
Abstract
The term 'black holes' has been applied to anatomical areas needing further study. The historical account On Growth and Form (D’AW Thompson, Cambridge University Press, 1917) applies mathematical principles to explain how physical forces influence the morphology of bone, tendon, ligament and fascia. The trabecular structure of bone is maintained by force loading, providing a map of strain concentrations to better understand fracture fixation and bone healing. The flexor tendon is classified in zones of sheath pathology, but the site of tendon injury depends on the posture at the moment of injury. Ligaments are multistranded structures and in maintaining joint stability throughout motion different fibrous strands must share load by ligament buckling. The fascial description of Guimberteau visualizes the human body as consisting of microvacuoles, quantifiable from the physical Laws of Plateau. Human morphology can be understood through mathematics. These ‘black holes’ offer possible research avenues to manage some of the unsolved problems in hand surgery or to optimize outcomes.
Introduction
Despite centuries of anatomical study of the human hand, there remain many areas where structures have either been described quite differently by different authors, or perhaps wrongly described or not described at all. Those deficient areas where confusion or traditional views persist may be considered by application of the cosmological term ‘black holes’. Anatomy is often mistakenly considered by the inexperienced to be a dead subject where everything worthwhile has been discovered and recorded, but our knowledge has been constantly improving through the centuries and there is much more to explore.
One example of the potential for age, study and experience to improve the accuracy of anatomical description is that of Leonardo da Vinci. Firstly, as anatomist and later artist, he drew the brachial plexus in his early career with one nerve root absent, and a more accurate depiction followed in later years (da Vinci, n.d.). Another of Leonardo’s drawing of the human hand illustrates the flexor tendons as robotic machine-like forces, pointing the way to future biomechanical science, emphasizing that structure and function must be studied together.
Charles Bell (1833) made many contributions to description of anatomical structure and physiology, some of which have earned his eponym, e.g. long thoracic nerve of Bell’s palsy, Bell’s phenomenon, etc. He undertook many dissections working with his brother-in-law, publishing many illustrations, books and atlases. However, Bell’s analytical understanding extended far beyond structure into the realms of analysing why our morphology is the way it is. The role of environmental influences is apparent in his published Bridgwater Treatise: The Hand, Its Mechanism and Vital Endowments as Evincing Design (Bell, 1833). Bell understood that our anatomy conforms to physical forces: ‘The magnitude of the earth determines the strength of our bones, and the power of our muscles; so must the depth of the atmosphere determine the condition of our fluids, and the resistance of our blood vessels’. The influence of physics on evolving morphology in this Treatise published in 1833 did not capture the same attention as Charles Darwin’s Descent of Man 26 years later, and advocacy of the importance of physical forces had to await many decades.
The influence of physical forces was also noted by D’Arcy Wentworth Thompson (Figure 1), anatomist, polymath and philosopher working at the University of Dundee, who published in 1917 On Growth and Form, in which he used examples from many species including fossil evidence to propose that laws of physics and mathematics could explain morphology of plants and animals (Thompson, 1917). Considering bone in general, Thompson noted that tubular bone was thickened in the midshaft area to withstand stress and tapered towards the joints proximally and distally where stress was less. He noted that bone was essentially plastic, and the trabecular anatomy was reconfigured after fracture.

D’Arcy Wentworth Thompson 1860–1948. Author of On Growth and Form, 1917, which introduced mathematics to biology, explaining the role of physical forces in anatomical morphology.
In looking for black holes to develop there is a great deal of knowledge that predates Index Medicus, and therefore modern cell and molecular techniques if applied to historical observations can provide new understanding. And so, if as surgeons we can understand why we are the way we are, there is potential to capitalize much more on physical force and biological mechanisms to improve current therapies, including rehabilitation regimes.
The drive to define precise structure and function has been encouraged by new concepts in surgical reconstruction, and new imaging technologies such as four-dimensional MRI have stretched the boundaries of our understanding. Black holes do, however, remain and it is difficult to write a review with extensive literature as chosen examples seem to have insufficient literature. My apologies are due to scientists who may have written extensively on a subject of which I am unaware. In this article, examples of dark hole areas seeming to require new insight will be described with illustrations by a selection of tissues: bone, ligament tendon and fascia.
Bone
Bones are generally recognized and named from their outside morphology and there is a general appreciation of the influence of muscle origins and insertions on their shape and strength. In comparative anatomy there seems to be a pattern of limb skeletal structures which curiously appear to have common parameters across species. The mouse and the elephant each have five digital rays, as did many dinosaurs (BBC, 2024). It appears that all mammals have evolved from a pentadactyl ancestor and it remains hard to believe that five-fingered species have benefits over other numbers of digits that make them the fittest survivors, or perhaps there are physical laws as well as genetic influences at work. However, given the skeleton that nature has provided, there is a tendency to overlook the complexity of precise internal bone structure, which is important for understanding fracture patterns and means of internal fixation. Each bone is in fact a complex structure with a quite unique arrangement of its rigid internal component parts.
The German surgeon and anatomist Julius Wolff (1836–1902) (1986 translation of 1892 publication) noted that bone in a healthy animal will adapt to the load under which it is placed. The remodelling of bone in response to loading is a process of mechanotransduction by which mechanical forces are converted to biochemical signals (Huiskes et al., 2000). A common example is the map of the trabecular pattern in the head and upper shaft of the femur often described as complying with Wolff’s law. In a letter to Nature, Huiskes et al. (2000) considered the distribution of trabecular bone based on their understanding that the trabecular bone pattern was not genetically mapped but maintained and adapted in response to mechanical forces, as had been suggested by Thompson’s On Growth and Form (Thompson, 1917). Pursuing this concept by computer simulation, they considered that osteocytes were likely to be mechanosensors reacting to strain energy density, recruiting osteoblasts with the effect that the lamellae of bone are laid down in a laminated pattern that complies with strain. In Chapter XVI of On Growth and Form, Thompson (1917) further outlined an extensive bibliography over 100 years in multiple languages on the physical effects on tissues, but much of the signalling still requires clarification today. Different cells in different tissues respond to different mechanical influences, with each single cell influenced by its immediate mechanical environment. Much of the detail is beyond the scope of this review
Internal bone architecture
The upper limb has had less focus than weight-bearing bone (i.e. the lower limb or spine), but precise mapping of the internal structures of the distal radius has been described in detail by Bain et al. (2017) along with its relationship to fracture pattern. Using microcomputer tomography on dry bone specimens, they noted a strong subchondral bone plate supported by arches of trabecular bone. The trabecular structure of the metacarpals and phalanges, however, has not had the same attention from clinicians, although the internal structure of bone has been of interest in anthropology where the detailed morphology and therefore strength of bone have been of interest in defining hand and limb usage in extinct primates (Saers et al., 2022).
In preparing imaging data for ‘The Interactive Hand’, a teaching aid in CD-ROM format (McGrouther and O’Higgins,1997), a detailed series of MRI images with millimetric slices showed patterns of trabecula in great detail. Passing from proximal to distal across the proximal interphalangeal (PIP) joint a series of figures demonstrates the individual arrangement at each cross-section (Figure 2). Commencing in the mid shaft proximal phalanx region there is a well-defined thick cortex with very few if any trabecular structures within the medullary canal (Figure 2(a)).
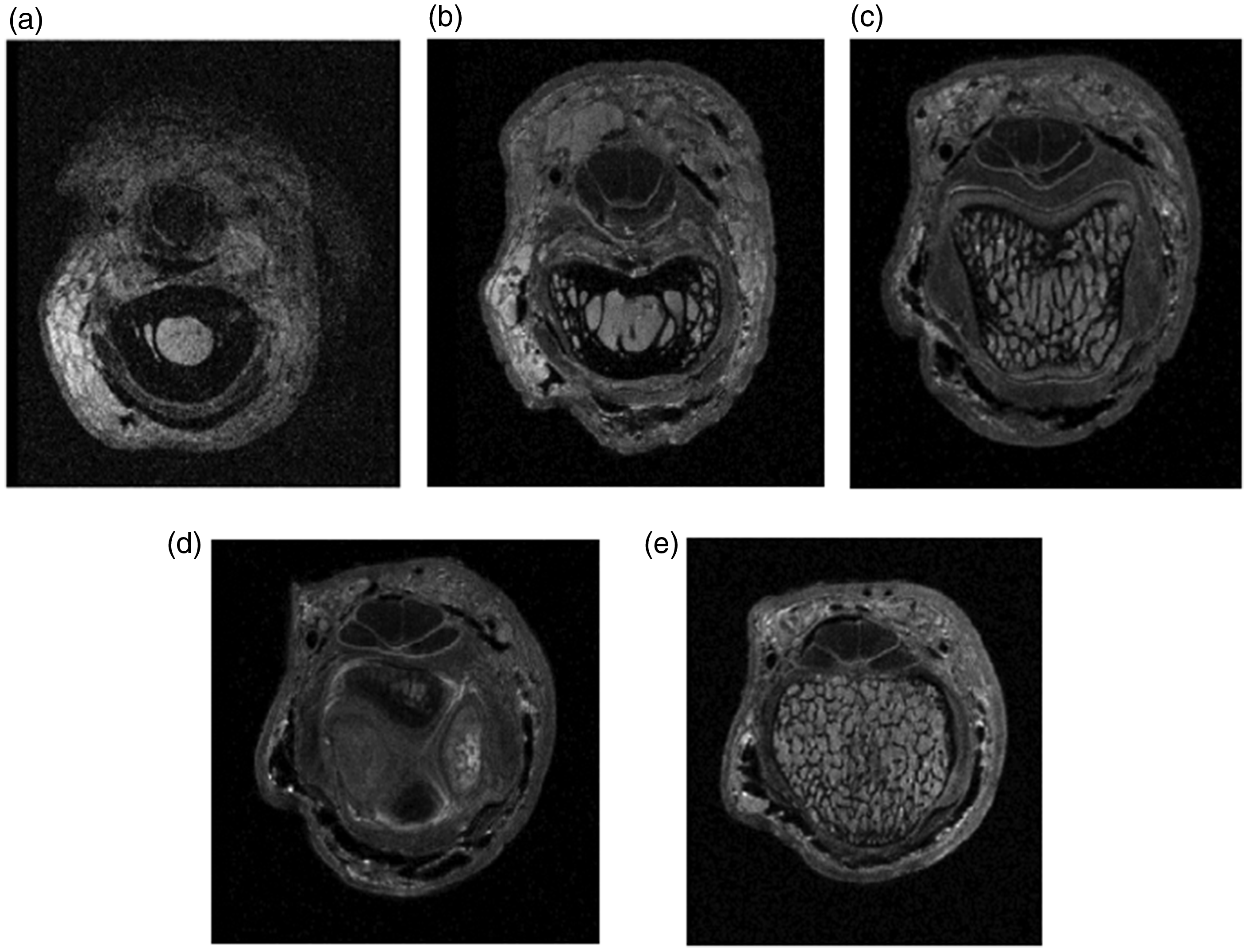
Bone morphology in relation to stress and strain. Cross-sections from a detailed MRI scan from proximal to distal. The Interactive Hand CD-Rom. First edition, 1997. (a) Mid proximal phalanx; cortex has a well-defined tubular form and there are few trabecula. (b) Distal proximal phalanx; commencement of transfer of load bearing from cortex to trabecula and trabecula more obvious. (c) Head of proximal phalanx; cortex dramatically thinned and trabecula well developed and longitudinally orientated. The collateral ligaments are thickened at this point where they are arising from the sulci in the head of the proximal phalanx. (d) Cross-section through articular contact surface of the proximal interphalangeal joint. High spots of proximal phalanx medial and lateral. High spots of middle phalanx flexor and extensor surfaces and (e) cross-section through base of middle phalanx; cortex very thin and trabecula orientated longitudinally and transversely forming multiple small cuboidal cellular structures.
Passing distally towards the PIP joint, the cortex progressively thins (Figure 2(b)) with a suggestion of delamination of the inner margin; as a result, the cortex becomes less sharply defined. The trabecular component becomes much more defined and longitudinally orientated in the head of the proximal phalanx (Figure 2(c)). At the joint region the cortex is just a thin shell (Figure 2(d)) covered by cartilage on the contact surfaces, comprising a structure for load transmission with some flexibility. In contrast, distal to the proximal interphalangeal joint line the trabecular pattern in the base of the middle phalanx has finer laminations (Figure 2(e)) orientated both longitudinally and transversely, displaying a very different load bearing pattern amounting to multiple cubic compartments. These different patterns map out the strain pattern in the proximal phalanx head as longitudinal whereas the compressive strain in the base of the middle phalanx appears multidirectional.
The process by which trabecular pattern is achieved has been discussed by Huiskes et al. (2000) with evidence that the maintenance and adaptation of form are dependent upon stimulation of trabecular bone formation, essentially owing to shearing strains in the bone. Using computer simulation, they describe how given a chosen direction of strain energy density bone hypertrophy is likely to reinforce the format of architecture rather than forming different patterns. These strain patterns determine that traumatized bones will fracture in different ways. Excess longitudinal strain in the proximal phalanx head is likely to result in a unicondylar or bicondylar fracture whereas the bone at the base of the middle phalanx will crumple and collapse under impact loading.
These anatomical trabecular structures also give clues to the deformity patterns in degenerative joint diseases. A detailed study of hand osteoarthritis by Simon et al. (2020) using high-resolution peripheral quantitative tomography has shown decreased trabecular density and increased cortical density in the metacarpal heads in hand osteoarthritis. A trabecular bone score using X-ray diffraction studies has been described as a predictor of compression fractures in vertebrae, but does not seem to have been applied to hand arthritis (Schousboe et al., 2002).
There is a need for a more extensive study and mapping of the inner anatomy of hand bones to give a clearer picture of fracture patterns, and consequently, more information on the best means of fracture management. We know from experience that collapsed fractures of the base of the middle phalanx may require traction or bone grafting (Kolovich and Heifner, 2023), but it is not yet known how we should best deliver the strain necessary to stimulate new bone generation. We should understand that areas of thin cortex adjacent to joints may not provide firm screw fixation. There are many other applications of better trabecular knowledge in trauma management.
Another potential application is in small joint arthroplasty. The difficulty in developing successful long lasting joint replacements in the hand is due in part to a lack of understanding of the mechanical influences that preserve and strengthen bone stock. Despite recent advances, modern arthroplasty designs may result in stress shielding rather than bone promotion. There is a need for further detailed study of microanatomy and strain-driven mechanisms of regeneration in our pursuit of the perfect prosthesis.
Tendon
The subject of flexor tendon injury and repair has been considered at length by this author in the conference book of the 2022 International Federation of Societies for Surgery of the Hand (McGrouther, 2022). As these comments were based on more than half a century of personal experience of tendon anatomy, it would be surprising to suddenly write about a new series of blackholes. There will inevitably follow a repetition of concepts and text but with further pointers to the younger generation of gaps that need to be filled.
Synovial sheaths
The macroanatomy of digital synovial sheaths was beautifully demonstrated by Alan Kanavel (1922) by injection of barium in cadavers using a pump with controlled pressures. He showed that there are separate synovial sheaths in the index, middle and ring digits distal to the A1 pulleys, but sheaths in continuity from digit to carpal tunnel in the case of the thumb and little finger, which incidentally have the least flexor tendon excursion of the digits. Cases of horseshoe abscesses of the flexor sheaths support the likelihood that thumb and little finger sheaths are connected in the carpal tunnel, at least in some patients. With the development of new mini-arthroscopic techniques there must surely be potential for future sheath imaging and intrasynovial repairs (Figure 3).

Arthroscopic examination of ring finger flexor sheath and tendon arrangement. (a) Instrumentation setup; and (b) image showing A1 pulley, with flexor digitorum superficialis (FDS) lying above the flexor digitorum profundus (FDP).
Focussing on the dynamic mechanisms of the tendon within the sheaths, many concepts of the arrangement of tendon synovium are based upon the description of Mayer (1952) in relation to his original study which was of the tendon of the tibialis anterior. That tendon had a loose fascial covering which he described as paratenon when situated outside a sheath. Within the sheath at the ankle, the tendon had a compact outer layer which he described as an epitenon and internal tissue within the substance of the tendon described as an endotenon. At the proximal pole of sheaths of many different tendons he described a loose fold, termed a plica, allowing invagination on movement. Within this sheath and on the non-contact surface of the tendon he described a thin membrane to supply blood vessels described as a mesotendon. Despite these studies, a full description of the blood supply of tendons is still lacking and there is little evidence for an avascular zone in flexor tendons. There remains considerable variability in tendon sheath descriptions and subsequent attempts by many authors to describe a common format and nomenclature for synovial sheaths of different tendons have been unconvincing. Terms such as epitenon, endotenon, paratenon and mesotenon have considerable variation in different anatomical sites and should be clearly defined or better replaced.
Tendon gliding and resistance
In the uninjured finger, flexor tendons have a frictionless excursion within the flexor sheaths and as tendons glide from intrasynovial to extrasynovial areas it is necessary to appreciate the connective tissue arrangements that allow this smooth motion at the junction to take place. Both Kanavel (1922) and Doyle and Blythe (1975) illustrated sleeve-like arrangements reminiscent of a bowel intussusception as a mechanism at the margins of synovial sheaths, but neither this arrangement nor Mayer’s plica would seem to allow enough free movement for the 35 mm of flexor tendon excursion in the mid palmar region. Guimberteau (2001) has demonstrated multiple fascial layers gliding upon one another in his videotape recordings (see below).
As mentioned, direct observation of tendon gliding using modern micro-endoscopy techniques may help our understanding of tendon pathology (Figure 3). A common description of the outer surface of the flexor tendon as a visceral synovial layer and a lining of the sheath as a parietal layer is an oversimplification as both tendons and sheath have many different areas of specialization which require further detailed cellular description. In areas of contact between tendons and between tendons and annular pulleys the synovium is quite different and needs better description. There is a clear need for new studies of tendon synovium employing cell and molecular technology rather than relying upon dated concepts derived from histopathology or naked eye observation (Cohen and Kaplan 1987). The A1 pulley is one area that has had detailed study (Sampson et al.,1991) and has been shown to have a covering of amorphous extracellular matrix including hyaluronan and lubricin (Taguchi et al., 2009). This surface is fragmented and partially absent in trigger finger, suggesting a shearing injury which in turn suggests that trigger finger may be initiated by a ‘degeneration’ of the delicate lubrication system. Where the cellular pathology lies in trigger finger remains to be seen; I know of only two papers on tendon biopsy of living patients and no functional problems were reported, yet there are many papers on A1 pulley biopsy.
Another area for research pertains to the palmar and dorsal surfaces of the tendons which have very different function and structure. The delicacy of the synovium has made histological study of the surface of the flexor tendon challenging. Edwards (1995) noted that synoviocytes are specialized fibroblastic lineage cells which he stained by uridine diphosphoglucose dehydrogenase (UDPGD). He described a synovial intima of overlapping cells rarely more than three cells thick without a basal lamina or tight junctions, and this cell pattern was incomplete in several areas. More recent studies (Kemble and Croft, 2021) on joint synovium have confirmed this general arrangement of a monolayer or near monolayer of synoviocytes (a basement membrane is now identified), supported by a subsynovial layer of loosely packed fibroblasts and macrophages. This work needs to be repeated on tendon synovium; standard histological preparation has been found to largely remove the surface layers but previous work in our laboratory has managed to image surface cells and our interpretation was that surface cells have numerous surface flagellate processes which appear to bind a layer of hyaluronidase and lubricin which in turn bind water to give a fluid film, thus reducing friction (Kakar et al, 1998). Given the delicacy of this arrangement it would seem necessary for it to have cellular mechanisms of repair and regeneration, possibly owing to macrophages. Tenocytes, which are tendon stem progenitor cells, have been identified but their role and their niche remain uncertain. A recent paper by Chen et al. (2025) has identified a tendon stem/progenitor cell residing in peritendon with self-renewal and multipotent differentiation potentials. These cells were shown to emigrate into the tendon mid substance and differentiate into tenocytes during tendon healing.
Finally, a general finding in various species is that compression areas within tendons have a chondroid appearance and this is also apparent, for example, on weight-bearing areas of the feet of rodents. It is rather less widespread in man but contact areas between two flexor tendons have a chondroid appearance (Walbeehm and McGrouther, 1995). Merrilees and Flint (1980) employed a series of experiments in which they modified the musculotendinous anatomy in various ways; by dividing most of rabbits’ tendons in the leg but sparing one tendon and then allowing all of the proximal muscles side by side to apply traction through the residual tendon they managed to place this tendon under excess lateral force as it passed through a retinaculum. The effect was that the tendon area under lateral compressive force changed its glycosaminoglycans towards a much more chondroid pattern. They then had an escape experiment whereby on allowing the tendon to bowstring and straighten up the chondroid biochemistry reverted to a tensional tendon profile. These were ingenious experiments performed before the terms cell biology and molecular biology were in common use and these experiments need to be repeated with up-to-date technology to refine the molecular changes.
Annular pulleys
The A1 pulley has been shown to have chondroid metaplasia (Sampson et al., 1991) whereas the dorsal surface of the tendon has loose gliding synovial tissue with abundant blood supply fed by vincular vessels reinforcing an intratendinous longitudinal supply This vascularized tissue appears to have the function of producing an ultrafiltrate of serum to provide fluid for tendon nutrition or lubrication. Tendon also has an important longitudinal blood supply which seems likely to be disrupted by any method of suture in tendon repair. Synovial surfaces need to be an area of focus for the future to understand and manage ‘adhesions’, which are currently the greatest challenge in improving outcomes.
Synchronized flexor and extensor tendon biomechanics
A particular Black Hole is our understanding of the complex biomechanics of integrated flexor and extensor function. If an engineer were to devise a machine to drive four linked mechanisms of unequal length it would not work and it is not clear how the hand does this. Of note, the flexor digitorum profundus muscle complex has a double innervation, with a general pattern of index and middle innervated by the anterior interosseous nerve and the ring and little by the ulnar nerve, suggesting an evolutionary process as separate muscles but presenting a neurological challenge in control of simultaneous motion. With so many bones and joints motored by so many different muscles under the control of three peripheral nerves it is amazing that the hand functions as it does in such a co-ordinated fashion.
Flexor tendon zones
Zones are an arbitrary concept which corresponds to a grouping of certain chosen lengths of the synovial sheaths (Figure 3). They do not however adequately describe tendon anatomy within the sheath and a very different pattern of tendon division may be found depending upon whether the wound occurs in flexion or extension (Figure 4). Moreover, the exact degree of digital flexion at the moment of injury (Figure 5) will alter other parameters of tendon anatomy − blood supply, innervation, chondroid specification and degeneration and lubrication. Some new system of classification is needed to characterize what factors are really important to determine clinically relevant flexor zones.
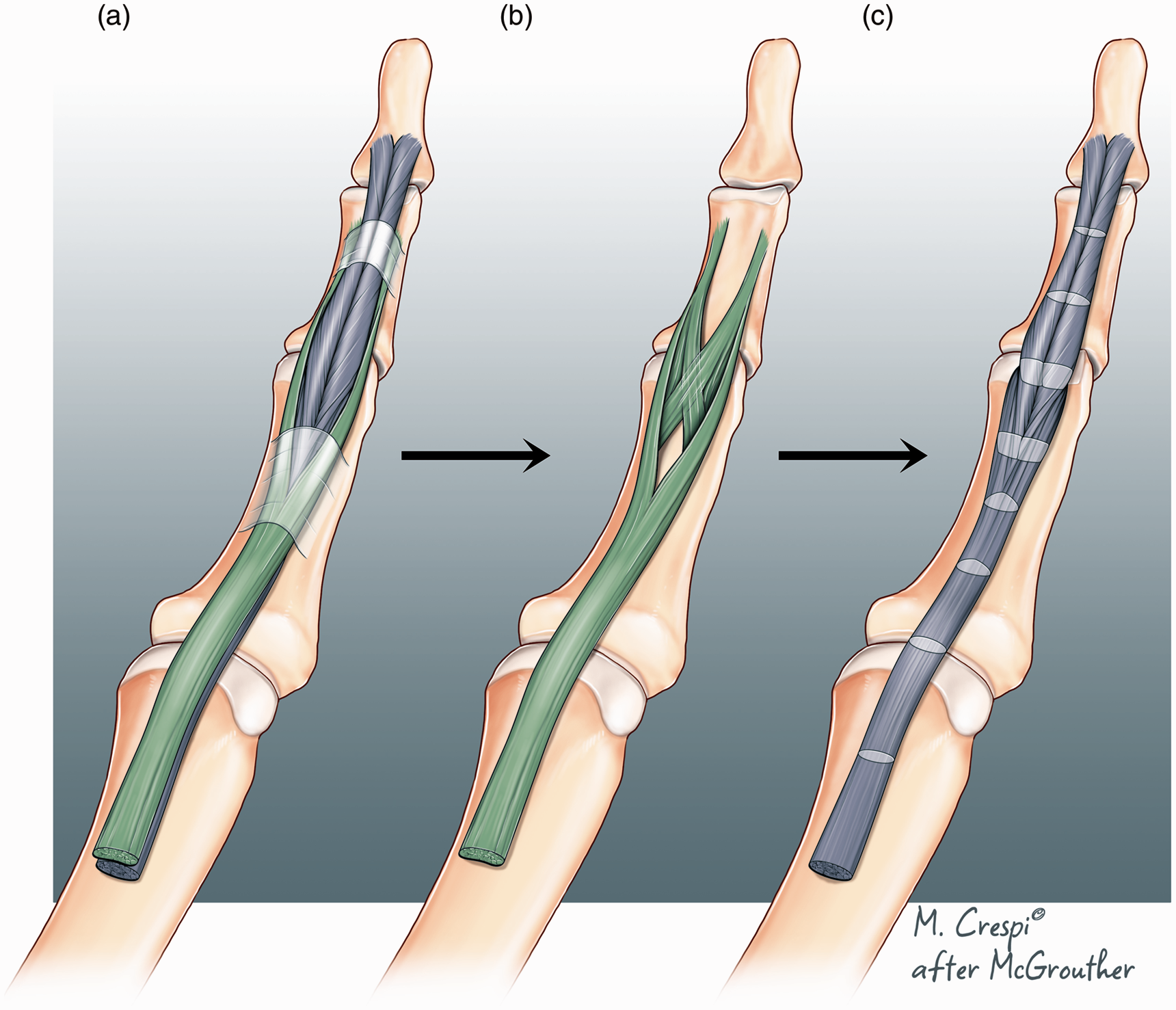
Flexor tendon anatomy. (a) Normal anatomy of flexor digitorum superficialis (FDS) and flexor digitorum profundus (FDP) in a relaxed digital posture. Distal FDP is bifid. Redrawn from dissections and sketches kindly provided by Professor Erik Walbeehm. (b) Normal anatomy of FDS. Modelled after Schmidt et al. (1994) and (c) normal anatomy of FDP alone. Note that the fibres spiral within the FDP tendon in a pattern which parallels the spiralling pattern of FDS. On passing from proximal to distal most lateral fibres curve around the outer tendon from anterior to posterior. The lighter coloured bands show different cross-sectional profiles at different points.
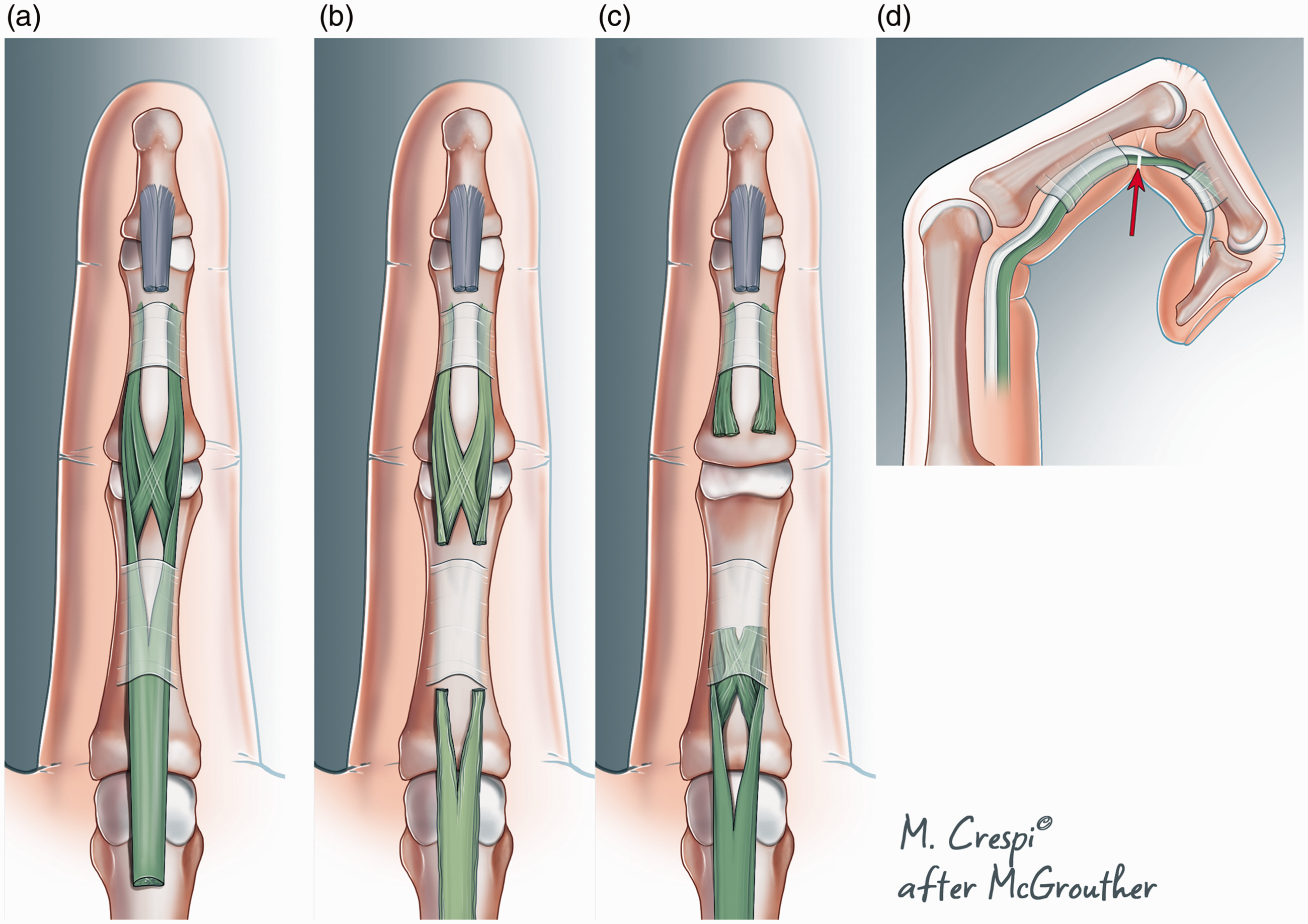
Different pattern of flexor digitorum superficialis (FDS) injury with digit in extension or flexion. (a) Normal FDS modelled after Schmidt et al. (1994). FDP has been divided distally and removed. (b) Pattern of FDS location after division with finger extended, just distal to the A2 pully. (c) Site of FDS injury with finger flexed. There is less proximal retraction when the finger was flexed during injury. However, the amount of proximal retraction in situation (b) or (c) will also be determined by the applied load at the moment of injury and (d) the site of tendon laceration with the finger flexed.
Tendon adhesions
‘Adhesions’ (Wong et al., 2010) are a very non-specific clinical term after a tendon trauma as surface adhesion is only a small part of the zone of cellular injury. Deep to the surface adhesion there is a dense zone of cellular inflammatory change in both damaged tendon and surrounding tissues. An outdated concept is the theoretical potential for intrinsic vs. extrinsic healing. Many cell populations participate in tendon healing inevitably from both within the tendon and neighbouring structures. The surface cells have been shown to migrate into a wound defect and also through the tendon substance. Distant stem cells may also be involved as many tenocytes will necrose after injury and many more after surgical repair (Wong et al., 2010). There is a need for us to know much more about the cell and molecular participation, both local and distant, in order to optimize our management of tendon injuries.
Degenerate tendon conditions
Tendons suffer a range of ‘degenerative’ tendon pathologies around the fifth decade of life. When encountered in significantly younger patients there is usually a history of physical overuse activities in the gym, repetitive job stresses or carrying heavy objects or young children. Tendinopathy conditions have in common an anatomical basis of a tendon being under pressure, but the exact pathology differs. The tendon in trigger finger may have little to show when explored other than some yellowish discolouration or a dry surface, whereas in de Quervain’s tenosynovitis the tendons can be visibly and painfully inflamed, swollen and hyperaemic. It is not clear if the different pathology is due to different mechanical stress or different cell biology. De Quervain’s involves two different tendons undergoing multiple different axes of wrist movement whereas trigger finger has a single bending axis. Triggering occurs (Chuang et al., 2017) at an area of anatomical thickening of the combined flexor tendon mass where the flexor digitorum superficialis splits and encircles the flexor digitorum profundus. The trigger occurs when the tendon overall is swollen for reasons yet to be determined.
Ligament
The third structure we will look at is ligaments. Ligaments are condensations of viscoelastic fibrous tissue with roles in influencing motion and resisting unwanted motion in skin anchorage or joint stability. Joint ligaments are fibrous structures that maintain contact of articular cartilaginous surfaces during a range of movements. Some ligaments seem to have a role throughout joint motion while others seem to have a major role in checking the end of a range of motion; the anterior cruciate ligament in the knee is a good example.
Ligament in hand joints
In considering what is required in the hand the interphalangeal joints have a very specific requirement of preventing subluxation or dislocation not just at the end of a range of motion but contributing to stability throughout the range of joint flexion or extension. Thinking about this geometrically and understanding that the PIP collateral ligaments have broad areas of origin and insertion on the phalanges (Figure 6), any individual fibre would only be under tension at one point of the range of joint motion and otherwise it would be slack. It might be expected that the dorsal edge of the ligament would be tense on flexion but slack on extension, but this was not apparent in cadaveric studies (Allison, 2005). Clearly a different mechanism was required to maintain stability in this complex three-dimensional ligament structure. Allison (2005) also noted that as the PIP joint flexed, bulges appeared in the ligaments with the fibres of one part of the ligament rolling over its neighbours (Figure 7). Rather than any fibres becoming slack they maintained their tension by adopting a curved course from origin to insertion bunching up in a more curved alignment.
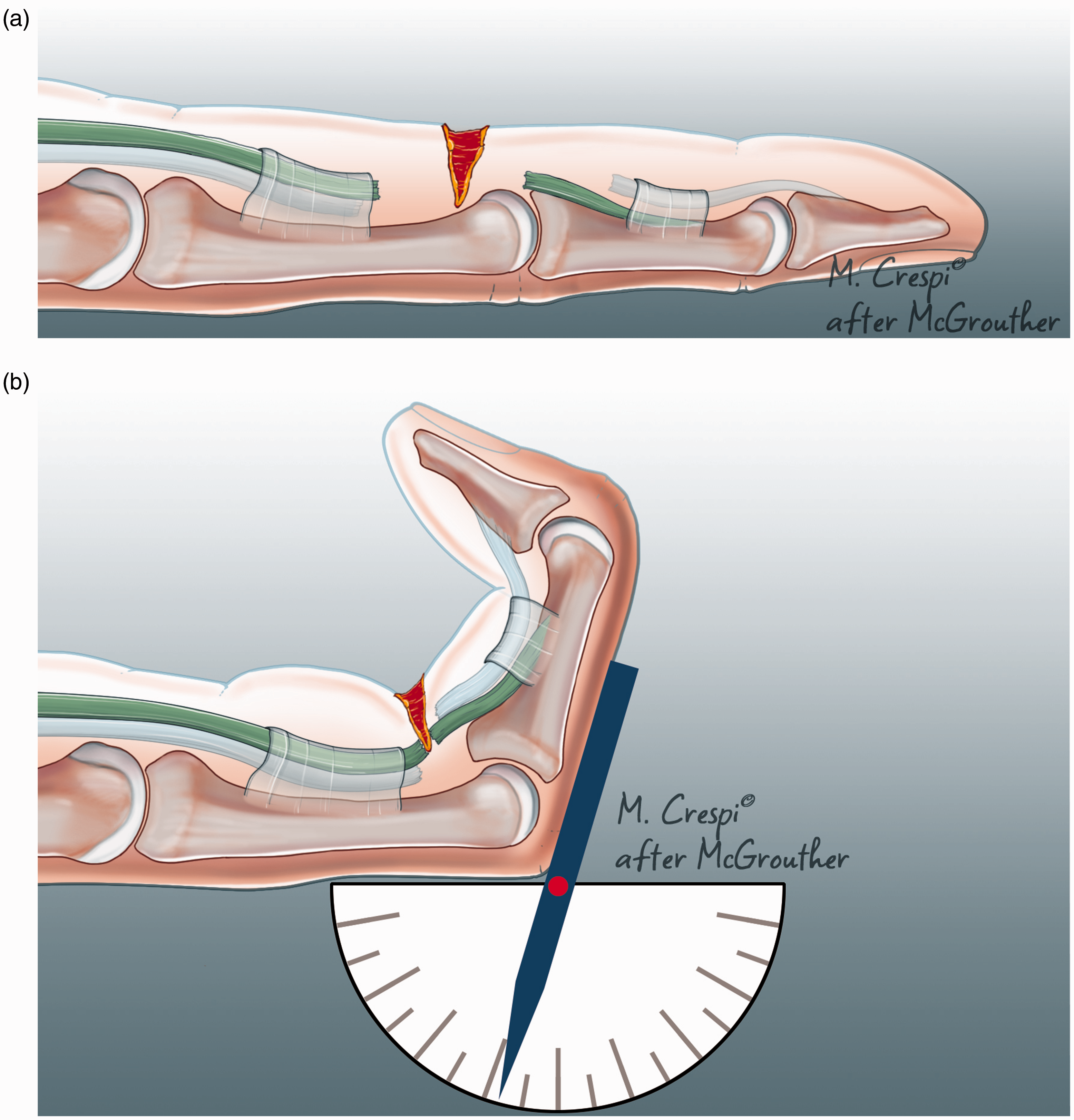
Recreation of the digital posture at the moment of injury. (a) When the patient with a flexor tendon injury presents at the A&E department it is not possible to know whether the injury was sustained in flexion or extension although there may be clues from the history and (b) it is possible to recreate the posture that prevailed at the moment of injury by flexing both interphalangeal joints until the distal cut ends of FDS and FDP are aligned with the wounds of skin and sheath, This will aid in locating the tendon ends, planning the incision and by recording the joint angles this provides potential data for outcome analysis.
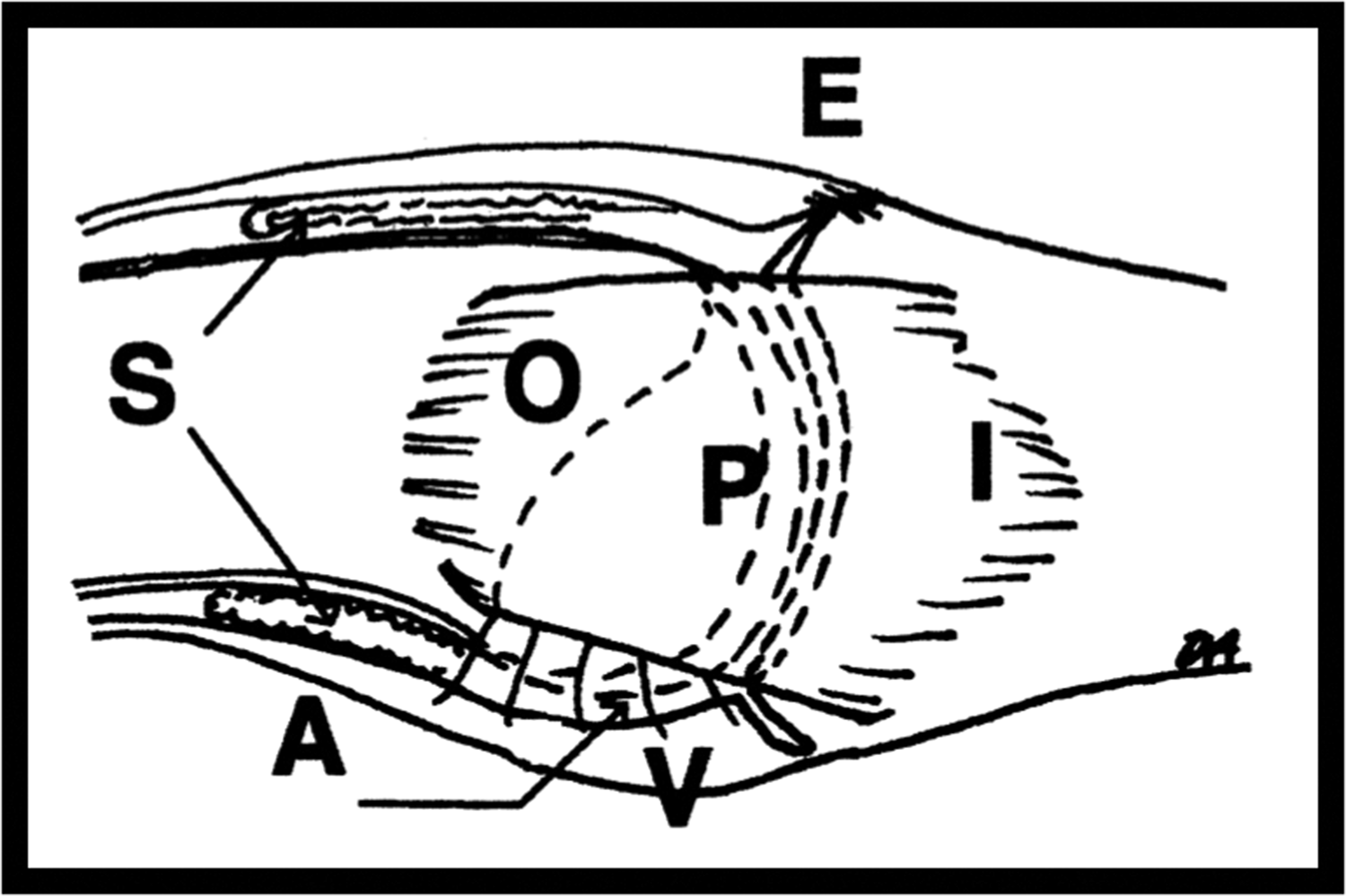
Collateral ligaments of the proximal interphalangeal joint (PIP joint) showing attachment to bone. E, Extensor insertion; V, volar plate; O, origin of collateral ligament proximal to dotted line; I, insertion on middle phalanx; P, proper collateral ligament; A, accessory collateral ligament. Illustrations by kind permission of Mark Allison. J Hand Surg 2005; 30A: 1026–1031.
Figure 2(c) illustrates the remarkable thickness of the collateral ligaments, a detail that had not been made plain in previous illustrations. The bulging of the collateral ligaments increases their strength such that trauma is more likely to result in fracture rather than total ligament disruption although partial tears are not uncommon. The ligament pattern of the PIP joint requires further study of the microanatomy and dynamics of movement, although every ligament in the hand is a candidate for microfibrous analysis during joint movement. This could be an excellent computational study which could greatly contribute to our understanding of fracture patterns around the hand joints.
Fascia
The last structure we will examine is the humble fascia. Fascia is an anatomical term which encompasses many different types of tissues for static or dynamic movement, restraint or lubrication (Guimberteau et al., 2010). The most obvious macroscopic structures have been named, including the deep fascia, fascia lata and palmar fascia, but their mechanical roles of restraint and lubrication have long been underplayed. Traditional anatomical cadaveric dissection has often removed delicate fascial tissue to access solid organs without considering its static structure or dynamic behaviour.
A detailed anatomy of fascia
In the hand, Jean Claude Guimberteau (2001) has given us an entirely new concept of fascial structures by high-magnification studies in living patients at operation. He has used video as a medium to explain his innovative concepts of fascia dynamics of movement rather than relying upon more traditional and static forms of academic literature publication. Relying upon his operation fields as the basis for his studies he has avoided the problems inherent in cadaveric dissections of desiccation and specimen deterioration. Much of the delicate tissue he describes can be damaged at open surgical procedures owing to surface drying, chemical toxicity and traumatic surgery. Offset against these benefits of video recording of dynamic movement are the problems of rapid change of format in electronic teaching media limiting dissemination of knowledge.
Guimberteau’s new concept of the structure of the human body explains many confusing historical findings, such as the fact that the human body is 60% water. Essentially, we are a mass of fluid-filled spaces of different dimensions both intracellular and extracellular. Guimberteau (2001) views fascia as a network of fine fibrous filaments (Figure 8) in a fractal configuration, best explained as similar patterns recurring at progressively smaller or larger scales. The tiny three-dimensional spaces between delicate fibres are described as microvacuoles. The whole structure of our microvacuolar system is therefore analogous to the structure of soap bubbles where round bubbles packed together become facetted in shape and comply with the Laws of Plateau (they meet in threes at angles of 120°). These laws were formulated in the nineteenth century by the Belgian Physicist Joseph Plateau and explained recently by the science author Philip Ball (2011).
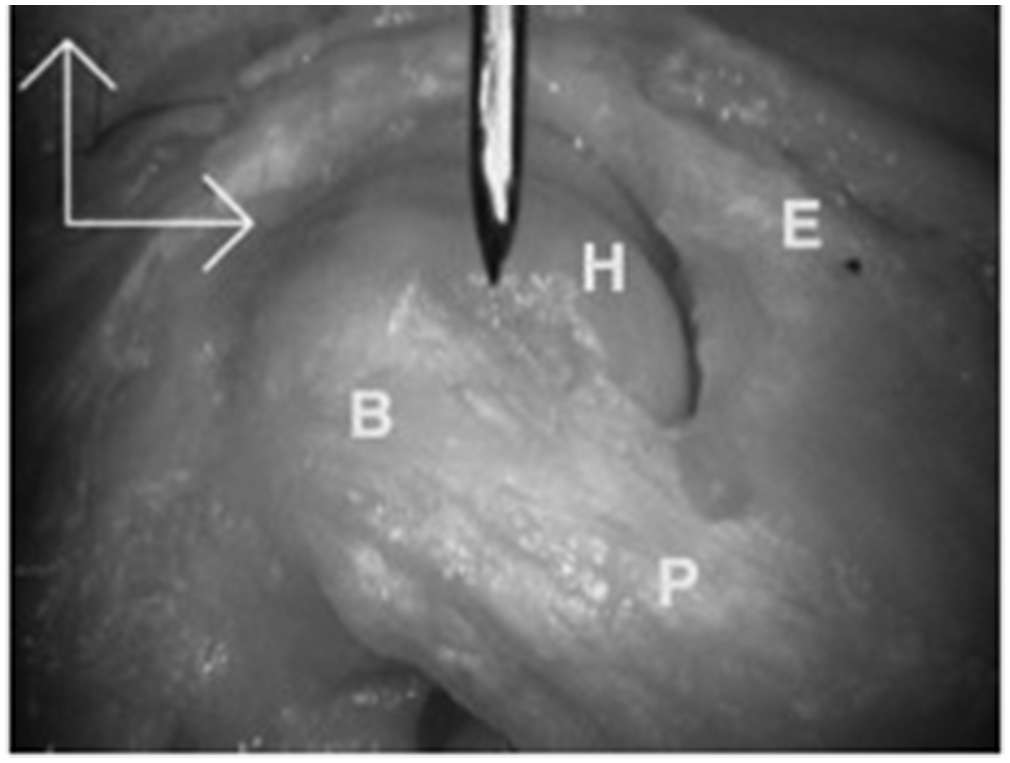
Lateral view of the proximal interphalangeal joint (PIPJ) in flexion. E, Extensor partly removed to expose H, proximal phalanx condyle. On PIPJ flexion, a bulge, B, appears in the proper collateral ligament, P, owing to folding as fibres roll over their neighbours.
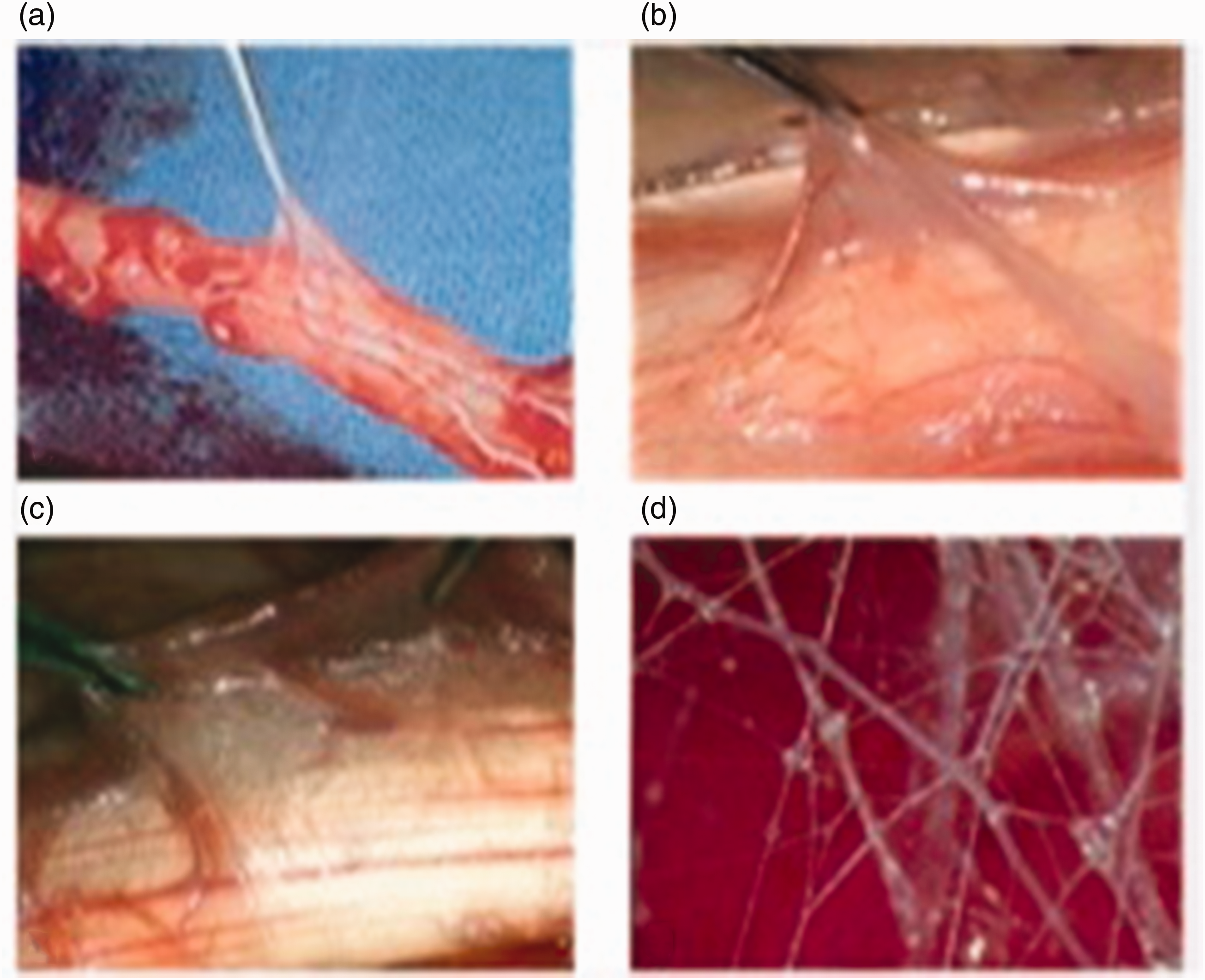
Fascia after the observations of Jean Claude Guimberteau: (a) low magnification image of fascial structure; (b) higher magnification view of mobile fascial structure; (c) higher magnification – bubbles of microvacuoles seen in junctional areas; and (d) individual connective tissue fibres criss crossing to form boundaries of microvacuoles.
Blood vessels in a zig -zag pattern course through this fascia plexus with enough laxity to allow motion between layers of microvacuoles. Three-dimensional movements of any area of tissue result in the fibres changing orientations and thus altering the shape of the microvacuoles. On this pattern, Guimberteau et al. (2010) describes the flexor tendon synovial sheath as a larger macrovacuole with blood vessels as vincula traversing the vacuoles. Such a complex arrangement in a structure often considered insignificant and removed without hesitation has been demonstrated by Guimberteau (2001) to play a key role in the frictionless excursion of flexor tendons. It is therefore unfortunate that Guimberteau’s tissue dynamics and fascial science have not had the recognition and wide understanding that they deserve. There should be no black hole here but only something waiting to be discovered and announced.
Conclusion
The framework of Guimberteau meets with the philosophy of D’Arcy Wentworth Thompson, where this article commenced, about the role of physics and mathematical laws and quantitation of our tissues. Even as modern surgery advances, any procedure will continue to be blighted by scar, fibrosis and adhesions. The supreme challenge in innovation of repair of all tissues is to be able to regenerate Guimberteau’s microvacuolar fascia. An enhanced appreciation of the different internal bony architecture will allow us to understand more about fracture patterns. A fresh look at the microanatomy of tendons will help us achieve better outcomes following injury. The same can be said for ligaments, where mere assumptions must give way to novel biomechanical studies. It is hoped that these few examples will hopefully show where black holes still exist in hand surgery, with potential research areas for our enlightenment of this wonderful organ.
Footnotes
Declaration of conflicting interests
At the time of first drafting of this review the author was Senior Hand Surgeon at Singapore General Hospital and is now retired. There are no conflict of interests, no funding or grant support. Ethical approval not sought as no patient contact or referral to patient records. This paper is a single author review.