
Abstract
Most women with type 1 diabetes mellitus (T1DM) have increased insulin requirements during pregnancy. However, a minority of women have a fall in insulin requirements. When this occurs in late gestation, it often provokes concern regarding possible compromise of the feto-placental unit. In some centres, this is considered as an indication for delivery, including premature delivery. There are, however, many other factors that affect insulin requirements in pregnancy in women with type 1 diabetes mellitus and the decline in insulin requirements may represent a variant of normal pregnancy. If there is no underlying pathological process, expedited delivery in these women is not warranted and confers increased risks to the newborn. We will explore the factors affecting insulin requirements in gestation in this review. We will also discuss some novel concepts regarding beta-cell function in pregnancy.
Keywords
Introduction
The insulin requirements of a woman with type 1 diabetes mellitus during pregnancy are variable. We will explore the reasons underlying this. Many women have increasing insulin requirements throughout gestation. A perplexing clinical issue is the woman with falling insulin requirements, particularly in late gestation. Whether this represents a true pathological process or a variant of normal is uncertain.
Insulin release, clearance and metabolism
Carbohydrate metabolism is predominantly influenced by insulin and its opposing hormone, glucagon. The incretin system, gastrointestinal factors and other counter-regulatory hormones also influence glucose metabolism. Insulin lowers blood glucose levels (BGL) by stimulating glucose uptake into the liver, muscle and adipose tissue, promoting glucose storage as glycogen and inhibiting glucose production. Pancreatic beta cells produce proinsulin. Proinsulin is enzymatically cleaved to insulin and C-peptide which are released in equimolar concentrations. 1 The metabolic activity of proinsulin is 10% that of insulin. C-peptide has traditionally been regarded to have a minor role in carbohydrate metabolism. 2
Glucose is transported into the beta cell through the glucose transporter, type 2 (GLUT2). It is phosphorylated to glucose-6-phosphate by glucokinase. The subsequent metabolism via glycolysis and the respiratory cycle increases the adenosine triphosphate (ATP) to adenosine diphosphate (ADP) ratio. This results in closure of the potassium-sensitive ATP channel, membrane depolarisation, calcium influx and exocytosis of insulin containing granules (see Figure 1). Glucose and, to a lesser extent, amino acids are insulin secretagogues. High concentrations of free fatty acids (FFA) induce insulin resistance. An acute elevation of FFA causes a compensatory increase in insulin secretion. Glucose disposal also occurs via insulin-independent pathways. Glucose effectiveness describes the ability of glucose to facilitate its own uptake and suppress hepatic glucose production in the basal insulin state.
Pathway of insulin secretion.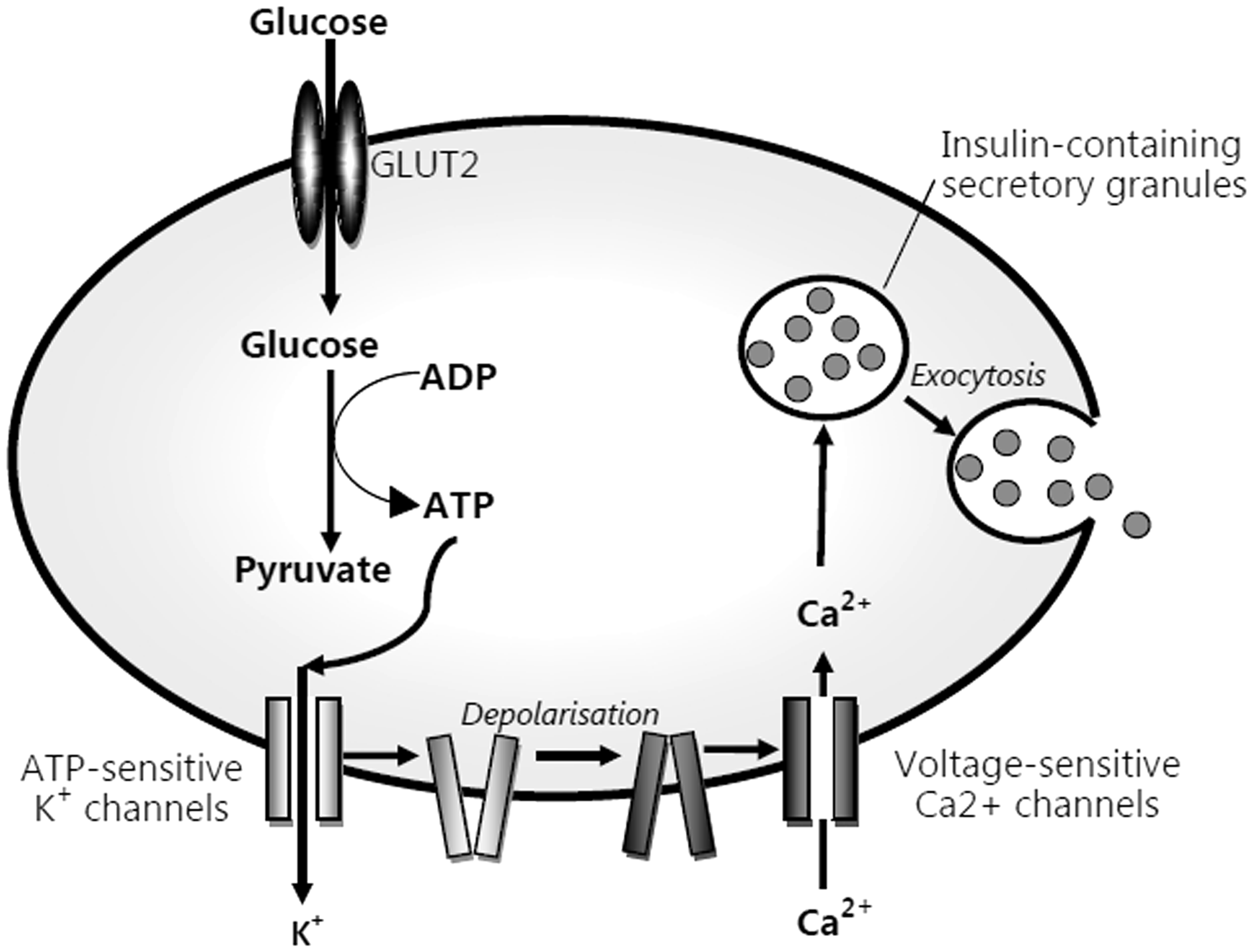
Insulin levels are lowest overnight. A rise in insulin secretion occurs at 4–6 am (dawn phenomenon) secondary to transient insulin resistance from increased cortisol and growth hormone concentrations. 3 Subsequently, insulin secretion decreases throughout the day. 3 A total of 40–60% of insulin undergoes first pass metabolism in the liver. 4 C-peptide is not extracted by the liver 4 and is predominantly renally excreted. The fractional hepatic extraction of insulin is inverse to endogenous insulin secretion. 5 The effects of insulin are mediated by a cell-surface receptor. Internalisation and dephosphorylation of the receptor terminates the action of insulin.
Insulin requirements in non-pregnant women with type 1 diabetes mellitus
Factors affecting insulin requirements in type 1 diabetes mellitus (T1DM).
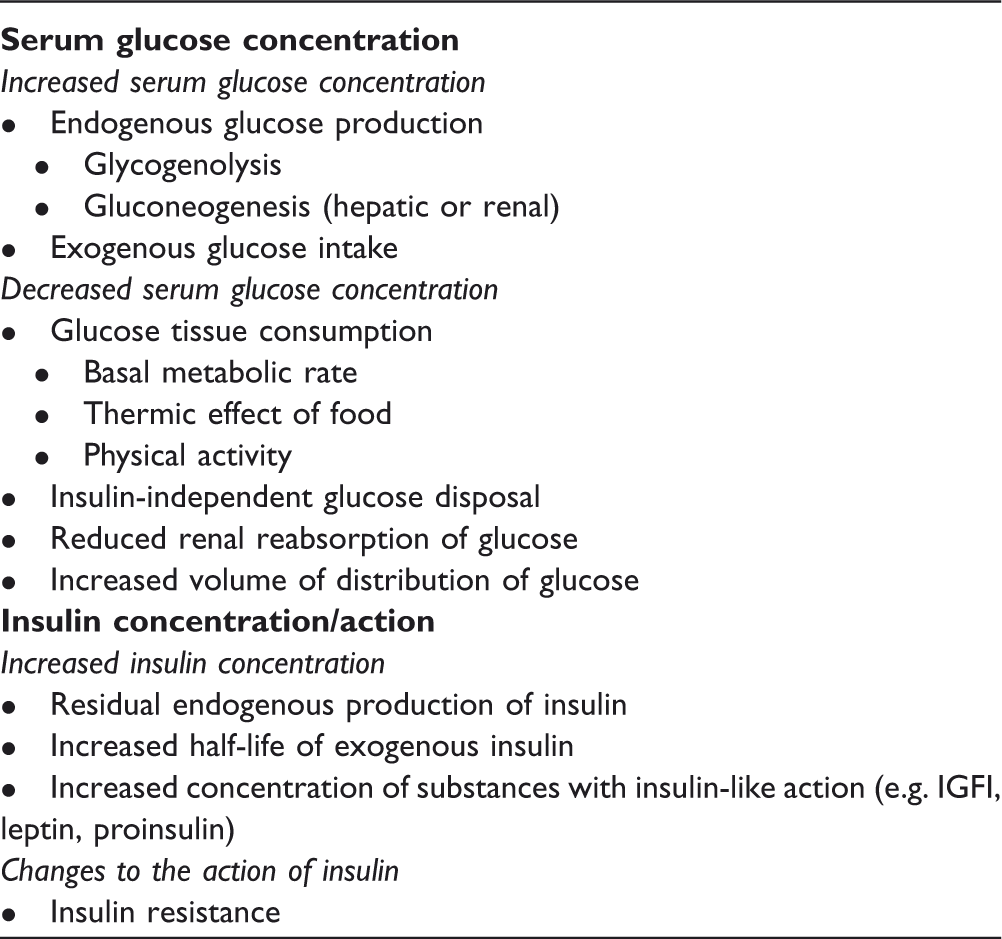
During pregnancy, changes in maternal metabolism facilitate the production and transfer of nutrients to the fetus.
The fetus primarily utilises maternally-derived glucose and an equilibrium exists between the fetal and maternal glucose pools. 10 In non-diabetic women, the plasma glucose concentration is decreased after an overnight fast from 6 to 10 weeks gestation and is unchanged throughout pregnancy.11,12 Despite the fall in fasting glucose, hepatic glucose production is increased in the basal state.3,12,13 The rate of gluconeogenesis in vivo can be estimated by labelled gluconeogenic precursors and deuterated water. In human pregnancy, these are limited by invasiveness and fetal exposure. The most robust evidence for increased glucose production is a trial of six healthy women using a primed constant infusion of 6-6 2H2 glucose pre-pregnancy, in early and late gestation. 13 This showed a 30% increase in basal endogenous production by late gestation after correction for fat-free mass. Studies using stable isotopes or deuterated water are limited by small numbers, different women as controls or data extrapolated from the postpartum period.10,14,15 However, using invasive techniques in pregnant dogs, Connolly et al. 16 showed that the net hepatic uptake of gluconeogenic substrates and total glycogenolysis was unchanged in pregnancy. This suggests that changes in the post-glycogenolytic partitioning of glucose (including futile cycling) account for the increased hepatic glucose output.
In normal pregnancy, insulin resistance facilitates the placental transfer of glucose. Using a hyperinsulinaemic euglycaemic clamp, the increase in insulin resistance is greatest between 16 and 26 weeks, with minimal increases thereafter. 17 It is attributed to a post-receptor defect in the action of insulin 18 induced by human placental lactogen (hPL), progesterone, oestrogen, cortisol, prolactin, leptin, placental growth hormone (PGH) and possibly tumour necrosis factor α (TNFα).19–23 To counter the insulin resistance, insulin production is increased. 24 This is enabled by structural and functional changes in the islets of Langerhans. 25 Animal studies indicate that both beta-cell hypertrophy and hyperplasia occur.26–32 Autopsy studies in non-diabetic women who died peri-partum suggest that the fractional beta cell mass is increased 1.4 fold because of an increase in small islet populations. 33 The increase in insulin-positive cells in ducts and scattered beta cells implies that this is due to beta-cell neogenesis. Animal studies,28,34 in vitro human islet studies 34 and limited human trials 35 indicate that hPL and prolactin, not GH or IGF1, are the likely mediators.
Fatty acids and ketones can serve as an alternative fuel source. Glucose and fatty acids interact and compete to serve as the predominant fuel source (Randle cycle). In the first half of gestation, increased insulin concentration with unchanged insulin sensitivity promotes lipogenesis. 36 Later in gestation, central leptin resistance facilitates ongoing fat acquisition in the fed state. 37 In the fasting state, increasing insulin resistance favours fatty acids as the fuel source.
Insulin resistance is greatest during the second trimester resulting in a marked increase in insulin requirements. During the first trimester, insulin resistance is less pronounced, however most women still have increased insulin requirements. A minority have a transient fall, typically at week 10.19,38–44 Possible factors include increased endogenous insulin production,35,45 the luteal-placental shift of progesterone production,46,47 increased insulin sensitivity, 48 hyperemesis or siphonage of glucose. The decrease is observed regardless of the adequacy of pre-pregnancy metabolic control. 43 Insulin resistance usually predominates in the third trimester. A subgroup of women has stable or slightly decreased requirements.43,49 A smaller percentage have a significant fall in insulin dose.19,50 Historically, this was attributed to failure of the feto-placental unit. However, there is no association between a fall in insulin requirement and adverse fetal outcome.19,50
Feto-placental factors affecting insulin requirements in pregnancy in women with T1DM
Two studies have explored the significance of falling insulin requirements in late gestation. The first, by McManus et al., 50 was a retrospective review of 32 women with T1DM. From 36 weeks until delivery, 62% had a fall in insulin dose. There were no adverse neonatal outcomes, including in the six women with a more than 15% decrease in insulin dose. Two years later, Steel et al. 19 published a prospective trial of 237 pregnancies. Almost 8% had a fall in insulin requirements of 30% or more within seven days in the third trimester. There was no association with adverse fetal outcome although 61% of these women delivered more than two weeks after the initial decline in insulin. However, the applicability to current practice is debatable as recruitment occurred from 1976 to 1991 and routine induction at 38 weeks was standard practice.
Maternal factors affecting insulin requirements in pregnancy in women with T1DM
Women with T1DM are at risk for developing gastroparesis during pregnancy. However, gastrointestinal symptoms have not been reported in the women with significant falls in insulin requirements in late gestation.19,50 Conversely, an increase in carbohydrate intake with increasing gestation has been described in non-diabetic women. 51 If this occurred in women with T1DM, an increase in insulin requirements would result.
Limited data suggests that gluconeogenesis and basal glucose turnover is comparable between diabetic and non-diabetic pregnancies.23,52 These studies are limited by small cohorts, failure to evaluate women pre-conception or in late gestation 52 or the use of ammonia excretion as indirect measure of renal gluconeogenesis. 53 The kidneys contribute about 40% to the total rate of gluconeogenesis. Consequently, overt or subclinical diabetic nephropathy could reduce gluconeogenesis. Using 6-6 2H2 glucose infusions pre-conception, in early and late gestation in women with GDM, there was no difference in the rate of gluconeogenesis. 13 The different pathophysiology limits the applicability to T1DM.
Maternal glucose consumption comprised physical activity, basal metabolic rate (BMR) and food-induced thermogenesis. Pregnant women, particularly during late gestation, have reduced physical activity.51,54,55 In sedentary individuals, approximately 80% of the daily energy expenditure is attributable to the BMR and food-induced thermogenesis. 56 There is a 6–7/kg% increase in BMR evident from early pregnancy with little change thereafter.51,57 Little available data suggests that food-induced thermogenesis is unchanged in pregnancy. 51
The fetus is largely incapable of producing glucose and is dependent on maternal glucose.10,58 This could impact on maternal insulin requirements if maternal glucose production was insufficient or the fetus was large for gestational age. A small retrospective study showed a non-significant trend toward greater birth weight in women with T1DM and the largest falls in insulin requirements in late gestation. 50 This was not validated in a larger prospective study. 19
During pregnancy there is an increase in the plasma volume with a corresponding change in the volume of distribution. The plasma volume in the third trimester is increased by 50% 59 compared to pre-pregnancy. However, it remains constant during the third trimester, 59 so should not explain a late fall in insulin requirements.
In pregnancy, the tubular maximum for glucose reabsorption is reduced by up to 50%, resulting in increased glucosuria.60–62 Animal studies suggest that changes to glucose handling by the distal nephron are responsible. 63 However, the fraction of the filtered glucose load reabsorbed by the kidneys shows no change at different gestational ages. 60 Changes in glucose tolerance, insulin sensitivity, physical activity and weight affect non-insulin-mediated glucose disposal.64,65 The effect of pregnancy on non-insulin-mediated glucose disposal or its parameters of glucose effectiveness at zero insulin and basal insulin effect is unknown. Research using a frequently sampled oral glucose tolerance test is required. Arachidonic acid (AA) and docosahexaenoic acid (DHA) are the major metabolites of essential fatty acids. Their concentration affects glucose and insulin concentrations.66,67 In animal studies long-chain PUFA enhance insulin sensitivity.68,69 A lower membrane DHA concentration is also associated with increased insulin resistance. Limited human data suggests a relationship between insulin sensitivity and long-chain PUFA, particularly AA, in skeletal muscle. 70 This is the major site of insulin-mediated glucose disposal. However, these patients had coronary disease and fasting insulin was used as a surrogate for insulin sensitivity, which lacks the sensitivity of clamp studies.
The fetus is reliant on the maternal transfer of essential fatty acids for growth and development. Women with pre-existing diabetes, particularly T1DM, have reduced red blood cell (RBC) DHA and cord plasma and RBC AA and DHA concentrations.71,72 This may affect fetal growth and development. Given the relationship between PUFA concentrations and insulin resistance, this may also affect maternal insulin requirements. However, non-esterified fatty acids are increased in pregnancy, which may reduce hepatic glucose uptake and increase glucose production.73,74
Insulin resistance is mediated by many hormones, most placentally-derived. A reduction in these would reduce insulin resistance. Progesterones, oestrogens, hPL and PRL continue to rise throughout gestation, especially during the second trimester with a plateau in the final weeks of pregnancy.75–77 Cortisol, PGH and TNFα show dramatic increases in late gestation.21,36,78 Therefore, in the absence of dysfunction of the feto-placental unit or fetal compromise, these hormones remain elevated in late gestation. Placental insufficiency would reduce the production of the hormones that mediate insulin resistance, and thereby decrease insulin requirements. Placental compromise can be attributable to hypertensive disorders in pregnancy, for example, pre-eclampsia which is more common in women with T1DM. In one study, women with the greatest decrease in insulin requirements in late gestation were more likely to have hypertensive disorders compared to those with stable or increased insulin requirements. 79
Studies concerning the clearance of insulin during pregnancy are discordant (see Appendix Table 2).24,80–84 This is partly attributable to failure to correct for changes in volume of distribution and small sample sizes. Most studies suggest no change to insulin clearance.24,81,84,85 Reduced clearance has been demonstrated in women with T1DM 80 and reduced hepatic insulin extraction in women with GDM. 86 The magnitude of reduction of hepatic extraction in the GDM study (30%) was similar to the overall reduction of clearance in the T1DM study (24%). This suggests that a change in hepatic extraction accounts for the difference in clearance rate. Changes in body composition and blood volume distribution can also affect BGLs. A correlation between the volume of distribution and clearance of insulin with insulin sensitivity has been observed in late gestation. 87 The mechanism for this is uncertain.
Animal studies suggest that proinsulin does not cross the placenta.88,89 The peripheral metabolism of proinsulin is unchanged by pregnancy in rodents. 89 Finally, the lower metabolic activity of proinsulin makes it unlikely that small changes would significantly alter insulin requirements. The concentration of leptin is increased during normal pregnancy.90,91 Leptin levels correlate with insulin sensitivity in pregnancy in women with normal glucose tolerance and GDM.22,91,92 Further, leptin administration to a mouse model of T1DM reduced BGLs, HbA1c and hyperketonaemia.93,94 Leptin also increases hepatic glycogen 93 and suppresses glucagon and growth hormone.93,94 In normal pregnancy, the peak leptin concentration is around 28 weeks 91 with little change from 20 to 30 weeks gestation. 95 However, in T1DM, there is a significant decline in free leptin levels from 20 to 30 weeks gestation. 95 It is therefore unlikely that leptin accounts for late changes in insulin requirements. The diabetogenic effect of IGF-1 is well documented. During pregnancy, PGH gradually replaces pituitary GH. Due to the structural similarity, both hormones are thought to mediate similar effects. Fuglsang et al. 49 demonstrated that hPGH levels rise especially after week 25 of gestation. 49 This correlated with birth and placental weights. However, from week 35 there was a non-significant decrease in hPGH. Although insulin requirements also decreased there was no correlation with hPGH, IGF-I, IGF-II levels or GH binding protein.49,96 However, the study was insufficiently powered to detect this relationship. Secondly, Nielsen et al. 97 showed that women with T1DM during pregnancy have lower IGF-1 and higher hPGH levels compared with healthy women. 97 Further lower IGF-1 levels were associated with more frequent and severe hypoglycaemia.
Novel concepts to explain altered insulin requirements in late pregnancy
Two studies have raised the possibility of increased endogenous insulin production in pregnancy.35,45 Ilic et al. 45 studied 10 pregnant women with long-standing T1DM and undetectable C-peptide pre-pregnancy. They documented a rise in C-peptide at 10 weeks gestation in all women with a fall in insulin requirements in 90%. The radioimmunassay used can be affected by cross reactivity with other hormones. The authors attempted to circumvent this by using three different antibodies. A more specific method is liquid chromatography-mass spectrometry isotope-dilution assay.98,99 Further, the mean concentration of C-peptide was 0.2 nmol/L (0.6 ng/mL), which approaches the lower limit of detection (0.1 ng/mL). At this point, the sensitivity of the assay is reduced.
A second study by Nielsen et al. 35 documented a gradual rise in C-peptide throughout pregnancy, with a rapid fall postpartum. At 8 weeks gestation, C peptide was detected in about 40% of women compared with 97% by 33 weeks. The absolute increase in C-peptide was associated with improved glycaemic control but not a decrease in insulin requirement. However, in other studies decreased requirements were observed after 36 weeks. Further, analysis of the entire group may have nullified a subgroup effect. Finally, the samples were not taken under standardised conditions.
About 50% of C-peptide undergoes renal extraction. Most is then reabsorbed and the remaining degraded by the kidney or secreted into the urine. 100 In pancreatic transplant recipients impaired renal function reduces the clearance of C-peptide.101,102 All patients in the Nielsen study and 8/10 in the Ilic study had normal renal function. However, a subclinical diabetic nephropathy could prolong the half-life of C-peptide. Secondly, in rodents, insulin, but not C-peptide, undergoes enhanced peripheral metabolism. 89 Whether this occurs in humans is unclear. Thirdly, in rodents, C-peptide does not cross the placenta 89 however, in other animal studies, small immunoreactive C-peptide fragments do cross the placenta and undergo degradation. 88 A recent study undertaken using a closed-loop insulin delivery system did not show any change in C-peptide from early to late gestation in T1DM. 103 However, 6 of the 10 women did not have fasting or stimulated C-peptide detectable in early or late gestation. Therefore, the results were available for four women. In addition, the immunochemiluminometric assay has a lower limit of detection of 5 pmol/L. However lower levels of C-peptide remain functional (see below).
Fetal-maternal microchimerism, that is the entry of fetal cells into the maternal circulation, may explain an increase in maternal insulin production. Some of these have stem-cell-like properties and may serve as the source for new beta cells. 104 Alternatively, these mesenchymal stem cells may re-educate maternal lymphocytes. Zhao et al. 105 used a “stem cell educator” which comprised a closed loop system that co-cultured lymphocytes from T1DM patients with cord-blood stem cells in vitro. These re-educated lymphocytes were returned to the patient’s circulation. A single treatment improved C-peptide levels, reduced A1c and lowered insulin requirements regardless of residual beta-cell function.
The proposed mechanism for increased endogenous insulin production is that the immunosuppression of pregnancy facilitates enhanced function of existing beta cells or an increase in islet cell mass via hypertrophy or hyperplasia, reduced apoptosis or neogenesis from duct cells. In the post-partum period, the autoimmune process flares and destroys the new or hyperfunctioning beta cells. This relies on the persistence of beta cells or new beta-cell formation in long-standing T1DM. Patients with residual beta-cell function have fewer diabetic complications.106–110 An early autopsy study showed persistence of insulin positivity in 37% of patients with T1DM of more than 21 years duration. 111 Similarly, beta cells have been identified in 88% of patients with T1DM 67 years duration. 112 Recently, Wang et al., 113 using an ultrasensitive assay, detected C-peptide in 10% of individuals with T1DM 30–40 years after disease onset. These beta cells remained functional as levels of 2.8 pmol/L responded to hyperglycaemia with increased C-peptide production. However, thymidine analog incorporation and radiocarbon dating indicated that beta-cell turnover is confined to the first three decades of life. 114 In T2DM and non-diabetic patients, intracellular lipofuscin body accumulation suggests that adult beta cells are established by 20 years. 115 In T1DM, the underlying autoimmune state is ongoing. It is possible that newly formed beta cells are continually destroyed, thereby stimulating beta-cell mitosis or neogenesis. In support of this, the rates of beta-cell apoptosis in T1DM is double that of controls. 112 This suggests that new beta cells are being produced and subsequently destroyed.
Another possibility is that of a dormant but potentially functional population of beta cells. Olsson and Carlsson 116 showed that in rats, 20–25% of all islets had low levels of oxygenation which correlated with metabolic activity and insulin production. This percentage doubled after pancreatic transplantation and almost disappeared after partial pancreatectomy. New beta cells may arise from neogenesis from duct cells as occurs in vitro in response to epidermal growth factor and gastrin. 117 Other growth factors also can cause neogenesis.118–121 In rats with new onset diabetes, leptin prolongs the survival of transplanted syngeneic islets. 122 Therefore, if new beta-cell formation occurs, the higher leptin levels in pregnancy may contribute to their persistence.
Against the idea of beta-cell restoration is the temporal relationship between immunosuppression in pregnancy and changes in insulin requirements. Many autoimmune conditions show a gradual improvement in pregnancy and usually a gradual deterioration weeks to months postpartum. However, the changes in insulin requirement can be abrupt. The fall in insulin requirement in late gestation can occur over a few days and the requirement for insulin postpartum can be within hours. A possible explanation is that there is a critical mass of beta cells that produces a clinically significant amount of insulin that acutely reduces insulin requirements in late gestation. Post-partum, the destruction of beta-cells may reduce the numbers below this critical mass, resulting in the recurrent dependence on insulin.
Conclusion
In summary, pregnancy is a state of insulin resistance. However, the action of insulin and the degree of insulin resistance is only one of the many factors that influence the requirement for insulin in women with T1DM during pregnancy. Other factors that affect the serum glucose concentration and the concentration of insulin or insulin-like substances are also highly relevant. A significant decline in insulin requirements, particularly in late gestation, historically provoked concern regarding compromise to the feto-placental unit. However, this change may not signify an underlying pathological process given the poor correlation with adverse clinical outcomes. Finally, the role of increased endogenous insulin production and stem cell modulation of the maternal immune system are intriguing concepts that warrant further investigation.
Footnotes
Conflict of Interest
None declared.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Guarantor
Naomi Achong
Contribution to authorship
NA was primarily responsible for the literature search, analysis and composition of the paper. HM and LC contributed to the planning of the paper, editing of the manuscript and analysis of the data.
Appendix
Clearance of insulin during pregnancy in diabetic and non-diabetic women.
Patients
Study size
Timing of evaluation
Technique
Result
Ref 23
GDM, Non-diabetic pregnant women
9 GDM, 7 non-diabetic
Pre-pregnancy, 12–14 weeks gestation 34–36 weeks gestation
Hyperinsulinaemiceuglycaemic clamp
Increased with advancing gestation with no difference between GDM and controls
Ref 80
T1DM
10
3rd trimester, 5–13months post-partum
Hyperinsulinaemic hypoglycaemic clamp
24% lower during pregnancy than postpartum
Ref 81
Non-diabetic pregnant women, non-diabetic non-pregnant women
8 pregnant, 5 non pregnant
>36 weeks gestation
Single injection of bovine insulin
Unchanged
Ref 82
Insulin treated diabetics
7
3rd trimester, 1–3 weeks post-partum
Euglycaemic insulin clamp
Unchanged
Ref 83
Non-diabetic pregnant women
20
3rd trimester, 4–8 weeks postpartum
Fasting molar c peptide/insulin ratio
Unchanged
Ref 84
Non-diabetic pregnant women, GDM
15 non-diabetic, 15 GDM
3rd trimester 6–8 weeks postpartum
Single intravenous injection of insulin with disappearance rate characterised by a two compartment model
Unchanged
Ref 85
Non-diabetic pregnant women, Non-diabetic non-pregnant women
11 pregnant, 5 non-pregnant
> 34 weeks gestation, 8–0 weeks postpartum
Single dose of IV porcine insulin with minutely measurements of insulin
Unchanged
