
Abstract
Innate γδ T cells expressing Vγ6 produce IL-17A at an early stage following infection with Mycobacterium bovis Bacillus Calmette-Guérin (BCG). In this study, we used IL-21 receptor knockout (IL-21R KO) mice and IL-21-producing recombinant BCG mice (rBCG-Ag85B-IL-21) to examine the role of IL-21 in the regulation of IL-17A-producing innate γδ T-cell response following BCG infection. IL-17A-producing Vγ6+ γδ T cells increased in the peritoneal cavity of IL-21R KO mice more than in wild type mice after BCG infection. In contrast, the number of IL-17A-producing Vγ6+ γδ T cells was significantly lower after inoculation with rBCG-Ag85B-IL-21 compared with control rBCG-Ag85B. Notably, exogenous IL-21 selectively induced apoptosis of IL-17A-producing Vγ6+ γδ T cells via Bim. Thus, these results suggest that IL-21 acts as a potent inhibitor of a IL-17A-producing γδ T-cell subset during BCG infection.
Introduction
Conventional Th17 cells are exported from the thymus as naive cells, and then acquire effector functions, and produce IL-17A upon Ag encounter in the periphery. However, some γδ T cells are functionally committed to be IL-17A-producing effector cells within the thymus. Therefore, these γδ T cells are also called ‘innate γδ T cells’.1–5 Innate γδ T cells are disproportionately distributed in mucosal tissues such as the uterus, lung and peritoneal cavity,3–5 and spontaneously produce IL-17A at an early stage following infection with Mycobacterium bovis Bacillus Calmette-Guérin (BCG),6–8 Citrobacter rodentium, 9 Salmonella enterica serovar Typhimurium, 9 Listeria monocytogenes,10,11 Escherichia coli 12 or Candida albicans. 13 Thus, innate γδ T cells bridge the gap between innate and adaptive immunity in terms of the kinetics of the immune response following microbial invasion.
Various exogenous signals, via the common gamma (γc) chain receptor (R), play an important role in regulating γδ T-cell IL-17A or IFN-γ production. The IL-15/CD125 (IL-2Rβ) axis is important for γδ T-cell IFN-γ production, 3 while the IL-7/CD127 (IL-7Rα) axis is critical to IL-17 A production.14,15 IL-21 is a pleiotropic γc cytokine secreted by activated CD4+ T and NKT cells. The IL-21 R is widely expressed on various cell types within the immune system, including macrophages, dendritic cells, NKT, CD4+ and CD8+ T cells, and B cells.16–22 The widespread lymphoid distribution of IL-21 R leads to pleiotropic actions of IL-21 in the innate and adaptive immune responses, including host defense against infection, antitumor activity, and pathogenesis of inflammatory diseases such as autoimmunity and allergic diseases.20–22
Do et al. reported that IL-21 was dispensable for γδ T-cell IL-17 A expression in lymph nodes (LN), 23 while Moser et al. reported that IL-21R signaling suppressed the IL-17A-producing γδ T-cell response in lung after influenza A virus infection. 24 Thus, the role of the IL-21/IL-21R axis in innate γδ T-cell response remains obscure. In this study, we examined the role of IL-21 in the regulation of IL-17A- producing innate γδ T-cell response following BCG infection, using IL-21R knockout (KO) mice and IL-21-producing recombinant BCG mice (rBCG-Ag85B-IL-21), which we constructed. We found that IL-17A-producing Vγ1−Vγ4− γδ T cells in rBCG-Ag85B-IL-21-inoculated mice significantly decreased, whereas they increased in BCG inoculated il21r−/− mice. IL-21 induced selectively apoptosis of IL-17A-producing γδ T cells via Bim. Thus, this suggests an immunoregulatory role for IL-21 in innate IL-17A-producing γδ T-cell response during BCG infection.
Materials and methods
Mice
Age- and sex-matched C57BL/6 (B6) male mice were purchased from Japan KBT Inc. (Tosu, Japan). Il21r−/− mice were obtained from Taconic (Hudson, NY, USA) and backcrossed for eight generations onto a B6 background. 17 Il17g/g mice, in which the IRES–eGFP was inserted into the Il17a locus without affecting IL-17A production, were described previously. 25 All mice were maintained under specific pathogen-free conditions and provided food and water ad libitum. All mice were used at 6–8 wk of age. The Committee of Ethics on Animal Experiments of the Faculty of Medicine, Kyushu University approved all animal experiments.
Microorganisms
Lyophilized M. bovis BCG (Tokyo strain) was purchased from Kyowa Pharmaceutical (Kyoto, Japan) and grown in 7H9 medium (Difco, Detroit, MI, USA) supplemented with albumin–dextrose–catalase (Difco). For construction of IL-21-producing rBCG, pNN2-mIL-21-Ag85B was introduced into BCG (strain Tokyo 172) by electroporation as previously reported. 26 Transformed BCG were plated on Middlebrook 7H10 agar supplemented with 10% oleic acid–dextrose–catalase, 20 µg/ml kanamycin, 100 U/ml penicillin G and 100 µg/ml cycloheximide (Difco). After growing for 3 wk at 37℃, single colonies were picked and grown in Sauton’s medium for 3 wk at 37℃. Mice were inoculated i.p. with viable M. bovis BCG, rBCG-Ag85B-IL-21 or rBCG-Ag85B in 0.2 ml PBS on d 0 at a sublethal dose of 1 × 106 BCG CFU or 2 × 106 rBCG-Ag85B-IL-21 or rBCG-Ag85B CFU. On the indicated days after inoculation, mice were euthanized, and peritoneal exudate cells (PECs) were subjected to lavage with 1 ml ice-cold Hank’s balanced salt solution and harvested after gentle massage. Samples were spread on Middlebrook 7H10 medium enriched with oleic acid–dextrose–catalase, and colonies counted after incubation for 3 wk at 37℃.
Detection of IL21-Ag85B fusion protein
rBCG-Ag85B-IL-21, rBCG-Ag85B or BCG were lysed in lysis buffer for 30 min or cultured in 7H9 medium for 1 wk. After centrifugation, supernatants were harvested. Cell lysate or supernatant protein quantification was performed using Ultrospec 3300 pro (Amersham Biosciences, GE Healthcare Biosciences, Piscataway, NJ, USA). Samples were diluted to the same amount of protein, separated by SDS-PAGE, and proteins blotted and detected with anti-IL-21 mAbs, (Biolegend, San Diego, CA, USA). Culture supernatant IL-21 was measured using a DuoSet ELISA (R&D Systems, Minneapolis, MN, USA), according to the manufacturer’s instructions.
Flow cytometry
Single cells were isolated from PECs as previously described. 3 Briefly, PECs were obtained by lavage with 5 ml HBSS. After washing, PECs were pre-incubated with an FcRγ-blocking mAb (CD16/32, clone 2.4G2; BD Biosciences, San Jose, CA, USA) for 20 min at 4℃ to prevent Ab binding to the FcR. Cells were then stained with various combinations of mAbs. We purchased the following mAbs: anti-CD11b-fluorescein isothiocyanate (FITC) (clone M1/70), anti-NK1.1-phycoerythrin (PE) (clone PK136), anti-MHC class II (clone M5/114.15.2), anti-CD3e-APC-Cy7 (clone 145-2C11), anti-CD4-peridinin chlorophyll protein complex (PerCP) and allophycocyanin (APC) (clone RM4-5), anti-CD8α-PerCP and FITC (clone RM53-6.7),), anti-Gr1-PE (clone RB6-8C5) (all BD Biosciences), anti-CD45R-APC (clone RA3-6B2), anti-F4/80-APC (clone BM8), anti-MHC class II-PerCP-eFluor 710 (clone M5/114.15.2), anti-TCRβ-PE (clone H57-597) (all eBioscience, San Diego, CA, USA), anti-CD3e-PE (clone 145-2C11), anti-TCRγδ-APC (clone GL3), anti-TCRγδ-BrilliantViolet421 (clone GL3), anti-TCRγδ-PE/Cy7 (clone GL3), anti-TCR Vγ1-PE and FITC (clone 2.11), anti-TCR Vγ4-PE and FITC (clone UC3-10A6), and anti-IL-21R-PE (clone 4A9) (all Biolegend). Stained cells were analyzed on a FACSCalibur and a FACSVerse flow cytometer (BD Biosciences). The data were analyzed using CellQuest (BD Biosciences) and FlowJo software (Tree Star, Ashland, OR, USA). γδ T-cell apoptosis was determined by staining with Annexin V–APC conjugate according to the manufacturer’s instructions (eBioscience).
Intracellular cytokine staining
Freshly isolated PECs were re-suspended in RPMI 1640 containing 10% fetal bovine serum (FBS), 100 U/ml penicillin, 100 µl/ml streptomycin and 50 µm 2-ME. PECs were cultured for 24 h in 96-well plates in the presence or absence of rIL-21 (25 or 50 ng/ml; R&D Systems) and further incubated without any stimulation for indicated time at 37℃ and 5% CO2. Brefeldin A (BFA) (Sigma-Aldrich) (10 µg/ml) was added for the last 4 h of culture and the cells washed with FACS buffer and surface stained with various combinations of mAbs for 30 min at 4℃. Intracellular staining was then performed according to the manufacturer’s instructions (BD Biosciences). Briefly, 100 µl Cytofix/Cytoperm solution (BD Biosciences) was added to the cell suspension, mixed and the mixture placed on ice for 20 min at 4℃. Fixed cells were washed twice with 500 µl of Perm/Wash buffer (BD Biosciences) and stained intracellularly with anti-IL-17 A-PE (clone TC11-18H10), anti-IFN-γ-FITC (clone XMG1.2) (both BD Biosciences), anti-IL-17 A-PerCP-Cy5.5 (clone eBio17B7; eBioScience) and anti-Bim-PE (clone C34C5; Cell Signaling, Danvers, MA, USA) for 30 min at 4℃.
Statistics
Statistical significance was evaluated by Student’s t-test using Prism software (GraphPad Software Inc., La Jolla, CA, USA). P < 0.05 was considered statistically significant.
Results
Kinetics of IL-17 A-producing γδ T cells in the peritoneal cavity of il21r−/− mice after BCG inoculation
We examined the kinetics of γδ T cells producing IL-17 A in il21r−/− mice after i.p. inoculation of 1 × 106 CFU BCG as assessed by intracellular cytokine flow cytometory analysis without stimulation. Typical flow cytometory profile and absolute numbers are shown in Figure 1. The number of IL-17 A-producing γδ T cells, especially IL-17A-producing Vγ1−Vγ4− γδ T cells, significantly increased in both groups after BCG infection, with Vγ1−Vγ4− γδ T cells increasing more in il21r−/− mice than in WT mice on d 7 and 14 (Figure 1A–C). These results suggest that inhibition of IL-21R activates IL-17A-producing γδ T cells, primarily Vγ1−Vγ4− γδ T cells at an early stage after BCG infection.
Kinetics of peritoneal γδ T cells in il21r−/− (IL-21R KO) mice after BCG infection. (A) Kinetics of peritoneal γδ T cells and Vγ1−Vγ4− γδ T cells in IL-21R KO mice at the early stage of BCG inoculation. IL-21R KO and age-matched WT (B6) mice were inoculated i.p. with 1 × 106 CFU BCG. PECs were harvested on the indicated days and surface stained with mAbs. (B) Kinetics of peritoneal IL-17A+ γδ T cells or (C) IL-17A+Vγ1−Vγ4− γδ T cells in IL-21R KO mice at the early stage BCG infection. PECs were harvested on the indicated days, cultured with BFA for the last 4 h, stained intracellularly for IL-17A or IFN-γ and analyzed by flow cytometry. Data are shown as mean ± SD (n = 8) from a representative of three separate experiments. Statistically significant differences are shown (*P < 0.05, **P < 0.01, ***P < 0.001).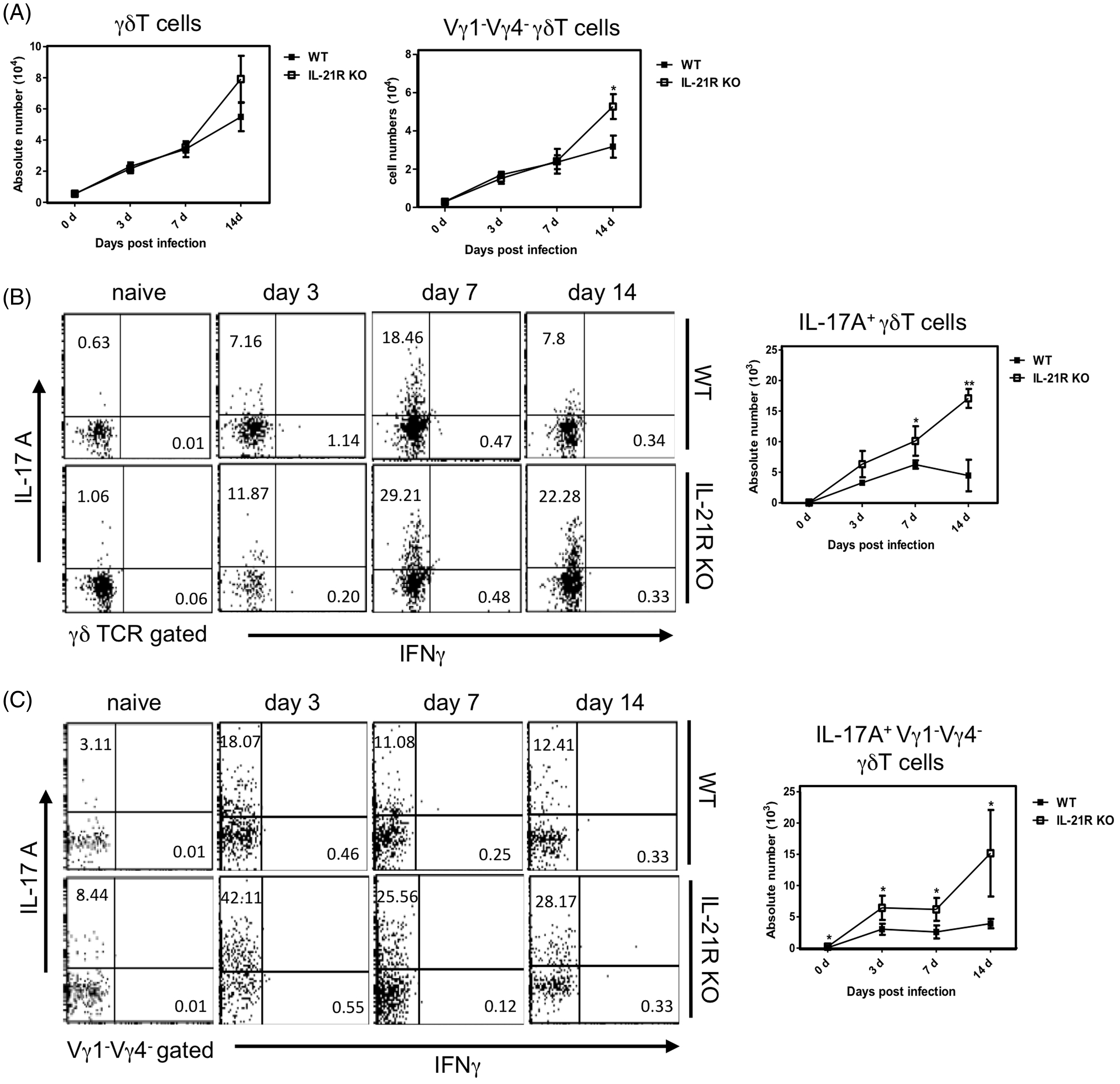
Detection of Ag85B-IL21 fusion protein from rBCG-Ag85B-IL-21
We constructed an rBCG expressing Ag85B-IL-21 fusion protein using the pNN2-mIL-21-Ag85B vector (Figure 2A). Lysates or culture supernatants of rBCG-Ag85B-IL-21, rBCG-Ag85B or BCG were blotted and reacted with anti-IL-21 mAbs to detect Ag85B-IL-21. Protein was detected at ∼47 kDa in the cell lysates and culture supernatant of the rBCG-Ag85B-IL-21 lane but not in the rBCG-Ag85B or the BCG lanes (Figure 2B). Thus, Ag85B-IL-21 was confirmed in rBCG-Ag85B-IL-21. rBCG-Ag85B-IL-21 and rBCG-Ag85B were cultured in 7H9 medium for 6 wk. Notably, IL-21 production peaked during the second wk, as determined from the rBCG-Ag85B-IL-21 culture supernatant (Figure 2C). Together, these data suggest rBCG-Ag85B-IL-21 was successfully constructed.
Construction of the mIL-21 expression plasmid. (A) Construction of pNN2-Ag85B-mIL-21. (B) Bacteria were cultured in 7H9 medium for 1 wk and supernatants harvested after centrifugation. After supernatant and lysate protein quantitation, samples were separated by SDS-PAGE, then blotted and reacted with anti-IL-21 mAb. Arrows indicate Ag85B-IL-21 fusion protein at ∼47 kDa. (C) The concentration of IL-21 in culture supernatants at the indicated days after in vitro culture of rBCG was measured using ELISA kits. The data are representative of two separate experiments and are expressed as the means ± SD of three mice from each group.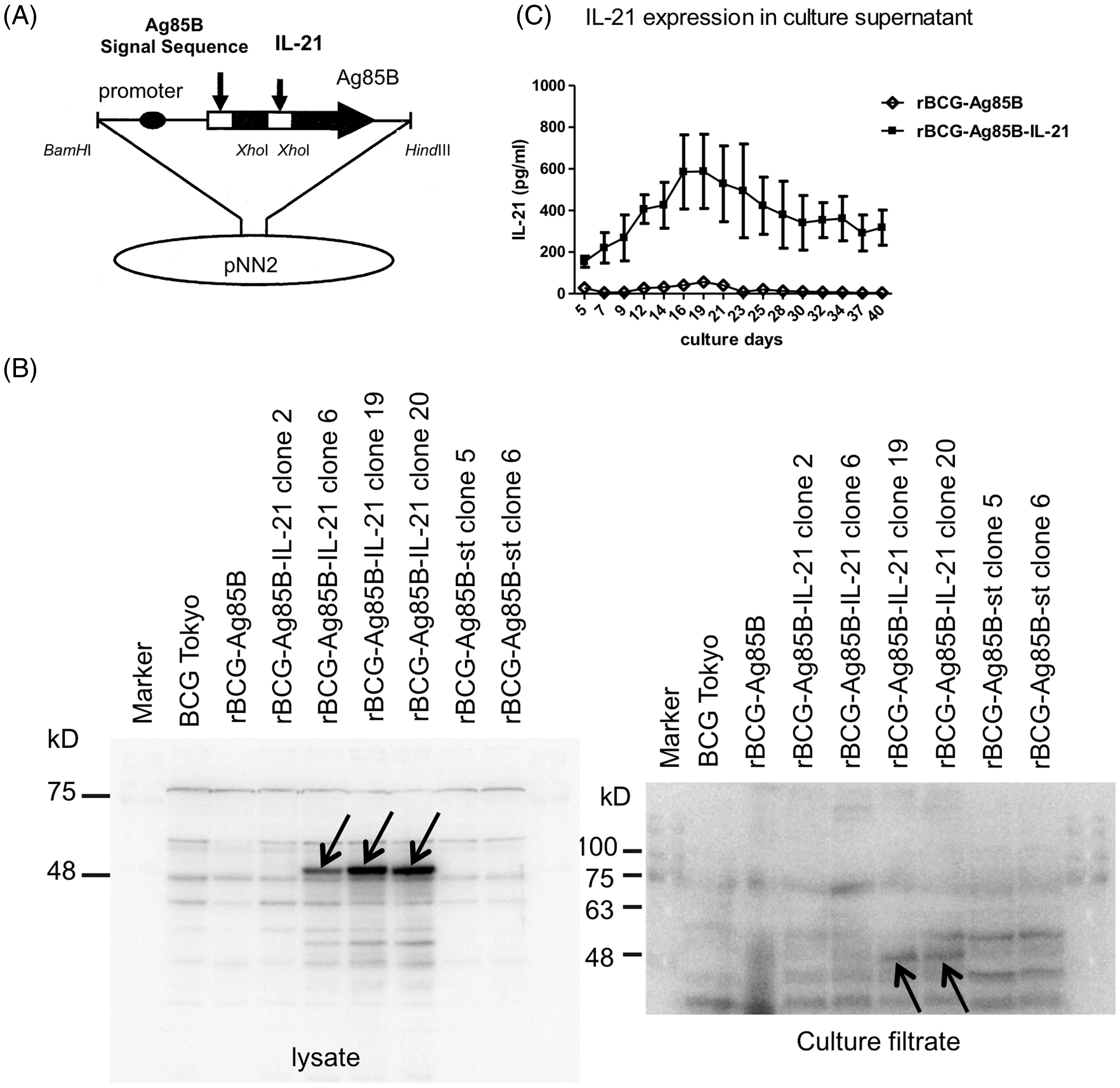
Kinetics of PEC IL-17A-producing γδ T cells in rBCG-Ag85B-IL-21-inoculated mice
We examined the kinetics of bacterial growth and PECs in the peritoneal cavity of WT mice at the early stage after i.p. inoculation with 2 × 106 rBCG-Ag85B-IL21 or rBCG-Ag85B. No obvious differences were observed in the number of bacteria in the peritoneal cavity between rBCG-Ag85B-IL-21- and rBCG-Ag85B-inoculated mice (Figure 3A). However, neutrophil number was significantly higher on d 7 and 14 after rBCG-Ag85B inoculation than after rBCG-Ag85B-IL-21 inoculation (Figure 3B, C; Figure S1). There was no difference in the number of macrophages (CD11b+ Gr-1low F4/80high) between rBCG-Ag85B-IL-21- and rBCG-Ag85B-inoculated mice (Figure 3C).
Kinetics of bacterial growth and PECs in the peritoneal cavity of WT mice after rBCG-Ag85B-IL-21 inoculation. WT mice were inoculated i.p. with 2 × 106 CFU of rBCG-Ag85B-IL-21 or rBCG-Ag85B. (A) The number of bacteria recovered from the peritoneal cavity of inoculated mice was determined on the indicated days. (B) Flow cytometry analysis of peritoneal neutrophil (CD11b+ Gr-1high F4/80low) kinetics and total numbers from inoculated mice. PECs were harvested on the indicated days after rBCG-Ag85B-IL21 or rBCG-Ag85B inoculation and surface stained with various mAbs. (C) Kinetics of peritoneal neutrophils shortly after rBCG-Ag85B-IL21 or rBCG-Ag85B inoculation. Neutrophil (CD11b+ Gr-1high F4/80low) number was calculated by multiplying the total PEC number by the percentages of each PEC subset. Data are represented as the mean ± SD (n = 8) from a representative of three separate experiments. Statistically significant differences are shown (*P < 0.05, **P < 0.01, ***P < 0.001). Kinetics of peritoneal γδ T cells at an early stage of rBCG-Ag85B-IL21 or rBCG-Ag85B inoculation. PECs were harvested on the indicated days and surface stained with mAbs. Kinetics of peritoneal γδ T cells (A) or peritoneal Vγ1−Vγ4− γδ T cells (C) at the early stage rBCG-Ag85B-IL21 or rBCG-Ag85B inoculation. The absolute number was calculated by multiplying the total PEC number by the percentages of each PEC subset. (B) Data are represented as the mean ± SD (n = 8) from a representative of three separate experiments. Statistically significant differences are shown (*P < 0.05, **P < 0.01, ***P < 0.001).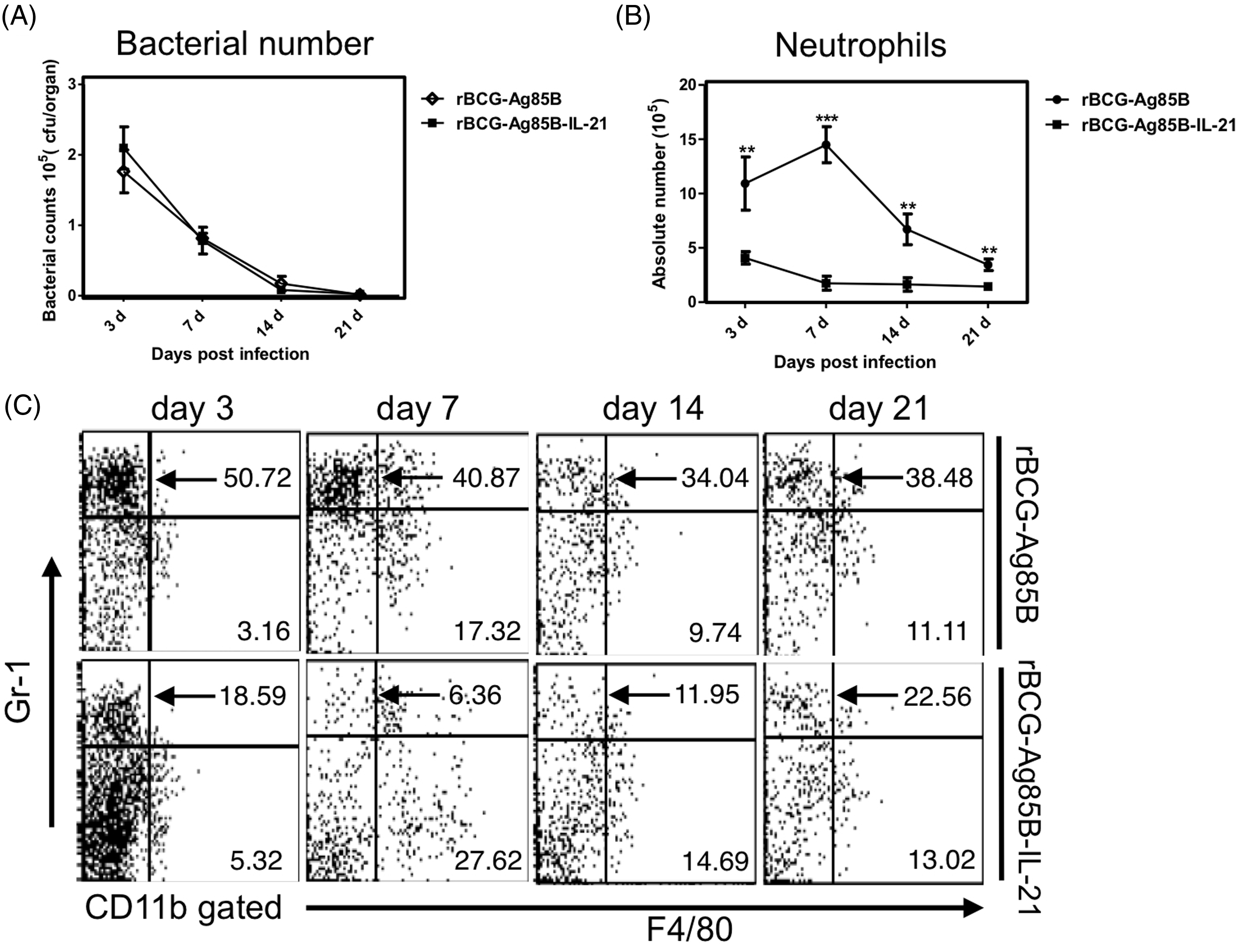
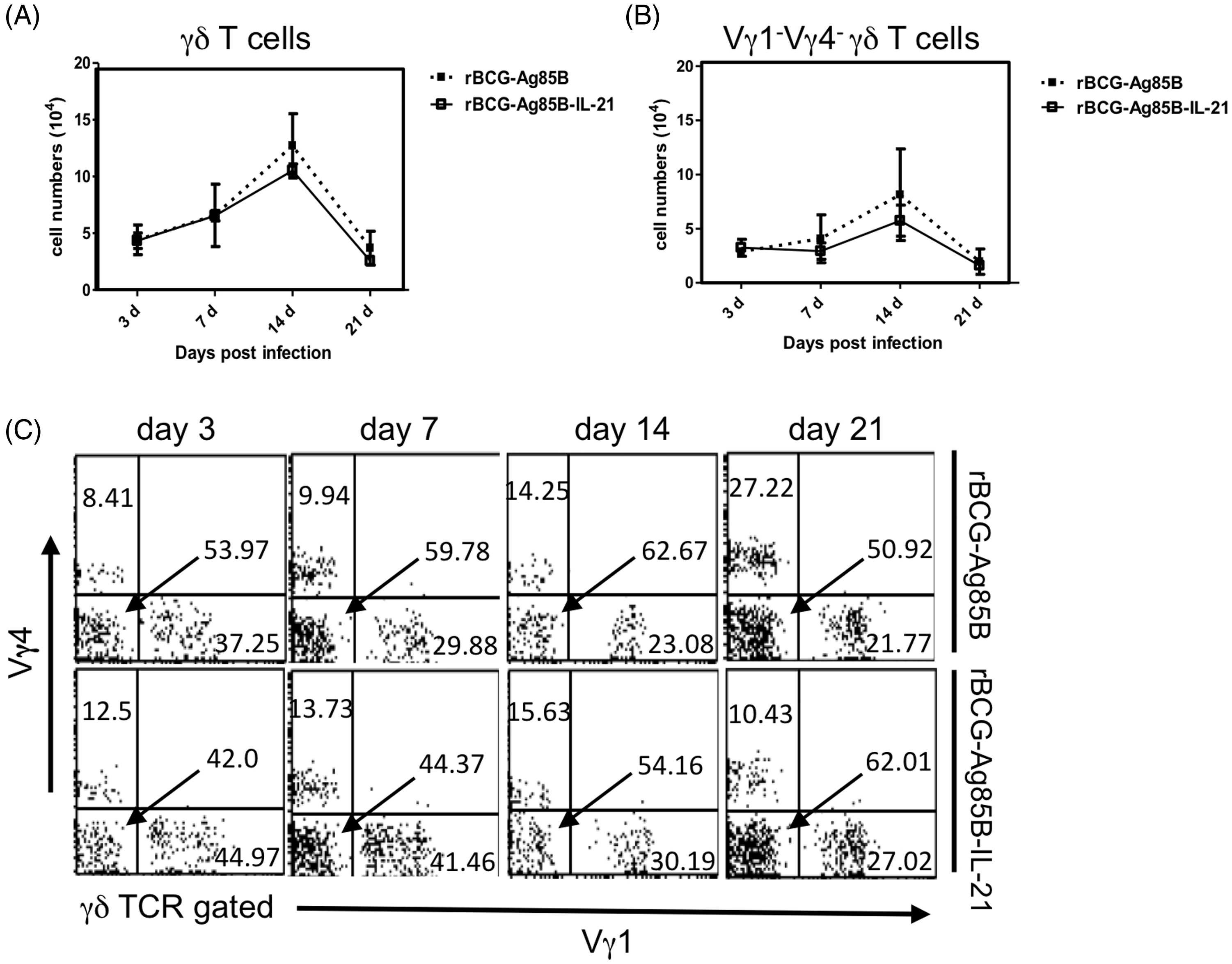
We next examined the kinetics of innate IL-17A-producing γδ T cells after rBCG-Ag85B inoculation, by intracellular cytokine flow cytometory analysis without stimulation. There were also no significant differences in the absolute number of γδ T cells and Vγ1−Vγ4− γδ T cells between rBCG-Ag85B-IL-21- and rBCG-Ag85B-inoculated mice on d 3, 7 and 14 (Figure 4A–C; Figure S2). Innate γδ T cells preferentially appear at an early stage after BCG inoculation, detectable by intracellular cytokine flow cytometry analysis without ex vivo stimulation.
27
We have previously reported that the Vγ1−Vγ4− subset expressing Vγ6/Vδ1 mRNA is the major subset of IL-17A-producing cells in the peritoneal cavity after BCG infection.
27
Consistent with this, the numbers of IL-17A-producing γδ T cells, especially IL-17A-producing Vγ1−Vγ4− γδ T cells, significantly increased after rBCG-Ag85B inoculation. However, the number of IL-17A-producing Vγ1−Vγ4− γδ T cells was significantly lower on d 7 and 14 after rBCG-Ag85B-IL-21inoculation (Figure 5A, B). These results suggest that overproduction of IL-21 may down-regulate activation of IL-17A-producing Vγ1−Vγ4− γδ T cells in mice after BCG infection.
Kinetics of IL-17A+ γδ T cells shortly after rBCG-Ag85B-IL-21 or rBCG-Ag85B inoculation. PECs were harvested on the indicated days, cultured with BFA for the last 4 h, stained intracellularly for IL-17A or IFN-γ and analyzed by flow cytometry. Kinetics of IL-17A+ γδ T cells (A) and IL-17A+ Vγ1− Vγ4− γδ T cells (B) in PECs during early stage rBCG-Ag85B-IL-21 or rBCG-Ag85B inoculation. Data are represented as the mean ± SD (n = 8) from a representative of three separate experiments. Statistically significant differences are shown (*P < 0.05, **P < 0.01, ***P < 0.001).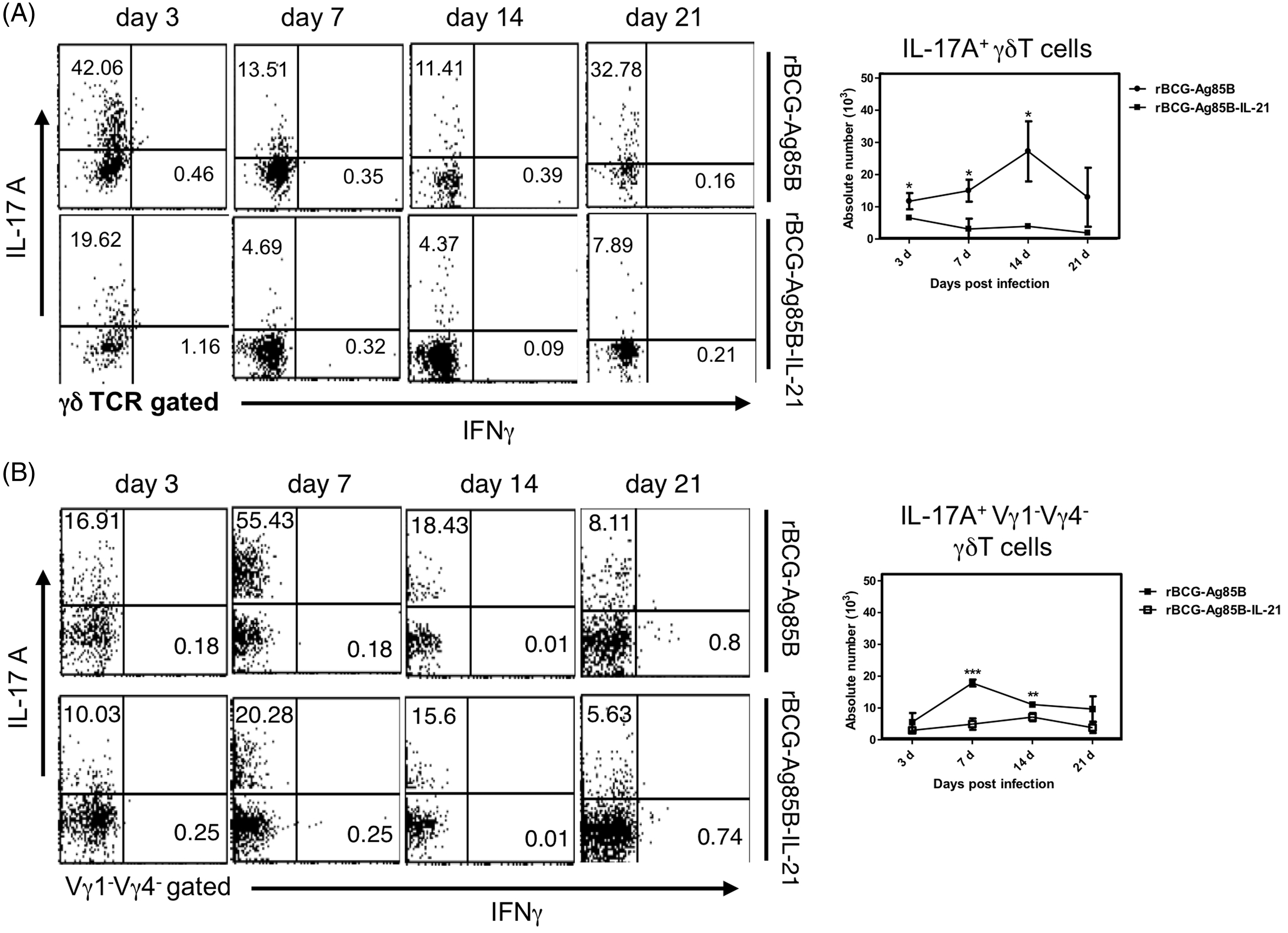
Effect of IL-21 on apoptosis of IL-17A producing γδ T cells in vitro
To further examine how IL-21 suppresses the response of IL-17A-producing γδ T cells, we examined the effect of rIL-21 on IL-17A-producing γδ T cells in vitro from mice on d 7 post BCG infection. We confirmed stimulation of IL-21R on Vγ1−Vγ4− γδ T cells, on d 7 post-BCG infection (1 × 106 CFU) (Figure 6A). Addition of rIL-21 to in vitro γδ T cells for 24 h reduced the frequency of IL-17A-producing γδ T cells (Figure 6B) or Vγ1−Vγ4− γδ T cells (Figure 6C).
Effect of exogenous IL-21 on IL-17A+ γδ T cells in vitro. (A) Expression of IL-21R on γδ T cells. PECs were harvested from mice post-7 d BCG inoculation and stained with anti-IL-21R mAb and anti-Cδ mAb. (B) Effect of IL-21 on IL-17A+ or IFN-γ+ γδ T cells. PECs were harvested from WT mice on d 7 after inoculation i.p. with 1 × 106 CFU of BCG, cultured in 24-well plates for 24 h in the presence or absence of IL-21 (25 or 50 ng/ml), and further incubated without any stimulation for 6 h at 37℃ and 5% CO2. BFA (10 µg/ml) was added for the last 4 h and the cells washed with FACS buffer and surface stained with various combinations of mAbs for 30 min at 4℃. The percentage of intracellular cytokine-producing cells was assessed by flow cytometry after gating on γδ T cells. (C) Effect of IL-21 on Vγ repertoire of γδ T cells. PECs were cultured in 24-well plates for 24 h in the presence or absence of IL-21 (25 or 50 ng/ml), and surface stained with mAbs.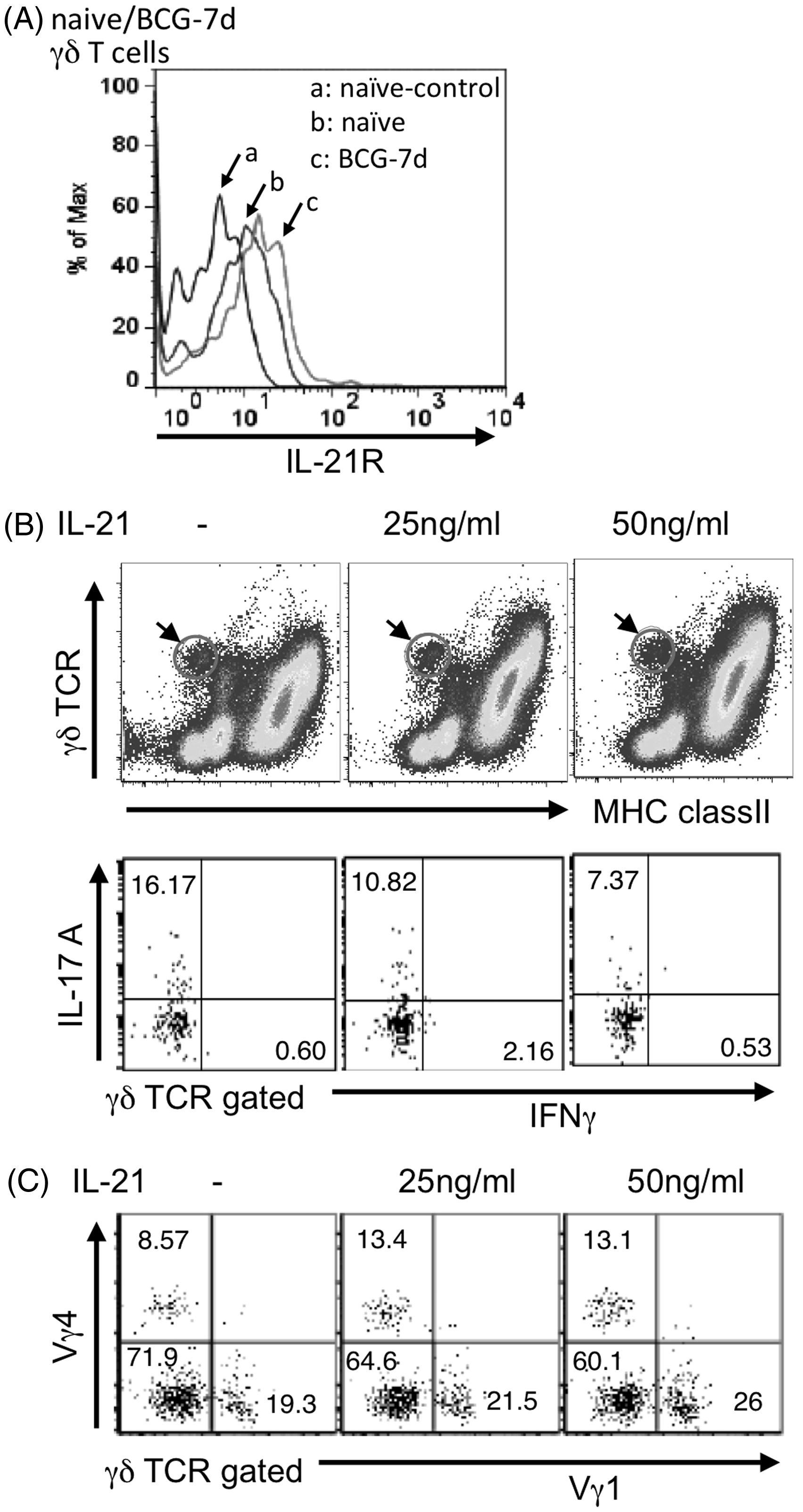
IL-21 induces apoptosis of some lymphoid and myeloid cells.
28
To further examine whether IL-17A-producing cells underwent apoptosis upon IL-21 stimulation, we used IL-17A-reporter (il17g/g) mice, which had been inoculated with BCG 7 d earlier. The frequency of GFP+ γδ T cells undergoing apoptosis (Annexin V+) increased in a dose-dependent manner (Figure 7A). The expression of Bim, a member of the pro-apoptotic Bcl2 family, was up-regulated by addition of rIL-21 (Figure 7B). Taken together, the data indicated that IL-21 selectively induces apoptosis of IL-17A-producing Vγ1−Vγ4− γδ T cells via Bim expression.
Effect of exogenous IL-21 on apoptosis of IL-17A+ γδ T cells in vitro. (A) Apoptosis of IL-17A+ γδ T cells. (B) Bim expression in IL-17A+ γδ T cells. PECs were harvested from IL-17A-reporter (il17g/g) mice on d 7 after inoculation i.p. with 1 × 106 CFU of BCG, cultured in 24-well plates for 6 h (B) and 12 h (A) in the presence or absence of IL-21 (25 or 50 ng/ml), then intracellularly stained with mAbs for apoptosis. Solid gray line represents isotype control (representative of two independent experiments).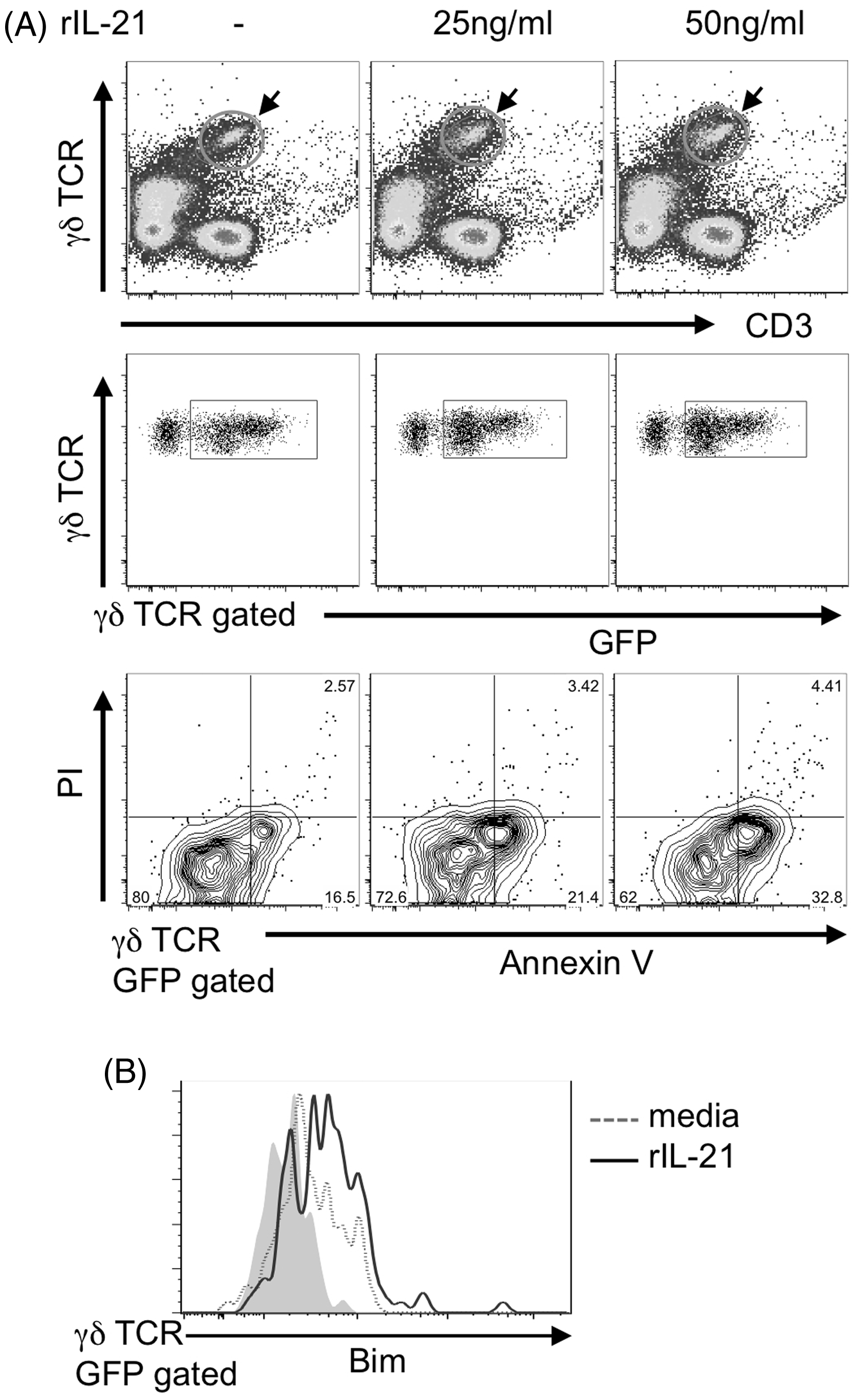
Discussion
We have shown that the IL-21/IL-21R plays an important role in regulating innate IL-17A-producing γδ T cells after BCG infection. The role of IL-21 in regulating IL-17A-producing T cells is controversial. Several lines of evidence suggest IL-21 plays a critical role in the expansion of IL-17A-producing αβ T (Th17) cells, although it is not required for the differentiation of Th17 cells from naive CD4+ T cells.29,30 In contrast, Ertelt et al. reported that IL-21 suppressed IL-17A production by Listeria Ag-specific CD4+ T cells during in vivo infection with L. monocytogenes. 31 Do et al. reported that IL-21 was required for Lymph node γδ T cell IL-17A expression. 23 In addition, WT and il21−/− mice exhibited LN γδ T cell IL-17A expression in response to phorbol 12-myristate 13-acetate and ionomycin. In contrast, Moser et al. reported that IL-21R signaling suppressed lung IL-17A-expressing γδ T cells following influenza A virus infection. 24 Consistent with this, we found that the number of IL-17A-producing Vγ1−Vγ4− γδ T cells decreased significantly after inoculation with rBCG-Ag85B-IL-21, while IL-17A-producing Vγ1−Vγ4− γδ T cells increased in il21r−/− mice after BCG infection. Exogenous IL-21 selectively induced Bim expression and reduced IL-17A-producing Vγ1−Vγ4− γδ T cells by apoptosis. Thus, IL-21/IL-21R may act as a potent inhibitor of the IL-17A-producing γδ T cell subset during BCG infection.
The mechanism whereby IL-21R signaling induces apoptosis of IL-17A-producing γδ T cells remains to be determined. IL-21R is composed of two subunits, IL-21R and Cγ. 19 IL-21R activates Jak1 and Jak2, causing phosphorylation, dimerization and nuclear translocation of STAT3, and, to a lesser extent, STAT1, STAT5a and STAT5b.19–22 IL-21 induces B-cell apoptosis through a Bim- and caspase-dependent pathway, although it stimulates B cells in the context of a co-stimulatory T-cell signal.17,32,33 Similarly, IL-21 induces NK cell apoptosis, although it co-stimulates cytokine-activated NK cells.34,35 Recently, it has been reported that IL-21 induces apoptosis of conventional dendritic cells via a STAT3 and Bim-dependent mechanism. 36 Consistent with these reports, our present study suggests that IL-21 reduces IL-17A-producing γδ T cells by apoptosis via Bim expression. An important finding in the present study is that only Vγ1−Vγ4− γδ T cells were affected by IL-21. We previously reported that the Vγ1−Vγ4− subset expressing Vγ6/Vδ1 mRNA in the peritoneal cavity produced IL-17A after BCG infection without ex vivo stimulation. 27 An important finding in the present study is that only Vγ1−Vγ4− γδ T and Vγ6 γδ T cells were affected by IL-21. The Vγ6/Vδ1 γδ T cells bear invariant TCRs, even to the nucleotides in the TCR gene junction.37,38 The canonical sequence is very simple, with no apparent N-region contribution. This characteristic has led to the hypothesis that this subset of γδ T cells are pre-programmed to recognize a limited set of stress or environmentally-induced self-Ags derived from commensal microflora.1,2 We have previously reported that IL-17A-producing Vγ6 γδ T cells selectively express CD30L/CD30, which are induced by TCR signaling,11,27 supporting this hypothesis. At present, the specificity of these γδ T cells remains unknown. However, Vγ1−Vγ4− γδ T cells bearing Vγ6 may be stimulated by TCR upon recognizing an unknown Ag in the medium or auto-Ags expressed by neighboring γδ T cells. In chronic viral infection, Ag-specific CD8+T cells become exhausted and lose function, along with increased expression of PD1. IL-21 restores the effector function of exhausted CD8+T cells, promoting viral clearance and resulting in immunopathology.39–41 IL-21 signaling induces Blimp-1, a transcriptional factor, which mutually regulates T-cell BCl6 expression.33,42,43 In the case of αβ T cells, terminally differentiated effector T cells express a high level of Blimp-1 but a low level of Bcl6.44,45 Thus, we speculate that IL-21 signaling plays an important role in terminating the IL-17A-producing γδ T-cell response by promoting activation-induced cell death. Further studies are needed to clarify these possibilities.
γδ T cells are innate sources of IL-17A, closely linked to the recruitment of neutrophils after bacterial infection, including E. coli, 12 C. albicans 13 and BCG6–8 infections. Importantly, the neutrophil-dependent inflammatory response decreased after rBCG-Ag85B-IL-21 inoculation. Such a regulatory role for IL-21 in the neutrophil-dependent inflammatory response may be important in the shift of innate to adaptive immunity. It is also useful for controlling inflammatory diseases caused by IL-17A-producing γδ T cells.
Conclusion
We have demonstrated that the number of IL-17A-producing γδ T cells, particularly Vγ1−Vγ4−γδ T cells, significantly decreased after inoculation with rBCG-Ag85B-IL-21, while IL-17A-producing Vγ1−Vγ4−γδ T cells increased in il21r−/− mice after BCG infection. IL-21 selectively stimulated Bim expression and induced apoptosis in IL-17A-producing Vγ1−Vγ4−γδ T cells. IL-21/IL-21R may act as a potent inhibitor of IL-17A-producing γδ T cells, controlling neutrophil-dependent inflammatory responses mediated by IL-17A-producing γδ T cells.
Footnotes
Acknowledgements
We thank Akiko Yano, Miki Kijima and Yumiko Kitada for their secretarial assistance and all the members of our laboratory for helpful discussion.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Grants-in-Aid (25670213; 26293098) from the Japan Society for the Promotion of Science, Yakult Bioscience Foundation (Y.Y.), and the Takeda Science Foundation (Y.Y.).