
Abstract
Preaxial polydactyly is a congenital hand anomaly predominantly of sporadic occurrence, which is frequently associated with abnormalities of the Sonic hedgehog signalling pathway. In experimentally induced preaxial polydactyly, radial aplasia is also frequently observed. To determine if there is a correlation between preaxial polydactyly and radial aplasia, we induced ectopic Sonic hedgehog signalling during chicken limb development with application of a smoothened-agonist (SAG) or retinoic acid. Application of SAG caused malformations in 71% limbs including preaxial polydactyly (62%) and forearm abnormalities (43%). Retinoic acid application induced malformations in 56% of limb including preaxial polydactyly (45%) and forearm abnormalities (50%). Radial dysplasia and ulnar dimelia were observed in both experimental conditions. We demonstrate that ectopic Sonic hedgehog signalling may cause both preaxial polydactyly and predictable forearm anomalies and that these conditions could potentially be classified as one embryological group. We propose a unifying model based on known models of ectopic Sonic hedgehog signalling.
Keywords
Introduction
Preaxial polydactyly (PPD) is a common congenital hand anomaly predominantly of sporadic occurrence. Wassel (1969) proposed a radiological classification for PPD conditions, which has since been modified numerous times (Buck-Gramcko, 1998; Wood, 1978), most recently by Zuidam et al. (2008). These continuing efforts to ‘classify’ complex PPD conditions may reflect a lack of understanding into the underlying disease process.
Our current understanding remains that PPD is frequently caused by mutations that result in either the ectopic expression or activation of the Sonic hedgehog (SHH) signalling pathway (Anderson et al., 2012); a pathway that acts during embryonic limb development (Riddle et al., 1993) to specify both digit number and anterior-posterior (radial-ulnar) digit identity (Tickle and Towers, 2017). Examples of mutations include alterations in the long-range limb specific enhancer of SHH (the zone of polarizing activity regulatory sequence or ZRS) where altered anatomies are confined to the limb (Anderson et al., 2012), more systemic features such as Greig syndrome (OMIM #175700) or ciliopathic conditions such as Short rib polydactyly syndrome (Huber and Cormier-Daire, 2012; OMIM #616546). SHH signalling acts to induce formation of GLI-Activator proteins over GLI-Repressor proteins and in many syndromic conditions associated with PPD, activity of the GLI transcription factors downstream of SHH signalling are altered (Davey et al., 2006; Tickle and Towers, 2017; Vortkamp et al., 1991). Experimental manipulations of SHH signalling in animal models have shown that SHH acts in a time and dose dependent manner (Tickle, 1981). Thus, it is predicted that PPD severity in humans could be due both to the strength and duration of ectopic SHH expression in the developing limb bud. The developmental stage at which SHH is ectopically expressed also influences the severity of PPD; ectopic SHH expression in early limb buds creates more severe defects than expression in more mature and patterned limb buds (Dunn et al., 2011; Yang et al., 1997).
Other than the autopod (hand), the ulna is also dependent on the SHH pathway for normal development. The ulna is partially derived from SHH expressing cells (Harfe et al., 2004) and in mice and chickens where SHH expression is depleted, the ulna is lost while the radius is maintained (Chiang et al., 2001; Pagan et al., 1996; Ros et al., 2003). Precisely timed genetic disruption of SHH signalling demonstrated that the ulna requires early/high levels of SHH signalling, equivalent to that of the most posterior digit, in order to develop (Zhu et al., 2008). In addition, strong prolonged ectopic anterior SHH signalling can cause ulnar dimelia (duplication) (Duprez et al., 1999; Yang et al., 1997). More commonly, however, radial aplasia without dimelia is observed in conjunction with induced PPD in experimental models, either through anterior application of SHH protein or retinoic acid (SHH agonist) (Eichele et al., 1985; Tickle et al., 1975) or through early deletion of GLI3 (Bowers et al., 2012). So far from these experiments, it would suggest that a spectrum of effects could be elicited by ectopic SHH signalling, resulting in PPD and radial aplasia/ulnar dimelia.
We suggest a possible unifying hypothesis based on our understanding of the criteria of the limb morphogen, including developmental stage of action, time and strength of ectopic SHH signal, to explain the various degrees of PPD, radial aplasia and ulnar dimelia.
Methods
Source and incubation of chicken eggs
ISA Brown chicken lines were maintained at The Roslin Institute under UK Home Office licence. Fertilized chicken eggs were incubated at 38℃. Stage of embryonic development was determined by Hamburger and Hamilton (HH) (1951).
Smoothened agonist (SAG) application and mechanism of action
One microgram of smoothened agonist (SAG, Calbiochem, Darmstadt, Germany) was diluted in water and applied in 5 uL applications so that embryos received 1 µg, 2.5 µg or 5 µg doses between stages 15–21 HH (see Supplementary Data 3 for optimization of SAG dose). SAG was injected directly onto the embryo within the amniotic membrane. For information on the action of smoothened see Supplemental Data 1 and Supplemental Figure 1.
Radial aplasia is observed within the spectrum of ectopic SHH signalling induced limb malformations. (B–Q) Examples of limbs in which ectopic SHH signalling had been induced with through RA. (B–J) Application of SAG (K–M) or application of SHH protein (P,Q). Limbs induced by RA were categorized by observation of the polydactylous phenotype, i.e. ‘Mild PPD’, ‘Severe PPD’, Severe PPD (Mirror Hand-like), Dimelia. In all categories, limbs with radial aplasia/dysplasia (F–I, M) or limbs with a transformation from radius-ulna were observed (partial distal ulna) (M), except in Dimelia (E). (J) Limb with a recording of location of RA bead and whether it caused limb malformation (red). Control limbs (A,N,O). H: humerus; r: radius; u: ulna; 1: digit 1; 2: digit 2; 3: digit 3; o: olecranon; ra/it: radialus/intermedium; it: intermedium; pi: pisiform; slc: semilunate carpal; b: blip, *: missing bony element; PPD: preaxial polydactyly; SAG: smoothened agonist; SHH: Sonic hedgehog; RA: retinoic acid. Changes in anatomy are markers in red annotation.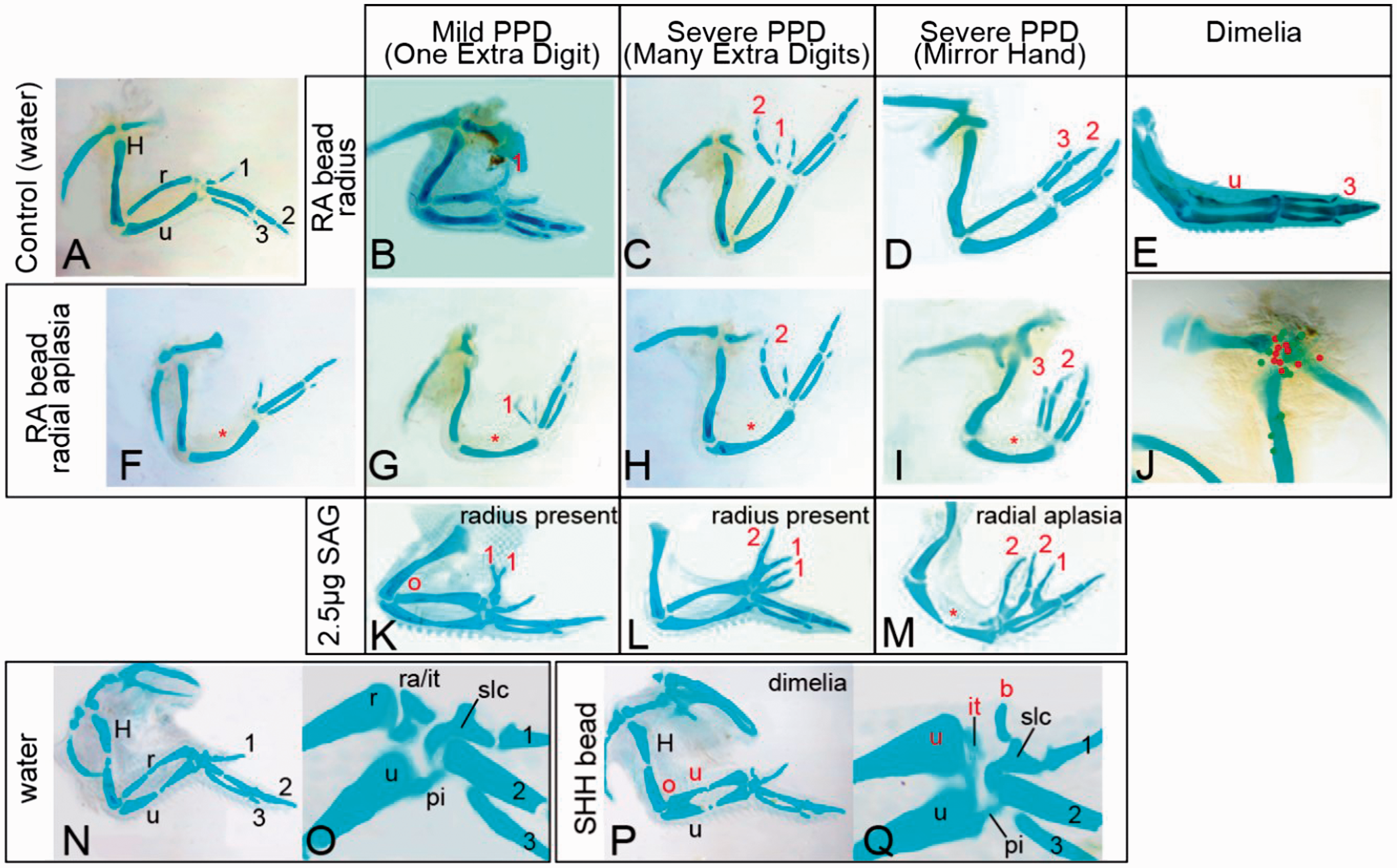
Retinoic acid (RA) application
AG1-X2 ion exchange resin beads were loaded with all-trans-retinoic acid (RA) as per Johnson et al. (2014). RA beads were implanted into a slit created under the base of the anterior distal apcial ectodermal ridge of the limb bud at stages 17–21 HH. This application of RA in this location is known to induce SHH in a precise and repeatable way in the limb mesenchyme (Johnson et al., 2014; Riddle et al., 1993).
SHH protein application
Affi-Gel Blue Beads were soaked in SHH protein (Recombinant N-Terminus Mouse SHH protein; 461-SHH RandD Systems) and applied as per Tiecke and Tickle (2007).
Skeletal staining and anatomy
Embryos were incubated at 38℃, fixed at E10 (RA and SAG manipulations) or E12 (SHH bead manipulations) in 5% trichloroacetic acid, stained overnight in 0.1% Alcian green in 70% acid alcohol and dehydrated in graded steps to 100% ethanol and cleared in methyl salicylate. Anatomy is described as per Towers et al. (2011) for digits and Botelho et al. (2014) for carpals. For further details of bird limb anatomy see online Supplemental Data 2.
Results
In preliminary experiments using SAG applied to the developing limb, we observed 4/76 independent incidences of loss or reduction of the radius, one of which was associated with very mild preaxial polydactyly (see Supplemental Data 3). As a loss of the radius in association with PPD has also been previously demonstrated, although not well described, in classical experiments in which PPD has been induced through application of posterior limb mesenchyme expressing SHH, SHH expressing fibroblasts or RA to the anterior wing bud (Eichele et al., 1985; Tickle, 1981; Yang et al., 1997), we therefore set out to examine if a loss of the radius could be directly associated with induction of PPD upon abnormal activation of SHH signalling. We undertook two approaches; application of 2.5 ug SAG to stage 16 HH to induce widespread SHH-related signalling (Group 1) or application of RA to the anterior limb bud at stages 17–21 HH to induce localized ectopic SHH signalling (Group 2).
Group 1
The number of limbs with manipulations and incidence.
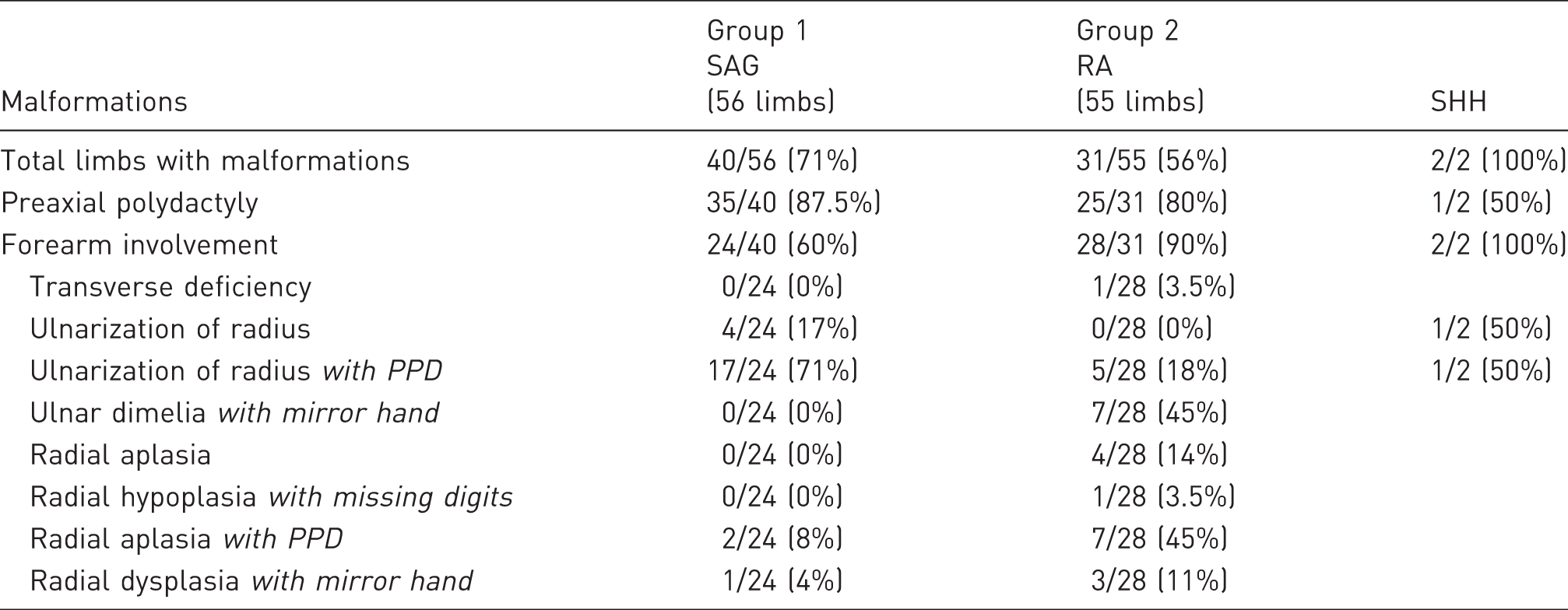
SAG: smoothened agonist; RA: retinoic acid; SHH: Sonic hedgehog; PPD: preaxial polydactyly.
Group 2
In Group 2, following RA application to the anterior limb bud, the majority of limbs again exhibited limb malformations (Figure 1(B–I); Table 1). The incidence of PPD in the autopod was similar to the SAG-treated embryos, but there was a much higher percentage of forearm involvement including radial aplasia or hypoplasia with or without associated PPD (Figure 1(F,G,H,I)). In addition, there were seven cases of true ulnar dimelia with the anterior forearm bone displaying both an olecranon proximally and a loss of the chicken radial carpal bone (radialus) distally (Figure 1(E), Supplemental Data 2). Thus, utilizing two methods for activation of SHH signalling in the limb, we have shown consistently that the zeugopod, as well as the autopod, is affected if SHH signalling is ectopically activated.
Stage and location dependence of RA effect on polydactyly
Both the stage of embryonic development and the precise location of RA beads can lead to variation in the extent of PPD that is induced (Eichele et al., 1985). To determine if the embryonic stage of RA application and location of RA also influenced the outcomes of forelimb malformations, we re-analysed our data both using subsets of samples in which the embryonic stage of manipulations were precisely recorded (n = 31) and in which we could locate and map the final location of the bead within the E10 limb as an indication of initial bead placement (n = 23). While limb truncations (2/30) including transverse deficiency and digit truncations were only observed at the earliest time point (stage 17 HH), PPD of both ‘mild’ forms (one extra digit) and severe forms (mirror hand) were observed at both early and latter stages: 19 HH (two mild and one severe) and 20 HH (three mild and two severe), while stage 21 HH exhibited only severe PPD (2/2 atypical mirror hand). Radial deficiency, however, was also observed at all stages: 17 HH, 19 HH, 20 HH and 21 HH (n = 7) associated with no PPD (2/7), mild PPD (2/7) and severe PPD (3/7). The location of bead placement also played an important role in alteration of limb anatomy and beads, those. Beads centred on the shoulder joint were more likely to result in a change in limb anatomy (red locations gave rise to limb malformations) (Figure 1(J)).
SHH protein manipulation of limb bud
Finally, to demonstrate that manipulation of the developing limb bud with SAG or RA is comparable with manipulation of SHH signalling, we directly manipulated anterior developing limb buds at stage 20 HH with recombinant SHH protein. Surviving embryos showed induction of mild PPD (n = 2) (Figure 1(P,Q)) and associated alterations to the radius, including distal thickening and shortening of the radius (n = 1). Transformation to an ulna (n = 1) included loss of the anterior radialus (compare Figure 1(O) with 1(Q)). The radial-ulna transformation was akin to the morphology observed in SAG and RA application. With SHH protein application we did not observe a loss of the radius, although this has been previously reported (Yang et al., 1997).
Discussion
PPD, radial aplasia and ulnar dimelia are seen clinically as separate entities, not only because of differences in location of aberrant anatomy, but also their different treatment regimes. Our results have suggested that these conditions could be points on a continuum of aberrations caused by abnormal SHH signalling pathway induction in the embryonic arm, similar to a hypothesis first proposed by Al-Qattan (2013).
In human, mouse and chicken limbs, digit identity is thought to be determined by the SHH pathway. According to the Growth/Specification Model (Towers et al., 2008) and based on the French Flag hypothesis (Wolpert, 1968), digit number and identity has been shown to be dependent on the strength and duration of normal SHH signalling. These hypotheses also predict that a spectrum of PPD defects would be induced by differences in strength and length of ectopic SHH expression. As SHH signalling acts also during zeugopod development, we hypothesize that ectopic SHH expression may also lead to forearm pathologies. Despite the numerous studies focusing on digits, relatively little attention has been on zeugopod formation and abnormal phenotypes caused by aberrations in the SHH signalling pathway.
In our stage-compared experiments, interpretation of results is complex but some causes of variation may be hypothesized. Early limb buds exhibited either severe truncation defects or mild PPD. The truncations are similar to X-radiation-induced cell death in the chicken limb with both anterior and posterior axes equally effected (Galloway et al., 2009). This suggests that young limb buds (stage 19 HH) are either more vulnerable to cell death caused by RA application or that the distal limb elements have been re-specified (Roselló-Díez et al., 2011) as RA specifies proximal patterning. Early application did not induce strong PPD; possibly as the stage 19 HH anterior limb bud may not be competent to express strong SHH, that is, the ‘priming phase’ may be longer (Eichele et al., 1985). On the other hand, application of RA to stage 21 HH (latter stage) induced PPD in a manner that was comparable with a mirror-hand (Eichele et al., 1985), a phenotype considered inducible only by the strongest ectopic SHH signal (Yang et al., 1997). At stage 20 HH, induced PPD phenotypes were of an intermediate type as compared with those induced between 19 HH and 21 HH, demonstrating that there is a continuum based on ectopic SHH expression.
While the severity of PPD could be correlated over time (early, intermediate and late), a reduction of the radius was seen in all stages. This suggests that the time during which the radius is susceptible to SHH signalling is both earlier and more prolonged than the digits. This is in agreement with the Summerbell hypothesis that the zeugopod is patterned before the autopod (López et al., 1995; Summerbell, 1974) or that the chondrogenic precursors of the radius are vulnerable to changes in signalling before digit precursors (Galloway et al., 2009). Our results are consistent with mouse studies in which an early loss of GLI-Repressor activity causes tibial loss in conjunction with PPD, but later timed deletions only cause PPD (Bowers et al., 2012). In addition, our results also suggest that radial aplasia/hypoplasia can be induced by a wide range of strength in SHH signalling, although ulnar dimelia was only induced in limbs manipulated directly with SHH/RA and not with SAG (weaker agonist). The RA-induced PPDs were the most extreme, where anterior digits of the ‘index type’ were completely lost. As RA is known to induce a large area of anterior ectopic SHH signalling, this suggests that dimelia only occurs when there are high ectopic levels of SHH signalling. Indeed, application of SHH only caused a brief but strong activation of the SHH pathway. With this brief activation, we were able to produce only a forelimb phenotype (dimelia), confirming that the radius is susceptible to SHH signalling at an earlier time point. Finally, dimelia was only induced by RA/SHH in manipulations of older limb buds at stage 20 HH+, showing that the conversion of identity is a later occurrence than specification of an anterior zeugopod element.
We therefore suggest that there are two time-dependent phases in the induction of the zeugopod pattern (Figure 2). Phase 1 is dependent on low concentrations of SHH and during this time period, the zeugopod tissues become ‘primed’ to form two zeugopod elements with no set identity. Additional anterior SHH signalling during this phase ablates first the presumptive anterior zeugopod element and at higher levels, the posterior zeugopod element, resulting in a limb truncation. This illustrates that even when they are first established, the presumptive radius and ulna have different developmental potentials and sensitivity to SHH signalling. If not exposed to additional SHH signalling, both zeugopod elements would differentiate into a ‘default’ radius, as is observed in the replicated anterior zeugopod mouse in which lowered SHH signalling results in two radii (Krebs et al., 2003). During Phase 2, additional SHH signalling does not ablate the radius but instead acts to transforms it into an ulna. We suggest this is a prolonged developmental phase, illustrated by the partial ‘ulna transformations’ in which the radius becomes shorter and proximally retains a radius articulation with the elbow but distally becomes ulna-like. Both phases can overlap with the period of digit patterning, although our evidence suggests Phase 1 occurs slightly earlier than digit patterning.
Hypothesized correlation between longitudinal deficiencies and SHH signalling. (a) We propose, based on our experimental evidence and that of others, that the strength of ectopic SHH signalling causes a spectrum of forelimb malformations. Partial radial to ulna transformations occur at lower levels of ectopic SHH signalling and dimelia as a result of high levels of ectopic SHH signalling. (b) We further propose that limb malformations induced by ectopic SHH expression are determined both by the strength of SHH signalling and the time that they are expressed and that SHH may be an underlying cause of a variety of limb malformations not currently classed as related. Early ectopic SHH signalling can induce first transverse deficiency, then radial deficiency, then radius-ulna transformations and finally polydactyly. Depending on the length and strength of ectopic SHH signalling, a number of different malformations can be induced that model human limb malformations. We propose radial longitudinal deficiency with PPD is theoretically part of this spectrum and would be due to early strong and prolonged ectopic SHH expression.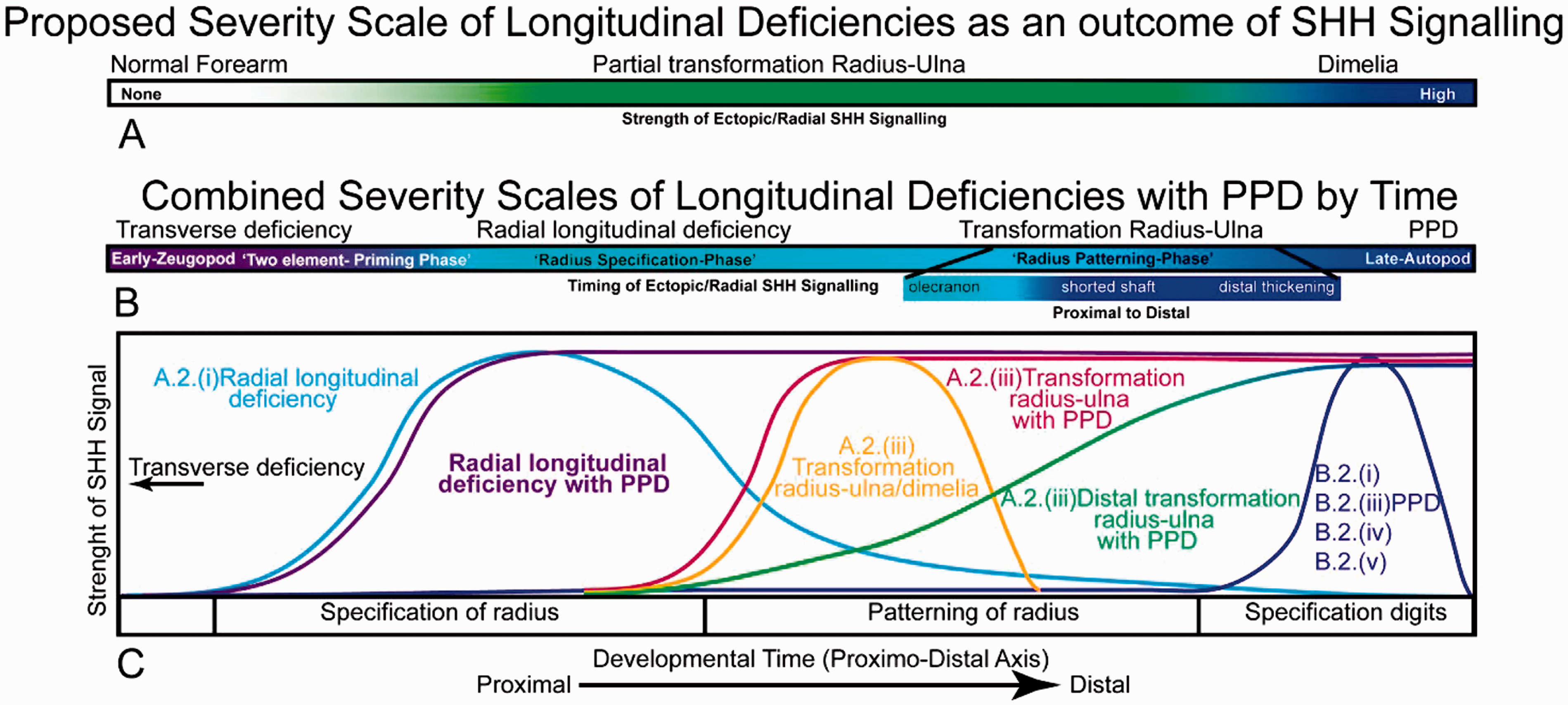
A classification system of human PPD based on strength and timing of ectopic SHH can therefore be proposed based on our work and that of others. For example, our results coincided with the hypothesis of Al-Qattan (2013) that minor to moderate ectopic SHH signalling results in PPD and its variants. Furthermore, we conclude that radial dysplasia and ulnar dimelia can occur in conjunction with ectopic SHH signalling and are likely to be associated with higher and earlier expression of ectopic SHH. Our main conclusion remains, however, that PPD, radial aplasia and ulnar dimelia are part of a single disease spectrum likely involving ectopic SHH activities.
Excerpt from the current OMT classification of congenital anomalies of the hand and upper limb (Oberg et al., 2010).

In summary, we found experimentally that PPD, radial dysplasia and ulnar dimelia can be part of a single spectrum involving aberrations in the SHH pathway. A unifying theory of disease pathogenesis may inform the hand surgeon about surgical anatomy and lead to new avenues of research.
Supplemental Material
Supplementary Figure -Supplemental material for Experimental evidence that preaxial polydactyly and forearm radial deficiencies may share a common developmental origin
Supplemental material, Supplementary Figure for Experimental evidence that preaxial polydactyly and forearm radial deficiencies may share a common developmental origin by Wee L. Lam, Julia D. H. Oh, Edward J. Johnson, Sandra Poyatos Pertinez, Chloe Stephens and Megan G. Davey in Journal of Hand Surgery (European Volume)
Supplemental Material
Supplementary Data -Supplemental material for Experimental evidence that preaxial polydactyly and forearm radial deficiencies may share a common developmental origin
Supplemental material, Supplementary Data for Experimental evidence that preaxial polydactyly and forearm radial deficiencies may share a common developmental origin by Wee L. Lam, Julia D. H. Oh, Edward J. Johnson, Sandra Poyatos Pertinez, Chloe Stephens and Megan G. Davey in Journal of Hand Surgery (European Volume)
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this work was primarily supported by The Roslin Institute via Institute Strategic Grant funding from the BBSRC research grant to MD. We also acknowledge funding from BSDB Gurdon Summer Studentship, the British Society for Surgery of the Hand and the Rooney Plastic Surgery Foundation.
This work was previously presented orally at the World Symposium on Congenital Malformations of the Hand and Upper Limb, May 2015 (best paper), European Association of Plastic Surgeons meeting, May 2015 and British Society for Surgery of the Hand Meeting, October 2015.
Supplementary material
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.