
Abstract
Bile acid metabolism is partially regulated through the activity of the gut microbiota. Primary bile acids can be deconjugated and modified by bacteria expressing bile salt hydrolase and other enzymes, changing bile acid recycling by changing the interactions between enterocytes and hepatocytes. The modified bile acids can also activate signalling in cells regulating metabolism including colonic L-cells, skeletal muscle cells and brown adipocytes. In pregnancy, both bile acid metabolism and gut microbiota composition are altered. In women with intrahepatic cholestasis of pregnancy, the changes in bile acid metabolism are exacerbated and there is some evidence that the gut microbiota composition is also altered. Here we review the crosstalk between the liver and the gut especially in women with intrahepatic cholestasis of pregnancy, with a focus on the role of the gut microbiota in this crosstalk.
Introduction
The human gut microbiota has a major impact on its host, from the fermentation of dietary fibre to the generation of metabolites, which affect metabolic and immunological processes. The gut microbiota is highly dynamic and is quick to respond to changes in dietary intake as well changes in host physiology and pathology. For instance, in pregnancy the composition of the gut microbiota is altered, becoming less diverse and more pro-inflammatory, and likely contributes to the increases in insulin resistance and weight. 1 The gut microbiota in humans also has a critical role in bile acid metabolism, altering the composition of the bile acid pool and thereby affecting cellular signalling in the host. This is an example of how the gut microbiota is part of the language by which the gut and the liver communicate both in health and in disease. Intrahepatic cholestasis of pregnancy (ICP), a condition characterised by increased bile acid concentrations, may be a condition where the crosstalk between gut and liver is altered. While the contribution of the gut microbiota in ICP is not yet clear, there is some evidence indicating that the gut microbiota is involved, which may provide additional avenues for treatment in the future.
In this article, we will discuss the crosstalk between the gut and the liver in bile acid metabolism, focusing on changes occurring in healthy pregnancies and those affected by ICP.
Bile acid synthesis
Bile acids are derived from cholesterol and their synthesis occurs in the liver. In humans, there are two pathways for bile acid synthesis, the classical and alternative pathways. The classical pathway converts cholesterol into 7α-hydroxycholesterol with catalysis through the enzyme CYP7A1. The 7α-hydroxycholesterol is then oxidised to 7α-hydroxy-4-cholesten-3-one and is then, by a multistep pathway, mainly converted to cholic acid (CA). In the alternative pathway, cholesterol is converted to 27-hydroxycholesterol with the help of CYP27A1 in the liver or extra-hepatic tissues, and is then mainly converted into chenodeoxycholic acid (CDCA). The final reaction for both CA and CDCA production in each of the pathways is catalysed by CYP27A1 (Figure 1).
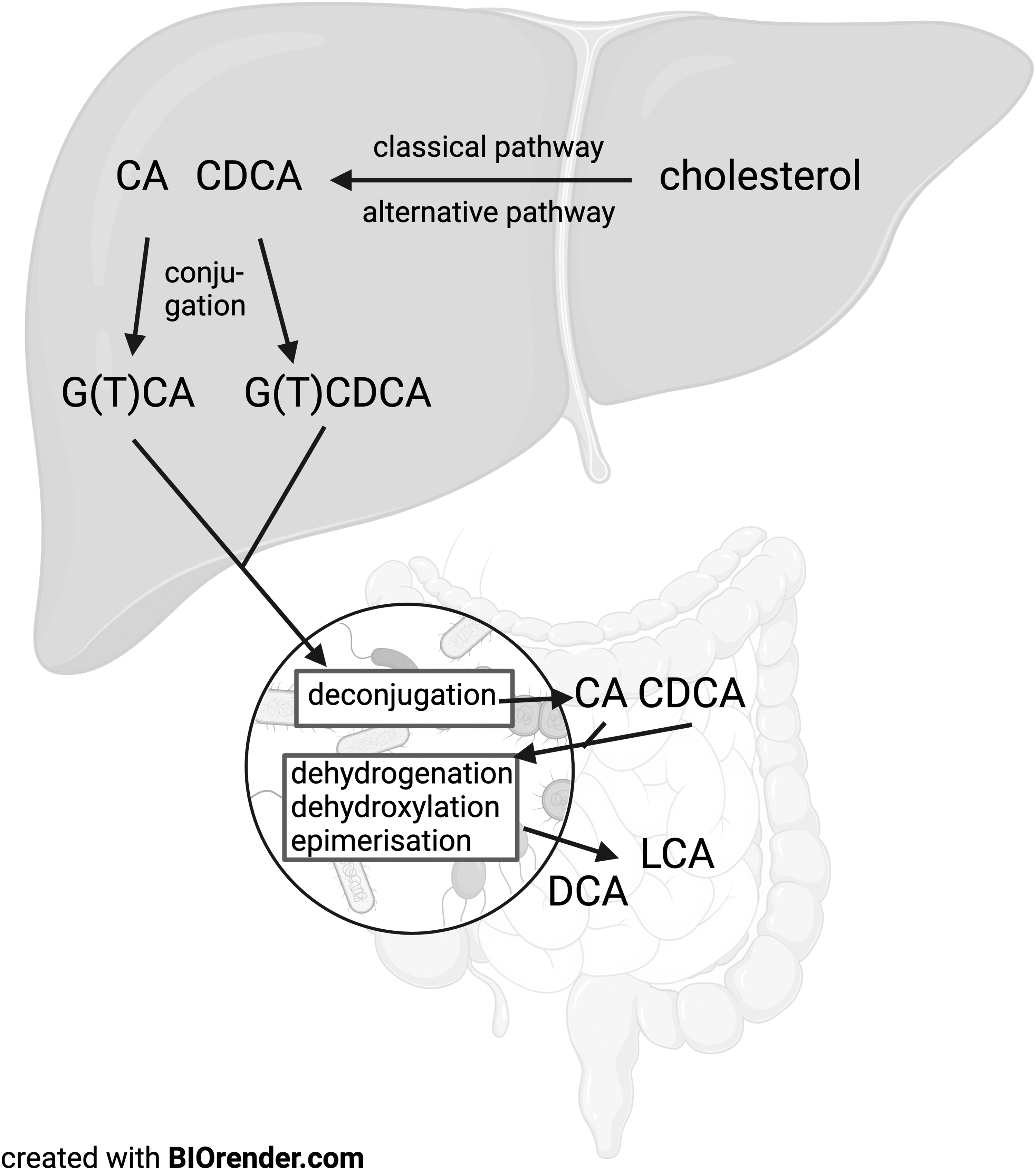
Overview of bile acid synthesis. The primary bile acids CA and CDCA are produced from cholesterol via the classical and alternative synthesis pathway respectively in the liver. CA and CDCA are then conjugated to glycine or taurine and exported from the liver to the lumen of the gastrointestinal tract where they can be deconjugated and further altered by dehydrogenation, dehydroxylation and epimerisation to secondary bile acids.
CA and CDCA are then conjugated to either the amino acid glycine or to taurine, with glycine-conjugation dominating in humans in a ratio of 2:1 to 3:1.2,3 The conjugated bile acids, or bile salts as they are called after conjugation, are known as glycine/taurine cholic acid (G/TCA) or glycine/taurine chenodeoxycholic acid (G/TCDCA). The bile salts are then actively transported into the bile ducts by transporters such as bile salt export pump (BSEP) and multi-drug resistance-associated protein 2 (MRP2) where they are mixed with bilirubin, cholesterol, fatty acids and lecithin as well as inorganic salts and water. Conjugated bile acids can be stored and concentrated in the gall bladder from where they are released upon stimulation by cholecystokinin (CCK) and FGF19 into the duodenum. 4 The gall bladder thereby contributes to the amount of available bile acids in the intestine and thus the enterohepatic circulation.
In mice, the synthesis pathway of primary bile acids is similar to that in humans, but α/β-murideoxycholic acid predominates with CA; there are only small amounts of CDCA present. 5 In mice, bile acids are predominantly conjugated to taurine and not glycine. These differences need to be kept in mind when generalising the results of murine studies to human (patho)physiology. 5
The conjugation of the bile acid to the amino acid increases the hydrophilic nature of part of the bile acid, while the other part is more hydrophobic. This makes it possible for the bile salts to make micelles and help with fat absorption. This altered hydrophobicity also makes conjugated bile salts less permeable to diffusion across plasma membranes, enabling high concentrations to be stored within bile; once secreted, the majority of reuptake of bile salts therefore occurs by membrane-bound bile acid transporters in the terminal ileum.
After a meal, bile is secreted into the gastrointestinal tract where it is critical for fat breakdown and absorption. The majority of the primary bile salts are recycled via the enterohepatic pathway in the distal ileum. However, a small portion of the bile salts are hydrolysed by bacteria containing bile salt hydrolases, which deconjugate the primary bile acids by removing their glycine or taurine residue, yielding unconjugated primary bile acids. 6 These can be further altered by bacterial dehydrogenases, dehydroxylases and epimerases, giving rise to a wide range of secondary bile acids. 6 Unconjugated primary and secondary bile acids can be taken up in the portal circulation by passive transport across the enterocytes, from which they travel in the portal circulation often bound to albumin. At the liver, they are actively transported back into hepatocytes, enabling them to be conjugated and secreted back into the intestine. Here, they can undergo more alterations by bacteria, resulting in an even more complex bile acid pool. Secondary bile acids are, in general, more hydrophobic and have a higher affinity for membrane lipids than primary bile acids. 7
Bile salt hydrolases
Bile salt hydrolases are present in the cytoplasm of some bacteria, especially Gram-positive bacteria belonging to the Firmicutes (recently reclassified as Bacillota), Bacteroidetes and Actinobacteria including the genera Clostridium, Bifidobacterium, and Lactobacillus8–10 but also in some Gram-negative genera9,10 and especially Bacteroides, which is known to express bile salt hydrolases. 11 The hydrolase activity is optimal in a slightly acidic environment (pH 4.5–6.0), 12 which can be found in the duodenum where the concentration of conjugated primary bile acids is high. There are multiple isoforms of bile salt hydrolases that differ more in their preference for the conjugated amino acid than for the type of BA to which glycine or taurine is conjugated. 13
The deconjugation by bile salt hydrolases is thought to reduce the toxicity of the bile acids for the bacteria given that bile acids are acidic detergents. 3 It also yields amino acids, which can be used by the bacteria to support growth or for energy supply, with glycine breakdown yielding ammonia and carbon dioxide, and taurine additionally yielding hydrogen sulphide. Sulphide availability is thought to increase the synthesis of DCA (deoxycholic acid, a secondary bile acid) and has been linked to increased cell proliferation and cancer. Bile salt hydrolase activity may also help to prevent overgrowth of pathogens, some of which use conjugated primary bile acids as a source for spore germination, for example Clostridium difficile. 14
Differences between biliary and faecal BA composition
The effects of the alterations to the bile acid pool by the bacteria in the gut are particularly demonstrated by comparing the composition of the biliary bile acid pool and the faecal bile acid pool. 7 The human biliary pool consists of about 35% CA and 35% CDCA, with the secondary bile acid DCA present at around 25% of the total pool. All other bile acids make up the remaining 5% of the pool. However, in the faecal pool, CA and CDCA only make up 4% of the pool. Thirty-four percent of the faecal bile acid pool is DCA, 29% is lithocholic acid (LCA) with the remainder consisting of a large range of different secondary bile acids. Ursodeoxycholic acid (UDCA) is present at similar proportions (2%) in both the biliary and the faecal pool.
Bile acid signalling
Bile acids bind to the apical sodium bile acid transporter located on the apical membrane of ileal enterocytes (Figure 2); although able to bind all bile acids, the transporter has a preference for conjugated bile acids.15,16 The bile acids then transverse the enterocyte and are secreted into the portal vein by the organic solute transporter heterodimer OSTα/β. 17 They are then taken up by OATP (organic anion transporter polypeptides) or NTCP (sodium/taurocholate cotransporter polypeptide) on the plasma membrane of hepatocytes 18 and are transported to the nucleus, where they can bind to the nuclear bile acid receptor FXR (farnesoid X receptor), which is often in a heterodimer with the RXR (retinoid X receptor). 19 Activation of the FXR/RXR complex increases expression of the nuclear receptor SHP (small heterodimer partner), which binds to nuclear LRH-1 (liver receptor homologue 1), resulting in inhibition of the transcription of CYP7A1. 20 This reduces the first step in bile acid synthesis from cholesterol, and thereby prevents excessive bile acid synthesis.
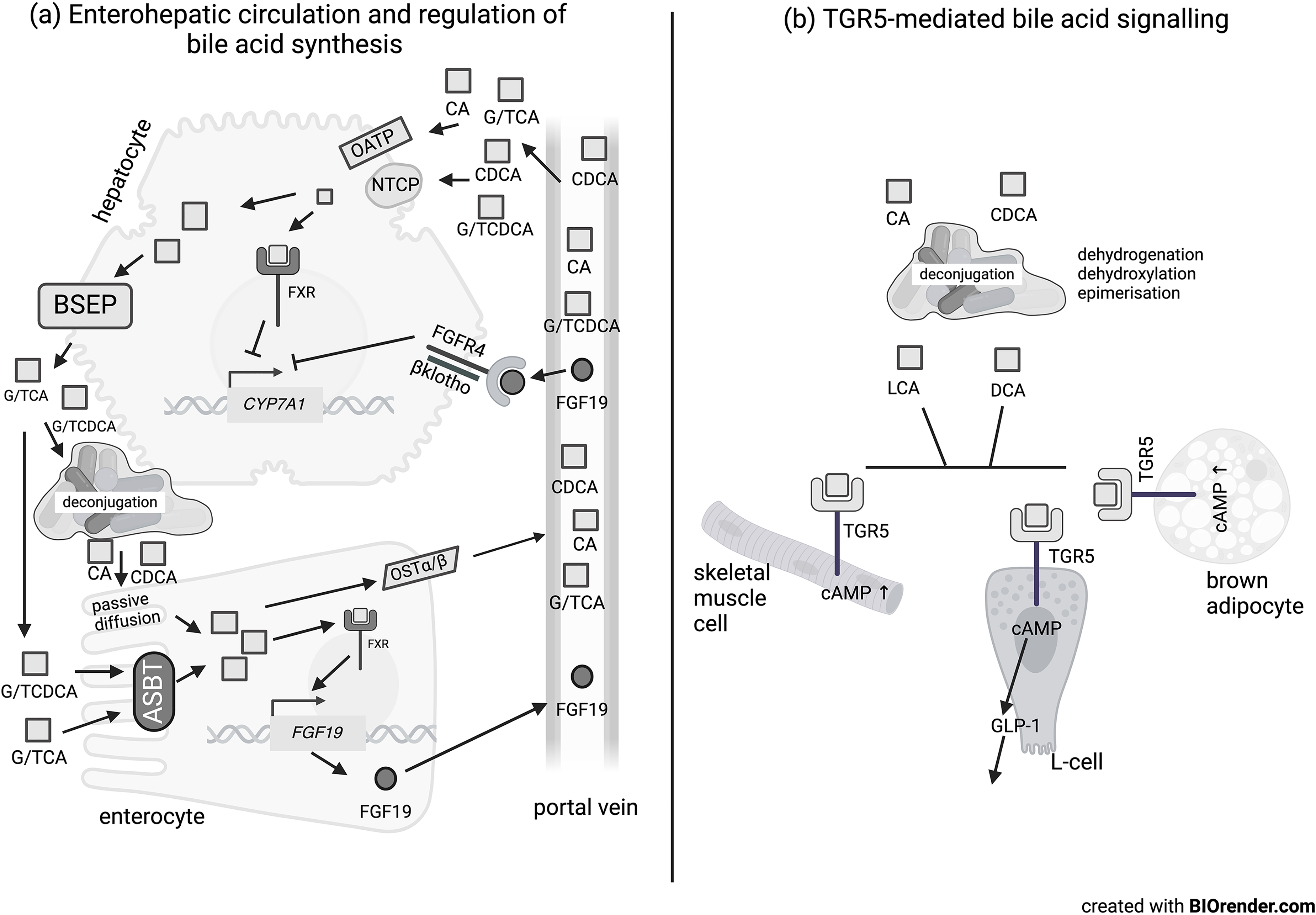
Overview of bile acid metabolism and signalling. (a) Enterohepatic circulation of bile acids between liver and gut via the portal vein. Conjugated primary bile acids are taken up by active transport in the enterocyte by the ASBT transporter whereas deconjugated primary bile acids are taken up to a lesser extent by passive diffusion. In the enterocyte, this stimulates FGF19 expression and secretion into the portal vein via stimulation of the FXR receptor. The bile acids can also be transported out into the portal vein via the OSTα/β transporter. The primary bile acids from the portal circulation bind the transporters OATP and NTCP on the hepatocyte and be transported out again via the BSEP. They can also activate FXR and also inhibit bile acid synthesis. FGF19 can bind to FGFR4/β-klotho in the hepatocyte and inhibits the expression of CYP7A1, which reduces bile acid synthesis. (b) Signalling by secondary bile acids through the TGR5 receptor in skeletal muscle cells, colonic L-cells and brown adipocytes.
Bile acids in the ileal enterocytes can also bind to the nuclear FXR/RXR receptors. 19 Binding of bile acids to FXR leads to transcription of fibroblast growth factor 19/15 in humans or rodents respectively, which is then secreted into the portal vein. 19 FGF19 (15) bind to the FGFR4/β-klotho receptor complex and leads to an intracellular signalling cascade that reduces expression of CYP7A1. 21 This again prevents bile acid concentrations from becoming detrimentally high through a negative feedback loop. Activation of FXR not only affects bile acid homeostasis but also other metabolic processes, including glucose and lipid homeostasis, insulin signalling and immune responses. 22
Bile acids also act as signalling molecules at cell surface receptors (Figure 2). They bind to the TGR5 or M-BAR (membrane bile acid receptor), a G-protein coupled receptor, on many different tissues, including colonic L-cells, skeletal muscle and brown adipocytes. 8 In skeletal myocytes and brown adipocytes, TGR5 activates deiodinase 2 and helps to convert T4 into active T3. 23 In colonic L-cells, activation of TGR5 leads to release of the incretin hormone GLP-1. 24 Thus, signalling by bile acids is also associated with regulating energy metabolism, thermogenesis, insulin signalling and inflammation. 23 GLP-1 secretion is also stimulated by short-chain fatty acids that are produced by the gut microbiota through fermentation of dietary fibre, 25 indicating that different microbial metabolites can work synergistically or antagonistically to affect host function.
Bile acids in pregnancy and in ICP
In pregnancy, total serum bile acids average around 4 μmol/L throughout pregnancy with a range of 0.3 to 16.7 μmol/L. 26 Across pregnancy, the proportion of conjugated bile acids versus unconjugated bile acids increases, with the change appearing between 28 and 31 weeks' gestation. 27 In ICP, non-fasting total serum bile acids are ≥19 μmol/L with concentrations in severe ICP being ≥40 μmol/L and ≥100 μmol/L in very severe ICP. 28 In human pregnancy, there is reduced enterohepatic feedback, demonstrated by decreased post-prandial circulating concentrations of FGF19. 29 This is associated with increased overall production of bile acids, as measured by increased fasting concentrations of C4 (7α-hydroxy-4-cholesten-3-one) – an intermediate in the pathway of de novo bile acid synthesis. 29 As discussed above, women with ICP have similar concentrations of FGF19 to pregnant women with normal bile acids, 30 though in women with recurrent ICP, β-klotho concentrations are decreased. 30 Given that both FGF19 and β-klotho are required for activation of the FXR in the liver, it is possible that, despite normal FGF19 concentrations, its signalling is altered in women with (recurrent) ICP. However, women with ICP who are treated with UCDA have higher FGF19 concentrations and lower C4 concentrations, 31 indicating that the regulation of FXR in the liver may be restored.
Gut microbiota in ICP
The gut microbiota changes across gestation, becoming more pro-inflammatory and less diverse according to some studies,1,32,33 whereas others report no such or only limited changes.34–36 This may reflect differences in study size and populations, sampling frequency, and method of gut microbiome assessment. Studies transplanting the gut microbiota of healthy women in their third trimester of pregnancy to germ-free mice resulted in the mice becoming more insulin-resistant and gaining weight compared with counterparts who received a transplant from the first trimester of pregnancy. 1 This suggests that the gut microbiota may be driving some of the physiological changes observed in pregnancy. It is possible that the microbiota also drives pathological processes in ICP. Indeed, a study comparing gut microbiota composition with 16S rRNA gene amplicon sequencing in 21 women with ICP, of whom 17 were treated with UCDA, and 14 healthy controls showed that UCDA treatment increased the abundance of bacteria belonging to the Bacteroidetes phylum and altered the Bacteroidetes:Firmucutes ratio. 31 Women with a lower Bacteroidetes:Firmucutes ratio had lower bile salt hydrolase activity and also had higher faecal concentrations of UCDA and LCA as well as unconjugated bile acids. 31 In vitro treatment of mouse colonic enterocytes with deoxycholic acid or LCA caused increased GLP-1 release as compared with controls, 31 indicating that altered bile acid composition in the colon may alter the secretion and thus the activity of enteroendocrine hormones.
A recent, larger study of 50 women with ICP and 50 without ICP comparing the gut microbiota composition using metagenomic sequencing showed no differences in diversity between the groups. 37 There was a small number of bacterial species with increased abundance in the women with ICP, including Bacteroides fragilis, several Klebsiella species and Enterobacter cloacae. B. fragilis abundance was especially increased in women with moderate to severe ICP (total bile acids ≥40 μmol/L). B. fragilis can degrade human glycoproteins and mucus, and it can ferment sucrose, glucose, fructose and inulin, which leads to the production of acetate, propionate and succinate. 38 It also secretes antibacterial compounds and carries a wide variety of antibiotic resistance genes and efflux pumps. 39
Transplantation of a stool sample from patients with ICP or just of B. fragilis into mice treated with antibiotics to remove their gut microbiota resulted in the mice developing altered total bile acids, abnormal liver function and altered hepatic morphology. It also affected pregnancy outcomes, including lower pup weight and fetal loss, thereby resembling the ICP state. 37 Transplantation in microbiota-depleted mice with B. fragilis resulted in decreased concentrations of conjugated bile acids, especially glycine-conjugated deoxycholic acid (GCDA), likely through its expression of a specific BSH enzyme isoform. 37 B. fragilis colonisation also increased circulating C4 concentrations, decreased circulating FGF15 (the murine equivalent of FGF19) concentrations and inhibited the mRNA expression of FXR-regulated genes in the liver and the ileum, most likely in response to altered bile acid metabolism. 37 The changes were reversed by supplementation with GDCA in the mice. 37 These results indicate the importance of B. fragilis in the pathogenesis of ICP, when tested in a mouse model where the normal microbiota was depleted. However, whether the effect is similar when B. fragilis abundance is increased in the context of a full microbiota needs confirmation. In addition, the role of increased B. fragilis abundance needs to be confirmed in metagenomic analyses of other human cohorts although increases to BSH-producing Bacteroides species (including B. fragilis) have been reported to be associated with changed bile acid composition in people with colorectal cancer 40 while the order/class that B. fragilis belongs to was found to be positively associated with ICP in two 16S rRNA gene amplicon sequencing studies.41,42 It is also not yet clear if women predisposed to ICP have altered B. fragilis abundance already prior to pregnancy, and whether the changes in B. fragilis abundance precede symptoms. If this is the case, strategies to alter the gut microbiota composition could be employed prior to the start of pregnancy in high-risk women.
Gut microbiota and circulating mediators
There are also interactions between the role of the gut microbiota in bile acid and choline and carnitine metabolism. Dietary protein from meat is a major source of choline and carnitine in the intestine. The gut microbiota converts choline and carnitine into the gas trimethylamine (TMA). This can be taken up by the enterocytes and be converted by flavin monooxygenase 3 (FMO3) in the liver into trimethylamine-N-oxide (TMAO), which is associated with increased cardiovascular disease. Women have slightly higher concentrations of hepatic FMO3 than men due to a negative regulation of expression by testosterone. 43 It is not clear if pregnancy changes FMO3 abundance or activity. There is however some evidence that FMO3 is activated by hepatic FXR, which itself is regulated by bile acids, 43 providing a good example of the crosstalk between gut and liver.
Treatments and effects on microbiome
UCDA, which is a more hydrophilic bile acid, 44 is present in low concentrations in humans and is also used as treatment for ICP. UDCA acts as a signalling molecule by inhibiting the FXR receptor activation and replaces the more hydrophilic bile acids within the bile acid pool, comprising approximately 60% of circulating bile acids in treated patients with ICP. 45 UCDA also improves biliary bicarbonate (HCO3−) secretion, which maintains bile flow and helps to protect the cholangiocytes from protonated hydrophobic bile acid monomers and thereby apoptosis.46,47 However, UCDA treatment is not effective for itch reduction in many patients, and did not reduce maternal bile acids nor did it reduce adverse perinatal outcomes in women with ICP in the PITCHES randomised controlled trial. 48 This could be partially due to differences in the maternal gut microbiota composition, similar to what is observed in patients with primary biliary cholangitis treated with UCDA. 49
Obeticholic acid (OCA) is a synthetic FXR agonist, which has similarity to CDCA but with a two orders of magnitude greater potency for binding to FXR. 50 It reduces bile acid production and promotes biliary flow 51 and could perhaps be an alternative treatment for ICP. 52 However, OCA use is associated with marked pruritus and risk of liver injury, and may therefore not be appropriate for use as a treatment of ICP, thus alternative treatments are required.
Rifampicin, which is a broad-spectrum antibiotic and agonist of the pregnane X receptor, can stimulate the enzyme CYP3A4 to hydroxylate bile acids. 53 It can also reduce the expression of CYP7A1 in vitro, which reduces hepatic production of bile acids. 53 However, it is not yet clear if rifampicin reduces pruritus and if it changes the composition of the gut microbiota in women with ICP, though the ongoing TURRIFIC RCT 54 will provide these insights.
Conclusion
As we have demonstrated, there is extensive communication between the liver and the gut through bile acid metabolism and gut microbiota metabolites (Figure 3). In women with ICP, the role of the gut microbiota and, with that, the possibilities to target the microbiota either before or during pregnancy, needs further exploration. Further studies need to explore if the gut microbiota is altered prior to the development of symptoms as, if so, this would emphasise the importance of additional treatment strategies targeting this aspect of ICP pathogenesis.
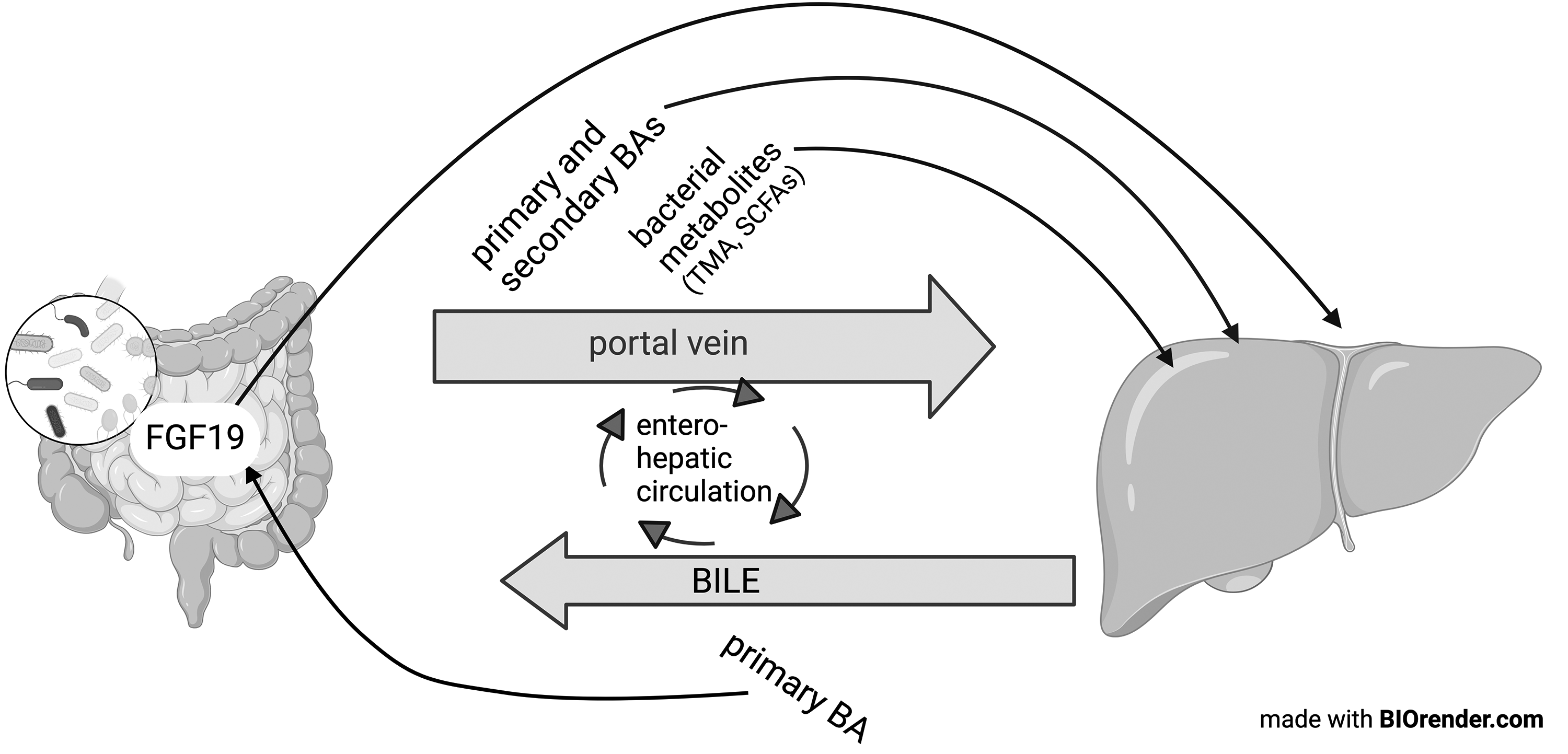
Overview of crosstalk between the liver and gut and the involvement of bacterial metabolites.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Guarantor
MDN.
Contributorship
MDN and CO researched the literature. MDN wrote the first draft of the manuscript. All authors reviewed and edited the manuscript.