
Abstract
Although more than 60 hypotheses predict associations between characteristics of nest architecture and at least one measure of breeding success, reproductive consequences of nest architecture remain quantitatively understudied. In addition, most detailed studies of animal constructions in natural conditions were conducted on species built inside human-made nest-boxes that facilitate monitoring. Here, we present the first review of methodologies and research findings in studies that examined the reproductive consequences of nest characteristics in wildlife species that build nests outside cavities, mainly focusing on nest-size components, animal-derived nest material, anthropogenic nest material, or fresh greenery (i.e., fresh vegetative materials placed in the nest). We found 84 published field studies that were conducted on 51 non-cavity nesting bird species from 10 different avian orders, mainly members of the Passeriformes (69%). We discuss why most investigations reported weak relationships between aspects of nest design and breeding success. We propose suggestions for future research on bird nests, and also highlight the importance of poorly studied research problems.
Keywords
Introduction
To maintain population size, breeding birds should produce on average at least one breeding offspring of the same sex per lifetime, regardless of the nest types they build or the number of eggs or offspring they produce. 1 To anticipate offspring loss in unpredictable living environments, maximizing the individual lifetime production of offspring with phenotypes that allow post-fledging survival (e.g., high fledgling weight) should always be considered ‘adaptive’. 2 This implies that the time-window of egg and offspring development inside the nest is an important life-history stage. However, most life-history investigations have ignored the possibility that architecture of the nest could influence the abilities of a parent to rear its offspring to independence. For instance, quantitative intraspecific wildlife studies of associations between avian nest architecture and breeding success remain rare, even though numerous hypotheses are postulated to explain how and why nest structure characteristics should be correlated with at least one measure of reproductive success.3–9
Field studies that have quantitatively investigated reproductive consequences of intraspecific variation in avian nest design have been guided by hypotheses that describe the potential benefits, costs or constraints of nest characteristics.7–11 Most of these hypotheses resulted from interpretations of intraspecific or interspecific correlations between nest architecture and biotic or abiotic factors without quantifying breeding success, sometimes using results of experimental studies with collected nests and nests in situ.3,7,8,10,12–35 For instance, based on the observations of changes in nest characteristics with latitude, altitude, ambient temperature, or time of the year, it has been repeatedly suggested that bigger nests with thicker, denser, nest walls and more animal-derived nest material improve nest insulation efficiency favouring the development of ectothermic eggs or offspring in colder living conditions, especially in smaller bird species.7,8,10,15,20,21,25,30,36–40 On the other hand, smaller nests with less animal-derived nest material might prevent lethal hyperthermia 41 when endothermic nestlings are exposed to a hot breeding environment.8,40,42 That birds try to avoid hyperthermia is consistent with field observations that nest placements avoid direct sun exposure, or that greenery (i.e., fresh vegetative materials placed in the nest) or roof constructions might be used to create shade.14,31,43–46 Nest structures most probably evolved to achieve control of the impact of dynamics in nest temperature, humidity or airflows during contact incubation and so minimize energetic costs.7,8
According to Deeming, 39 the ‘bird-nest incubation unit’ is required to maintain egg temperatures during incubation in small bird species. However, as the species-specific body size increases, the relative species-specific clutch mass decreases. This means that larger bird species are less reliant on the nest to confer any insulation – normally the body and body feathers can do this. 39 On the other hand, smaller, concealed, nests that are more difficult to find might lower risks associated with nest predation or brood parasitism in smaller species.26,37,47–51 By contrast, the larger flighted species (e.g., diurnal raptors, storks, herons) can only breed in big but conspicuous nests and these species may also be more effective at repelling potential predators, or they reuse nests in inaccessible sites (top of trees, cliffs, aquatic environments, or man-made platforms). It has also been hypothesized for years that nest structures are not only shaped by climatic factors or interspecific interactions, but also can play a significant role in intraspecific sexual selection involving territorial defense, mate attraction or mate stimulation, where exaggerated nest characteristics have been defined or identified as pre-mating or post-mating ‘signals’ of parental performance abilities addressed to mates or potential competitors.5,52–66
Therefore, the nest characteristics that maximize reproductive success within species should probably be adjusted to accommodate a mixture of factors that also depend on species-specific life histories and the environmental conditions when breeding.37,67 In this framework, the species-specific nest design is the outcome of an evolutionary shaped ‘compromise solution’10,49 that takes different opposing selection pressures into account (e.g., minimising energetic costs versus climate versus predation risk versus signaling of breeder characteristics reflecting abilities to take care of eggs or offspring). However, nest design is also constrained by phylogeny-based characteristics (e.g., species-specific body size influencing physical strength) and environmental factors (e.g., food availability influencing physical body condition) that together will determine the ability to carry heavier nest material and construct larger nests that can support heavier incubators and broods. 68 In addition, the strength of the local selection pressures might differ between years or sites, for instance when cold spells, heat waves, rich food abundance, or strong intraspecific or interspecific pressures do not occur every year in local study plots. This would imply that within local study plots the strength of the quantified associations between nest architecture and breeding success should exhibit significant spatiotemporal variation and often cannot be predicted in advance. 9 All this means that species-specific nest structures will never be perfectly adapted or adjusted to a single factor (e.g., energetics or climate or predation or signaling of breeder characteristics reflecting abilities to take care of eggs or offspring) which explains why nests often do not ensure total breeding success, irrespective of their design.
Most intraspecific studies of reproductive consequences of nest architecture have been conducted in secondary cavity-nesting avian species that breed in artificial boxes. Nest-box studies in the most well-studied species generally showed that nest-size components are most often weakly associated with breeding success, whereas animal-derived material (e.g., feathers, fur, hair), or greenery (e.g., aromatic herb fragments), might sometimes influence nestling characteristics that reflect abilities to survive.6,9 However, nest-box studies can suffer from methodological biases caused by the position and design of the boxes that impose constraints on when, where, and how the nest will be built. For instance, in cavity-nesting species the size of the nest is often closely associated with the size of the nest-box chamber.11,69,70 On the other hand, cavity nests built outside of human-made boxes are often constructed in situations that are less accessible for research. The nests of species that routinely nest in open environments are often abandoned at the end of the breeding attempt allowing them to be collected for analysis. However, not all such nests are necessarily readily available for analysis of composition or architecture. For instance, open nests of large bird species can be reused during more than one breeding attempt so their removal to allow them to be dismantled to determine their composition, and associated success of former breeding attempts, would compromise further breeding attempts. Thus, the costs and benefits of field research practices will depend on the species-specific life histories and associated nest types investigated.
The reproductive consequences of nest-box design in avian studies of secondary hole-nesters have been reviewed multiple times during the last four decades.9,71–77 However, there are no detailed review papers dealing with reproductive consequences of nest types other than cavity nests, probably because these kinds of studies remain rare. Therefore, following the lead from a previous review, 9 here, we review the methodologies and research findings of intraspecific wildlife studies of a wide variety of birds (e.g., passerines, raptors, waterbirds) that quantitatively examined reproductive consequences of nest-size components, animal-derived nest material (ADNM), anthropogenic nest material (ANM), or greenery, in species that build nests outside cavities. One of the goals of this study is to highlight research problems associated with breeding success that have been ignored for decades. Moreover, comparing the findings of studies dealing with the non-cavity nesters with those of cavity nesters might help to extract general patterns shared by species with different nest types. For instance, as observed in nest-box studies,9,78,79 one might expect for the intraspecific field studies of non-cavity nesting species that: (i) most investigations will examine nest-size components without providing details of nest composition, (ii) most passerine studies will report weak associations between aspects of nest size and breeding success, and (iii) passerine studies will report associations between the amount of ADNM or greenery on the one hand and nesting success or nestling characteristics on the other hand.
Literature and search methods
The literature search methods were the same as those presented in Lambrechts and Deeming 9 The most recent publications in English were found on the electronic platforms HAL, Web of Science, ResearchGate, Google Scholar, JSTOR, and journal websites. We verified the most frequently cited review and original papers, and the contents of the publications that cited these papers. The underlying assumption was that a new publication should always cite at least one of the frequently cited papers. We only considered publications of field studies that provided information on the association of at least one physical measure (e.g., linear dimension, weight) of nest architecture and one measure of breeding success as defined in Lambrechts and Deeming. 9 Correlative studies that only focused on presence/absence of nest components (e.g., presence/absence of ANM) were not considered. Other studies on bird nests might have been published in inaccessible local journals in languages other than English. However, we noticed that the verified publications dealing with reproductive implications of nest architecture regularly cited the same publications, which indicates that the bibliographic searches were exhaustive.
Components of nest architecture measured in 57 different studies of avian species breeding outside of cavities for passerine species and in 27 different studies for all other avian orders combined. Values indicate the number of studies that reported the measure of nest architecture. Mean (± SD and range) values are provided for the number of nest components per study and for the number of years per study.

Hypotheses and associated predictions about benefits and costs of nest characteristics that have been published for species (listed alphabetically in their respective orders) breeding outside nest cavities.
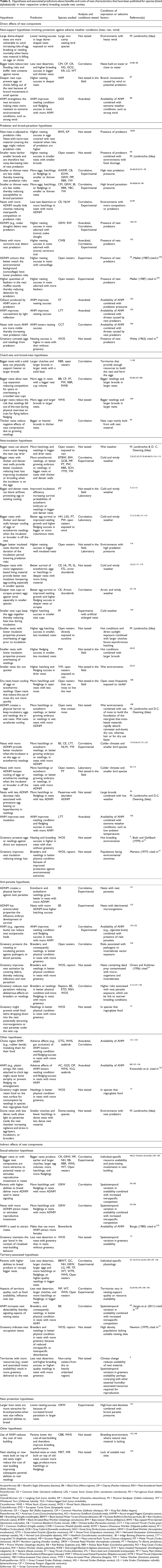
Accipitriformes: BE = Bonelli’s Eagle (Hieraaetus fasciatus), BK = Black Kite (Milvus migrans), OP = Osprey (Pandion haliaetus), RSH = Red-shouldered Hawk (Buteo lineatus).
Anseriformes: CE = Common Eider (Somateria mollissima), LSG = Lesser Snow Goose (Chen caerulescens caerulescens), FD = Ferruginous Duck (Aythya nyroca).
Charadriiformes: LG = Laughing Gull (Larus atricilla), KP = Kentish Plover (Charadrius alexandrinus), PS = Pectoral Sandpiper (Calidris melanotos), WT = Whiskered Tern (Chlidonias hybrida), YLG =Yellow-legged Gull (Larus michahellis).
Ciconiiformes: WHS = White Stork (Ciconia ciconia), WOS = Wood Stork (Mycteria americana).
Columbiformes: ETD = European Turtle Dove (Streptopelia turtur).
Gruiformes: CR = Clapper Rail (Rallus longirostris), CM = Common Moorhen (Gallinula chloropus), KR = King Rail (Rallus elegans).
Passeriformes: AC = American Crow (Corvus brachyrhynchos), AG = American Goldfinch (Carduelis tristis), AWMP = Azure-winged Magpie (Cyanopica cyanus), BB = Brambling (Fringilla montifringilla), BBWT = Black-backed Water Tyrant (Fluvicola albiventer), BF = Eurasian Bullfinch (Pyrrhula pyrrhula), BS = Barn Swallow (Hirundo rustica), BTBW = Black-throated Blue Warbler (Setophaga caerulescens), BW = Baya weaver (Ploceus philippinus), BWS = Bay-capped Wren-Spintail (Spartonoica maluroides), CB = Common Blackbird (Turdus merula), CBB = Chinese bulbul (Pycnonotus sinensis), CCT = Clay-colored Thrush (Turdus grayi), CF = Common Chaffinch (Fringilla coelebs), CWB = Common Waxbill (Estrilda astrild), EOW = Eastern Olivaceous Warbler (Hippolais pallida elaeica), FF = Fieldfare (Turdus pilaris), GCB = Gray Catbird (Dumetella carolinensis), GGS = Great Grey Shrike (Lanius excubitor), GRW = Great Reed Warbler (Acrocephalus arundinaceus), GTG = Great-tailed Grackle (Quiscalus mexicanus), HF = House Finch (Carpodacus mexicanus), HH = Helmeted Honeyeater (Lichenostomus melanops cassidix), HL = Horned Lark (Eremophila alpestris), HW = Hawfinch (Coccothraustes coccothraustes) , IB = Indigo Bunting (Passerina cyanea), LTT = Long-tailed Tit (Aegithalos caudatus), MP = Eurasian Magpie (Pica pica), MRT = Many-colored Rush Tyrant (Tachuris rubrigastra), MT = Mistle Thrush (Turdus viscivorous), NLW= Northern Lapwing (Vanellus vanellus), ORW = Oriental Reed Warbler (Acrocephalus orientalis), PT = Penduline Tit (Remiz pendulinus), PW = Prairie Warbler (Setophaga discolor), RB = Red Bishop (Euplectes orix), RBR = Rufous Bush Robin (Cercotrichas galactotes), RO = Ring Ouzel (Turdus torquatus), RW = Rufous Warbler (Aedon galactodes), RWB = Red-winged Blackbird (Agelaius phoeniceus), SCH = Spiny-cheeked Honeyeater (Acanthagenys rufogularis), SL = Skylark (Alauda arvensis), ST = Song Thrush (Turdus philomelos), STL = Short-toad Lark (Calandrella brachysactyla), TP = Tawny Pipit (Anthus campestris), WR = Wren-like Rushbird (Phleocryptes melanops), WFF = White-browed Fantail Flycatcher (Rhipidura aureola), WTR = Wood Thrush (Hylocichla mustelina), YBC=Yellow-breasted Chat (Icteria virens), YTM = Yellow-throated Miner (Manorina flavigula), YW = Yellow Warbler (Dendroica petechia).
Podicipediformes: GCG = Great Crested Grebe (Podiceps cristatus), HG = Horned Grebe (Podiceps auratus), LG = Little Grebe (Tachybaptus ruficollis).
Sphenisciformes: CP = Chinstrap Penguin (Pygoscelis antarctica).
Suliformes: GC = Great Cormorant (Phalacrocorax carbo).
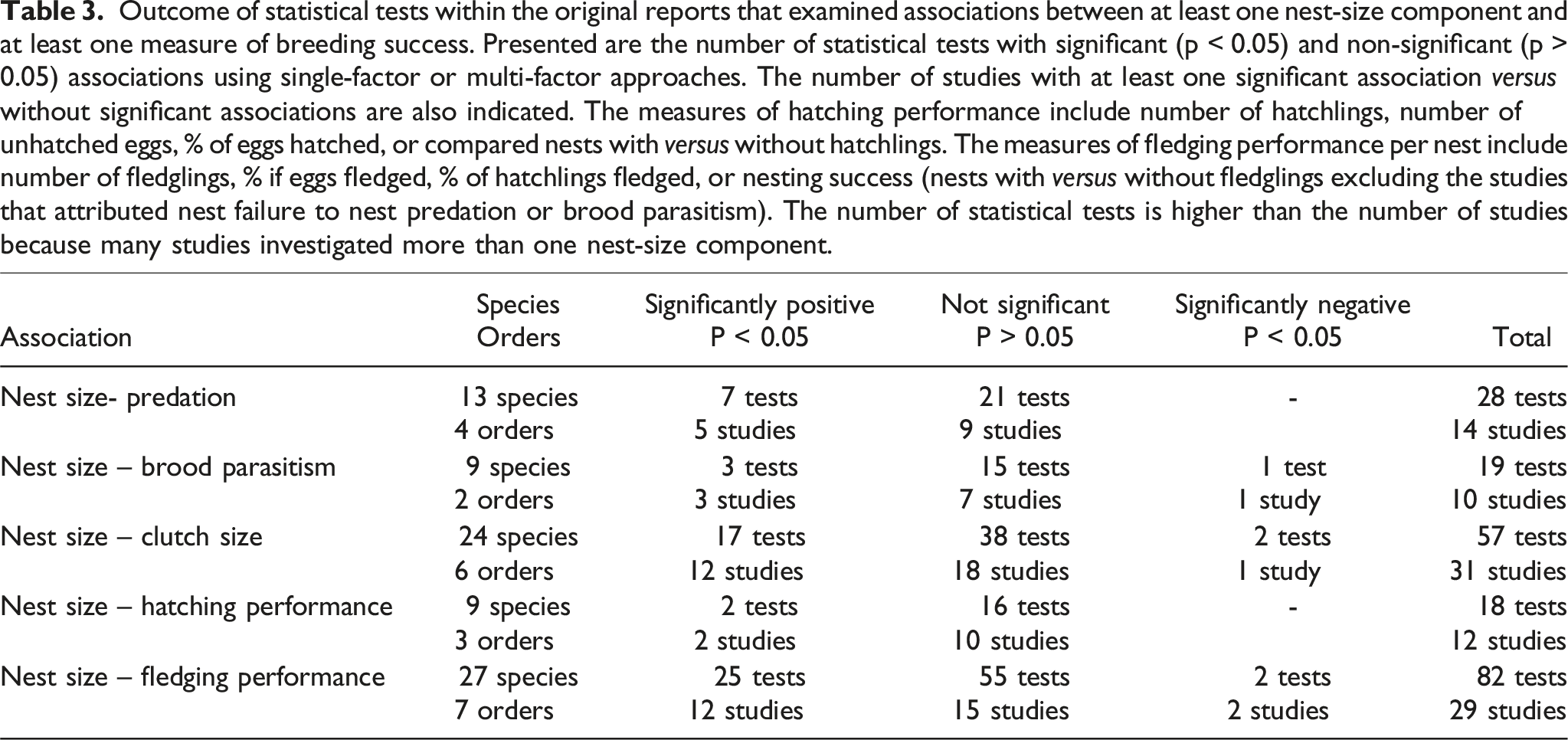
Outcome of statistical tests within the original reports that examined associations between at least one nest-size component and at least one measure of breeding success. Presented are the number of statistical tests with significant (p < 0.05) and non-significant (p > 0.05) associations using single-factor or multi-factor approaches. The number of studies with at least one significant association versus without significant associations are also indicated. The measures of hatching performance include number of hatchlings, number of unhatched eggs, % of eggs hatched, or compared nests with versus without hatchlings. The measures of fledging performance per nest include number of fledglings, % if eggs fledged, % of hatchlings fledged, or nesting success (nests with versus without fledglings excluding the studies that attributed nest failure to nest predation or brood parasitism). The number of statistical tests is higher than the number of studies because many studies investigated more than one nest-size component.
Methodologies and research findings
Methodologies
Small open nests are more difficult to find than nests constructed in accessible and known nest-boxes. Finding nests requires identification of the position of song posts, binocular observations of birds carrying nest material or feeding chicks, and systematic searches in potential territories.40,91 These nests are more likely to be found during the later stages of breeding, for instance at the time the parents feed the noisy chicks. However, a fast-growing brood might stretch the nest cup increasing the cup diameter and compacting the nest wall, perhaps causing positive associations between nest-size measures and breeding success when nests differ in the timing of nest measurement, and can explain why successful nests of some species have a wider nest-cup diameter than unsuccessful nests.40,48,49,150 In addition, nests built inside nest-boxes can be easily extracted from the boxes and investigated in more detail at different moments of the breeding cycle.151,152 However, the base of a nest built outside a cavity is often tightly attached to branches or other physical supports, for instance to avoid strong winds or branch movements that lower nest stability increasing the risk that eggs or chicks fall out of the nest.87,110,153,154 Therefore, research-associated removal of nests tightly attached to a physical support might create nest damage, for instance when some of the extracted nest material is falling on the ground, 155 perhaps changing the physical structure of the occupied nests after research-associated verifications. Frequent nest inspections might increase risks linked to nest predation or brood parasitism, for instance when nest predators exploit cues left by humans.99,156 In addition, information on the original structures of deserted nests may not be available when nest predators destroy nests or conspecifics remove material from deserted nests to construct new nests.40,157 For instance, Antonov 49 excluded the data from nests that were noticeably damaged by predators in analyses that looked for negative associations between nest size and nesting success. Thus, research practice might alter the associations between nest architecture and breeding success. Some of the shortcomings of research practice that have been discussed in more detail for field studies of secondary cavity-nesting passerines 9 will probably also apply for studies of the non-cavity nesters. This includes the rarity of experimental approaches in natural conditions, and the lack of replication of research protocols and data analysis procedures at the within-species level.
Nest-size components
The nest components that have been linked to at least one measure of breeding success (e.g., nesting success: failed versus at least one young fledged, number of eggs hatched or fledged, % of hatchlings fledged, % of egg fledged) were mainly linear or weight measures that reflect aspects of nest size (Table 1). Nest size represents all the physical measures of the entire nest, which include diameter, dry mass, surface area, floor area, bulk or volume.11,22,25,50,138,158 Nest-size components include the measurements of the depth and diameter of the nest cup or bowl, the thickness of the nest wall expressed as the difference in diameter between the nest cup and the entire nest, the height or depth of the nest wall as the distance from the base to the top rim of the nest, the thickness of the nest under the eggs, floor thickness, and calculations of nest-cup volume, the difference in nest volume and cup volume as a measure of material volume, or the percentage of the area of the base of the nest in contact with the branches.22,40,48,49,52,91,114,159,160
The types and number of nest-size components investigated substantially differed across field studies and model species mainly involving linear measures (e.g., diameter) (Table 1). Linear measures of nest size were taken with digital callipers (± 0.5 mm), 38 rulers, or tapes (accuracy 1 mm – 0.5 cm),45,55,155,161 or a clinometer, 127 whereas the dry nest mass was measured with a Pesola spring balance48,155 or electronic balance. 38 Nest thickness under the eggs was measured with wire inserted through the centre of the egg chamber. 48 Some of the nest-size measures were replicated across studies, such as the way the nest volume and nest-cup volume were calculated by Soler et al., 162 Palomino et al., 48 Antonov, 49 Møller et al., 11 and Tabib et al. 163 Akresh et al. 40 used principal component analyses to reduce the number of nest-size components in the statistical analyses involving breeding success.
Field experiments that investigated reproductive consequences of nest size or nest-size components involved tests with (i) collected nests (e.g., swapping of nests differing in size within or between species),37,50,91 (ii) enlarged nests to examine incubation efficiency 120 or mate investment,61,63 or (iii) nests from which nest material was removed to examine adjustments of clutch size or incubation patterns in response to material removal. 53 However, manipulating the amount of nest material in active nests might induce parents to adjust nest size in response to the human-induced nest changes, so that the experimental conditions cannot always be fully controlled in some model species. 164
Animal-derived nest material (ADNM)
Although more than 70% of the European passerine species use ADNM (feathers, fur, hair) to line the nest, 36 and the importance of ADNM for breeding has been repeatedly stressed during the last four decades,8,17,103,165 there are few quantitative studies of the mass of ADNM (but see Biddle et al. 159 and Dickinson et al. 160 ), probably because separating ADNM from the other nest material is a time-demanding activity. 9 Kubelka et al. 34 used photographs to estimate the size of the lining material in Northern Lapwings (Vanellus vanellus), a technique that has also been applied in cavity nesters 166 and open nesters. 167 Field experiments that verified reproductive implications of experimental manipulations of ADNM remain an exception in non-cavity nesters.100,121,122 Six out of the 84 verified studies that looked for associations between aspects of nest architecture and breeding success examined reproductive consequences of ADNM (Table 1).
Anthropogenic nest material (ANM)
Presence or absence of ANM has been reported in many open-nesting species, and include plastic strings or foils, fragments of plastic bags, coloured linen or cotton, baling twine, man-made polyester, angling gear, bottle tops, plastic straws, rubber bands or cigarette butts.88,104,105,124,131,132,168,169 Experimental studies involving ANM and its reproductive implications remain rare.124,125,132 Correlative studies provide useful hints about their impacts (e.g., entanglement that prevents fledging as reported by Seacor et al. 134 for the Osprey [Pandion haliaetus]). However, studies that quantified the number or weight of ANM per breeding attempt are limited to a few species (e.g., White Storks [Ciconia ciconia], Ospreys).131,134
Greenery
Following Lambrechts and Deeming, 9 we considered greenery as fresh vegetative material that is added to the nest surface mostly after the onset of egg laying without having functions to strengthen the nest base.3,14,128 Use of greenery is a population-specific or species-specific trait. For instance, a comparative study reported higher avian blood parasite prevalence in open-cup nesting passerines than in cavity-nesting or ground-nesting passerines, 126 perhaps because of lower presence of greenery repelling flying nest parasites in open-cup nesters. In starlings (Sturnidae), greenery is more frequently used in cavity-nesting species than in open-nesting species. 29 This was attributed to greenery-induced volatile concentrations being higher inside cavity nests and not exposed to an airflow than in open nests exposed to wind, possibly implying that open-nesting passerines rarely use greenery to adorn the nest. On the other hand, it has been hypothesized that the abundant use of greenery by raptors (Accipitriformes and Falconiformes) might have many roles, such as: masking host-generated odours as a nest-protection strategy, repelling parasites via the action of chemical volatile compounds, covering nest debris in a nest sanitation context, concealing eggs or nestlings from predators, regulating nest humidity, protecting eggs and nestlings against direct sun exposure, or altering conspecific sex-associated interactions via the identification of active nesting status.3,14,170 Greenery (e.g., fresh conifer branches) might repel nest parasites that are attracted to the odours of dead prey fragments accumulating inside the raptor nest, although Wimberger 3 did not find an association between diet and greenery use across raptor species. Greenery might stimulate immune functions in adults or nestlings in cavity-nesting species,171,172 but this possibility has been ignored in published studies of open-nesting species. In raptors, greenery is quantified with binoculars to obtain estimates of the % of greenery added to the nest. For instance, green branches can represent >70% of the weight of Bonelli’s eagle (Aquila fasciata) nests. 129 In contrast, small passerines add herbaceous greenery in small quantities relative to the mass of the whole nest. 6 The lack of greenery in nests examined ex situ may relate to the open nests of the small bird species being inspected after the greenery fragments dried out and disappeared.27,127 In addition, cryptic nests built in dark environments (e.g., bushes, shrubs) might complicate the detection and quantification of small greenery fragments in natural conditions to explain why there are no quantitative studies of reproductive implications of the mass of herbaceous greenery in small, non-cavity nesting landbirds. It does not exclude the possibility that absence of greenery might chemically conceal nests to prevent nest predation.
Research findings
As expected, most short-term field studies of reproductive consequences of nest architecture in the non-cavity nesting species focused on nest-size components that can be easily measured in natural conditions. Few studies have reported correlations between aspects of nest composition on the one hand and nesting or breeding success on the other hand.117,173 As in cavity nesters, intraspecific repeat studies conducted by different research teams were limited to a few non-cavity nesting model species (e.g., Common Blackbird Turdus merula; Eurasian Magpie Pica pica; and White Stork). These studies are described in more detail below.
Nest size, predation and brood parasitism
Non-cavity nesters are expected to be more vulnerable to predation or brood parasitism than cavity-nesters given that active open nests are potentially more detectable than active cavity nests.47,51,147 Thus, the idea that predation pressures or brood parasitism should favour the construction of smaller, less conspicuous, open nests at the intraspecific level has been proposed for decades (Table 2).37,47,91,93,147,174 We found 14 field studies that examined associations between aspects of nest size and nest predation risks, and 10 studies that examined associations between nest size and brood parasitism (Table 3). Whereas eight out of 24 verified studies reported that depredated or brood parasitized nests were on average larger than non-predated or non-brood parasitized nests,37,49,92,96,147 67% of the studies showed that neither nest size, nor its components, were linked to the risk of predation or brood parasitism (Table 3). In 2004, Antonov 49 claimed that nest size has not been found to be related to predation probabilities within any species. This may reflect the point that in most studies the impact of nest-size associated predation was masked by other factors, such as breeder characteristics that allow the construction of larger nests in territories that also provide better protected nest sites and the adults have more effective anti-predator behaviours.34,40,48,59,91,92,158,174–176 For instance, Grégoire et al. 91 compared predation pressures on natural Common Blackbird nests monitored during the breeding season with that of collected conspecific nests monitored after the breeding season. The contents of artificial nests with a larger external diameter were more frequently depredated than the artificial nests with a narrower diameter. However, this association was not found in the natural nests, perhaps because the parents modified the association between nest size and breeding success in the natural nests via effective anti-predator behaviour, such as concealment or defence of nests. In Great Reed Warblers (Acrocephalus arundinaceus), breeders with larger nests seem to be more effective in avoiding brood parasitism because of higher discriminatory abilities against Common Cuckoo (Cuculus canorus). 59
In a study of Yellow Warblers (Setophaga petechia), birds in the subarctic produced larger, and better insulated nests than conspecifics in temperate regions. Smaller, poorly insulated, temperate nests were exchanged with nests from the subarctic; in the subarctic region the smaller, non-local, temperate nests suffered less from nest predation than larger, local, subarctic nests. 37 Also, the larger subarctic nests contained more lining material (e.g., white feathers) and therefore were more potentially detectable from a distance by nest predators (e.g., see also Møller 177 ). By contrast, in the temperate zone, the nest predation rates were similar for large subarctic nests and small local nests. Therefore, the strengths of the quantified relationships between nest size and nesting success in Yellow Warblers were study-area dependent.
If visually-based nest detectability per se is a key factor influencing predation risks, passerine species with smaller nests should suffer less from nest predation than passerine species with larger nests, which has been observed in some multi-species studies.50,176,178,179 If visually-based nest detectability per se in combination with benefits of nest-size selection are key factors influencing brood parasitism, brood parasites should prefer larger than smaller host nests in choice experiments, which has been observed in a field experiment with Common Cuckoos visiting nests of Oriental Reed Warblers (Acrocephalus orientalis). 147 Nest-site selection can disassociate nest size from breeding success, for instance when nests are built in sites that are inaccessible to predators180,181 or in areas where predation pressures are low. 78 Wildlife studies interested in impacts of predation pressures that simultaneously investigated nest-site characteristics and nest characteristics concluded that the nest-site characteristics (e.g., nest height above the ground, substrate, vegetation density) had a more important impact on nesting success than the nest characteristics per se107,163,182 (but see Tsogt et al. 179 for an inter-specific study). It is evident that the outcome of such a study should depend on the composition of the predator community present on the study plots, which can vary between years or study areas.37,78 For instance, absence of nest predators, combined with a positive effect of breeder characteristics on nest size, might explain why Sovrano et al. 141 found a positive relationship between the external nest width and the daily nest survival rate in Black-backed Water Tyrants (Fluvicola albiventer).
Nest size and clutch size
The ‘clutch-size’ hypothesis of nest size predicts that: (i) parents that are physically able to build larger nests are also physically able to lay more eggs, (ii) parents adaptively adjust clutch size to nest size to prevent overcrowding inside the nest cup, or (iii) males build larger exaggerated constructions to induce females to lay more eggs (Table 2).47,53,107 This hypothesis has been tested in 31 field studies (Table 3). Although Møller et al. 11 concluded that the nest-base area is positively associated with clutch size in several cavity-nesting and open-nesting species, they found huge intraspecific and interspecific variation in the slope estimates of these relationships. Twelve of the 31 verified studies that focused on nest-size components other than those investigated by Møller et al. 11 reported positive associations between aspects of nest size and clutch size (Table 3).48,61,109,138,139,141,142,183,184 For instance, four studies found that White Storks with larger nests produced more eggs per breeding attempt.109,138,139,185 In White Storks, nest size reflects past nest-building efforts in reused nests and the males preferentially occupy the larger nests. Therefore, nest size might become a proxy of the characteristics of the territory or the physical strength or other characteristics of the male breeder.109,138,139
In Eurasian Magpies, an experimental reduction of nest size via a reduction in the thickness of the roof layer prior to the onset of egg laying was associated with a reduction in clutch size. 53 Soler et al. 53 proposed that nest size might be a post-mating signal that reflects the male-builder’s willingness to invest in reproduction and is aimed to stimulate egg laying in mates. However, experimentally removing roof material might have induced more light penetration inside the nest chamber that also might have increased stress that reduced investment in egg formation or incubation.
By contrast, other studies dealing with other species reported weak associations between nest size and clutch size or clutch volume.48,49,64,114,155,186 For instance, Teglhøj 161 found in Barn Swallows (Hirundo rustica) that smaller clutches and smaller broods were found in newly built natural nests compared with artificial nests. This was even though the volumes of the nest cup and entire nest were larger and more variable in the newly-built natural nests than in the artificial nests. Increased physiology or energetics-associated costs of nest building might have lowered the reproductive performance in the birds that built a natural nest.
The strength of the associations between nest-size components and clutch size also depended on how the data were analysed and which nest-size components were considered (nest-cup volume versus nest mass versus nest diameter, e.g., Suárez et al. 142 ). For instance, Palomino et al. 48 reported that clutch size increased with the mass of the non-deconstructed nest collected after nest desertion, whereas clutch size was disassociated from nest-cup volume. However, this study did not control for the amounts of debris produced by growing nestlings and trapped inside the non-deconstructed nests. Nestling skin dust can substantially increase nest weights, which correlates with brood size, therefore possibly creating positive associations between nest weight components and clutch-size associated brood size.79,152,187
Whiskered Terns (Chlidonias hybrida) build plant-based, platform nests on rafts of floating vegetation and Paillisson and Chambon 64 reported a weak correlation between the male-built nest volume and the mean egg volume per clutch. The characteristics of the supporting nest substrate seemed to be essential for the construction of large tern nests, so the relative importance of the direct versus indirect effects of nest-size components on egg production abilities remains to be identified. Of course, large nests might be better insulated than small nests, which may perhaps improve the energy budgets of females to form eggs in egg-layers that use well-insulated nests as nocturnal roosting sites around the egg-laying period.
Nest size and breeding success
Characteristics of the individual, territory and/or nest per se might indirectly or directly cause positive relationships between nest size and aspects of breeding success (Table 2). However, only 12 out of 29 field studies reported at least one statistically significant positive association between nest-size components and different aspects of breeding success, including the success of nesting (failed versus at least one fledgling), hatching or fledging (Table 3).37,40,89,94,107,111,112,163,188 In addition, only 30.5% of the 82 statistical tests were significant at an alpha threshold at 0.05. This is consistent with the findings of most field studies in the secondary-cavity nesting passerines.9,79 However, there are some exceptions. In the subarctic study of Yellow Warblers, smaller, and less insulated, non-local temperate-zone nests produced slower-growing nestlings than larger, better insulated, local subarctic-zone nests, but the average number of fledglings per nest was higher for the smaller, non-local, temperate-zone nests that suffered less from nest predation. 37 In a study of female-built nests of Great Reed Warblers, nest size was positively associated with nestling weight, brood size and the probability that nestlings return to their natal site, and nestlings were fed more by the male in experimentally enlarged nests compared to control nests. 61 In a study of Iberian Bullfinches (Pyrrhula pyrrhula iberiae), the nest volume of successful nests was larger than that of unsuccessful nests, 155 but the study did not report associations between nesting success (failed versus successful) and the other five nest-size components measured (i.e., nest weight, nest and cup diameter, nest and cup depth). In a study of Penduline Tits (Remiz pendulinus), nest size was positively associated with hatching success and brood size at fledging, 186 but there are no repeat studies in natural field settings.
In contrast to intraspecific studies that claim that larger nests are built by more experienced breeders that also occupy better-protected and richer territories, Antonov 49 found in Eastern Olivaceous Warblers (Hippolais pallida elaeica) that smaller and denser nests had higher fledging success, whereas the nest volume was not related to hatching success. It was argued that, besides natural selection, sexual selection might have favoured smaller and denser nests, although 78% of the nest-size variation remained unexplained in this non-experimental study. 49
In Eurasian Magpies from Iran, nests with a smaller internal diameter had higher nest productivity. 189 Moreover, in a study of Turkish Eurasian Magpies, nest diameter and nest volume were weakly related to breeding success in two habitat types. 188 However, the Eurasian Magpie studies in Spain postulated that a large nest is a reliable indicator of parental willingness to invest in reproduction or abilities to rear offspring that cause positive relationships between nest size and aspects reflecting breeding success.53,55,96 For instance, De Neve et al. 55 reported a positive relationship between magpie nest size and nestling immunocompetence (T-cell mediated) in control nestlings, not in nestlings that were experimentally fed to reduce the impact of variation in parental abilities to rear nestlings. In addition, the difference in nestling immune response between fed and control nestlings within a nest was larger in parents with smaller nests, perhaps because parents from larger nests delivered richer and more food to the control nestlings than parents from smaller nests. However, in the same study, nest size was weakly related to the nestling body condition index when the statistical effect of brood size was not considered.
In a study of White Storks in Spain, larger nests were associated with higher breeding success and with the number of times that the nest was used in the past. 139 However, in a repeat study with White Storks in Algeria, the size of the nest surface area was not related to hatching or fledging success. 138 Thus, as in secondary cavity-nesting passerines, 9 the strength and direction of the associations between nest size and aspects of breeding success can differ in intraspecific repeat studies, also depending on the number and types of variables included in the statistical analyses or the environmental factors (e.g., food availability) considered.
The associations between nest size and nesting success (failed versus successful) seem to be stronger in larger aquatic bird species. For instance, investigations in Chinstrap Penguins (Pygoscelis antarctica) showed that larger stone-based nests suffered less from flooding than smaller stone-based nests, indicating a positive relationship between nest size and nesting success (failed versus successful, number of eggs/chicks lost).80,81 In addition, the physically (and presumably energetically) costly stone-provisioning behaviour was intensified when the risk of perceived nest flooding was experimentally increased. 190 Larger floating nests (nest height and/or nest diameter) of the Horned Grebe (Podiceps auratus) and the Great-crested Grebe (Podiceps cristatus) in Finland flooded less than smaller nests. Here, the larger nests were more robust in response to changes in water levels and waves, and therefore were more successful. 82 All this is consistent with additional observations in King Rails (Rallus elegans) that adjusted nest height in response to fluctuations in the level of variation in water level, 83 or Laughing Gulls (Larus atricilla) that repaired nests in response to experimental changes in nest humidity. 84 Cheriet et al. 191 found that the larger nests of Great-crested Grebes in Algeria were more productive than smaller ones, but they ignored whether the larger nests were built by older, more experienced, individuals. Gaines et al. 85 found in Clapper Rails (Rallus longirostris) that the active nests had a larger diameter than the non-active nests. In the Little Bittern (Ixobrychus minutus), nest width was larger in nests used for breeding than in nests not used for breeding 192 and there was also a positive association between the nest diameter and the brood size at fledging, 193 again supporting the idea that building larger nests might be beneficial in aquatic environments. However, Amininasab et al. 86 did not find that larger nests were more productive in the Common Moorhen (Gallinula chloropus). In contrast, in the smaller terrestrial open nest types, nest flooding should not be a problem given that the porous nest material with properties that speed up drying will not allow water to accumulate inside the nest cup. In addition, parents using their body and feathers to cover eggs or nestlings can provide protection against heavy rainfall.7,8,110
One study of open-nesting species has explored the relationship between measures of thermal insulation of the nest wall and breeding success. Neither nest mass nor thermal conductance of Song Thrush (Turdus philomelos) nests collected over a period of seven years correlated with clutch size, hatchability, or fledging success, although sample sizes were very small given that 14 out of 22 nests were depredated. 194 Unfortunately, the effect of differences in thermal insulation for Common Blackbird nests in different locations in Britain on breeding success was not reported. 25 Similarly, studies of variation in open-nests and insulation in North American species have not reported breeding success.21,22
Animal-derived nest material (ADNM)
To the best of our knowledge, there are no recent field studies that investigated the reproductive consequences of ADNM in non-cavity nesters. 103 In an older study of Common Eiders (Somateria mollissima), artificial nests with exposed eggs were more vulnerable to predation than those covered with down. 99 Another experimental study with Common Eiders compared nests lined with down with that of hay and did not find substantial differences in incubation behaviour or hatching success between the two nest types, although sample sizes were small. 122 It does not exclude the possibility that down accumulating inside Eider nests might be a by-product of brood patch formation rather than an adaptive decision to improve nest insulation in cold nesting environments. Finally, more than three decades ago, Møller 121 experimentally manipulated nest feather loads in Barn Swallows to show that feather removal caused a decrease in nestling body mass being consistent with the findings of more recent experiments in nest-box breeding Tree Swallows (Tachycineta bicolor), also indicating a direct effect of feather presence on nestling development. However, feather load manipulations did not affect the number of swallow fledglings per breeding attempt. However, the study of Møller 121 was based on small sample sizes and never replicated in the same species.
Anthropogenic nest material (ANM)
Anthropogenic nest material has been found in many open-nesting species 132 most often having detrimental effects via ingestion or entanglement. For instance, consumption of rubber bands by French White Storks has been frequently observed sometimes causing gut occlusion, but the number of chicks per breeding attempt was not associated with the presence or absence of rubber bands inside the nest. 131 Artificial fibres attached to chick legs might occasionally cause leg bone atrophy and prevent fledging in White Storks (Kwieciński et al. cited in Girao et al. 137 ), American Crows (Corvus brachyrhynchos), 135 or Ospreys. 134 Entanglement due to and ingestion of ANM (mainly plastics) has also been frequently recorded in seabirds (Gall and Thompson, 2015 133 ). However, Lopes et al. 136 reported that the average hatching success in Yellow-legged Gulls (Larus michahellis) was not lower in urban study areas where ANM was frequently found than in natural study areas where ANM was rare. In addition, incorporation of smoked cigarette butts into nests of urban House Finches (Carpodacus mexicanus) significantly reduced nest parasite loads potentially favouring breeding success.124,132 In addition, Sergio et al. (2011) cited in Jagiello et al. 132 reported that Black Kites (Milvus migrans) use ANM to signal individual age and the ability to defend a territory and to successfully rear offspring (see also Canal et al. 62 ).
Greenery
Field studies report that many raptor species use greenery to adorn the nest 3 without quantifying reproductive implications of this behaviour. 128 As an exception, Ontiveros et al. 130 reported in Bonelli’s Eagles that nests that had a higher % of tree branches with greenery (Maritime Pine, Pinus pinaster) in the nest produced more fledglings per year and contained fewer ectoparasites (i.e., blowfly larvae, Protocalliphora spp.). Studies that examined the impact of fresh herbaceous greenery on reproductive success in smaller open-nesting bird species are lacking, perhaps because small plant fragments that rapidly dry out are more difficult to quantify and identify to species’ level in natural conditions (see above).
Concluding remarks and perspectives
The primary function of the vast majority of the avian-built structures is to receive the eggs in order that they can be incubated. 8 Incubation efficiency can be enhanced by at least five functions: providing thermal insulation to reduce energetic costs, providing an optimal humidity during incubation, ensuring that there is appropriate exchange of respiratory gases, contributing to the optimal positioning of the eggs or egg turning, and protecting nests against predators12,195 This does not exclude the possibility that the architecture of successful nests might vary among individuals within local study populations, 27 that different species that occupy the same habitat will build successful nests in a species-specific way (e.g., Alambiaga et al. 196 ; Deeming 8 ), or that nests might have evolved because of reasons disassociated from incubation efficiency. 10 Table 2 summarizes circa 60 hypotheses that explain how and why intraspecific variation in nest-size components, ADNM, ANM, or greenery might cause intraspecific variation in at least one measure of reproductive success. Most of these hypotheses have been mentioned in publications dealing with the architecture of nests built outside cavities. The list might have been extensively longer if other aspects of nest design had also been considered (e.g., texture, fibre size, fibre flexibility, fibre density, fibre chemistry, fibre tensile strength, nest colour, nest shape, nest asymmetry, material-associated wave patterns, camouflage patterns, contrasts between eggs or nestlings and nest material, optical effects of nest surfaces).26,65,173,197–200 However, the number of field studies that quantitatively investigated reproductive consequences of nest constructions in non-cavity nesting species remains surprisingly low. For instance, studies in which results have been published several decades ago were rarely or not replicated at the intraspecific level (but see the repeat studies of Common Blackbirds, Eurasian Magpies or White Storks, as indicated above). Experimental approaches are very rare, so the relative importance of the direct versus indirect effects of nest architecture on breeding success remain to be identified in nearly all bird species, and often can only been revealed under laboratory conditions in which birds are not provided with natural nest materials.26,201
In addition, the most cited hypotheses of functional aspects of nest design were often not supported by the data involving breeding success. For instance, in contrast to predictions of frequently cited hypotheses, many studies reported poor associations between nest architecture and breeding success in natural conditions (summarized in Table 3), which has been attributed to several potential causes. First, parental behaviour can modify the associations between nest characteristics and breeding success via nest protection strategies, such as parents covering eggs or chicks during heavy rain or direct sun exposure, anti-predator behaviour preventing predators approaching the nest or identifying the nest location or adaptive nest-site selection exposing nests less often to predators10,181 or extreme environmental conditions, such as high temperatures.7,8,10,20,91,110 Therefore, breeder behaviour that buffers the negative effects of environmental dynamics might allow nests that substantially differ in design to host similar reproductive outcomes. However, it does not exclude the possibility that the eggs or chicks from nest structures with atypical architectures might endure costs (energetic, survival) in adverse environmental conditions (e.g., not enough material causing hypothermia versus too much material causing hyperthermia).9,37
Secondly, in many bi-parental mating systems, only one of the two partners will construct the nest whereas the reproductive effort will be the outcome of the efforts of the two parents.9,10 For instance, in uniparental nest-builders (e.g., only the female constructs versus only the male constructs), the breeding efforts and associated successes do not only depend on the building performances of the nest builder, but also on the breeding investments of the partner that are not under full control of the nest builder. This implies that reproductive success will also depend on the level of mate collaboration not necessarily depending on aspects of nest architecture per se. For instance, the level of mate collaboration in nest building and breeding might significantly reduce the costs of reproduction during the later stages of breeding so that the physical shortcomings of nest structures might not always penalize breeders in species where the two sexes contribute to nest building, incubation and/or brooding.
Thirdly, the selection pressures that impact on the breeding performance in nests varying in architecture can express important spatiotemporal variation when detrimental factors (e.g., nest predation, extreme weather) are not experienced during every breeding season or in every study plot. For instance, in some years nest predators might be locally so efficient that nearly all the nests of a local study plot are discovered and destroyed, whatever the nest design. Consequently, the breeding performance associated with a given nest structure might be lowered or improved in some, but not in other, environments (e.g., habitats with versus without nest predators). 78 Moreover, because most nest studies are short-term lasting 1-3 years at most, and the environmental characteristics (e.g., weather patterns) of breeding cannot be predicted in advance by researchers planning study designs inside local study plots, the strength of the relationship between nest-architecture components and reproductive success is expected to vary significantly across local studies at the within- and between-species levels.11,37 In addition, unmeasured drivers of breeding success, such as food availability, 55 might mask the existence of associations between nest characteristics and aspects of breeding success. All this implies that the biology-based features of the study systems might result in weak associations between nest architecture and breeding success, even when field studies have been conducted in accordance with good research practice.
Finally, it is possible that variability of nest construction between years serves to optimise the nesting conditions and so maximise breeding success. As Lambrechts and Deeming 9 suggested, the normal nest-building behaviour of the birds may be preventing any obvious effects of nest composition or architecture on breeding outcomes. This view is supported in part by the observations that the performance of nest-building and breeding success are often lower in first-time breeders and both variables improve with breeding experience.139,202,203 In light of these possibilities, the question arises: how can we improve future studies researching avian reproduction and involving non-cavity nests?
Future research directions
Most of the future research directions that have been recently proposed for the nest studies of secondary cavity-nesting passerines 9 can also be recommended for non-cavity nesting species. This includes the need for more (i) intraspecific repeat studies to examine replicability of research findings at different spatiotemporal scales, also including species that build more than one nest type, (ii) long-term studies of dynamic living conditions aimed at identifying and selecting the environmental conditions during which a specific hypothesis can be adequately tested, and (iii) field experiments to identify in greater detail the relative importance of direct and indirect effects of nest architecture on breeding performance. However, there are other understudied research problems that deserve more attention, which are highlighted below.
How to build a ‘good’ nest remains an understudied research topic. 204 For instance, the restraints hypothesis predicts that nest-builders optimize, not maximize, the mass of the different nest components in the presence of abundant nest material. 9 To construct a nest, individuals can only select from the materials available inside the territory, but few studies have examined quantitatively where birds search for their nest material.199,205 It is known from species like the European Pied Flycatcher (Ficedula hypoleuca) and Eurasian Nuthatch (Sitta europea) that birds select particular nest materials, such as leaves or bark, on the basis of the availability in the immediate environment.206,207 Some studies placed material close to nests to examine nest-material choice,208,209 which might provide hints about non-random selection of nest material that significantly contributes to the functional properties of bird nests. Others added material inside the nest showing that the bird adjusts nest design in response to experimental manipulations of the nest components (e.g., Szentirmai and Szekely 164 ). However, to the best of our knowledge, there are no published field experiments that added ‘maladaptive’ nest material to active nests to examine how the breeders would behaviorally respond to the addition of such material. At least one study showed that birds are willing to accept insecticide-treated feathers to line the nest and significantly improve breeding success because of a significant reduction in nest parasites. 210 However, contaminating preferred nest material with pesticides has also been used to eliminate avian pest species that harm agricultural activities. 211 These studies, and the fact that birds are willing to adorn the nest with ANM that might vary in the levels of toxicity, reveal that nest-builders also construct nests with mal-adaptive human-modified materials. These materials might also have been contaminated with agriculture or urban-associated pesticides potentially having detrimental effects on embryos or nestlings. However, the causal association between breeding performance and the chemical toxicity of these kinds of nest material remains to be identified.
It is astonishing that the reproductive consequences of the composition of entire nests remain poorly investigated, irrespective of the avian nest type. Nests have been dismantled to measure the dry weight of each nest component in less than 50 out of more than 11,000 bird species, 8 but there are currently no quantitative studies that link information about the composition of entire nests to aspects of reproductive success in non-cavity nesting species. Many hypotheses dealing with different aspects of nest design predict that different nest components have similar reproductive consequences (Table 2), but the relative importance of the different nest components in the expression of breeding success remains to be identified because of a lack of information on details in nest composition or structure. For instance, larger nests with denser nest walls and more ADNM might have similar breeding success to smaller nests with more porous nest walls and less ADNM, depending on the thermal conditions of nesting. 9 Therefore, experiments that examine reproductive consequences of nest architecture should ideally take simultaneously the details of nest composition, nest structure, and habitat characteristics (e.g., predation pressures, ambient temperature, rainfall) into account, which is rarely or not done. Moreover, the environmental dynamics inside local study plots, such as weather fluctuations or interspecific interactions (e.g., predators, parasites) cannot be predicted in advance so that the planning of field tests of specific hypotheses becomes complicated. However, southern and northern study populations are exposed to different climates or predator communities at a macrogeographic scale. 36 Therefore, there is the option to examine the reproductive consequences of experimental nest exchange between study populations that face distinct environmental conditions, as was done in one study of Yellow Warblers. 37 However, when nests from different geographic regions differ in more than one nest component (e.g., nest size combined with wall thickness and lining material), additional experiments altering only some aspects of nest composition will also be required to identify key aspects of nest design influencing breeding success (e.g., altering lining material without changing nest size or wall thickness). To the best of our knowledge, this has never been done in experimental studies that swapped nests at a macro-geographic scale. In this framework, the application of new techniques might also quantify in more detail understudied aspects of nest organization without dismantling nests. 198 For instance, it would be interesting to link aspects of the design of active nests to the composition of the nestling microbiome assumed to impact nestling development and survival. 212
The physical properties of nests (e.g., insulative or hydrological properties, gas exchange abilities, structural support) in association with measures of nest architecture have usually been conducted in laboratory or natural conditions without taking aspects of breeding success into account33,37,40,165,213–215 Thus, nearly all wildlife studies of reproductive consequences of nest architecture used easily measurable nest-structure components as potential proxies of physical properties of nests. Therefore, there was no direct link between intraspecific variation in breeding performance and intraspecific variation in the nest architecture-induced dynamics of nest temperature, nest humidity, nest-cup air flows, or material characteristics (e.g., flexibility or density of fibres) in an active nest.7,8 On the other hand, several field studies quantified the breeding consequences of nest orientations or experimentally manipulated nest temperatures inside nest-boxes without simultaneously quantifying the details of nest architectures (e.g., Burton 216 and references therein). Use of infrared cameras or data loggers placed next to or inside active nests could track dynamics in nest temperature or humidity in active nests of which both the nest architecture and breeding performance are quantified. 37 Most examinations of the physical properties of bird nests have been conducted ex situ in standardized laboratory conditions focusing on the cooling or heating of collected non-deconstructed nests or artificial objects (e.g., plastic eggs or temperature loggers) or non-specific eggs (e.g., eggs of non-domesticated Galliformes) placed inside the nest cup.40,165,215,217,218 A rarely used laboratory approach is to simulate the natural conditions of breeding by examining heating or cooling in fluctuating thermal conditions and to measure cooling or heating of collected eggs also taking species-specific incubation patterns into account. 215 These laboratory tests should control for the potential influences of debris (e.g., dust caused by nestling feather development) trapped inside the non-deconstructed nests.219,220
It is evident that the list of propositions for new research activities could be considerably extended by proposing studies of unstudied species, under-studied nest characteristics (e.g., texture, structural strength, nest chemistry), or under-studied fitness components (e.g., recruitment rates of locally born offspring, unstudied nestling physiology or chemistry). However, there is little point in suggesting a range of ideas that have not yet been (or may never be) investigated in a robust way. It is hoped that this review has demonstrated the range of factors that can affect nest functionality and its effects on Darwinian fitness. Moreover, it is hoped that, despite the challenges of working in the field, it will stimulate meaningful research into investigating how nest characteristics affect aspects of Darwinian fitness of species that nest outside cavities.
Supplemental Material
Supplemental Material - Nest design and breeding success in intraspecific investigations of non-cavity nesting avian species
Supplemental Material for Nest design and breeding success in intraspecific investigations of non-cavity nesting avian species by Marcel M. Lambrechts and D. Charles Deeming in Avian Biology Research
Footnotes
Acknowledgements
We are grateful to Jim Reynolds and an anonymous referee for the constructive comments on a previous version of the manuscript.
Author contributions
Writing -Original draft preparation M.M.L.; Writing – Review and Editing D.C.D. and M.M.L.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.