
Abstract
Emotions can have both enhancing and impairing effects on various cognitive processes, from lower (e.g., perceptual) to higher level (e.g., mnemonic and executive) processes. The present article discusses emerging brain imaging evidence linking these opposing effects of emotion, which points to overlapping and dissociable neural systems involving both bottom-up and top-down mechanisms. The link between the enhancing and impairing effects is also discussed in a clinical context, with a focus on posttraumatic stress disorder (PTSD), where these opposing effects tend to co-occur, are exacerbated, and are detrimental. Overall, the present review highlights the need to consider together enhancing and impairing effects of emotion on cognition in studies investigating emotion–cognition interactions.
Keywords
Background
Recent research using behavioral, brain imaging and lesion methods emphasizes the intimate relationship between emotion and cognition, and shows that emotions can have complex influences on various cognitive processes. Specifically, emotions may affect both lower level (e.g., perceptual) and higher level (e.g., mnemonic and executive) cognitive processes, and the effects could be either beneficial/enhancing or detrimental/impairing. On the one hand, emotional stimuli/events can benefit from enhanced perceptual processing due to their ability to “capture attention,” and hence through prioritized processing they can be better encoded and remembered. On the other hand, task-irrelevant emotional information may lead to increased distraction and impaired performance in ongoing cognitive tasks. Moreover, these effects of emotion on cognition could be immediate and long-lasting and involve both automatic/bottom-up and controlled/top-up mechanisms. The emphasis of this review article is on investigations examining the neural mechanisms of enhancing and impairing effects of emotion and linking them in both healthy and clinical populations. The first two sections summarize evidence from studies separately examining the enhancing and impairing effects. The third and fourth sections discuss emerging evidence from studies linking enhancing and impairing effects, both in healthy and clinical groups, respectively. The review ends with concluding remarks and a brief presentation of open issues and future directions.
Enhancing Effects of Emotion on Cognition
Available evidence shows that emotional stimuli can benefit from enhanced perceptual processing due to their ability to “capture attention,” and hence because of this prioritized processing they can be better encoded and remembered (Dolcos & Denkova, 2008; Dolcos, Denkova, & Dolcos, 2012; Dolcos, LaBar, & Cabeza, 2006; Phelps, 2006; Pourtois, Schettino, & Vuilleumier, 2013; Vuilleumier, 2005). This section discusses evidence of enhancing effects of emotion on perception and episodic memory (EM), or memory for specific personal events.
The Enhancing Effect of Emotion on Perception
There is evidence of facilitated perceptual processing of emotional information reflected in better and/or faster detection of emotional stimuli, as compared with neutral stimuli (Ohman, Lundqvist, & Esteves, 2001; Pourtois, Grandjean, Sander, & Vuilleumier, 2004). At the neural level, these facilitating effects involve the amygdala (AMY), an almond-shaped group of nuclei located within the medial temporal lobe (MTL). Activity in the AMY was also found to correlate with activity in perceptual brain regions, typically according to the category of the stimuli (e.g., lateral extrastriate cortex for visual stimuli, in general; fusiform gyrus [FG], particularly for faces; superior temporal sulcus for voices; Grandjean et al., 2005; Vuilleumier, Armony, Driver, & Dolan, 2001). While traditionally this region has been associated with processing of negatively valenced emotions, particularly fear (Adolphs, Tranel, Damasio, & Damasio, 1995; Calder, Lawrence, & Young, 2001; Morris et al., 1996; Whalen et al., 1998; Zald, 2003), a number of functional neuroimaging studies involving both positive and negative stimuli, equated for emotional arousal, have also reported AMY responses to various positively valenced stimuli, including verbal (Garavan, Pendergrass, Ross, Stein, & Risinger, 2001; Hamann & Mao, 2002; Kensinger & Schacter, 2006b), pictorial (Dolcos, Graham, LaBar, & Cabeza, 2003; Hamann, Ely, Hoffman, & Kilts, 2002; Kensinger & Schacter, 2006b), olfactory (Anderson, Christoff, Stappen, et al., 2003) and gustatory (Small et al., 2003) stimuli.
The latter evidence is also emphasized in meta-analyses identifying valence-independent effects in AMY (Sergerie, Chochol, & Armony, 2008; Wager, Phan, Liberzon, & Taylor, 2003; but see Straube, Pohlack, Mentzel, & Miltner, 2008). However, hemispheric dissociation in the AMY has been linked to emotional valence (negative vs. positive), with right AMY showing preferences for negatively valenced stimuli (Canli, Desmond, Zhao, Glover, & Gabrieli, 1998; Dolcos et al., 2003) and left AMY for positively valenced stimuli (Canli et al., 1998). Of note, hemispheric asymmetries in the AMY have also been attributed to the level of processing (automatic vs. elaborated), with right AMY being more involved in initial, automatic detection of emotions, and the left AMY being involved in more elaborated, cognitive representations of emotions (Glascher & Adolphs, 2003; Morris et al., 1998; Morris, Ohman, & Dolan, 1999; Phelps et al., 2001). Finally, regarding the timing of emotion processing beyond the issue of lateralization aforementioned, traditionally it has been considered that emotion processing has privileged access to processing resources, and thus it is processed fast and automatically. However, whether processing of emotional information is automatic or depends on the availability of attentional resources is a matter of current debate (Anderson, Christoff, Panitz, De Rosa, & Gabrieli, 2003; Pessoa, 2005; Pessoa, McKenna, Gutierrez, & Ungerleider, 2002; Vuilleumier, 2005; Vuilleumier et al., 2001; Vuilleumier & Driver, 2007), as we will see in the section focusing on the impairing effect of emotion on perception.
The Enhancing Effect of Emotion on Episodic Memory
There is abundant evidence that emotional events are typically better and more vividly remembered than nonemotional ones (Bradley & Lang, 1994; Christianson, 1992b; Comblain, D’Argembeau, & van der Linden, 2005; Schaefer & Philippot, 2005; Talarico, LaBar, & Rubin, 2004). The enhancing effects of emotion on EM have been investigated at various stages of memory (from the early stages of memory formation—encoding and early consolidation of memory traces—to their later retrieval) and usually according to the two orthogonal affective dimensions, arousal and valence (Lang, Greenwald, Bradley, & Hamm 1993; Russell, 1980). To eliminate potential confounding effects of general perceptual processing and to specifically identify the enhancing effects of emotion on memory, the typical procedure used in brain imaging studies is to calculate the so-called difference in memory (Dm) effect—that is, the difference between brain activity for remembered versus forgotten items (Shafer, Iordan, Cabeza, & Dolcos, 2011). The extant research on the neural correlates of the memory-enhancing effect of emotion emphasizes the role of the AMY and memory-related MTL regions (hippocampus [HC] and the associated entorhinal, perirhinal, and parahippocampal cortices; see Figure 1) in the encoding (Dolcos, LaBar, & Cabeza, 2004b; Kensinger & Corkin, 2004; Kensinger & Schacter, 2006a; Ritchey, Dolcos, & Cabeza, 2008; Sergerie, Lepage, & Armony, 2006), consolidation (Ritchey et al., 2008), and retrieval (Dolcos, LaBar, & Cabeza, 2005; Kensinger & Schacter, 2005) stages. Importantly, evidence reveals that emotional memory enhancement effects emerge from interactions between AMY and the memory-related MTL regions (Canli, Zhao, Brewer, Gabrieli, & Cahill, 2000; Dolcos et al., 2004b; Hamann, Ely, Grafton, & Kilts, 1999; Kilpatrick & Cahill, 2003; Richardson, Strange, & Dolan, 2004), and that these interactions contribute to the persistence of emotional memory over time (Ritchey et al., 2008). Regarding the role of basic emotional properties of the stimuli, these investigations revealed that the memory-enhancing effects of emotion are driven by arousal rather than by valence, as similar effects are observed for positive and negative items matched for arousal (Murty, Ritchey, Adcock, & LaBar, 2010), although valence-related effects have also been identified (Mickley Steinmetz, Addis, & Kensinger, 2010; Ritchey, LaBar, & Cabeza, 2011).
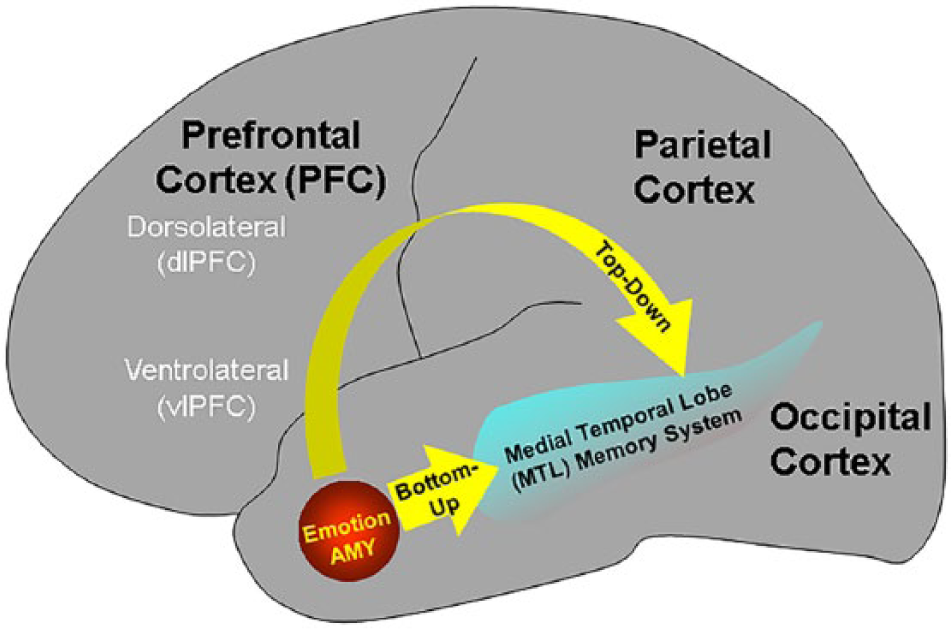
Direct/bottom-up and indirect/top-down neural mechanisms linked to the enhancing effects of emotion. Two main mechanisms involved in the memory-enhancing effect of emotion were identified: (1) direct/bottom-up mechanisms involving a basic emotion region (i.e., AMY) and memory-related MTL brain regions (e.g., hippocampus [HC]), and (2) indirect/top-down mechanisms involving modulatory influences from the prefrontal cortex (PFC; also including the parietal cortex [PC]) by enhancing strategic, semantic, working memory, and attentional processes. The visual cortical areas (occipital cortex) are also illustrated because they are susceptible to influences from emotion processing regions. Adapted from Dolcos et al. (2012).
In addition to the basic/direct AMY-MTL mechanisms, the memory-enhancing effect of emotion also benefits from the contribution of indirect/mediated mechanisms involving the prefrontal cortex (PFC) and the lateral parietal cortex (LPC; Dolcos, Iordan, & Dolcos, 2011). In the PFC, memory-enhancing effects of emotion have been linked to activity in both ventrolateral PFC (vlPFC) and dorsolateral PFC (dlPFC; Canli, Desmond, Zhao, & Gabrieli, 2002; Dolcos, LaBar, & Cabeza, 2004a; Kilpatrick & Cahill, 2003). Given the role of the left vlPFC regions in semantic processing (Kapur et al., 1996; Poldrack et al., 1999; Shallice et al., 1994), and the role of the dlPFC regions in working memory operations (D’Esposito, Postle, & Rypma, 2000; Owen et al., 1999; Petrides, 1995), it has been suggested that emotion enhances memory in part by promoting semantic (vlPFC) and working memory (dlPFC) processing (Dolcos & Denkova, 2008; Dolcos, LaBar, et al., 2006). Consistent with the role of the vlPFC, Ritchey et al. (2011) have shown that the vlPFC is associated with memory for emotional items that were semantically encoded (elaborate encoding) rather than perceptually encoded (shallow encoding). Interestingly, there is also evidence that the PFC can exert influences on amygdalar activity according to the current goals/focus (Smith, Stephan, Rugg, & Dolan, 2006; see also Cunningham, van Bavel, & Johnsen, 2008; Denkova, Dolcos, & Dolcos, 2013, 2014), which is consistent with recent research highlighting the importance of strategic and motivational factors in the impact of emotion on memory (Harmon-Jones, Gable, & Price, 2012; Kaplan, van Damme, & Levine, 2012; Levine & Edelstein, 2009).
Overall, available research provides evidence that the enhancing effects of emotion involve two mechanisms: direct/bottom-up involving AMY–MTL interactions, and indirect/top-down mechanisms involving modulatory influences from outside of the MTL (Dolcos & Denkova, 2008; Dolcos et al., 2011, 2012; Dolcos, LaBar, et al., 2006; see Figure 1). Bottom-up systems are assumed to be relatively automatic and guided by the stimuli, whereas top-down systems are assumed to be controlled and guided by the current goals.
Impairing Effects of Emotion on Cognition
There is evidence that emotion can not only enhance various cognitive processes (Dolcos et al., 2012; Dolcos et al., 2011), but also impair them (Iordan, Dolcos, & Dolcos, 2013; Johnson et al., 2005; Most, Chun, Widders, & Zald, 2005; see also Oaksford, Grainger, Morris, & Williams, 1996; Seibert & Ellis, 1991; Shackman et al., 2006). As we will discuss next, available evidence shows that emotion can have impairing effects on both lower level (e.g., perceptual) and higher level (e.g., mnemonic and executive) cognitive processes, which also involve contributions from bottom-up and top-down mechanisms.
The Impairing Effect of Emotional Distraction on Perception
The issue of automaticity in emotion processing mentioned previously has also been studied in the context of processing emotional information as task-irrelevant distraction presented in perceptual tasks (Anderson, Christoff, Panitz, et al., 2003; Pessoa, 2005; Pessoa et al., 2002; Vuilleumier, 2005; Vuilleumier et al., 2001; Vuilleumier & Driver, 2007). Unlike the traditional view which states that processing of emotional, especially threatening, information is automatic and does not depend on the availability of processing resources (Vuilleumier et al., 2001), according to a competing view emotion processing depends on attentional resources, which suggests a top-down regulation of emotion processing (Pessoa et al., 2002). Importantly, a recent investigation reconciled these competing views by providing direct behavioral and neuroimaging evidence that these two views are not mutually exclusive (Shafer et al., 2012).
Using a task with emotional distraction, Shafer et al. (2012) showed that processing of emotional distraction is both automatic and modulated by the availability of processing resources, although overall this study provided evidence consistent with the traditional view. This study investigated the impact of emotional distraction by manipulating both the degree of emotional charge of the distracting information and the attentional demands. Behavioral and fMRI data were recorded while participants performed a perceptual orientation discrimination task, involving pictures with various levels of arousal (high emotionally negative, low emotionally negative, neutral, and absolute neutral), which allowed more finer assessment of the emotional charge. The attentional demands were manipulated by varying the time of presentation (short vs. long duration) and task difficulty (low vs. high perceptual load). Behavioral findings revealed impaired performance by emotional distraction reflected in longer response times (RTs) for negative than for neutral items, regardless of manipulations of attentional demands. However, the detrimental effect of emotional distraction was strongest when the difference in emotional content was the greatest (highly emotional vs. absolute neutral), there was more time for distraction (long duration), and the attentional resources were most available (low load), hence suggesting that emotional information is also susceptible to attentional modulation. At the brain level, areas known to be involved in affective processing (including AMY, medial PFC, insula), as well as perceptual areas that are sensitive to affective modulation (FG, lateral occipital cortices) showed greater activations to emotional compared with neutral distracters. Paralleling the behavioral data and consistent with the traditional view, activity in the AMY was found regardless of manipulations of attentional demand. Alternatively, the engagement of higher level (cortical) emotion processing regions (i.e., medial prefrontal cortex [mPFC] and vlPFC) showed susceptibility to modulation by attention with increased engagement when there was more time for distraction and attentional resources were most available. In addition, analyses also dissociated between activity linked to impaired performance by distraction (dorsomedial PFC and vlPFC) versus coping with it (anterior cingulate cortex [ACC]) (Shafer et al., 2012).
Overall, this study highlights the interaction between the emotional charge and attentional resources in understanding the detrimental effects of emotional distraction on perception, and provides direct evidence that the two views concerning basic emotion processing (traditional and competing; compare Vuilleumier et al., 2001 with Pessoa et al., 2002) are not mutually exclusive. These findings show that emotion processing is both automatic and modulated by attention, but emotion and attention were only found to interact when finer assessments of emotional charge (comparison of most vs. least emotional conditions) were considered along with an effective manipulation of processing load (high vs. low). Hence, depending on the circumstances, emotional information can be processed automatically but is also susceptible to modulations linked to the availability of attentional resources (Shafer et al., 2012).
The Impairing Effect of Emotional Distraction on Working Memory
The impairing impact of task-irrelevant emotional distraction on more complex cognitive processes (working memory [WM]) has been also investigated in a series of fMRI studies by us and others (Anticevic, Repovs, & Barch, 2010; Chuah et al., 2010; Denkova et al., 2010; Dolcos, Diaz-Granados, Wang, & McCarthy, 2008; Dolcos, Kragel, Wang, & McCarthy, 2006; Dolcos & McCarthy, 2006; Iordan, Dolcos, Denkova, & Dolcos, 2013, reviewed in Iordan, Dolcos, & Dolcos, 2013). These studies have identified opposite patterns of response in brain regions that are part of a ventral neural system involved in “hot” emotion processing (HotEmo/Ventral Emotional System) versus brain regions that are part of a dorsal system involved in “cold” cognitive control/executive processing (ColdEx/Dorsal Executive System), which show increased activation versus deactivation, respectively, linked to a detrimental impact of emotional distraction (Dolcos & McCarthy, 2006, reviewed in Iordan, Dolcos, & Dolcos, 2013; see Figure 2).
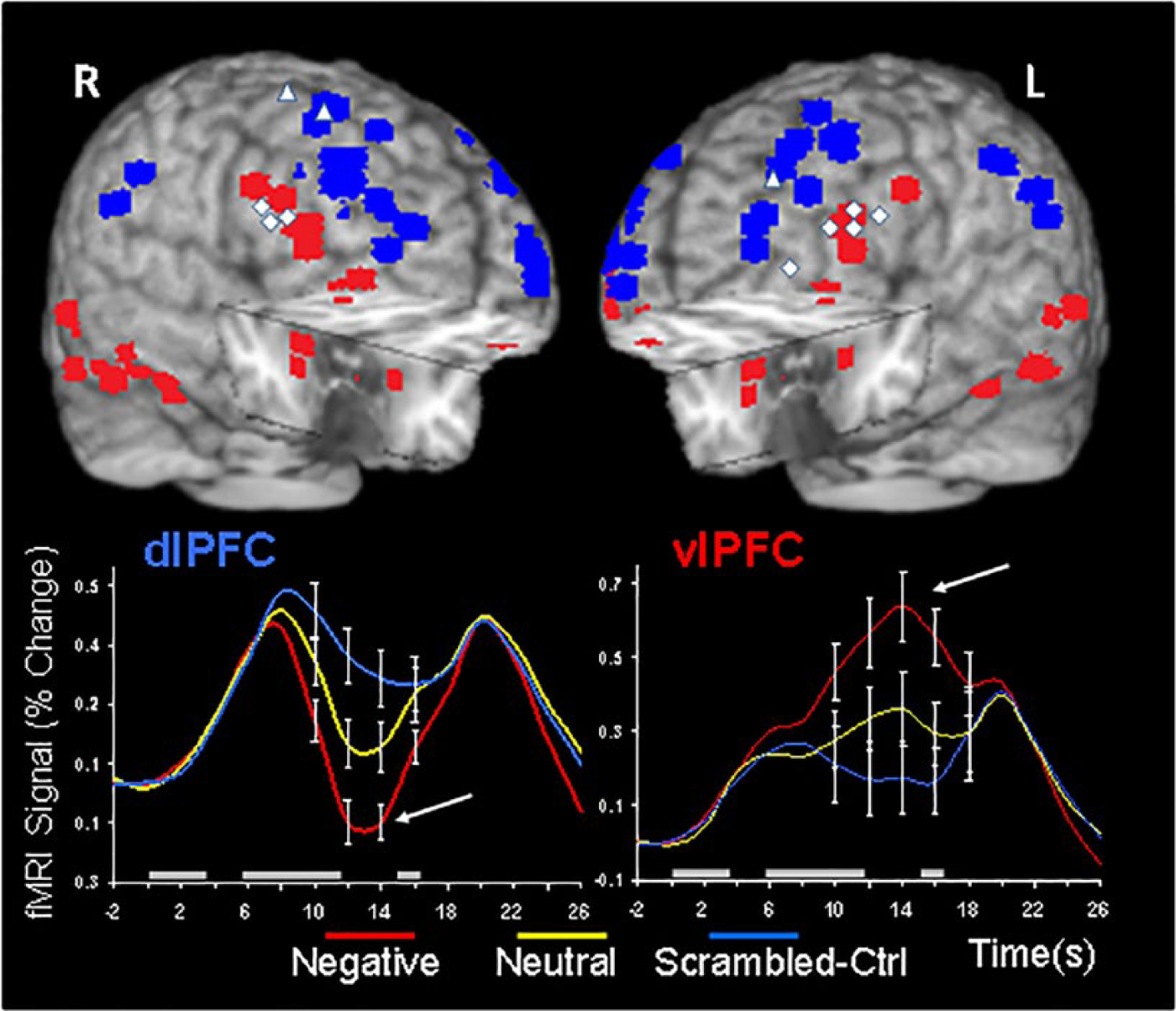
Dissociations between activity in dorsal executive and ventral affective networks. Displayed are peak activation voxels from ventral areas (red) showing increased and dorsal areas (blue) showing decreased activity to negative distraction, as identified by fMRI studies using versions of working memory (WM) tasks with emotional distraction. The line graphs show the time course of activity in typical dorsal (dorsolateral PFC [dlPFC]) and ventral (ventrolateral PFC [vlPFC]) areas. As indicated by the arrows, emotional distraction produced the most disrupting effect on delay activity in dlPFC, while producing the most enhancing effect in vlPFC. Noteworthy, subregional specificity within these larger areas has also been linked to coping with distraction (indicated by the white shapes), which tends to have a more ventral distribution in the lateral PFC. The peak activation voxels are superimposed on a high-resolution brain image displayed in a tridimensional view using MRIcro (Version 1.40 Build 1; Rorden, n.d.). R = right; L = left; Ctrl = control condition. Adapted from review by Iordan, Dolcos, and Dolcos (2013) and Dolcos and McCarthy (2006).
The basic approach in identifying these effects involved recording of brain activity using fMRI, while participants performed a delayed-response WM task with emotional distraction. The WM task involved keeping in mind a set of human faces (memoranda) for the duration of a short delay, and then answering whether a single face (probe) was part of the initial set or not. During the delay interval between the memoranda and the probe, high-arousing stimuli were presented as task-irrelevant distracters. The subjects were instructed to look at the distracters but maintain focus on the memoranda, and to make quick and accurate responses to the probes. Presentation of emotional distracters was associated with increased activity in emotion processing regions (AMY, vlPFC, and mPFC), together with decreased activity in dorsal executive brain regions (dlPFC and the LPC) leading to impaired WM performance. These findings suggest that activity in the affective and executive neural systems is strongly interconnected, such that increased activity in the ventral affective regions disrupts activity in the dorsal executive system and results in cognitive impairment.
Follow-up investigations (Denkova et al., 2010; Dolcos et al., 2008; Dolcos, Miller, Kragel, Jha, & McCarthy, 2007; Iordan, Dolcos, Denkova, et al., 2013) provided additional evidence that these patterns of neural activity are specific to emotional distraction, and further explored the specificity of this response to different types of distracters (reviewed in Iordan, Dolcos, & Dolcos, 2013). In addition, other investigations using similar (Anticevic et al., 2010; Diaz et al., 2011; Oei et al., 2012) or derived different tasks (Mitchell et al., 2008; Wang, McCarthy, Song, & Labar, 2005; Yamasaki, LaBar, & McCarthy, 2002) in healthy participants, and evidence from clinical research (Anticevic, Repovs, Corlett, & Barch, 2011; Morey et al., 2009) also support this ventro-dorsal dissociation in response to emotional distraction, hence highlighting the replicability and generalizability of this dissociation (Iordan et al, 2013).
An important factor influencing the impairing effect of emotion is the capacity to engage coping mechanisms in order to resist emotional distraction. Namely, brain imaging studies in which emotional information was presented as task-irrelevant distraction also provided evidence regarding the neural correlates of coping with distracting emotions, which entails increased activity in, and interactions between, brain regions involved in basic emotion processing (AMY) and cognitive control (PFC) brain regions, such as the vlPFC (Anticevic et al., 2010; Chuah et al., 2010; Denkova et al., 2010; Dolcos et al., 2008; Dolcos, Kragel, et al., 2006; Dolcos & McCarthy, 2006; Iordan, Dolcos, Denkova, et al., 2013). Of note, although the vlPFC shows overall increased activity to emotional distraction consistent with its inclusion in the HotEmo/Ventral Affective System, specific areas within this larger region have proven to be involved in coping with emotional distraction (see Figure 2), consistent with evidence regarding its role in top-down control (Aron, 2007; Aron, Robbins, & Poldrack, 2004). Moreover, the engagement of the vlPFC involves functional coupling with the AMY, which could reflect processing that originates in the AMY that signals the vlPFC through bottom-up mechanisms about the presence of emotional distraction, and thus the need to control its possible detrimental effects on cognitive performance (Dolcos, Kragel, et al., 2006). Overall, available evidence from research investigating the impairing effects of emotion highlights opposing patterns of response in, and interactions between, neural systems sensitive to modulation by emotional distraction. Top-down PFC control mechanisms seem to be engaged in order to counteract the bottom-up influences produced by emotional distraction, cope with its presence, and maintain cognitive performance.
Altogether, the evidence reviewed in first two sections of this article, resulting from studies separately investigating enhancing and impairing effects of emotion on cognition, suggests that emotion can exert beneficial or detrimental effects on cognition. These opposing effects have been linked to the involvement and interactions between neural systems associated with bottom-up and top-down processing. In the investigations of the enhancing effects, the bottom-up and top-down systems have been described as part of direct versus indirect mechanisms, which have joint contribution to the beneficial effect of emotion, whereas in the investigations of the impairing effects, the bottom-up and top-down processing have shown an opposing relationship leading to impairing effects of emotional distraction. Interestingly, at the brain level, the bottom-up and top-down effects seem to map onto similar ventral and dorsal neural systems, respectively, linked to enhancing or impairing effects of emotion. Also, some of the brain regions highlighted before (AMY, lateral PFC) seem to be involved in both effects, but their specific contribution is not clear—that is, is it exactly the same area involved in both effects, or is there subregional specificity within larger brain areas, with some subregions being involved in enhancing and others in impairing effects? Understanding these issues is important in both healthy functioning and disease, where the enhancing and impairing effects of emotion tend to co-occur. However, the link between these effects and the associated neural mechanisms is not clear, as they only recently have started to be investigated together. In the next two sections, we will discuss recent evidence addressing these issues.
Linking Enhancing and Impairing Effects of Emotion in Healthy Functioning
Although previous research has independently investigated the enhancing and impairing effects of emotion on cognition, very little is known about their interactions and the associated neural mechanisms. Concomitant investigation of enhancing and impairing effects of emotion and of their interaction is important because they tend to co-occur, at different levels, both within the same process (EM) and across different processes (Perception-EM, WM-EM). Regarding within-process effects, the same emotionally arousing event can lead to opposite effects on different aspects of memory: for example, hearing a gunshot might enhance memory for central aspects of what was happening at the time, while impairing memory for peripheral details (Christianson, 1992a). This phenomenon has been known as “emotional memory narrowing” (Reisberg & Heuer, 2007), “tunnel memory” (Safer, Christianson, Autry, & Osterlund, 1998), or “central/peripheral trade-off” (Kensinger, Garoff-Eaton, & Schacter, 2007). Similarly, emotion enhances memory for high-priority information, but reduces memory for low-priority information, with priority being assigned either by bottom-up perceptual processes or by top-down processes (Lee, Itti, & Mather, 2012; Lee, Sakaki, Cheng, Velasco, & Mather, 2014; Mather & Sutherland, 2011; Sakaki, Fryer, & Mather, 2014). Regarding the across-processes effects, the mechanisms responsible for the impairing effect of emotional distraction may also contribute to increased memory for the distracters themselves. For instance, seeing the scene of a tragic accident while driving may temporarily distract us from the main task (driving), and at the same time this could also lead to better memory for that particular driving experience. Similarly, in a social context, seeing disapproving expressions on the face of a person in audience during a talk may temporarily distract us and disrupt the flow of our presentation, while also leading to better memory for this particular person’s face compared to those of other people present in the audience. For the latter examples, the opposing effects of emotion could also be viewed in terms of a temporal dissociation between them—that is, immediate (impairing) versus long-term (enhancing) effect of distracting emotional information across different cognitive processes. The focus of this section will be on emerging evidence from studies linking immediate impairing and long-term enhancing effects of emotional distraction, as derived from studies in healthy participants.
Linking Opposing Effects of Emotional Distraction on Perception and Episodic Memory
Within-subject investigations of both immediate and long-term effects of emotional distraction have provided insight into the link between immediate (impairing) and long-term (enhancing) effects of emotion (Shafer & Dolcos, 2012; Dolcos et al., 2013). For instance, the aforementioned study by Shafer et al. (2012), in addition to manipulating the degree of emotional charge and attentional demands also included a surprise memory task for the emotional and neutral distracters themselves, while event-related fMRI data were recorded. By examining how emotionally distracting information at perception influences later memory enhancement, this study allowed identification of common and dissociable neural correlates associated with these opposing effects of emotional distraction on perception and memory encoding. As discussed before, task-irrelevant emotional information presented concurrently with a perceptual task impaired performance (reflected in longer RTs), especially in conditions when processing resources were most available. However, those conditions associated with the greatest immediate impairment in perceptual task did not lead to similarly strong enhancement of EM for the distracters. Instead, enhanced EM for distracters was strongest when processing resources were least available, which therefore provides a direct link between impairing effects of emotion on perception and enhancing effects of emotion on memory (see Figure 3A). In other words, a direct relationship between the impairing and enhancing effects was observed only under conditions of limited processing resources. At the brain level, this direct relationship was reflected in overlapping activations in ventral brain regions, including the AMY and HC, thus suggesting that the link between these effects involves bottom-up mechanisms (see Figure 3B).
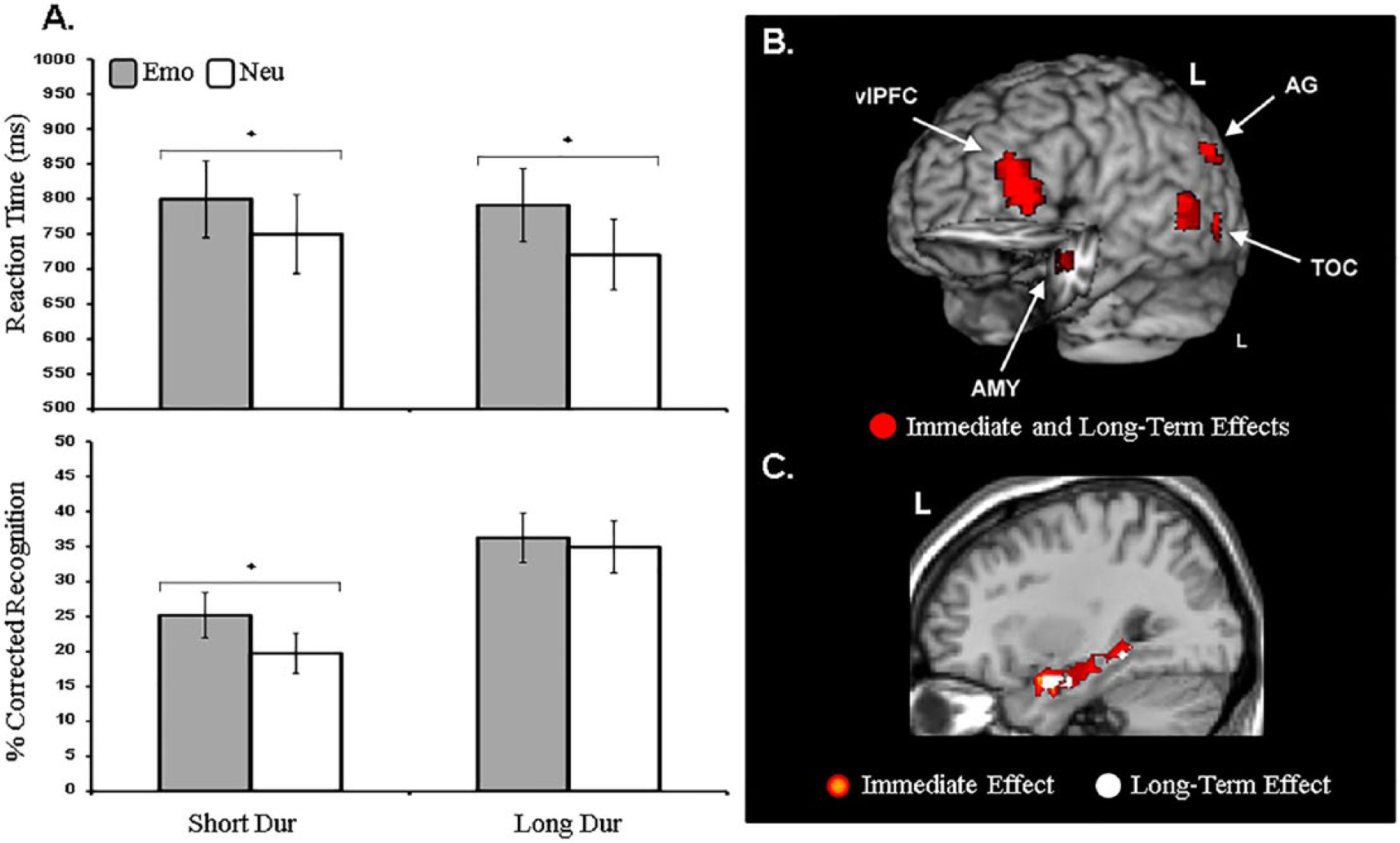
Direct relationship between impaired perceptual processing and enhanced episodic memory (EM), under limited resource availability. Behavioral data show item categories meeting both criteria—impaired perception (top bar graphs) and enhanced EM (bottom bar graphs) under condition of limited resources (short duration; ShortDur = short duration) (A). Neuroimaging data show common regions of response to the impairing effect of emotional distraction and the enhancing effect of emotional memory (B) and overlapping areas between the immediate and long-term effects in the left MTL (C). Emo = emotional; Neu = neutral; L = left hemisphere, vlPFC = ventrolateral prefrontal cortex, AG = angular gyrus, TOC = temporo-occipital cortices.
It should also be noted that, in addition to common mechanisms linking the immediate and long-term effects of emotional distraction, responses in specific brain regions also differentiated between these opposing immediate and long-term effects of emotion, both in terms of overall activation and covariation with performance. Specifically, activity in the medial frontal, precentral, superior temporal, and medial occipital gyri were specifically linked to the immediate impact (increased distraction by emotional information), whereas activity in the superior parietal cortex was specifically linked to the long-term impact (better memory for emotional distracters; Shafer & Dolcos, 2012).
Overall, these findings provided initial evidence that the link between immediate/impairing impact of emotional distracters during a perceptual task and their long-term enhancing effect is mediated by bottom-up direct mechanisms involving AMY and HC. In addition, dissociable brain mechanisms linked to these opposing effects were also identified.
Linking Opposing Effects of Emotional Distraction on Working Memory and Episodic Memory
The link between immediate and long-term effects of emotional distraction has also been investigated with tasks tapping into more complex cognitive processes, such as WM (Dolcos et al., 2013). Similar to investigating the link between opposing effects of emotional distraction on perception and EM (Shafer & Dolcos, 2012), the study by Dolcos et al. (2013) identified the neural correlates for trials meeting both criteria—impaired WM and enhanced EM. Interestingly, again, this study identified a role of bottom-up MTL mechanisms in linking the immediate impact of emotion on WM and the long-term enhancement of EM. In addition, it also identifies contributions from top-down PFC-based mechanisms. Notably, AMY has a central role in both effects, but its involvement is associated with opposing activity in other brain regions, linked to the direction of immediate (impairing) versus long-term (enhancing) impact of emotional distraction on WM and EM—that is, dlPFC deactivation coupled with HC activation, respectively (see Figure 4).
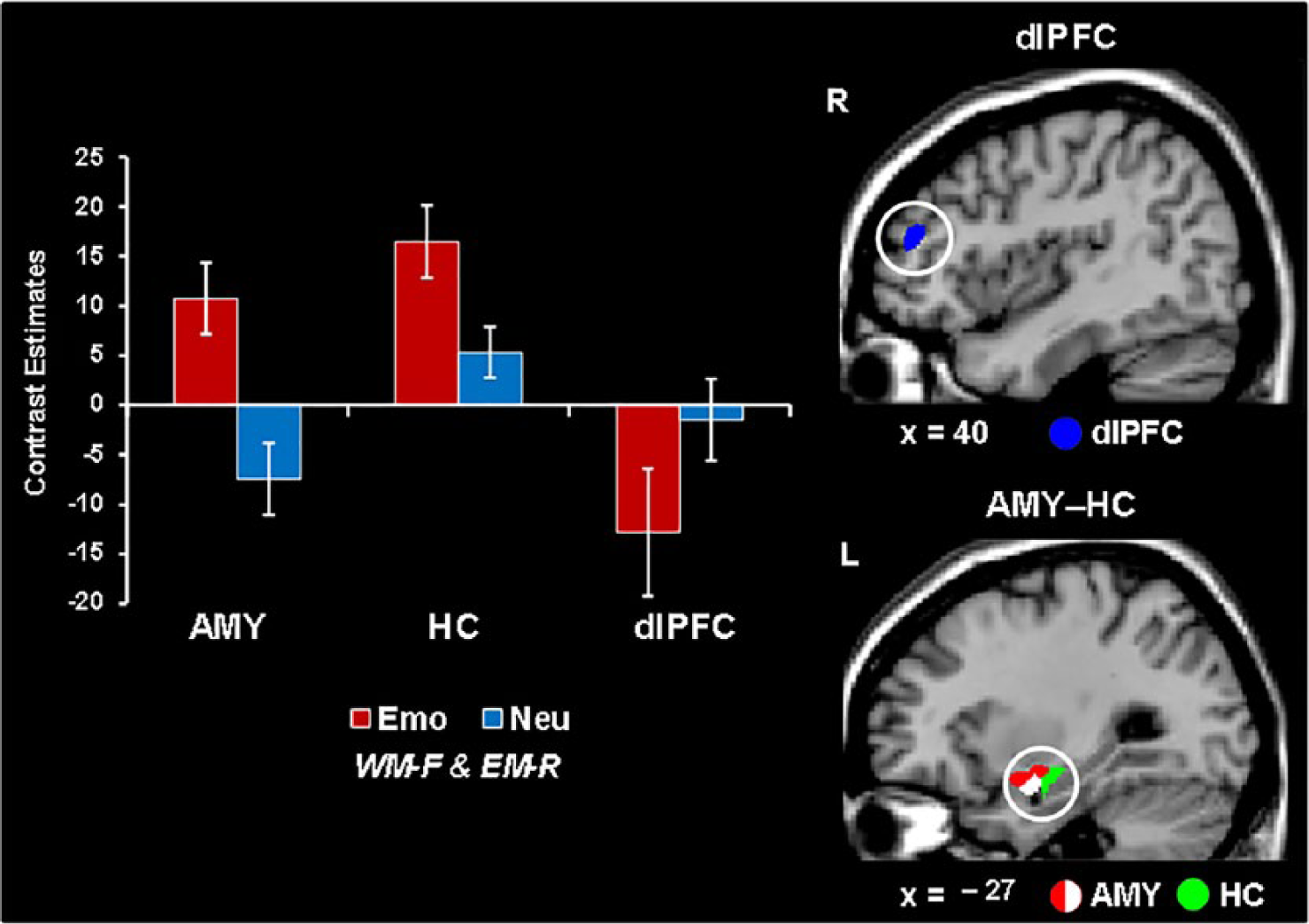
Opposing patterns of activity in AMY and HC versus PFC linked to working memory (WM) impairment and episodic memory (EM) enhancement by emotional distraction. Increased activity in both amygdala (AMY, red blob) and hippocampus (HC, green blob) and greater deactivation in the dorsolateral prefrontal cortex (dlPFC, blue blob) were observed in response to emotional compared to neutral distracters associated with impaired WM performance (WM-F) and later enhanced EM (EM-R). A negative correlation was also identified between left AMY activity and WM performance to emotional distracters (white blob). The bar graphs show the contrast estimates, as extracted from peak voxels of the targeted regions (AMY, HC, and dlPFC). WM-F = trials that were forgotten in the WM task, EM-R = trials that were remembered in the EM task; R = right, L = left. From Dolcos et al. (2013).
Possibly as a result of activating mechanisms signaling potential danger, processing of task-irrelevant negative distraction diverts processing resources from the main WM task to processing emotional distracters, which in turn lead to dlPFC deactivation (Dolcos & McCarthy, 2006), while simultaneously initiating processing that leads to enhanced EM for the distracting stimuli, via increased activation in an MTL-dependent route. This explanation is further supported by the finding of a negative relationship between AMY activity and WM performance, which suggests a bottom-up effect of emotional distraction, in which increased AMY activity in the presence of emotional distraction is associated with lower WM performance and increased EM.
Intriguingly, trials producing enhanced EM performance without disrupting WM performance were associated with increased involvement of top-down PFC mechanisms (i.e., right vlPFC; see Figure 5). This points to the engagement of emotion regulation (ER) strategies to cope with emotional distraction during WM, which then contributed to subsequent better EM for the distracters themselves, probably due to increased strategic influences on stimulus elaboration linked to a deeper level of processing (Dillon, Ritchey, Johnson, & LaBar, 2007). Related to this, studies investigating the impact of ER on memory provided evidence that some ER strategies can enhance memory, while other ER strategies impair memory (Dillon et al., 2007; Richards & Gross, 2000). Particularly, while reappraisal is associated with enhanced memory, suppression is associated with impaired memory and these dissociable effects could be linked to different levels of processing: deeper versus shallow, respectively. Interestingly, at the neural level, the beneficial effects of reappraisal on memory have been mediated by the involvement of vlPFC and coactivations between vlPFC and HC and AMY (Hayes et al., 2010), and interpreted in terms of a deeper level of processing leading to better memory through the involvement of top-down PFC mechanisms. Of note, as mentioned in the first section of this article (Enhancing Effects of Emotion on Cognition), deeper encoding of emotional information has been associated with the involvement of vlPFC and enhanced EM (Ritchey et al., 2011).
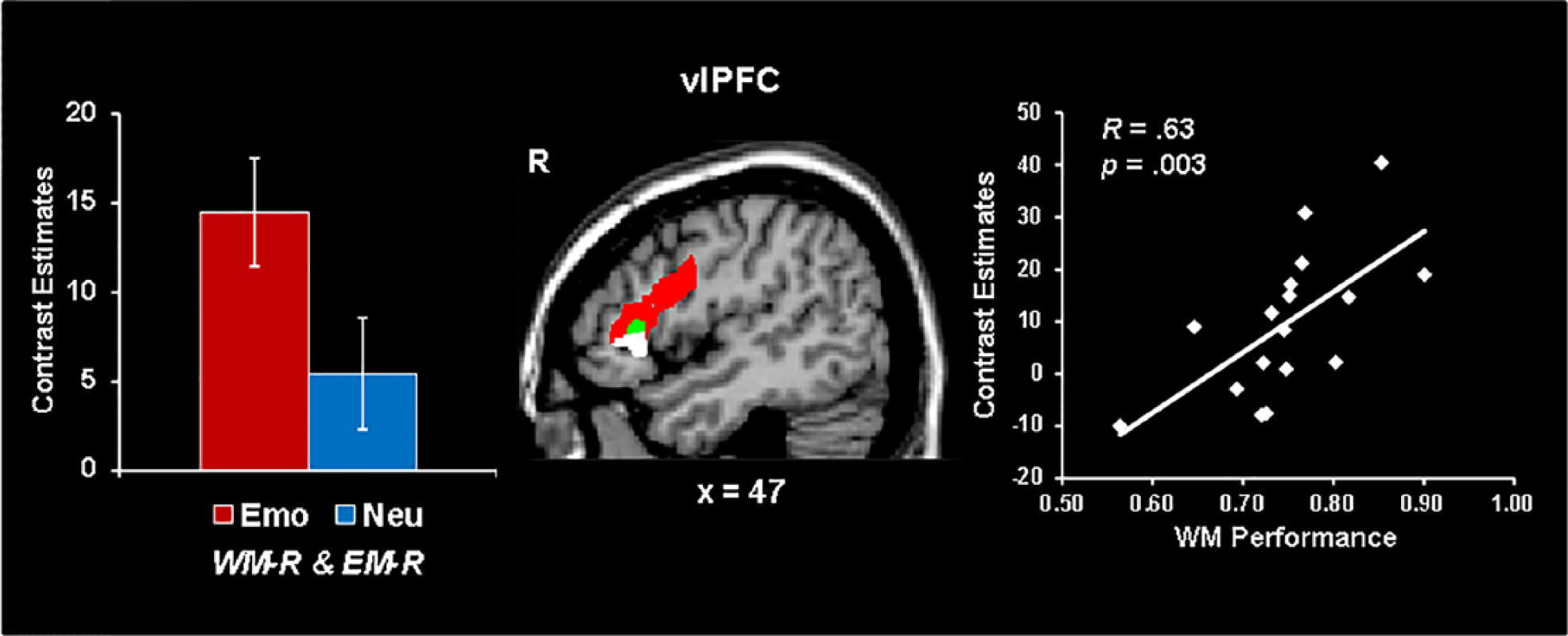
Increased right vlPFC activity linked to coping with emotional distraction during WM and later EM enhancement. Right ventrolateral prefrontal cortex (vlPFC) showed increased activity to emotional distracters associated with working memory success (WM-R) and episodic memory enhancement (EM-R), compared to both neutral distracters associated with WM success (red blob) and emotional distracters associated with WM impairment (green blob). In addition, a positive correlation was identified between activity in the right vlPFC area and WM performance for emotional distraction (white blob). The bar graph shows contrast estimates for the peak voxel in vlPFC for the comparison between emotional and neutral stimuli associated with WM success and enhanced EM. Error bars represent standard errors of means. The scatter plot shows the covariation between brain activity and WM performance, based on signal extracted from the white blob. WM-R = trials that were remembered in the WM task, EM-R = trials that were remembered in the EM task; R = right; Emo = emotional; Neu = neutral. From Dolcos et al. (2013).
Collectively, these findings demonstrate that the immediate impairing impact of emotional distraction on perception or WM and the long-term enhancing impact of emotion on EM are mediated by overlapping and dissociable neural systems, involving both bottom-up and top-down mechanisms. Specifically, bottom-up activity in MTL regions was common in linking opposing immediate/impairing and the long-term/enhancing effects in both perceptual and WM tasks with emotional distraction. In addition, in the case of the WM-EM task, there is also evidence for the involvement of PFC top-down mechanisms to reduce the impact of distraction, which also contributes to better EM, very likely as a result of deeper encoding due to more elaborate semantic processing of the distracters. Overall, linking enhancing and impairing effects has the potential to further our understanding of the mechanisms underlying the impact of emotion on cognition not only in healthy functioning, but also in affective disorders, where these effects are exaggerated and detrimental to mental well-being. As we will see in the next section, the link between enhancing and impairing effects in the case of PTSD, unlike healthy functioning, appears to occur in the opposite direction: increased remembering of traumatic memories, as internally generated distraction, may interfere with ongoing cognitive processing. Moreover, both effects seem to have a common cause, linked to the initial exposure to the traumatic event.
Linking Enhancing and Impairing Effects of Emotion on Cognition in Affective Disorders
The co-occurrence of enhancing and impairing effects of emotion is probably most evident in affective disorders, where both of these opposing effects are exacerbated. Particularly, affective disorders, such as depression and posttraumatic stress disorder (PTSD), are characterized by increased susceptibility to emotional distraction, either internal or external, and impaired cognitive control (Brown & Morey, 2012; Hayes, van Elzakker, & Shin, 2012; Rauch, Shin, & Phelps, 2006; Shin & Liberzon, 2009). Increased responsiveness to negative material can impair attention, memory, and cognitive control in depression and PTSD and this is typically associated with altered activity in emotion-related brain regions (AMY, rostral/ventral anterior cingulate cortex [rACC/vACC]) and cognitive control brain regions (PFC, dorsal ACC; Dolcos, 2013; Foland-Ross & Gotlib, 2012; Hayes et al., 2012). The focus here is on evidence from studies of PTSD, pointing to the importance of investigating both enhancing and impairing effects of emotion in elucidating the nature of alterations in the way emotion interacts with cognition in clinical conditions.
Of particular relevance is emerging evidence concerning the neural correlates of alterations associated with the encoding of emotional memories (Hayes et al., 2011) and with the responses to task-irrelevant emotional distraction (Morey et al., 2009). These changes are reflected in regions associated with functions that may be enhanced (EM) or impaired (WM) by emotion—that is, the MTL and dlPFC, respectively. One the one hand, recent findings from an EM fMRI study in PTSD patients showed reduced memory-related activity in the AMY-MTL system during memory encoding, and higher false alarm rates during retrieval, compared to trauma-exposed control (TEC) participants (Hayes et al., 2011). This provided evidence for nonspecific (gist-like) encoding of trauma-related material linked to disorganization of the MTL encoding mechanisms. One the other hand, findings from a WM fMRI study showed that the PTSD group had greater trauma-specific activation than the control TEC group in main emotion processing brain regions, including the AMY and vlPFC, as well as in brain regions susceptible to emotion modulation (e.g., FG; Morey et al., 2009). Importantly though, the PTSD group also showed greater nonspecific disruption of activity to combat-related and neutral task-irrelevant distracters in brain regions that subserve the ability to maintain focus on goal-relevant information, including the dlPFC. The undifferentiated dlPFC response to combat and noncombat distracters in PTSD is consistent with the hypervigilance hypothesis that may explain enhanced response to, and potentially distracting effect of seemingly neutral stimuli (see Figure 6).
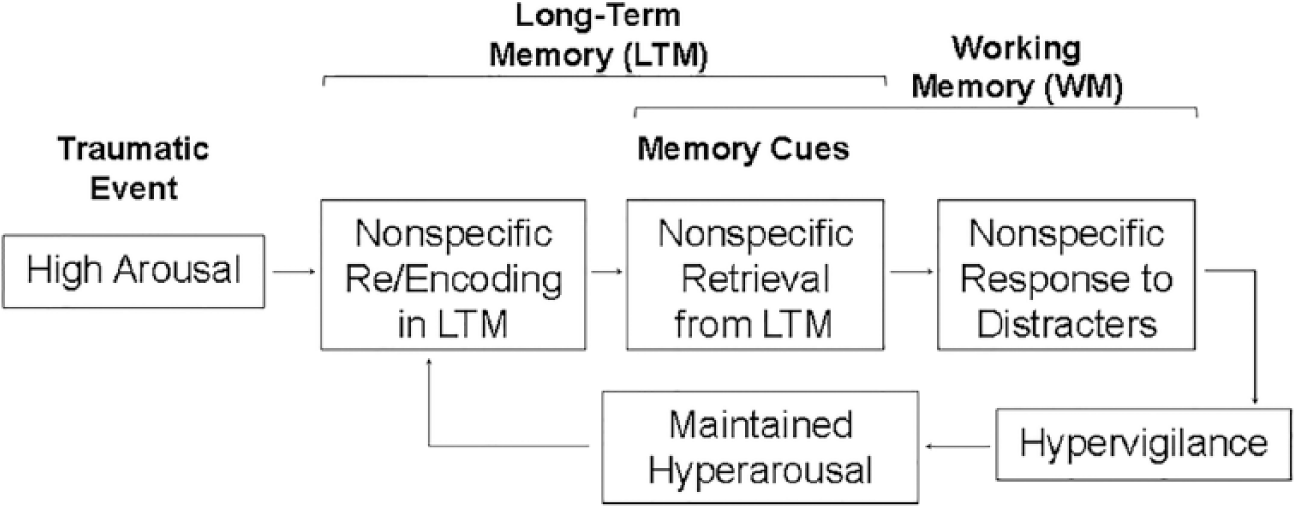
Diagram illustrating possible link between the impact of emotion on long-term memory and working memory in PTSD. From Dolcos (2013).
The evidence from separate lines of investigations discussed before, regarding the neural changes in PTSD linked to dysfunctions in the recollection of traumatic events and the response to emotional distraction, converge toward the idea that nonspecific response to emotional and neutral distraction may reflect retrieval distortions linked to inefficient initial encoding of trauma-related information (Dolcos, 2013). Namely, it is possible that the nonspecific disruption of the dlPFC activity by trauma-related and neutral distraction is linked to the retrieval of the traumatic memories triggered by nonspecific cues, which may also contribute to the perpetuation of the state of hyperarousal observed in these patients. Moreover, it is also possible that the source of these effects may be linked to elevated arousal during the initial exposure to traumatic events (Dolcos, 2013).
In summary, available evidence from investigations of PTSD patients points to general and specific emotional and cognitive disturbances that are linked to alteration in the neural circuitry underlying emotion–cognition interactions. This evidence suggests that reduction of AMY and HC signals for trauma-related cues may underlie nonspecific encoding of gist-based representations instead of specific and detailed contextual details of the trauma-related memories. This, in turn, may be linked to symptoms of hypervigilance and nonspecific responses to trauma-related distraction, which contributes to the maintenance of a hyperarousal state.
Conclusions and Future Directions
The overarching goal of the present review was to discuss findings from investigations examining enhancing and impairing effects of emotion and linking them in healthy functioning and clinical conditions. While much of the available evidence is coming from studies separately investigating enhancing and impairing effects, recent progress has been made in identifying the underlying neural mechanisms linking these effects. Emerging evidence from studies linking enhancing and impairing effects demonstrates that the immediate impairing impact of emotional distraction on perception or WM and the long-term enhancing impact of emotion on EM are mediated by overlapping and dissociable neural systems, involving both ventral/bottom-up and dorsal/top-down mechanisms. Moreover, these mechanisms appear altered in the case of affective disorders, such as PTSD, where it is possible that the defective long-term representations of the traumatic events can act as powerful internal distracters, which may impair current activities due to altered cognitive control. Interestingly, both effects seem to have a common cause in PTSD, linked to the initial exposure to the traumatic event. Overall, the present review highlights the need to consider and link together the enhancing and impairing effects of emotion and their immediate and long-term impact, in investigations of emotion–cognition interactions.
Despite significant progress in clarifying the mechanisms underlying opposing effects of emotion, several issues still need clarification in future research. (a) First, the link between these opposing effects has been investigated mainly using external emotional distracters (i.e., pictures depicting emotional scenes, faces with emotion expressions), and hence the impact of internal emotional distraction (i.e., recollection of personal memories; Young, Erickson, & Drevets, 2012) remains unclear. Investigation of this issue has relevance for understanding depression and PTSD, where ruminations on negative memories (Brewin, Reynolds, & Tata, 1999; Rubin, Berntsen, & Bohni, 2008) lead to an overall long-lasting negative state, which in turn impairs cognition and contributes to the maintenance of the clinical disorders. (b) Second, while there is evidence suggesting the engagement, very likely spontaneous, of coping mechanisms to deal with distraction, it is unclear how instructed engagement in various cognitive control strategies affects the impact of emotional distraction, and the underlying neural circuitry. Specifically, it remains unclear how instructed use of different coping strategies (e.g., reappraisal, suppression, attentional deployment, mindfullness) could influence the immediate and long-term effects of emotional distraction on cognition and the neural activity in bottom-up/hot/affective and top-down/cold/executive systems. Investigation of these issues has relevance for understanding reciprocal relationships between emotion and cognition in both healthy and clinical populations. (c) Third, while the influence of attentional resources on the impact of emotion on lower level perceptual processes has been investigated, it is unknown how manipulation of processing resources modulates the impact of emotional distraction on higher level cognitive processes. Based on evidence that variation in cognitive load may affect perceptual processing, it is expected that various levels of attentional demand can have differential impact also in the case of more complex cognitive processes (e.g., WM). Taking the example of the conference talk and the disapproving face of seeing person from the audience, it is possible that the impact of seeing disapproving faces (emotional distracters) on the ability to deliver the talk and later remembering those faces could be different depending on whether the talk is simple and overrehearsed (low load) or complex and presented for first time (high load).
Investigation and clarification of these issues is important for understanding mechanisms of emotion–cognition interactions not only in healthy functioning, but also in emotional disturbances, where the opposite effects of emotion are exacerbated and have detrimental consequences on everyday life functioning and well-being.
Footnotes
Author note:
During the writing of this review, Florin Dolcos was supported by the University of Illinois. The authors wish to thank Dr. Sanda Dolcos, Alex Iordan, and Andrea Shafer for comments on previous versions of the manuscript.