
Abstract
Neuroscience investigations of emotional influences on social cognition have been dominated by the somatic marker hypothesis and dual-process theories. Taken together, these lines of inquiry have not provided strong evidence that emotional influences on social cognition rely on neural systems which code for bodily signals of arousal nor distinguish emotional reasoning from other modes of reasoning. Recent findings raise the possibility that emotionally influenced social cognition relies on two stages of neural changes: once when emotion is elicited and a different set of changes at the time of social cognitive judgment. These findings suggest that affect infusion models may be a fruitful framework for bridging neuroscience and psychological understanding of the role of emotion in social cognition.
One of the early promises of social neuroscience and neuroeconomics was that inquires at the neural level of analysis might inform theories that explain the behavioral level of analysis (e.g., Beer, Mitchell, & Ochsner, 2006). Has this promise paid off for researchers interested in understanding how emotion influences, that is, motivates social cognition? This review considers an answer to this question by examining relevant neuroscience research. Whereas dual-process theories and a critical role of physiological arousal are not well supported at the neural level, a new empirical approach has generated findings that suggest a fruitful step forward will further draw on this approach and hypotheses generated by affect infusion models. It is important to note that many lines of research have looked at the neural underpinnings of emotion–cognition interactions but only a few have had the aim of understanding the role of emotion in social cognition. Inquires of this nature are important for understanding the parallels and differences between social-cognitive and nonsocial cognitive processing. For example, many studies have examined how experimentally manipulated emotion or individual differences in emotional deficits (e.g., patients with depression or anxiety) might change the neural processing required for executive functions (e.g., the emotional Stroop vs. the classic Stroop task; Okon-Singer, Hendler, Pessoa, & Shackman, 2015). These studies have laid important foundations for understanding how disorders such as depression or anxiety may impact neural processing.
In contrast, research on the somatic marker hypothesis and research on the neural basis of dual-process theories have more specifically tried to understand the role of emotion in social cognition (see Table 1). Although the processing underlying social cognition may or may not be distinct from non-social-cognitive processing, social cognition is distinct because it includes cognitive processes that emerge from interpersonal contexts, imagined or tangible. Currently, these lines of inquiry have been helpful in ruling out the theory that the physiological arousal associated with emotion is critical for optimizing decisions. Additionally, there is not strong evidence to substantiate the dual-process theory that an emotional mode of reasoning is somehow distinct at the neural level of analyses from other modes of reasoning. However, some initial studies that have not been conducted under the theoretical umbrellas of the somatic marker hypothesis or dual-process theories suggest possible support for affect infusion models. Specifically, at least one of these studies has shown that emotional influences on social cognition reflect two phases: changes that occur at the time of emotion elicitation and changes that occur at the time of the social cognitive process (Bhanji & Beer, 2012b). These lines of research are reviewed in what follows and are followed by recommendations on how future neuroscience research will need to depart from current empirical approaches in order to contribute to psychological theories of emotional influences on social cognition.
Neuroscience investigations into the role of emotion in social cognition.
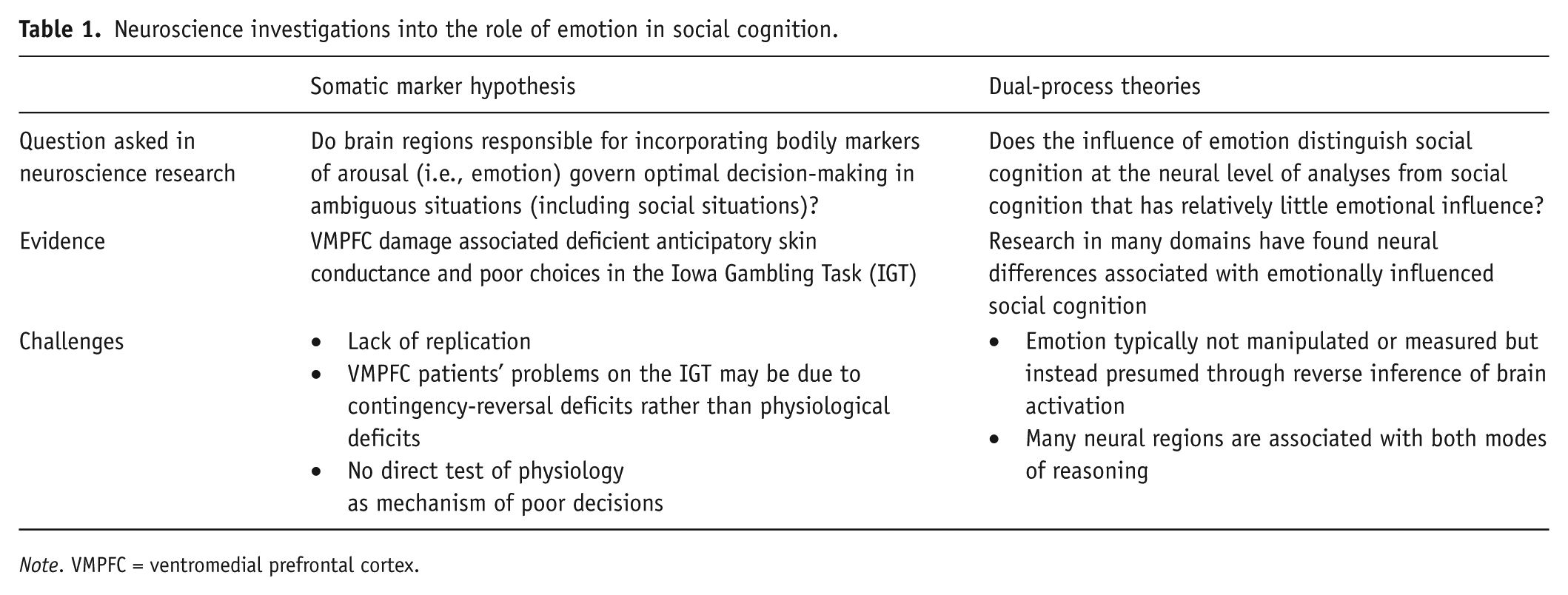
Note. VMPFC = ventromedial prefrontal cortex.
The Somatic Marker Hypothesis
Research on the somatic marker hypothesis opened the door for neuroscientists to study the role of emotion in social cognition (e.g., Bechara, Damasio, & Damasio, 2000). The somatic marker hypothesis proposed that optimal decisions in complex situations (including social situations) are guided by the integration of bodily markers of emotional arousal. In other words, wise decisions rely on emotional information (in this case, emotion is equated with physiological arousal) elicited by the situation. This integration and evaluation was theorized to take place in the ventromedial prefrontal cortex (VMPFC).
Evidence in Support of the Somatic Marker Hypothesis
The classic experiments testing the somatic marker hypothesis focused on the ability of patients with circumscribed brain damage to perform the Iowa Gambling Task (Bechara, Damasio, Tranel, & Damasio, 1997). The Iowa Gambling Task tests anticipatory bodily signals of arousal and risky decision. Participants are required to choose cards from four decks. Cards from each deck indicate whether the participant has won or lost money for that draw. Successful task performance is measured by the extent to which participants figure out which decks they should focus on to maximize their gains. Healthy participants typically figure out the optimal decks after an initial period of exploration (e.g., 25–50 trials). After the learning period, healthy participants tend to favor the optimal decks with the occasional draw from risky decks (those that have some large gains but also many large losses). In healthy participants, the draws from the risky decks are preceded by an increased skin conductance response (i.e., body markers of emotional arousal; Bechara et al., 2000).
In contrast, patients with VMPFC damage do not learn which decks to focus on to maximize gains nor do they show anticipatory arousal when making risky decisions. However, patients with VMPFC damage do show an increased skin conductance response once they learned they had lost on a particular trial (Bechara, Damasio, Damasio, & Lee, 1999). Therefore, these patients were considered to have impaired anticipatory responses to risk rather than a general deficit in arousal (e.g., Bechara et al., 2000). This pattern of findings led to the conclusion that anticipatory somatic markers optimized decisions in complex situations and the use of these anticipatory somatic markers relied on VMPFC function.
Challenges to the Somatic Marker Hypothesis
In recent years, a number of challenges have arisen for the somatic marker hypothesis (Dunn, Dalgleish, & Lawrence, 2006) and some of these challenges undermine the importance of emotional arousal for social cognition. Access to patients with circumscribed brain lesions tends to be limited (Beer, 2015) so the small samples in the original studies have led to questions about generalizability. The limited access to patients also means that there have been few opportunities for replication by independent labs. The clearest independent replication effort found that VMPFC patients’ poor performance on the Iowa Gambling Task may be largely driven by difficulty in contingency-reversal rather than a lack of anticipatory emotional arousal itself (Fellows & Farah, 2005). In the classic version of the Iowa Gambling Task, there is no counterbalancing of card positions within deck. The initial cards in the disadvantageous decks are actually dominated by gains (Dunn et al., 2006). Patients with VMPFC do not show poor performance on the Iowa Gambling Task once card position is counterbalanced such that there is no need to “unlearn” or reverse an initially learned contingency (Fellows & Farah, 2005). Additionally, studies of patients that do not have access to the afferent feedback of emotional arousal (i.e., have spinal cord injuries) find that these patients perform well on the Iowa Gambling Task (North & O’Carroll, 2001). If the processing of bodily markers of emotion was critical for making good decisions, it is difficult to understand why performance would not be impaired. Finally, researchers have pointed out that there is not strong evidence that the poor anticipatory arousal demonstrated in the VMPFC patients actually predicts their poor decisions (Dunn et al., 2006). In other words, physiological arousal is never tested as a mediator or even correlated with measures of optimal decision-making in this body of research making it difficult to strongly conclude that emotional arousal is key for optimal decision-making.
In summary, research on the somatic marker hypothesis was really the first systematic attempt in neuroscience to understand the relation between emotion and social cognition. This perspective defined emotion in terms of physiological arousal and focused on the importance of neural coding of physiological arousal as a mechanism for making optimal decisions. The VMPFC was identified as the brain region important for using physiological feedback to guide decision-making. However, a large body of subsequent research has called into question the importance of neural coding of physiological arousal as the means through which emotion affects social cognition.
Dual-Processes and Social Cognition
Neural research aimed at understanding the role of emotion in social cognition moved forward from the somatic marker hypothesis by focusing on the dual-process theories from the fields of social psychology and judgment and decision-making. There are several variations of dual-process theories but they generally posit that there are two modes of reasoning that shape judgments and decisions (e.g., Sloman, 1996; Tversky & Kahneman, 1974). One mode of reasoning is sometimes described as relatively primitive and characterized by automatic, effortless, fast, and associative processes. These processes tend to operate on particular types of content including emotion. In contrast, the other mode of reasoning is characterized by controlled, effortful, slow, and rule-based processing. These processes tend to operate on particular types of content including emotionally neutral information. Social neuroscience and neuroeconomics became interested in understanding whether the two modes of reasoning were truly distinct even at lower levels of analyses (e.g., Beer & Ochsner, 2006; Sanfey & Chang, 2008). More specifically, researchers wanted to test whether emotion-driven reasoning was associated with the recruitment of a distinct cognitive processing system.
Neural Evidence for Dual-Process Theories in the Realm of Social Cognition
If one were to look for examples where social cognition driven by emotion differs from social cognition less influenced by emotion at the neural level of analyses, there are many to draw upon. Providing an exhaustive review of this literature is beyond the scope of this article. Some examples are provided in the following lines to give the reader a flavor for the kind of findings that demonstrate a neural difference between social cognition presumed to be driven by emotion and social cognition presumed to be less influenced by emotion. It is important to note that examples can be found in a wide variety of domains. Differential neural activation is illustrated here in the domains of economic decision-making, moral reasoning, and self-evaluation. Although economic decision-making may at first seem controversial to include as an example of social cognition, it is mentioned because the meaning of money is dependent on a social context. On a deserted island, a dollar bill cannot be exchanged for goods.
Economic decision-making
Many of the early claims supporting the hypothesis that emotion-driven social cognition differs from social cognition less influenced by emotion comes from research on economic decision making (Sanfey & Chang, 2008). Sometimes the economic decisions are made in the context of an interpersonal interaction (e.g., the Ultimatum Game; Feng, Luo, & Krueger, 2015; Mehta & Beer, 2010; Sanfey, Rilling, Aronson, Nystrom, & Cohen, 2003) and other times participants are simply asked to make a series of decisions about different gambles (e.g., framing effects in gambling choices; DeMartino, Kumaran, Seymour, & Dolan, 2006). Taken together, this research suggests that regions such as the insula and amygdala are involved in economic decisions that are driven by emotion whereas regions like dorsolateral prefrontal cortex (DLPFC), anterior cingulate cortex (ACC), and ventromedial prefrontal cortex (VMPFC) are associated with decisions less influenced by emotion.
For example, a seminal study employed the Ultimatum Game to test whether decisions about unfair offers driven by emotion (i.e., rejection) differed neurally from decisions about unfair offers driven by mathematical calculation (i.e., acceptance; Sanfey et al., 2003). In the Ultimatum Game, participants are presented with an offer of how to divide a pool of money (e.g., $5:$5 split of a $10 pool). Participants’ task is to decide whether to accept or reject the offer. Accepted offers result in participants adding their portion of the pool to their bank whereas the proposer gets to keep whatever they did not offer to give to the participant. However, if the participant rejects the offer, neither the participant nor the proposer gets any of the pool. From a pure gains standpoint, it stands to reason to accept any offer as that is the only way to profit from the offer. However, participants are theorized to react with anger or sadness when they are offered less than a reasonable share of the pool of money (e.g., offered $1 of a $10 pool) and have high rejection rates in those cases. Participants made these decisions while in a scanner and the results showed that insula activation tracked the extent to which an offer was unfair and predicted the decision to reject. In this study and subsequent studies using the Ultimatum Game, regions such as VMPFC, ACC, and DLPFC have been associated with making decisions about unfair offers that are less influenced by emotional reactions (e.g., Feng et al., 2015; Mehta & Beer, 2010).
Other studies have investigated gambling and also find that less emotional decision-making relies on VMPFC activation whereas emotion-driven gambling choices are associated with amygdala activation (DeMartino et al., 2006). For example, participants were asked to make a series of forced choices in a gambling task. The forced choices always included one option to gamble. The second option was either a guaranteed loss or a guarantee to keep part of the current bank. Despite the economic equivalence of these two guaranteed options (e.g., when starting with a bank of 50, a guaranteed loss of 20 is the same as a guarantee to keep 30), participants were much more likely to choose the gamble option when the nongamble option was framed as a guaranteed loss. This decision pattern is interpreted as driven by fear of loss rather than a mathematical calculation of the economic consequence (Kahneman & Frederick, 2006). Amygdala activation was associated with the emotion-driven tendency to choose to gamble more often in the face of a guaranteed loss (DeMartino et al., 2006). However, VMPFC activation was associated with the extent to which participants made choices based on economic gain regardless of framing. As these examples illustrate, research on a wide variety of economic decision-making finds that emotion-driven choices are associated with neural activation (e.g., insula, amygdala) that is distinct from choices less influenced by emotion.
Moral reasoning
Another domain in which scientists have taken a dual-process view of the neural architecture of social cognition is moral reasoning (e.g., Greene, Nystrom, Engell, Darley, & Cohen, 2004; Greene, Somerville, Nystrom, Darley, & Cohen, 2001; Koenigs et al., 2007; Shenhav & Greene, 2014). In this domain, emotionally driven decisions tend to be associated with VMPFC activation whereas ACC and DLPFC activation are associated with decisions less influenced by emotion. For example, the seminal studies asked participants to make decisions about how to resolve moral dilemmas (e.g., Greene et al., 2004; Greene et al., 2001) while their neural activation was measured using fMRI. The dilemmas could be resolved by taking either a deontological or utilitarian standpoint. For example, participants might be asked if it is appropriate to pull a switch to reroute a runaway trolley. Pulling the switch would save the lives of five people stuck on the track. However, it would reroute the trolley such that one person on an alternative track would be killed. The decision to pull the switch is considered utilitarian, that is, a decision to promote the greater good even if there is some collateral damage along the way. In contrast, the decision to not personally interfere reflects a deontological orientation which is presumed to be driven by emotion. The revulsion of personally causing someone’s death shapes the decision to not act in this situation.
The extent to which participants make deontological decisions is associated with VMPFC activation (Greene et al., 2001) and increases VMPFC–amygdala functional connectivity (Shenhav & Greene, 2014). Consistent with the theorized importance of VMPFC for making emotional-driven responses, VMPFC damage decreases the likelihood of deontological responses (Koenigs et al., 2007). In contrast, utilitarian decisions theorized to involve the suppression of any emotional influences tend to be associated with activation in the ACC and DLPFC (Greene et al., 2004). Taken together, research on moral reasoning suggests that emotion-driven moral reasoning is associated with distinct neural processing. VMPFC activation is associated with emotion-driven moral reasoning whereas moral reasoning that is less influenced by emotion is associated with DLPFC and ACC activation.
Self-evaluation
A third illustrative domain which raises the possibility that emotion changes the neural underpinnings of social cognition is self-evaluation (Beer, 2014; Flagan & Beer, 2013). One of the most robust effects in self-evaluation is that people tend to evaluate themselves in a positive manner more so than is warranted by objective measures (Colvin, Block, & Funder, 1995; Sedikides & Gregg, 2008). Two explanations have been offered for this positivity bias. One is emotional in nature. Positivity biases are increased when feedback has threatened self-esteem (Taylor & Brown, 1988). On the other hand, positivity bias can also result when people try to cut cognitive corners when making a self-evaluation. For example, people may make evaluations with the first information that comes to mind rather than doing a more complete search for relevant information. The most easily accessible information about the self tends to be positive (Beer, Chester, & Hughes, 2013; Paulhus, Graf, & van Selst, 1989) and this positive information may give rise to positivity bias if is not countered by other information (Chambers & Windschitl, 2004). The neural underpinnings of positivity bias differ depending on whether the bias is driven by threat or the failure to use more than easily accessible information (Beer, 2014; Flagan & Beer, 2013).
When positivity bias in self-evaluation is driven by emotional reactions to threat, it is associated with a distinct functional network of regions (VMPFC, basal ganglia, and middle frontal gyrus; Flagan & Beer, 2013). This functional network mediates a person’s threshold for claiming something as self-descriptive (Hughes & Beer, 2012a) which may allow them to inflate self-associations with positive attributes and downplay self-associations with negative attributes (Flagan & Beer, 2013). For example, participants made social-comparative judgments while self-esteem threat was manipulated (Hughes & Beer, 2013). Participants were given feedback on how likable their peers found them; feedback was interspersed with blocks of social-comparative judgments. When faced with negative feedback, participants increased the extent to which they compared themselves favorably to an average peer. The more individuals coped with the self-esteem threat with exaggerated positivity, the more they exhibited increased VMPFC activation (Hughes & Beer, 2013). Follow-up analyses showed that VMPFC simultaneously increased functional connectivity with the basal ganglia and decreased functional connectivity to the middle frontal gyrus (Flagan & Beer, 2013). A further study used signal detection analyses and found that this functional network is associated with changing thresholds for claiming self-associations in a self-protective manner (Hughes & Beer, 2012a).
However, a different neural profile emerges when positivity bias in self-evaluation is examined in the absence of self-esteem threat. In this case, reduced VMPFC activation predicts positivity bias in social-comparative judgments (Beer & Hughes, 2010; Hughes & Beer, 2012b), overclaims of knowledge (Beer, Lombardo, & Bhanji, 2010), and discrepancies between self-evaluation and external observers (Beer, John, Scabini, & Knight, 2006; Beer, Heerey, Keltner, Scabini, & Knight, 2003). One fMRI study (Beer & Hughes, 2010) used the very same social-comparative judgments of personality traits that had previously been examined in relation to self-esteem threat as mentioned before (Hughes & Beer, 2013). When the self-esteem threat manipulation was not included, the less individuals exhibited VMPFC activation, the more they exhibited positivity bias in their social-comparative judgments. Furthermore, follow-up analyses suggest that VMPFC is working in a completely different functional neural network in these cases. The VMPFC region that shows a negative correlation with positivity bias does not show changes in functional connectivity with regions of the basal ganglia or the middle frontal gyrus. Instead, it shows decreased functional connectivity with a wide variety of neural regions including the regions in the temporal, occipital, and frontal lobes (Flagan & Beer, 2013). Therefore, research on positivity bias illustrates that differences can be found at the neural level when emotion influences social cognition compared to the absence of emotional influence.
Challenges for Distinguishing Social Cognition Driven by Emotion at the Neural Level
Although many studies of brain function have been interpreted as supporting the dual-process perspective, it is still premature to make a strong argument that a mode of reasoning that operates on emotion can be distinguished at the neural level from other modes of reasoning. There are number of problems that pervade current research on this topic.
One issue is that many of the studies interpreted in support of the dual-process perspective rely on reverse inference (Beer, 2015; Poldrack, 2006). In much of the research, emotion is presumed to be a driving force in social cognition but emotion itself is not experimentally manipulated or even measured. Instead, the fact that certain neural regions showed change in activation is often interpreted as proof that emotion was elicited. For example, regions within the limbic system are often presumed to reflect the operation of emotion. A reverse inference sometimes follows the form of “if the [limbic area such as the amygdala] shows increased activation, then that means the participant was experiencing [negative emotion].”
However, reverse inference is problematic. Regions within the limbic system support more than just emotion, emotion arises in relation to activation in regions within and outside the limbic system, and there is no way to infer a discrete emotion (e.g., fear vs. disgust) on the basis of brain activation in an individual participant. There is not a one-to-one correlation between the operation of emotion and limbic system activation (LeDoux, 1993). Furthermore, meta-analyses of emotion have shown that emotion is encoded in a widespread manner in the brain including brain regions that have other functions (Lindquist, Wager, Kober, Bliss-Moreau, & Barrett, 2012) nor is there a one-to-one correlation between discrete emotional states and brain activation (Hamann, 2012). The lack of experimental manipulation and measurement of emotion leaves open the possibility that the neural distinctions do not actually arise from differences in the operation of emotion.
Another problematic aspect of the current research is that it focuses on differentiating modes of reasoning in terms of neural regions. First, the focus on individual regions ignores the greater likelihood that psychological processes are reflected in patterns of brain activation and covariance of activation between brain regions rather than changes in isolated regions (Beer, 2015). Second, the existing data does not neatly divide into one set of regions associated with emotional reasoning and another set associated with nonemotional reasoning. Many regions have been implicated in both of the modes of reasoning described by dual-process theories. As mentioned before, the VMPFC has been characterized as a region that is important for mediating the influence of emotion on judgments (e.g., Greene et al., 2001; Hughes & Beer, 2013) as well as important for reasoning free from emotional influence (e.g., DeMartino et al., 2006; Mehta & Beer, 2010). In fact, many other neural regions have also been implicated in both modes of reasoning.
For example, the lateral orbitofrontal cortex (LOFC), predicts the use of emotion for gambling decisions when it is helpful and the suppression of emotion for gambling decisions when it is hurtful (Beer, Knight, & D’Esposito, 2006). In one study, participants were asked to make a series of gambling decisions and these gambling decisions were interspersed with pictures that were negative or neutral in emotional tone. Participants were instructed to ignore the pictures when making gambling decisions. Participants’ success in preventing emotional responses to the pictures from influencing their gambling decisions was predicted by increased activation in the lateral orbitofrontal cortex (LOFC). In a second study, participants performed the same procedure except instead of being asked to suppress reactions to the emotional pictures, they were told that the emotional information was useful for placing bets. In particular, negative emotion indicated a subsequent bet was likely to be particularly risky. The LOFC region from the first study was used as a region of interest for predicting emotion-driven bets (e.g., the tendency to risk less in the negative emotion condition compared to the neutral condition). The study found that this same subregion of LOFC that had been associated with preventing the influence of emotion on betting was now associated with using emotional information to place a bet (Beer, Knight, et al., 2006). Therefore, this series of studies show that the very same subcluster is activated when reasoning relies on emotion and when reasoning suppresses emotional influence. The common thread was that LOFC modulated emotional influence based on its utility for that situation.
Similarly, some neural regions persist in coding value regardless of whether an emotional or nonemotional mode of reasoning is activated (Bhanji & Beer, 2012a). In one study, participants first evaluated two food items and were then asked to choose between them. The mindset, that is, mode of reasoning driving the evaluation and choices was manipulated. Participants were sometimes prompted to think about taste (i.e., emotional reasoning) and other times prompted to think about health (i.e., rule-based reasoning). Participants were told that they would receive one of their choices at the end of the experiment which ensured that they maintained interest and gave honest answers. After leaving the scanner, participants rated the taste and health value of each food item. Brain activation measured during the evaluation phase found that it was not always the case that emotional reasoning triggered a distinct neural profile for value calculation (compared to the rule-based mindset). Instead, certain brain regions were modulated in relation to calculating taste or health value (e.g., medial orbitofrontal cortex [MOFC] for taste, LPFC for health) and these effects held across mode of reasoning.
In summary, a second manner in which neuroscientists have sought to understand the role of emotion in cognition is to test whether there is neural basis for dual-process theories. In particular, studies in a wide variety of domains including economic decision-making, moral reasoning, and self-evaluation have tested whether emotion-driven social cognition draws on a distinct neural system. Despite many studies being interpreted as supportive of the delineation of emotion-driven processing system, strong evidence that dual-process theories reflect the reality of neural operations remains elusive. Studies typically do not experimentally manipulate emotion nor measure it and instead favor problematic reverse inference to draw conclusions about the involvement of emotion. Similarly, many studies have now shown that focusing on individual regions and trying to assign them to one mode of reasoning or another cannot explain the existing body of findings. Many regions are implicated in both modes of reasoning.
What Do We Know About the Neural Underpinnings of Emotional Influences on Social Cognition?
While research on the somatic marker hypothesis and dual-process theories have dominated systematic investigations into the neural underpinnings of emotional influences on social cognition, a few recent studies have moved away from these approaches and tried to address the issues with emotion measurement and direct tests of neural mediation. The one study that has experimentally manipulated emotional experience separately from social cognitive judgment raises an intriguing possibility about emotional influences on social cognition. Specifically, this study suggests that emotional influence on social cognition may be reflected in two stages of brain activation: one set of changes in brain activation when emotion is elicited and another when the social judgment is executed (Bhanji & Beer, 2012b). Additionally, another study has taken the step of testing brain activation as a mediator of the relation between an experimentally manipulated emotional state and social-cognitive judgment (Harle, Chang, van’t Wout, & Sanfey, 2012). This study finds support for the role of insula in mediating emotional influences on social cognition.
The direct measurement or manipulation of emotion is a critical step in understanding how the brain mediates emotional influence on social cognition but this approach has not been used in many studies on this topic. The one study that has taken this approach asked whether emotional influence is marked by changes in brain activation arising at the point when emotional experience begins or at the point of the subsequently influenced judgment. In other words, is the brain doing something during the emotional experience that predicts whether the emotion will influence subsequent social cognition or do these differences appear as the social cognition is being executed? The study found that the answer was both.
This fMRI study experimentally manipulated emotional state and then used a separate probe to measure biases in social judgment (Bhanji & Beer, 2012b). Participants viewed words that either elicited negative emotion (e.g., murder) or were emotionally neutral (e.g., copper) and then had to identify the emotional expression of the first face they saw in an ambiguous picture (i.e., a picture of partially overlapping faces). Specifically, participants had to choose whether they first saw a happy face (not emotionally congruent in the emotional prime condition) or an angry face (emotionally congruent in the emotional prime condition). Most trials included one happy and one angry face, counterbalanced for position within the picture. However a subset of trials included two faces with the same emotional facial expression in order to check that participants were not simply stating they had seen a face that matched the word prime. As expected, participants were more likely to identify the angry face after seeing the negative word prime.
The design of this study permitted the measurement of neural activation at the time of emotion elicitation (i.e., the presentation of the word prime) as a function of whether it changed depending on whether a subsequent judgment would reflect emotional influence (i.e., be congruent or not with the negative emotion). It also permitted the measurement of neural activation at the time of judgment as a function of whether it had or had not been influenced by emotion. Results showed that emotional influence on social cognition is reflected in one set of neural changes at the time of emotion elicitation (i.e., the word prime) and another set of neural changes at the time of social cognitive judgment (i.e., which face they saw first; Bhanji & Beer, 2012b). Specifically, one subregion within VMPFC predicts whether an emotion will influence subsequent social cognition and a different subregion within VMPFC predicts whether a judgment has been influenced by previously elicited emotion. In contrast, an emotion prime that does not influence subsequent judgment is associated with greater activation in the dorsomedial prefrontal cortex, ventrolateral prefrontal cortex, and lateral OFC. By separating the experience of emotion from the judgment which it may influence, this study builds on our knowledge by showing that emotional influence on social cognition is reflected by two different patterns of neural activation at two different time points. One that is associated with the emotional experience itself and another that occurs when the judgment is executed.
Another study has advanced our understanding of the neural underpinnings of emotional influences on social cognition by directly testing brain activation as a mediator (Harle et al., 2012). In this study, participants were randomly assigned to a sadness or neutral condition. Specifically, participants watched movie clips designed to elicit sadness or a neutral emotional state before beginning a block of offers from the Ultimatum Game. As described earlier, the Ultimatum Game consists of offers to split a pool of money which vary from unfair to fair divisions of the money. Individual frames from the movies were further interspersed with the Ultimatum Game offers to boost the mood induction. As expected, participants in the sadness condition were less likely to accept offers including unfair offers and also reported greater sadness at the end of the study. Furthermore, the study found that insula activation mediated the relation between sad mood and rejection of offers. Although insula had been previously theorized as a region involved in using emotional reasoning, this study provides strong support because it is the first study which experimentally manipulates emotion, includes an emotion manipulation check, and tests neural activation as a mediator of the relation between emotion and social-cognitive judgment.
What We Can Do to Learn More
The existing neuroscience literature highlights the need for a different approach to understanding emotional influences on social cognition at the neural level. More research is needed to deeply understand whether two stages of neural processing capture emotional influence on social cognition and, if robust effects emerge, their psychological implications. This endeavor will require a theoretical model other than dual-process theories or theories focused on physiological arousal as a critical factor. Additionally, different experimental designs are needed to more rigorously test the bridge between psychological models and neural operations.
Two Stages of Emotional Influence?: Implications for Future Research, Guiding Frameworks, and Experimental Design
Future research and guiding frameworks
An important next step will be to more systematically investigate whether emotional influence on social cognition is reflected in two stages of neural changes as suggested by research which has not used reverse inference to interpret emotional influence. First and foremost, more research is needed to test the robustness and generalizability of these initial findings. So much of the existing work has been dominated with studies that have not considered the time points of emotion and social cognition separately. Therefore, it will be important to build a body of research that takes this distinct approach to understand whether the two stages of neural activation are consistent across studies.
Second, studies investigating the possibility of a two-stage model require a departure from the research questions and experimental designs typically used in studies focused on physiological arousal or dual processes. Neither dual process nor theories of physiological arousal predict that neural coding of emotional influence on social cognition would take distinct forms at the time when emotion is initially experienced and then when social cognition is executed. However, there are many other psychological models of emotional influences on social cognition (e.g., Bower, 1981; Clore, Schwarz, & Conway, 1994; Forgas, 1995; Keltner, Ellsworth, & Edwards, 1993) which make predictions that are consistent with a two-stage process at the neural level.
For example, the affect infusion model (AIM; Forgas, 1995) integrates several previous theories and makes predictions that are consistent with a two-stage process of emotional influence. In contrast to dual-process and physiological arousal theories, the AIM posits that there are two different kinds of events that are triggered by an initial emotional experience and each has a different means of influencing subsequent cognition. Therefore, the neural activation seen at the time of emotional experience which predicts subsequent emotionally influenced social cognition might reflect these different processes. One process is called “affect priming” and, in this case, the emotional experience triggers a set of memories, attentional priorities, and the depth of processing that is likely to follow the emotional experience. In contrast to dual-process theories, AIM does not equate emotion with shallow processing. Instead, positive and negative emotions are considered likely to engage shallow versus deeper processing in many cases. The “affect priming” process is contrasted with “affect-as-information” in which emotion is simply referenced in future judgments as the source of information relevant to the judgment. In other words, a person may be likely to consult their good or bad mood as means of judging whether they want to go see a movie rather than characteristics of the movie itself. It is important to note that AIM posits that although emotional experiences may predispose someone toward certain processing strategies (affect priming) or to certain information (affect as information) for a subsequent judgment, the degree of influence of emotion on the subsequent cognition is ultimately affected by situational and personal factors. Therefore, this perspective explains why there may be neural activation seen at the time of emotion that predicts emotionally influenced social cognition (either priming or information) but a second process would be required to operate as cognition gets underway to achieve emotional influence.
AIM also discusses emotional influences which are either integral or incidental to a subsequent judgment (and also see Blanchette & Richards, 2010; Johnson-Laird & Oately, 2000) which may explain why different neural operations are found at the time of emotional experience versus social cognition. Integral emotional influences are those that are relevant for making a social cognitive judgment whereas incidental influences are those that come from an unrelated context (e.g., a bad mood elicited by a difficult commute influences judgments of a spouse’s actions). It might be that the neural processing at either the time of emotion elicitation or social cognition reflects the expected utility of the emotion for that context. If the pattern that predicts emotional influence differs for integral versus incidental emotion, that would suggest that these two kinds of emotion impact social cognition through distinct psychological mechanisms.
Beyond its relation to AIM, a two-stage neural processing of emotional influences on social cognition would also have implications for the conceptualization of emotion regulation. Are different processes used to regulate the emotional experience versus its influence on subsequent social cognition? Are some more successfully applied at one stage versus another? If the emotional experience is successfully regulated after it arises, does that mean that it has no chance of influencing social cognition or is there still a trace that leaves open the possibility of influence at the time of judgment? These are just some examples of questions that would need to be understood at the behavioral level if it became clear that emotional influences occur through mechanisms that arise at the time of emotion elicitation and another set of mechanisms at the time of subsequent social cognition.
Experimental design considerations
In addition to the selection of a new theoretical model to test across the psychological and neural levels of analyses, there is also a more concrete need to modify current experimental design approaches to better understand the neural mediation of emotional influences on social cognition. These changes should take into consideration that differential processes may be happening when emotion first arises and then later when social cognitive judgment is engaged. To address this consideration, future studies should include all of the elements seen in the study which examined the manipulated effects of sadness on Ultimatum Game decisions (Harle et al., 2012) but also take a step further. Specifically, studies should experimentally manipulate emotion or, at the very least, measure emotion to substantiate that emotion rather than another variable creates differences in social cognition. Without the measurement of emotion it is not possible to test whether effects are explained by emotional influence (e.g., compared to other aspects of that mode of reasoning as described by dual-process theories such as associative processing, rapid processing, etc.). Additionally, testing brain activation as a mediator of the relation between emotion and social cognition is a fruitful way of identifying neural patterns of processing that reflect emotional influences on social cognition. Finally, future studies need to take a step further by not focusing on individual neural regions and test whether neural patterns of activation are consistently associated with emotion-driven social cognition (in comparison to social cognition not influenced by emotion).
The focus on patterns of neural activation (e.g., sets of regions that show covariation in their activation) is important for two reasons. First, patterns of neural activation (rather than activation within an isolated region) are more characteristic of how the brain operates in relation to psychological processes (Beer, 2015; see also Hamann, 2012). Second, the focus on networks also clarifies comparisons across studies. If insula activation is associated with emotional influences on social cognition in two different studies, then it is much more convincing that these two findings are related if insula activation is not only similar in location but also covarying in a similar manner with other brain regions across the studies. Additionally, network analysis has already proven important interpreting otherwise puzzling results. For example, the findings in the research on positivity bias in self-evaluation might appear paradoxical without the benefit of network analysis. In some studies, VMPFC activation predicts greater positivity bias while in other studies reduced VMPFC activation predicts greater positivity bias (i.e., Beer, 2014; Flagan & Beer, 2013). However, the addition of functional network analyses showed that this pattern does not reflect some kind of continuum of VMPFC activation but rather VMPFC is consistently acting within a particular functional network when positivity bias is driven by a need for self-protection and a different functional network when positivity bias is driven by other variables (Flagan & Beer, 2013). The examination of neural networks rather than isolated neural regions has much more promise in testing whether dual-process theories capture operations at the neurobiological level of analyses.
Finally, it is important to note that the investigation of whether emotion-driven social cognition differs at the neural level is not meant to negate any value that psychological theories have for explaining behavior. Instead, neuroscience research should be viewed as a complement which speaks to whether the characterization of emotional influence as a distinguishing feature of social cognition is rooted in neurobiology or a useful distinction that operates only at the psychological level of analyses.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The Funding Source number for this research is NSF BCS 1147776.