
Abstract
Ten cadaveric forearms were tested using a wrist testing apparatus specifically designed to investigate the mechanisms of muscle stabilization of the wrist. The specimens were set in a jig allowing the distal row to migrate proximally and rotate around the axis of pronosupination. The extensor carpi ulnaris (ECU) was loaded with specific weights. Reactive rotations of the scaphoid, triquetrum, and capitate were measured by an electromagnetic motion tracking device. Loading the ECU caused pronation of both proximal and distal rows. After sectioning its sheath, the overall direction of the movement remained unchanged, but there was a 40% and 50% decrease of the pronation power over the distal and proximal carpal row, respectively. In addition to stabilizing the distal radiolunar joint, the ECU is an important structure that contributes to the dynamic stability of the wrist. Furthermore, its sheath plays a crucial role in maintaining the effect of the ECU muscle on the carpus.
Introduction
Although the action of the extensor carpi ulnaris (ECU) on the global movement of the wrist and its effect on distal radiolunar joint (DRUJ) stability are well established, very little has been written about the specific roles of the motor muscles of the wrist, and specifically of the ECU and its sheath. Spinner and Kaplan (1970) noted the importance of the location of the ECU in its fibro-osseous canal on the dorsum of the ulna as an additional important stabilizing factor of the distal radiolunar joint. Since then, all articles have focused on the effect of the ECU on the dynamic stability of the DRUJ. The purpose of this study was to investigate the effects of simulating contraction of the ECU muscle in the three-dimensional (3D) alignment of specific carpal bones in order to better understand the role of the ECU and its sheath on dynamic carpal stability.
Methods
Ten fresh cadaver arms without pre-existing wrist pathology were used in these experiments. The use of human cadavers was in accordance with the regulations of our institutions. The average age of the specimens was 77 years (range 55–88). There were six male and four female wrists, all of which were unpaired. There were seven left wrists and three right wrists. Nine of the specimens had a type I lunate and one a type II lunate. Specimens were prepared by allowing them to thaw at room temperature and excising skin and superficial fascia around the wrist joint. Five muscles with tendons crossing the wrist joint (extensor carpi radialis longus [ECRL], ECU, flexor carpi ulnaris [FCU], flexor carpi radialis [FCR], and abductor pollicis longus [APL]) were transected at the middle-third of the forearm. Both the extensor and flexor retinacula were preserved. The proximal ends of the transected tendons were connected by heavy nylon strings to a system of pulleys, which allowed the tendons to be loaded with variable amounts of weights. The fingers were disarticulated at the metacarpophalangeal joints.
Arms were set vertically in a specifically designed jig and secured by two Steinmann pins drilled across the two forearm bones in neutral axial rotation (Figure 1). To facilitate setting sensors in the carpal bones, the extensor carpi radialis brevis tendon was removed and a capsular triangle between the dorsal radiocarpal and dorsal intercarpal ligaments was resected. Care was taken to avoid injuring any intrinsic or extrinsic ligaments. The position of the wrist was controlled with a pin inserted into the medullary canal of the middle finger metacarpal. The pin was held in a slot within an overhanging semicircular frame that allowed proximal-distal gliding of the frame along the pin. This ensured that the constrained rotation of the wrist occurred around an axis that roughly coincided with the centre of the capitate. Furthermore, the frame was connected to a horizontal rotating mechanism that controlled the plane of wrist rotation. In these experiments the frame was set in the sagittal plane and the pin held in a neutral vertical position throughout the test.
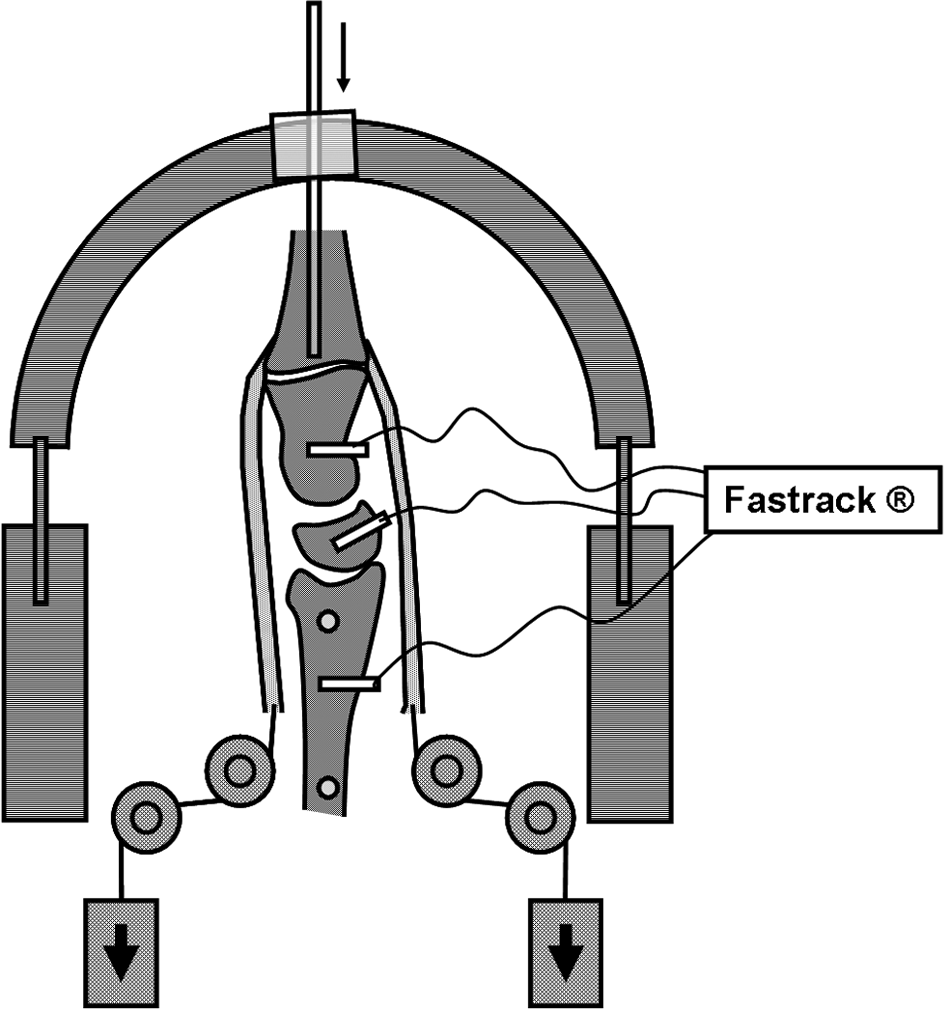
Arm set in the specially designed arm holder with sensors firmly connected to the dorsal aspect of the scaphoid, triquetrum, capitate, and radius by nylon rods. The position of the wrist was controlled with a pin inserted into the medullary canal of the middle finger metacarpal.
The 3D changes in the alignment of carpal bones were monitored using a FASTRAK motion tracking device (Polhemus Inc., Colchester, Vermont, USA). The system creates an electromagnetic field in which the 3D positions and orientations of specific moving markers relative to the fixed coordinate system can be calculated in six degrees of freedom. Four sensors operating at an update rate of 30 Hz and a short-range transmitter specially designed for small tracking areas spanning 2.54 to 60.9 cm were used. The static accuracy of the spatial tracking system was 0.8 mm for marker positioning and 0.15° for spatial orientation of the marker. Sensors were firmly connected to the dorsal aspect of the scaphoid, triquetrum, and capitate by nylon rods. The amount of load applied to each tendon was based on previous investigations that suggested using loads proportional to both the physiological cross-sectional area and relative electromyographic activity of each muscle when grasping an object with the wrist in neutral position (An et al., 1991; Brand, 1981; Kobayashi et al., 1997). To simulate muscle tone, all tendons were continuously loaded with a 1.5 N weight. To simulate muscle contraction, a 13.7 N weight was applied to the ECU.
The coordinate system of the transmitter and four sensors was constructed using the longitudinal axis of the radius as the Z-axis of reference. The Y-axis was set along the frontal plane, perpendicular to the Z-axis, and the X-axis was perpendicular to the Y- and Z-axis, along the sagittal plane. Rotation around the Z axis (yaw angle) indicated pronation-supination; rotation around the Y-axis (pitch angle) indicated flexion-extension; and rotation around the X axis (roll angle) indicated radial-ulnar deviation.
Two conditions of isolated loading of the ECU were analyzed before and after sectioning the ECU sheath, while the rest of the tendons had only the weight representing muscle tone. To assess the effects of tendon loading, the 3D positions of the scaphoid, triquetrum, and capitate were monitored before and after loading, and the average of five consecutive measurements was used to calculate the rotations sustained by the sensors. Because the goal of this study was to reproduce the conditions of an isometric type of loading — that is, with both flexion-extension and radioulnar inclination being constrained by the pin in the jig — this paper will mostly emphasize the intracarpal changes that occurred in the transverse plane (pronosupination) as a result of muscle loading.
Differences in spatial positioning of the scaphoid, triquetrum, and capitate sensors between the two loading conditions were statistically analyzed using the Wilcoxon test. p<0.05 was considered statistically significant.
Results
ECU loading caused changes in the spatial orientation of the scaphoid, triquetrum, and capitate bones. The isolated ECU loading provoked substantial pronation to both the distal row as well as the scaphoid and triquetrum. The scaphoid and capitate pronated in all cases: the scaphoid by an average of 1.48° and the capitate by 5.15° (Table 1). The triquetrum pronated in 7 out of 10 cases. After sectioning the ECU sheath, the overall direction of the movement remained unchanged, but there was a 40% and 50% decrease in pronation of the proximal and distal carpal rows, respectively. The scaphoid pronated by an average of 0.77° and the capitate by 3.01° (Table 1). The triquetrum pronated in most cases (9 out of 10). Differences in the pronosupination movements of the scaphoid and capitate before and after sectioning the sheath were statistically significant (p<0.01).
Effect of loading the isolated ECU before and after sectioning its sheath
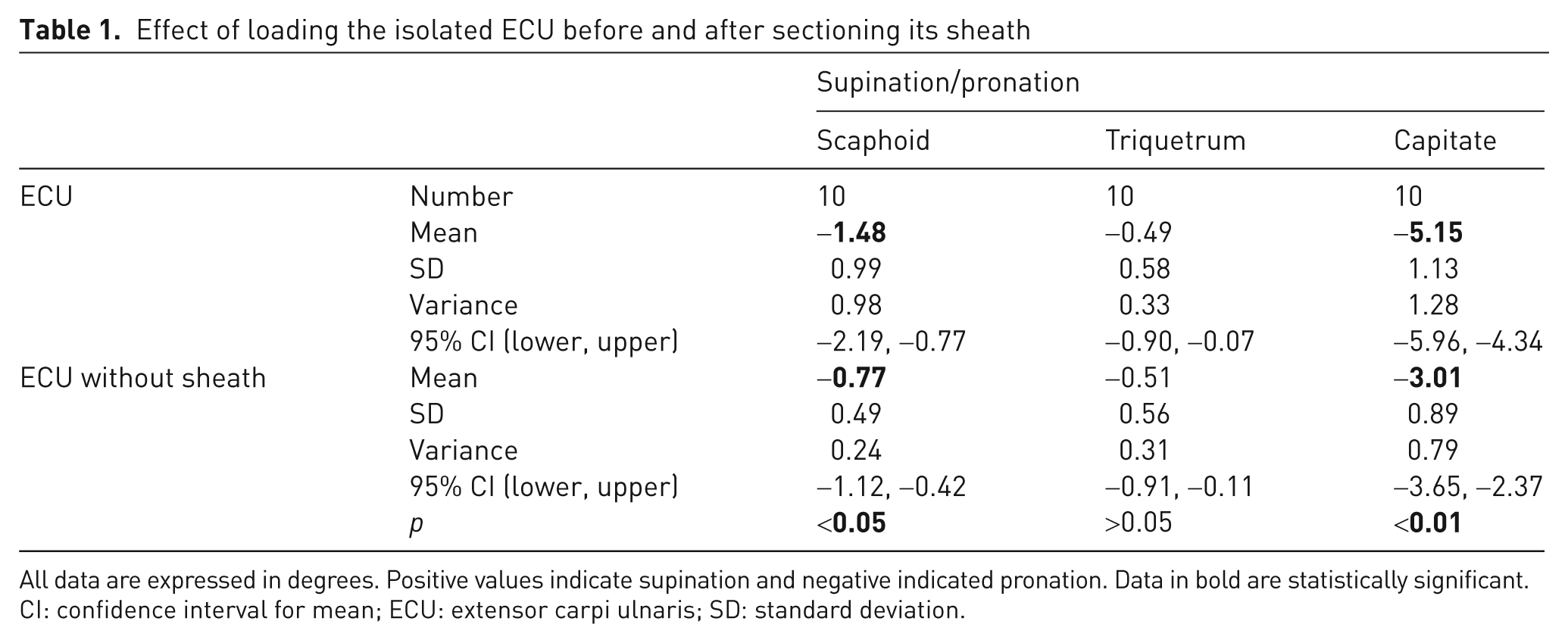
All data are expressed in degrees. Positive values indicate supination and negative indicated pronation. Data in bold are statistically significant.
CI: confidence interval for mean; ECU: extensor carpi ulnaris; SD: standard deviation.
Discussion
Previously published papers about the role of the ECU in carpal stability almost exclusively refer to the dynamic effect of muscle on the distal radioulnar joint (Adams and Berger, 2002; Berger, 2001; Kauer, 1991; Kihara et al., 1995; Linscheid, 1992).
The anatomy of the sixth dorsal compartment is unique and complex. The ECU has an angular path as it passes over the head of the ulna to its insertion at the base of the small finger metacarpal, which further increases in ulnar deviation. The ECU contributes to the stability of the DRUJ, and its anatomical relationship with the extensor retinaculum, which has no insertion to the ulna but surrounds it to attach to the pisiform and triquetrum, allows unrestricted rotation in pronation and supination (Spinner and Kaplan, 1970).
Several studies highlight the importance of the ECU being located in its sheath, because there could otherwise be symptomatic subluxations of the ECU, and the stability of the DRUJ could also be affected (Adams and Berger, 2002; Berger, 2001; Kauer, 1991; Spinner and Kaplan, 1970).
Little has been published about the role of the different wrist motor tendons on dynamic carpal stability. Ruby (1992) suggested that the musculotendinous units in their fibro-osseous canals had a dynamic effect on wrist stability, but he did not define their specific roles. Kauer (1980) recorded the electromyographic activity of different forearm muscles during wrist motion and observed minimal activity of the extensor pollicis brevis, APL, and ECU during unresisted wrist flexion. That led him to conclude that their function could be described as an adjustable collateral system, but no further mention of their role as wrist stabilizers was made.
In a previous publication, we presented the actions of five forearm muscles on the carpal bones in an experimental research study in cadavers (Salva-Coll et al., 2011). We proposed that forearm muscles could be divided into carpal pronators and supinators when axially loaded and suggested that dynamic wrist stability is related to the balance of wrist pronators (ECU and FCR) and supinators (FCU, ECRL, and APL). In this study we analyzed the specific effect of the ECU and the role of its sheath on rotation of the carpal bones when isolated axial loading is applied. As described in the results section, isolated ECU loading provoked substantial pronation to both the distal row, and to the scaphoid and triquetrum. In fact, among the tendons studied, it is the strongest pronator of the distal and proximal carpal rows.
The most likely explanation of the ECU action is that it is due to its position on the dorsal aspect of the ulna, having its own sheath, and divergence in the coronal and sagittal planes to its insertion at the anteromedial corner of the base of the small finger metacarpal (Figure 2a), using the distal ulna as a pulley to increase its mechanical advantage. The greatest amount of rotation after loading the ECU was observed in the distal carpal row. It has long been recognized that, if there is no muscle influence, the axially loaded scaphoid tends to rotate into flexion and pronation owing to its oblique alignment with respect to the longitudinal axis of the hand and its unique relationship with the obliquely oriented radioscaphocapitate ligament (Garcia-Elias, 1997; Garcia-Elias and Geissler, 2005; Kauer, 1980). If we take into account the effect observed in the carpus after loading the ECU, this muscle would enhance the natural tendency of the scaphoid to pronate, and thus, it could be potentially harmful for the scapholunate ligament, a phenomenon made evident by direct observation in all our specimens.
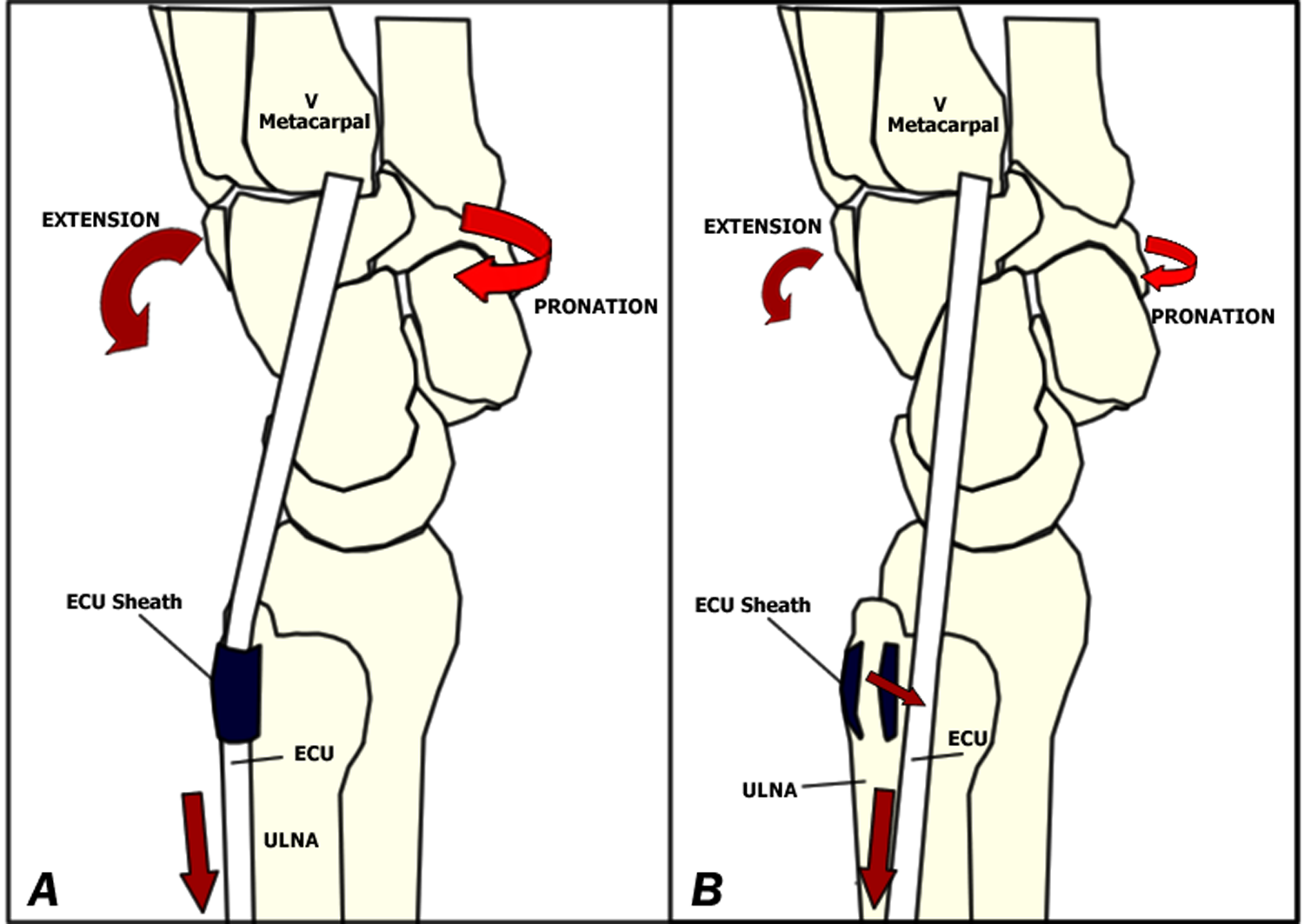
(A) Schematic drawing of the wrist and location of the ECU tendon, inserting at the base of the small finger metacarpal. (B) Medial subluxation of the ECU after sectioning the ECU sheath, causing loss of its mechanical advantage over the distal ulna.
After sectioning the ECU sheath, pronation of the scaphoid and capitate decreased significantly by up to 40% and 50%, respectively (Table 1). This is explained by the fact that the ECU displaces medially and loses its mechanical advantage (Figure 2b). This fact may be relevant in cases of partially incompetent ligaments, especially those located in the ulnar side of the wrist, where the carpus could be dynamically destabilized, making a previously asymptomatic ligament lesion become evident.
As a result of this study, we can conclude that the ECU tendon and its sheath are not only important in DRUJ stability, but also in the dynamic stabilization of the wrist, through its pronation effect on the distal carpal row. Hence, the importance of preserving and/or reconstructing the ECU sheath in cases of tendon subluxation.
Footnotes
Conflict of interests
None declared.
Funding
This study was supported by grants from the Spanish Society of Orthopaedic Surgeons (Sociedad Española de Cirugía Ortopédica y Traumatología [SECOT]) and the Catalan Society of Orthopaedic Surgeons (Societat Catalana de Cirurgia Ortopèdica i Traumatologia [SCCOT]).