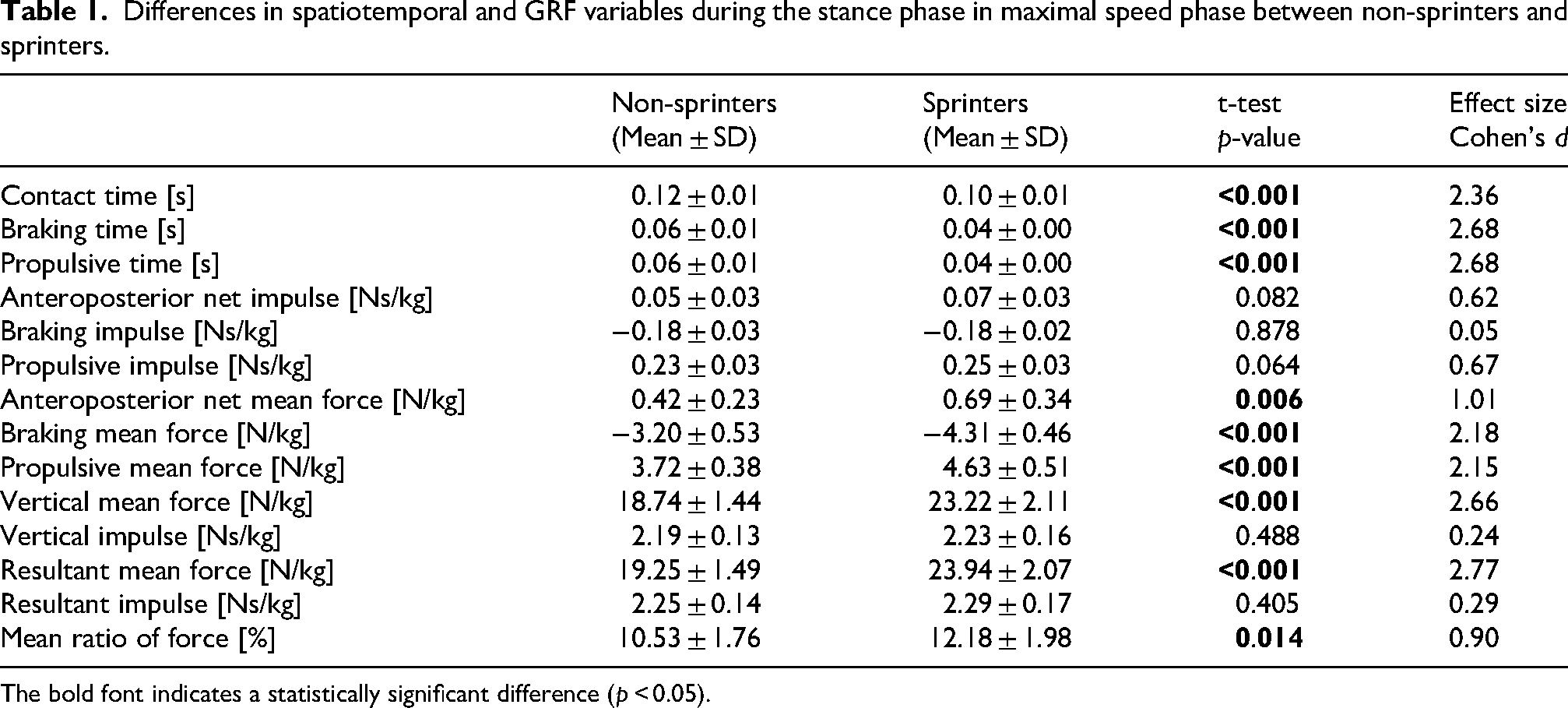
The bold font indicates a statistically significant difference (p < 0.05).
Non-sprinters were in a more flexed posture of the lower limb joints throughout the stance phase than sprinters (Figure 2(a)–(c)). Around the midpoint of the stance phase, non-sprinters exhibited larger peak values of knee flexion and ankle dorsiflexion angles than sprinters (Figure 2(b) and (c)). The knee flexion/extension joint angular velocity did not differ significantly between the two groups. In addition, a smaller hip extension joint angular velocity during the first half of the stance phase was observed in non-sprinters compared with sprinters (Figure 2(d)).
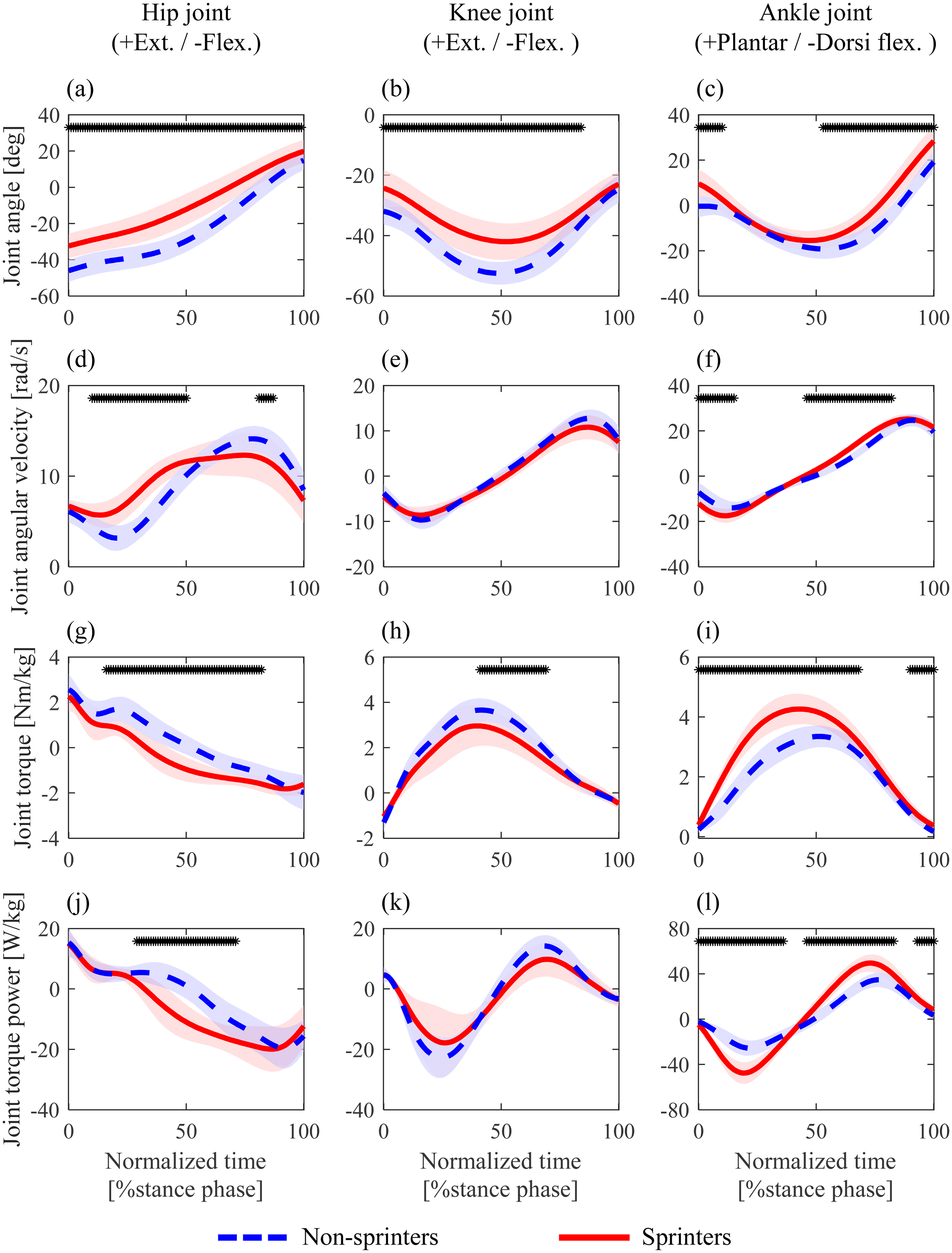
Mean of joint angles (a–c), angular velocities (d–f), torques (g–i), and torque powers (j–l) at the hip (a, d, g, j), knee (b, e, h, k), and ankle (c, f, i, l) during the stance phase in the maximal speed phase. Each shadow area represents one standard deviation. The joint torques and torque powers (g–l) were normalized by body mass. The black asterisk (*) at the top of the graph represents the time when significant differences were found between non-sprinters and sprinters.
Regarding joint kinetics (Figure 2(g)–(l)), a larger knee extension torque around the midpoint and a smaller ankle plantarflexion torque throughout the stance phase were observed in non-sprinters than in sprinters (Figure 2(h) and (i)). The negative and positive powers of the ankle joint throughout the stance phase were significantly smaller in non-sprinters than in sprinters (Figure 2(l)). Similar results were observed in the mean torques of the ankle and knee joints (Table 2). For the hip joint, a larger hip extension torque during the first half and a smaller flexion torque during the second half of the stance phase were observed in non-sprinters than in sprinters (Figure 2(g)). Furthermore, the duration of the hip extension torque was significantly longer in non-sprinters than in sprinters (Figure 2(g); non-sprinters, 51.80% ± 8.04% stance phase and 0.037 ± 0.076 s; sprinters, 34.06% ± 9.66% stance phase and 0.033 ± 0.010 s). Significant differences were observed by using an independent t-test to compare this duration (p < 0.001, d > 2.07).
Differences in mean values of lower-limb joint torque and power during the stance phase in maximal speed phase between non-sprinters and sprinters.
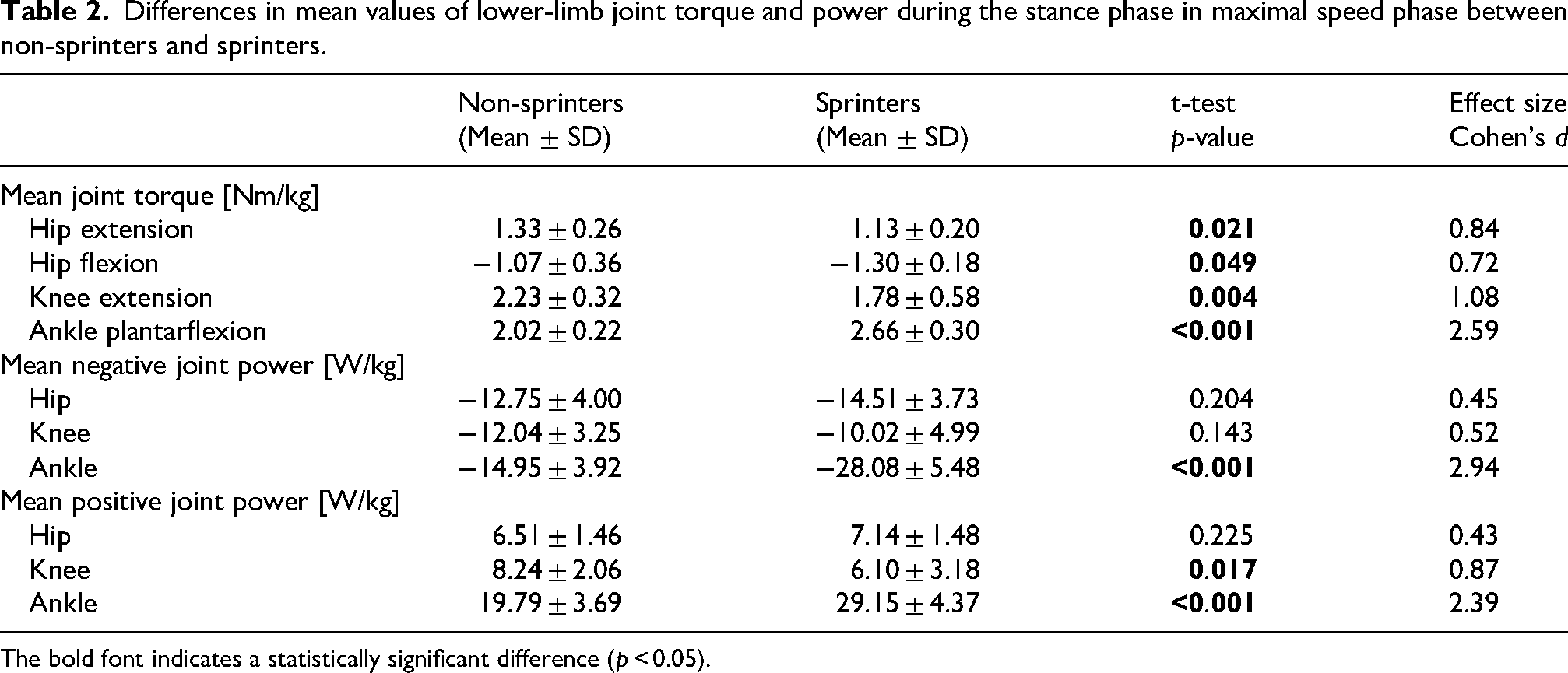
The bold font indicates a statistically significant difference (p < 0.05).
Discussion
In this study, differences in lower limb joint kinetics during the stance phase of maximal speed sprinting between non-sprinters and sprinters were identified. Non-sprinters exhibited a smaller ankle plantarflexion torque but larger hip and knee extension torques compared with sprinters, which supports our hypothesis. These findings indicate the different sprint mechanics in how GRFs are generated by the lower limb joints between non-sprinters and sprinters.
Non-sprinters exhibited smaller vertical and anteroposterior GRFs throughout the stance phase and longer contact times than sprinters, which is consistent with previous studies.4,6 The present study also showed that the differences in contact time and mean GRFs had large effect sizes (Cohen's d > 2.0). These findings indicate that non-sprinters have a lower ability to generate large GRFs within a short time than sprinters. Moreover, the vertical and anteroposterior impulses did not significantly differ between the two groups, suggesting that non-sprinters compensate for these impulses through longer contact times with smaller GRFs. Because GRFs were mainly generated by the lower limb extensors,13–16 these results imply that non-sprinters have a lower ability to produce muscle forces of lower limb extensors within a short time, which may lead to lower sprint performance. Furthermore, the braking force was smaller in the non-sprinters than in the sprinters due to their lower sprint speed. Despite the larger braking force in the sprinters, they exhibited a higher ratio of force than the non-sprinters, suggesting a more forward orientation of the GRF vector. This result indicates that the non-sprinters have an inferior force application technique compared with the sprinters, in line with a previous study. 5
This study found that the ankle plantarflexion torque and plantarflexion–dorsiflexion power were smaller in non-sprinters than in sprinters, suggesting that these smaller ankle joint kinetics may account for the smaller vertical and anteroposterior GRFs observed in non-sprinters. Previous studies have shown that the ankle plantar flexors mainly contribute to the generation of vertical and propulsive GRFs during running and sprinting.13–16 Thus, in non-sprinters, insufficient ankle plantarflexion torque would be associated with smaller GRFs. Furthermore, the differences in ankle plantarflexion torque between non-sprinters and sprinters were greater during the first half of the stance phase than during the second half. During the first half, downward displacement of the CoM and lower limb joint flexion were observed. The torques and negative power of the lower limb joints can resist excessive downward displacement of the CoM and joint flexion. Therefore, a smaller ankle joint torque and power may lead to greater flexion of the lower limb joints and subsequent extension motions, which could result in longer contact times. Consistently, in the present study, a more flexed posture and greater angular displacement from contact to toe-off were observed in non-sprinters than in sprinters. This larger angular displacement, along with a smaller joint torque, indicates lower joint stiffness. In particular, higher ankle joint stiffness might be associated with higher sprint performance.29,30 During the first half of the stance phase, higher ankle joint stiffness might reduce contact time by preventing excessive angular displacement. Taken together, insufficient ankle plantarflexion torque and power, especially during the first half of the stance phase, and lower ankle joint stiffness may be limiting factors for sprint speed. Since ankle plantar flexors act eccentrically during this phase, improving their eccentric function might enhance the sprint performance of non-sprinters.
Contrary to the smaller ankle plantarflexion torque, larger knee and hip extension torques were observed in non-sprinters than in sprinters, suggesting the different mechanisms of GRF generation. This finding implies that the knee and hip extension torques compensate for the insufficient ankle plantarflexion torque in non-sprinters. The knee and hip extension torques contribute to the generation of the vertical GRF following the ankle plantarflexion torque, although they also contribute to braking force.13–16 Thus, the predominant use of the knee and hip extension torques may increase braking forces relative to vertical force. Moreover, the present study showed that the knee extension torque was larger in non-sprinters than in sprinters during the propulsive phase. In this propulsive phase, the knee extensors would negatively contribute to producing the propulsive force, while contributing to generation of vertical force. 16 The larger knee extension torques in non-sprinters during the propulsive phase may be associated with a smaller propulsive force, which in turn may result in a smaller ratio of force. Consequently, predominantly utilizing the hip and knee extension torques to compensate for insufficient ankle plantarflexion torque would be a limiting factor for sprint speed in non-sprinters.
In addition to the larger hip extension torque, non-sprinters exhibited a longer duration of this torque than sprinters. Moreover, the hip joint showed a greater range of motion, defined as the angular displacement between contact and toe-off, in non-sprinters, which is consistent with a previous study. 10 Although non-sprinters might utilize hip extension torque to generate GRFs, the prolonged hip extension torque may limit the exertion time of hip flexion torque. The present study also found that flexion torque and negative power of the hip joint during the second half of the stance phase were smaller in non-sprinters than in sprinters. The hip flexion torque would generate the flexion angular velocity of the hip joint during the subsequent swing phase, thereby enhancing swinging leg motion. Accordingly, a smaller flexion torque could be a limiting factor for sprint performance by reducing the subsequent swing-leg speed. Given that swing time decreases with the increase of running speed, 12 a rapid swing motion is a fundamental skill for achieving a higher running speed. A previous study found that non-sprinters exhibited less forward swinging of the thigh segment than sprinters. 10 A longer duration of hip extension torque may also produce larger hip extension angles at toe-off, and then induce a delay in leg recovery during the subsequent swing phase. This sprint characteristic would be associated with the “ground-based” strategy.10,31 Moreover, from the perspective of muscular function, switching the activity from the biceps femoris to the rectus femoris was observed during the early swing phase.32,33 These findings suggest that rapid transitions in hip joint torque from extension to flexion could improve sprint performance by adapting to shorter swing time as running speed increases. Therefore, as a practical implication, sprint training that promotes quick hip flexion during the stance phase, such as mini-hurdle drills, 34 may improve sprint performance in non-sprinters through improving swing-leg motion.
The findings of this study indicate the different sprint mechanics between non-sprinters and sprinters. Consistent with previous studies,5,6,10 these differences in joint kinetics might be associated with not only sprint ability but also sports specificities. Compared with sprinters, non-sprinters predominantly utilize their hip and knee extension torques rather than ankle plantarflexion torque during the stance phase. This characteristic may result from the weakness of the ankle plantar flexors. Moreover, non-sprinters had shorter Achilles tendon moment arms than sprinters, 35 which may limit their ankle plantarflexion torque. Therefore, increasing muscle strength and improving neuromuscular function during the eccentric contraction of the ankle plantar flexors, to address this morphological disadvantage, could improve sprint performance in non-sprinters.
Furthermore, these different sprint mechanics between non-sprinters and sprinters may be attributed to the differences in sport-specific movement characteristics, especially their habitually practiced athletic motions. Non-sprinters frequently engage in movements involving greater lower limb joint flexion, such as changes of direction and landing. Maniar et al. (2019) 36 reported that the knee extensors primarily contributed to the vertical GRF during cutting maneuvers, compared with high-speed running.13–16 These differences in lower limb joint functions between sprinting and cutting maneuvers imply that non-sprinters may generate a greater knee extension torque during sprinting to prepare for subsequent direction changes, which are commonly required in their sports. For non-sprinters, understanding their unique strategy of GRF generation through lower-limb joint torques would be useful for the development of non-sprinter-specific training programs. As a practical implication, the findings of this study suggest that ankle plantarflexor function and a rapid transition of hip joint torque from extension to flexion may be key modifiable factors to improve sprint performance in the maximal speed phase. Nevertheless, since non-sprinters frequently require rapid responses to various situations during games, these suggested modifications may not be directly applicable to short-sprint situations involving frequent changes of direction. Therefore, carefully considering the optimal sprint mechanics for achieving a higher speed or faster changes of direction is necessary in non-sprinters. Further research is warranted to address this issue.
This study has some limitations. First, the longitudinal changes in lower limb joint kinetics were not investigated in both non-sprinters and sprinters. Although our findings highlight the sports-specific characteristics of sprint mechanics, careful planning of sprint training for non-sprinters is essential to achieve higher sprint performance. Second, the differences in sprint motion between spiked and non-spiked shoes or between starting methods with and without starting blocks were not confirmed. In addition, sprinters generally train on track surfaces, whereas non-sprinters practice on artificial turf and soil surfaces. These differences in running shoes, starting methods, and familiarity with track surfaces may have affected sprint motion, and the results of this study may have been influenced by these differences. Third, the present study did not examine the differences in sprint motion and GRF variables between non-sprinters and sprinters during the early acceleration phase. For non-sprinters, frequently engaging in shorter sprint running during games and achieving a higher sprint speed as quickly as possible is essential. Future studies are required to examine the differences in sprint motion and GRF variables during the acceleration phase of sprinting.
Conclusions
This study aimed to examine the differences in lower limb joint kinetics during the stance phase of maximal speed sprinting between non-sprinters (soccer and baseball players) and sprinters. Compared with sprinters, non-sprinters exhibited a smaller ankle plantarflexion torque but larger knee and hip extension torques. These results indicate that the knee and hip extension torques in non-sprinters compensate for the insufficient ankle plantarflexion torque in generating vertical and anteroposterior GRFs. These findings imply the differences in lower-limb joint kinetics, which contribute to the generation of GRFs between non-sprinters and sprinters. Understanding these kinetic differences may help develop training programs tailored to non-sprinters.
Footnotes
Acknowledgments
We would like to thank Sports Biomechanics Laboratory members at University of Tsukuba for their help with collecting data.
Ethical considerations
This study was approved by the Human Subjects Committee of the University of Tsukuba (reference number: 020–9).
Consent to participate
A written informed consent was obtained from all participants prior to the experiment.
Consent for publication
Not applicable.
Authorship contribution statement
Terumitsu Miyazaki: Writing – original draft, Writing – review & editing, Validation, Visualization, Software, Project administration, Methodology, Formal analysis, Investigation, Data curation, Funding acquisition, Conceptualization, Supervision. Hirotomo Kubota: Writing – review & editing, Data curation, Investigation, Formal analysis, Methodology. Norihisa Fujii: Writing – review & editing, Methodology, Conceptualization, Funding acquisition.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number [24K20565 for T.M. and 20H04067 for N.F.] and MIZUNO SPORTS PROMOTION FOUNDATION (for T.M.).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
Data will be made available on request.