
Abstract
Background
Consider this Carrion's disease (CD) is a biphasic illness—comprising acute and chronic phases—endemic to Peru and caused by Bartonella bacilliformis, a bacterium transmitted by sandflies. Despite its clinical relevance, the mechanisms underlying innate immune activation in response to B. bacilliformis remain poorly understood. Toll-like receptors (TLRs) play a central role in recognizing conserved molecular patterns present in pathogens, thereby initiating innate immune responses. The present study aimed to describe the expression patterns of TLR2 and TLR4, along with cytokine secretion profiles, during peripheral blood mononuclear cells (PBMCs) exposure to B. bacilliformis.
Methods
Peripheral blood mononuclear cells (PBMCs) from healthy donors were stimulated with heat-inactivated B. bacilliformis (ATCC 35685 strain), zymosan (TLR2 control), or E. coli LPS (TLR4 control). TLR2 and TLR4 gene expression was quantified by RT-qPCR at 0, 12, 24, 36, and 48 h. Cytokines were measured using a 17-plex panel. Analyses were descriptive, using non-parametric statistics.
Results
Results elicited measurable changes in the transcriptional expression of TLR2 and TLR4 during stimulation, with peak activation typically observed at 12 h, although in one case the maximal response was delayed to 24 h. These expression changes coincided with significant modulation of multiple cytokines, including pro-inflammatory mediators (TNF-α, IL-17, IL-12p70), Th1/Th2 cytokines (IFN-γ, IL-2, IL-5, IL-13), regulatory cytokines (IL-10), and growth factors (GM-CSF, IL-7).
Conclusions
This exploratory study describes the transcriptional expression patterns of TLR2 and TLR4 and the accompanying cytokine responses in PBMCs exposed to B. bacilliformis. These profiles expand current knowledge of the early innate immune signature elicited by this neglected pathogen and provide a foundation for future studies using receptor-specific functional assays. Importantly, these patterns likely reflect early innate immune response signatures rather than definitive evidence of functional receptor activation.
Introduction
Carriońs diseases (CD), ORPHANET 64692, is a neglected poorest-linked illness, endemic in areas of Peru, Colombia and Ecuador, primarily affecting populations in Andean valleys between 500 and 3200 masl. 1 Although, recently, new cases emerged in non-endemic areas. 2 B. bacilliformis is the causal agent of illness. It is a Gram-negative bacillus, pleomorphic, aerobic, intracellular, flagellate and non-fermentative.3,4 It is transmitted mainly by sand flies (Lutzomyia spp.) and clinically presents two phases: the Acute phase (most lethal) and the Chronic phase (called Peruvian wart as well). In addition, a high number of asymptomatic carriers has been reported. 3
Current knowledge of the innate immune response to B. bacilliformis is very limited, mainly due to the neglected nature of CD and the difficulty in accessing samples from endemic areas. 5 In a previous study evaluating cytokine responses after in vitro stimulation, it was described a pro-inflammatory profile in the early hours, high levels of TNF-α, IL-1α, IL-1β, IFN-α2, and IL-6, followed by an anti-inflammatory response led by IL-1RA. 6 Moreover, some aspects of the adaptive immune response have been previously described, such as elevated levels of IL-10 and abnormal counts of CD4⁺ and CD8⁺ T lymphocytes during the acute phase, increased levels of IFN-γ and IL-4 have also been reported in the chronic phase. 7 More recently, IL-10 overproduction has been associated with immunosuppression and may contribute to immune evasion by the bacterium. 8 However, data on pattern-recognition pathways, including TLR-associated transcriptional responses, remain scarce.
TLRs are proteins expressed in immune cells, such as macrophages and dendritic cells, as well as in non-immune cells, including fibroblast cells and epithelial cells. 9 These proteins recognize Pathogen-Associated Molecular Patterns (PAMPs) and Damage-Associated Molecular Patterns (DAMPs). There are two types of TLRs, the Cell surface TLRs that mainly recognize microbial membrane components such as lipids, lipoproteins, and proteins such as TLR1, TLR2, TLR4, TLR5 and TLR11, and intracellular TLRs that recognize nucleic acids derived from bacteria and viruses and also recognize self-nucleic acids in disease conditions such as TLR3, TLR7, TLR8 and TLR9. 10 From the few existing reports about B. bacilliformis, it has been found to evade TLR5, an essential receptor for recognizing flagellated bacteria. This evasion is associated with mutations in flagellin, that has different amino acid sequences in the recognition site. 11 In previous studies, B. henselae and B. quintana were associated with TLR2 and cytokine induction12,13; notwithstanding, the TLR associated with B. bacilliformis is currently unknown.
Given the increasing incidence of CD in recent years, understanding its innate immune signature is increasingly relevant. 14 This exploratory study aims to investigate the expression of TLR2/TLR4 and associated cytokines in peripheral blood mononuclear cells (PBMCs) stimulated in vitro with B. bacilliformis, over time.
Materials and methods
Culture, inactivation and quantification of B. bacilliformis
The ATCC 35685 B. bacilliformis strain was grown on 5% Sheep Blood agar (HiMedia, Mumbai, India) at 28°C under microaerophilic conditions. The presumptive colonies of B. bacilliformis were identified by Polymerase Chain Reaction (PCR) using the specific primers to detect the flagellin-encoding fla gene (F: 5’-ATAGAAAGAGCCTGAATACC-3’ and R: 5’-TGATGAAGCATGACAGTAACAC-3’). 15 Subsequently, B. bacilliformis cultures were resuspended in PBS and heat-treated for 60 min at 56 °C to prevent potential alterations in the results of subsequent assays, attributable to factors related to bacterial replication, metabolic changes, and thermolabile elements (such as enzymes or variations in antigenic proteins). 13 The heat-inactivated (HI) bacteria were stored at −80°C until the stimulation assay. The bacterial quantification was done by flow cytometry as described previously, 6 resulting in approximately 11.42 × 106 cells/μl, which were standardized per well until 1 × 106/ml.
Blood extraction, isolation and stimulation
Thirty millilitres of peripheral blood were collected by venipuncture in EDTA tubes from five volunteers. Peripheral Blood Mononuclear Cell (PBMCs) isolation was performed using the density gradient method with Histopaque (Sigma-Aldrich, Massachusetts, US). 16 Cryovials were initially placed in a controlled-rate freezing container at −80 °C to ensure gradual cooling and subsequently transferred to liquid nitrogen until the use in stimulation assays. In 96-well round-bottom trays, RPMI 1640 culture medium (Merck KGaA) supplemented with 5% heat-inactivated fetal calf serum (Gibco BRL, Auckland, New Zealand), Minimal Essential Medium 1X (Gibco BRL, Auckland, New Zealand), sodium pyruvate 20 mM and 100 mg/ml penicillin/streptomycin were stimulated PBMCs (1 × 106/ml) with HI B. bacilliformis (1 × 106/ml), Escherichia coli lipopolysaccharide O111:B4 (100 ng/ml, Merck KGaA) as a TLR4 positive control, 17 or zymosan (10 µg/ml, Cayman chemical) as a TLR2 positive control, 18 while non-stimulated PBMCs were cultivated in culture medium as negative control at 37°C in 5% CO2. PBMCs and the supernatant were collected at 0, 12, 24, 36, and 48 h after incubation.
RNA extraction and Rt-qPCR to detect TLRs
RNA was extracted using TriReagent (Molecular research center, US) following the manufacturer's instructions with some modifications as indicated below: the plate was centrifuged at 3000 R/min for 5 min at 4°C, then 500 µl of the sample from each volume of the well was transferred to a 1.5 ml tube where TriReagent was added in a proportion of 1:3, respectively, it was incubated for five minutes and 0.2 ml of isopropanol were added for every 1 ml of TriReagent used and centrifuged for 10 min at 13,000 R/min. After centrifugation, the supernatant was transferred with an equivalent volume of isopropanol and incubated for 15 min, followed by centrifugation at 13,000 R/min for 10 min at 4°C. Subsequently, the supernatant was decanted, washed with 75% ethanol, and centrifuged at 7500 g for 5 mins. After washing, the open tubes were air-dried for 15 min to evaporate residual ethanol and eluted in 25 µl of RNase-free water. Finally, thermal treatment was applied for 10 min at 60°C. The RNA quality was determined by 1% Agarose gel, and the OD260/280 ratio was measured by spectrophotometry (BioTek Epoch Microplate Spectrophotometer, Agilent). The qMAX™ First Strand cDNA Synthesis Flex Kit (Accuris, New Jersey, US) was utilized for reverse transcription according to the manufacturer's protocol. Gene expression analysis was assessed via qPCR using the primers described 18 to detect TLR2 (F: 5’-TTG TGA CCG CAA TGG TAT CTG-3’, R: 5’-GCC CTG AGG GAA TGG AGT TT-3’), TLR4 (F: 5’-GGC CAT TGC TGC CAA CAT-3’, R: 5’-CAA CAA TCA CCT TTC GGC TTT T-3’), and the housekeeping gene β-Actin (F: 5’-CCT GGC ACC CAG CAC AAT-3’, R: 5’-GCC GAT CCA CAC GGA GTA CT-3’). BlasTaq™ 2X qPCR MasterMix with SYBR Green (Applied Biological Materials, British Columbia, Canada) was used under the following qPCR conditions: 10 min at 95°C, followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. Also, the Melting curve analysis was performed using the following protocol: 95°C for 15 s, 60°C for 60 s and 95°C for 15 s. Each run was evaluated by duplicate. Relative gene expression levels were calculated using the 2−ΔΔCt method. 19
Cytokines analysis
After stimulation of PBMCs, the 96-well plate was centrifuged at 2000 R/min for 5 min, and the supernatant was collected and stored at −80°C. The cytokines analysis was performed with a commercial human cytokines panel (Bio-Plex Pro Human Cytokine 17-plex, BIORAD) using 30 µl of supernatant following the manufacturer's instructions to evaluate 17 biomarkers of cellular signaling of cytokines and chemokines including: granulocyte colony-stimulating factor (G-CSF), granulocyte macrophage colony-stimulating factor (GM-CSF), interferon γ (IFN-γ), interleukins IL-1β, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL- 12 (p70), IL-13, IL-17A, monocyte chemoattractant protein-1 (MCP-1/MCAF), macrophage inflammatory protein MIP-1β and tumor necrosis factor (TNF-α). Unstimulated PBMC were used as a control for every time evaluated. Samples were read by the MAGPIX instrument and analyzed with xPONENT software 3.1.
Data analysis
An exploratory study was conducted to examine gene expression levels of TLR2 and TLR4 at multiple times points 0, 12, 24, 36, and 48-h post-stimulation with HI B. bacilliformis A non-parametric paired Wilcoxon signed-rank was employed to compare each post-infection time point against the baseline control, within the same individual and was visualised using box plots. The p-values were considered statistically significant when ≤ 0.1
For cytokine analysis, a logarithmic transformation (log₁₀) was applied to cytokine concentrations to improve normality and reduce heterogeneity of variance; later, comparisons between experimental groups (control vs. HI B. bacilliformis) were conducted independently for each time point using the non-parametric Mann-Whitney U test. To explore potential relationships among TLRs and cytokine responses, the Spearman test was chosen. The p-values were considered statistically significant when ≤ 0.05. All statistical analyses and visualisations were performed using R (version 4.3.1).
Results
Characteristics of volunteers
The study included five participants, 80% male and 20% female, with a mean age of 27.3 ± 7.5 years. None reported allergies, immunological disorders, or a history of Bartonellosis. Only two participants (40%) had traveled to endemic areas in the past five years.
TLR expression
PBMC exposure to heat-inactivated B. bacilliformis led to distinct temporal changes in TLR2 and TLR4 transcription, both TLRs exhibited an increase at 12 h, suggesting an early transcriptional response. TLR2 expression peaked at 12 h, with transcriptional levels 2.73-fold higher than the control (p = 0.059). However, at 24 h, one individual exhibited a markedly higher peak, with expression levels 10.78-fold greater than the control (Figure 1). TLR4 expression followed a similar kinetic profile to TLR2, peaking at 12 h in four individuals, with a 2.36-fold increase relative to the control (p = 0.059) and, later, was downregulated 1.04-fold compared to the control (p = 0.059). One individual, however, displayed a peak at 12 h with a 13.27-fold increase in transcriptional expression (Figure 1).
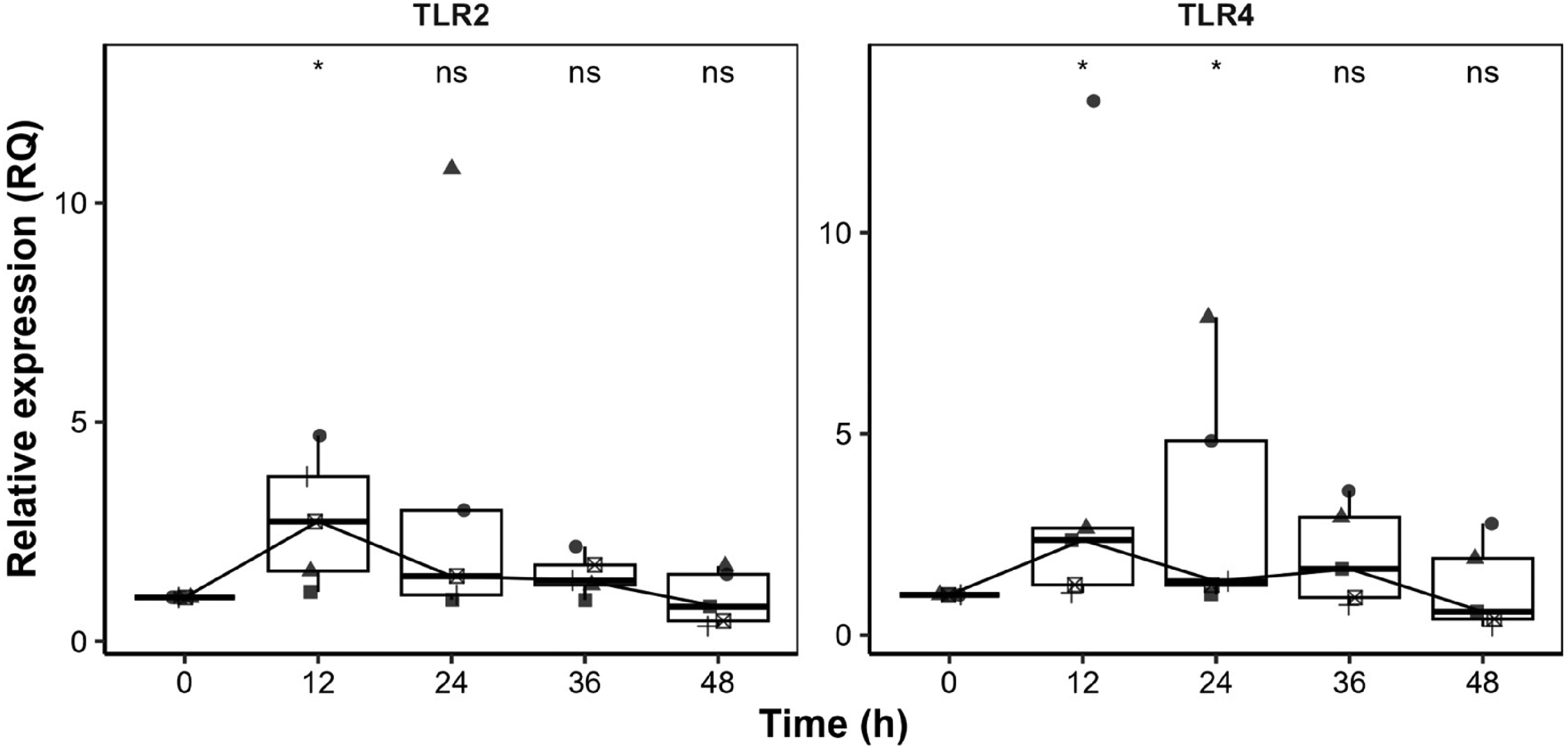
Kinetics of TLR2 and TLR4 expression in PBMCs stimulated with heat-inactivated Bartonella bacilliformis. Relative expression (RQ) over 0, 12, 24, 36, and 48 h was shown. Boxplots represent the median (horizontal line), interquartile range (Q1–Q3, boxes), and overall data range (whiskers), with individual data points overlaid using a jitter plot to illustrate sample-level variability. (*) indicate statistically significant differences between B. bacilliformis-stimulated and unstimulated controls (p < 0.1); ns, not significant.
Concerning the transcriptional expression of the TLR positive controls, TLR2/Zymosan peaked at 36 h, reaching 2.73-fold higher levels than the control, except in one case where expression peaked at 24 h (8.11-fold). Similarly, TLR4/LPS expression peaked at 24 h, showing a 3.48-fold increase (p = 0.059), except for one individual who exhibited markedly higher expression compared to the control, with a 10.3-fold increase at the same time point (Figure 2).
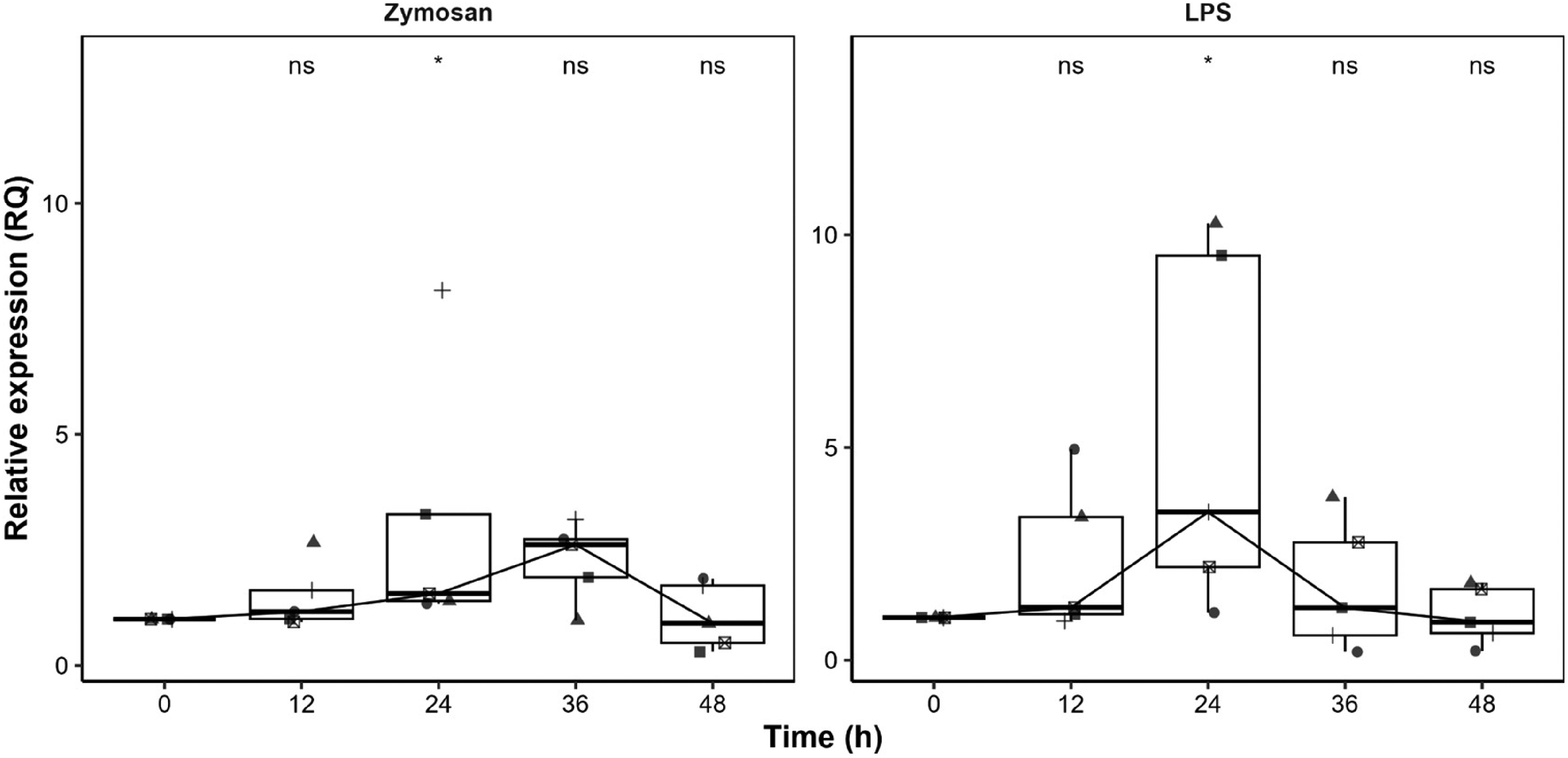
Relative expression of positive controls in PBMCs: zymosan for TLR2 and LPS for TLR4. Boxplots represent the median (horizontal line), interquartile range (Q1–Q3, boxes), and data range (whiskers), with individual data points overlaid to illustrate variability. (*) indicate significant differences compared with unstimulated controls (p < 0.1 ; ns, not significant).
Quantification of cytokines
Cytokine measurements revealed time-dependent modulation following B. bacilliformis stimulation. Early elevations included GM-CSF GM-CSF, IL-12p70, IL-13, IL-2, IL-5 and IL-7, but only GM-CSF, IL-13 and IL-7. Later, at 36 h, cytokines IFNγ and IL-10 were produced and finally at 48 h, TNF-α and IL-17 were produced. Specifically, GM-CSF showed a sustained increase starting at 12 h (p = 0.031), with consistently significant elevations at 24, 36, and 48 h (p = 0.008). IFN-γ expression became significant at 36 h and remained elevated at 48 h (p = 0.008). IL-10 increased significantly to 36 h (p = 0.016) and 48 h (p = 0.036). IL-12p70 levels were significantly elevated at 12 h (p = 0.016) and 48 h (p = 0.012). IL-13 showed progressive upregulation, with significant increases at 12 h (p = 0.012), 24 h (p = 0.036), 36 h (p = 0.021), and 48 h (p = 0.021). IL-17 was significantly upregulated at 48 h (p = 0.032). IL-2 exhibited significant increases at 12 h (p = 0.008), 36 h and 48 h (p = 0.016 each). IL-5 was significantly elevated at 12 h (p = 0.008) and 48 h (p = 0.032). IL-7 showed a progressive and consistent increase from 12 h (p = 0.034) to 48 h (p = 0.011). TNF-α became significantly upregulated at 48 h (p = 0.032) (Figure 3). Additionally, no differences in G-CSF, IL-1β, MCP-1, IL-6 and IL-8 concentrations have been reported.
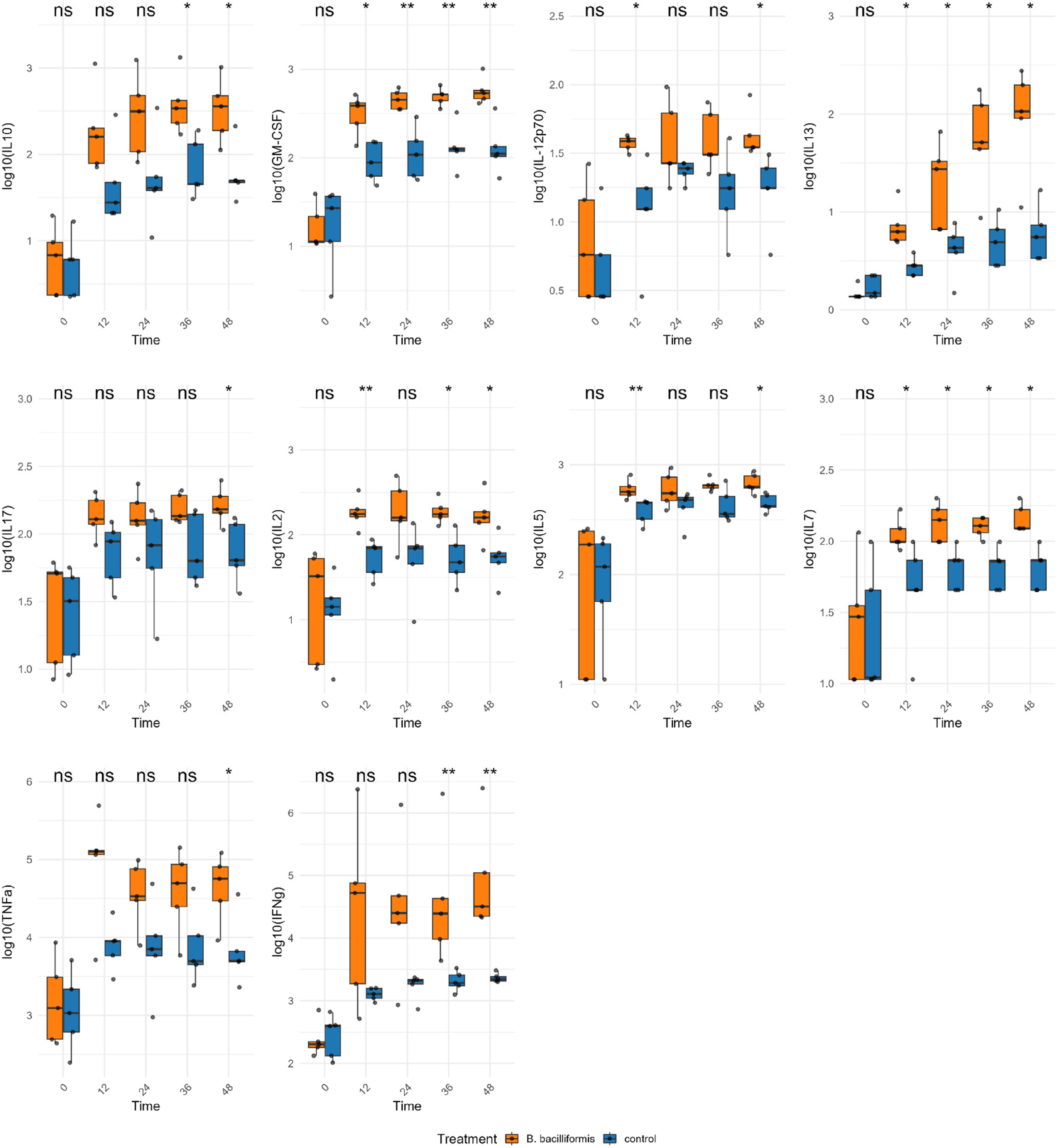
Cytokine expression of PBMC stimulated with HI B. bacilliformis (orange boxplots) and unstimulated (blue boxplots), both Q1 and Q3 interquartile range at different times. Statistical analysis was performed using the Mann-Whitney U test. * : p < 0.05, ** : p < 0.01; ns: not significant.
The correlation analysis between cytokine concentrations and TLR expression
The correlation analysis between cytokine concentrations and TLR expression (Figure 4) revealed distinct temporal patterns. At 0 h, correlations were low, reflecting baseline expression. At 12 h, when median TLR2 and TLR4 levels peaked, most cytokines showed weak or negative associations. Notably, TLR4 correlated positively with IL-12p40 (p = 0.35) and strongly with IL-8 (p = 0.80), suggesting early proinflammatory activation. By 24 to 48 h, distinct immune polarization signatures emerged, thus TLR2 expression correlated positively with Th2 and regulatory cytokines, including IL-10, IL-13, and IL-5, while TLR4 showed positive correlations with Th1-associated cytokines, such as IL-12p70, IFN-γ, and TNF-α. At 48 h, IL-7 exhibited strong positive correlations with both TLR2 (p = 0.56) and TLR4 (p = 0.46). TLR4 also showed positive correlations with MCP-1 (p = 0.60), IL-13 (p = 0.41), IL-8 (p = 0.41), IL-12(p70) (p = 0.36), and IL-6 (p = 0.60).
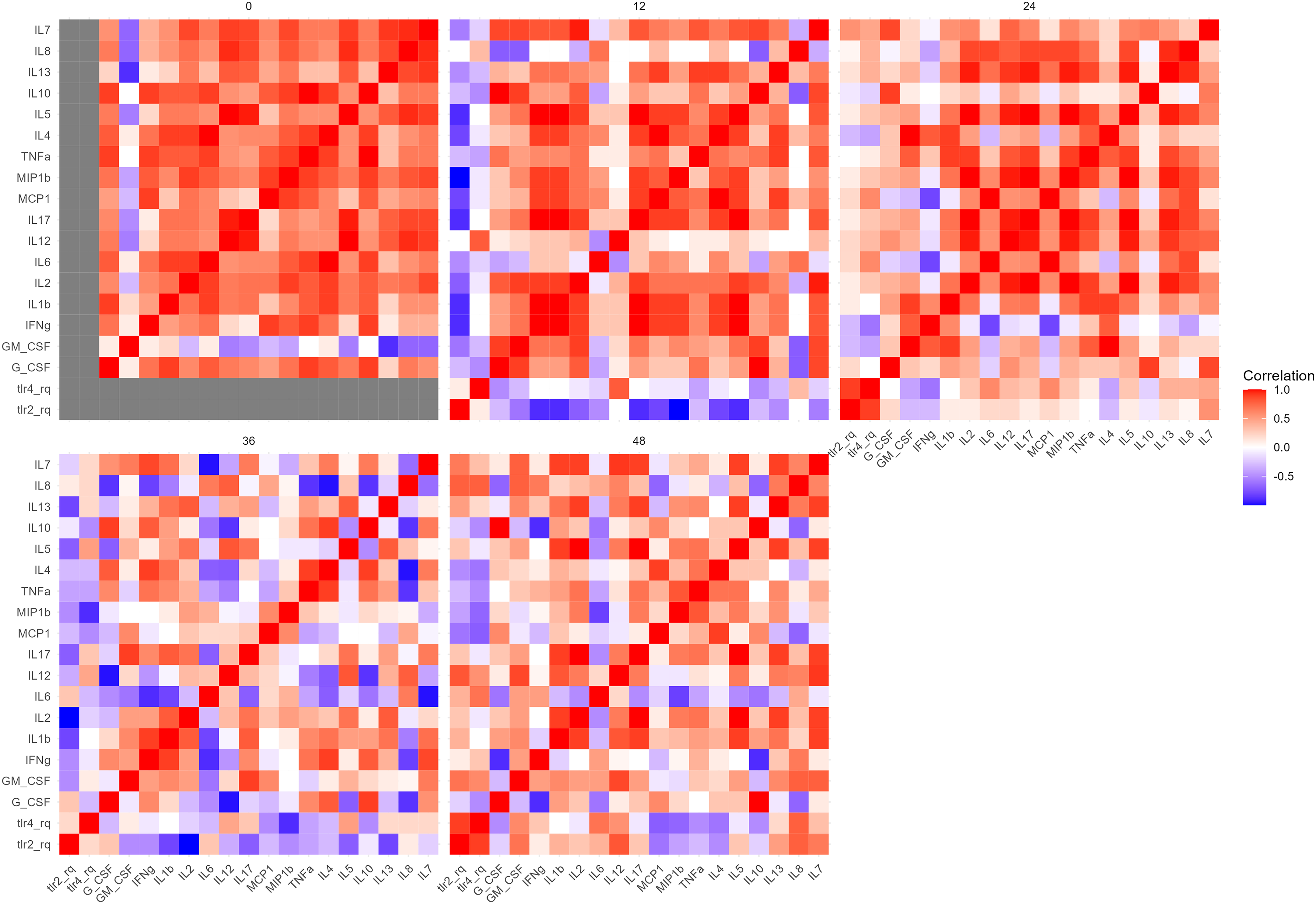
Correlation matrix between TLR2 and TLR4 transcriptional expression and cytokine levels over time in PBMC stimulated with HI B. bacilliformis. Spearman correlation was used.
Discussion
This study provides a descriptive characterization of the transcriptional patterns of TLR2 and TLR4 and the accompanying cytokine responses in human PBMCs exposed to B. bacilliformis. To our knowledge, this is the first dataset detailing the temporal behavior of these markers in response to this neglected pathogen. Given the exploratory nature of the work, the findings should be interpreted as preliminary transcriptional trends than evidence of functional receptor activation.
Our findings reveal that B. bacilliformis elicited measurable changes in TLR2 and TLR4 transcription, with most donors showing early peaks at 12 h. Cytokine patterns evolved over time, spanning pro-inflammatory, regulatory, and growth-factor pathways. These results outline an initial innate immune signature but do not allow mechanistic conclusions regarding downstream pathway activation. These observations contribute to defining the innate immune signature associated with PBMC exposure. Previous studies have reported that other Bartonella spp. may act as both agonists and antagonists of these receptors. 16 Thus, Popa et al. evaluated HI B. quintana in vitro in CHO cells transfected with TLR2 and TLR4, demonstrating that B. quintana activates the immune response exclusively through TLR2, inducing the production of TNF, IL-1β, and IL-6. Furthermore, the authors showed that B. quintana LPS functions as an antagonist of TLR4. 16 Regarding B. henselae, Scutera et al. reported that it induces TLR2, epidermal growth factor receptor, and nucleotide-binding oligomerization domain-containing protein 1 expression after exposure in mesenchymal stromal cells. 20 Additionally, McCord et al. (2005) studied the interaction of B. henselae with human microvascular endothelial cells to determine whether it promotes monocyte chemoattractant protein-1 (MCP-1), a chemokine involved in macrophage recruitment and known to be TLR4-mediated. Their findings indicated that MCP-1 induction was independent of TLR4 activation. 12
However, for B. bacilliformis, the information concerning TLRs is scarce; evidence suggests that B. bacilliformis may evade immune detection by TLR5, likely due to mutations in its flagellin that alter the receptor-binding domain, preventing proper recognition by TLR5. 11 Additionally, our results provide preliminary descriptive evidence of changes in TLR2 and TLR4 transcription during PBMC exposure to B. bacilliformis. This idea is consistent with TLR2 recognizing a wide variety of PAMPs, including lipoproteins, peptidoglycan and lipoteichoic acid, present in Gram-positive as well as Gram-negative bacteria. 21 On the other hand, TLR4 is primordial to recognize bacterial LPS. 22 Both TLRs are essential for detecting pathogens and initiating the innate immune response, serving as a critical component of the body's early defence against microbial invasion. 22 TLR4 is primarily associated with a Th1-type immune response and the production of pro-inflammatory cytokines such as IL-2, IFN-γ, and TNF-β. Notably, IFN-γ can suppress the Th2 pathway, reinforcing Th1 dominance. 23 In contrast, TLR2 is often linked to a Th2-type response, characterized by the release of anti-inflammatory cytokines including IL-3, IL-4, IL-5, and IL-10, and may inhibit Th1 polarization via IL-4, IL-10, or IL-13. 23 To date, the role of TLR4 in CD has only been reported in the context of vaccine development through immunoinformatics, where it has been proposed as a potential target for immunogenic design against B. bacilliformis.24,25
Coincidentally, the highest relative expression of both TLR2 and TLR4 was detected at 12 h, together with a significant upregulation of GM-CSF, IL-12, IL-13, IL-2, IL-5, and IL-7. This early cytokine pattern is consistent with responses described in other models involving TLR2 and TLR4 stimulation. The simultaneous increase of both receptors may reflect cooperative signaling, as atypical or low-endotoxic LPS has been shown to induce TLR2/TLR4 interaction and even heterodimer formation, potentially broadening PAMP recognition and contributing to mixed Th1/Th2 cytokine profiles. 26 Although our study cannot assess receptor interactions directly, this mechanism offers a plausible explanation for the transcriptional and cytokine responses observed. Notably, GM-CSF can be induced through both TLR2- and TLR4-mediated pathways in other bacterial stimulation models, aligning with our findings. 27 Importantly, IL-12 is a hallmark Th1-driving cytokine predominantly triggered by TLR4 linked to antigen-presenting cells. 28 IL-13 and IL-5, typically associated with type 2 immune responses, also show elevated levels upon TLR2 stimulation, reflecting innate immune skewing. 29 IL-2, aside from its classical T cell role, can be produced early by innate lymphoid cells in response to TLR2/4 ligation. 30 In addition, IL-7 and its early regulation are described as favoring lymphocyte survival in the context of innate immune activation, and it is associated with being a Th2-type cytokine. 31
The analysis of cytokine concentrations reveals significant temporal modulation, consistent with patterns previously reported in contexts where TLR2/TLR4 may participate, and could may reflect an initial innate immune response that potentially primes downstream adaptive immunity. Our correlation analyses demonstrated diverse immunological patterns over time: in the early phase (12 h), the positive correlation of TLR4 with IL-12p70 and IL-8 suggests exploratory temporal associations between TLR gene expression and cytokine levels in Th1 type responses. 32 The association with IL-8 is particularly relevant, since this chemoattractant is essential for the rapid recruitment of neutrophils and monocytes, key cells in the initial control of bacterial infections. Similarly, the relationship with IL-12p70 supports the classic function of TLR4 in the early polarization toward cellular responses. 32 At 24 h, the trend reversed, and positive correlations between TLRs and cytokines predominated, reflecting a parallel decrease in both variables. This finding could be interpreted as a period of regulation or adjustment of the response, in which the drop in TLR expression was accompanied by a transient decrease in the secretion of proinflammatory cytokines. However, negative correlations with IFN-γ, GM-CSF, and IL-4 indicate that certain mediators remained active, suggesting the onset of polarization toward Th1 and Th2 adaptive responses. 33 At 36 and 48 h, markedly different behaviors were observed for both TLRs, as these decreased, anti-inflammatory cytokines tended to increase, indicating mechanisms of cell regulation and recruitment. These observations motivate future mechanistic studies to test whether B. bacilliformis possesses immune-modulatory features that can not detect by the host easily such as B. quintana and B. henselae LPS, which had been reported as antagonist to TLR4. 34 Furthermore, the observed inter-individual variability in TLR2 and TLR4 expressions in response to B. bacilliformis stimulation, particularly the markedly elevated expression in certain individuals, may reflect biological variability, including potential genetic or epigenetic influences.21,35–38
This study has several limitations. Although the number of PBMC samples from healthy donors was limited, we performed a large number of measurements across multiple time points and a broad panel of biomarkers—including TLRs, cytokines, chemokines, and growth factors—which allowed us to generate new descriptive insights. Highlight that the use of heat-inactivated bacteria may alter PAMP structure and receptor interactions. Additionally, we did not perform functional assays (such as TLR neutralization, siRNA silencing, or TLR-specific reporter analyses), in order to elucidate the receptor-specific activation pathways. Moreover, the individual variability observed is biologically relevant and reflects the natural heterogeneity of human immune responses, especially in conditions of intracellular infection. In addition, the analysis was focused exclusively on TLR2 and TLR4, although other TLRs may also participate in the recognition of B. bacilliformis or contribute to the cytokine responses observed in this PBMC model. Additionally, TLR4 is known to undergo endosomal internalization after ligand engagement, where it can signal through alternative pathways that influence cytokine production. 39 Because our study did not assess intracellular receptor localization or the involvement of additional TLRs, the mechanistic interpretation of the transcriptional and cytokine patterns should be considered preliminary and warrants further investigation.
As conclusions, these findings contribute to a better understanding of the immune response in CD. This is the first study to characterize temporal transcriptional patterns of TLR2 and TLR4, together with cytokine secretion profiles, in human PBMCs exposed to B. bacilliformis, providing descriptive immunological signatures that may inform future mechanistic studies aimed at determining whether, and how, TLR2 or TLR4 participate in host responses during B. bacilliformis infection.
Footnotes
Acknowledgements
We would like to thank the technical staff at the laboratory of the Universidad Científica del Sur for their support during the execution of the project.
ORCID iDs
Ethical considerations
The study was approved by the Ethics Committee of the Faculty of Biological Sciences of the Universidad Nacional Mayor de San Marcos (051-2023-CBE-FCB-UNMSM).
Consent to participate
All participants signed the written informed consent form approved by the Ethics Committee of the Faculty of Biological Sciences of the Universidad Nacional Mayor de San Marcos (051-2023-CBE-FCB-UNMSM).
Consent for publication
Not applicable
Author contributions
AK, MJP, JEB, AMH, JR conceived and designed the experiments. AK, JEB performed the laboratory work and collected the data. AK, JEB, AMH, analyzed the results and performed statistical analysis. AK, MJP, JR, RGG wrote the manuscript. All authors reviewed and approved the final version of the manuscript.
Funding
We thank Programa Nacional de Investigación Científica y Estudios Avanzados (PROCIENCIA) [contract: N° PE501084497-2023] and Universidad Científica del Sur (087-2023-PRO99) for supporting this research.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.