
Abstract
Monitoring steroid hormone patterns via excreted metabolites is an effective, non-invasive method for assessing physiological states in wild animals. This method allows for the monitoring of individuals or social groups with minimal disturbance. Here we aimed at elucidating the measurable glucocorticoids and androgens in excreta, for monitoring stress and reproductive hormones in the common waxbill (Estrilda astrild), a small estrildid finch. We measured the excreted metabolites in response to physiologically induced stress and reproductive challenges, both in males and females. For this, we challenged 12 captive common waxbills with adrenocorticotropic hormone (ACTH) and another 12 with luteinizing hormone-releasing hormone (LHRH). Steroid concentrations in droppings were analyzed using liquid chromatography mass spectrometry (LC-MS) before and up to six hours after each challenge. We found that enzymatic hydrolysis prior to LC-MS increased the recovery of steroid metabolites from common waxbills excreta. The ACTH challenge resulted in a significant increase of excreted glucocorticoids with a cortisone standard within 2-3 hours, suggesting that metabolites with a cortisone-similar structure are a reliable measure for monitoring physiological stress in this species. Although the LHRH challenge was inconclusive for androgens, our results provide useful technical details for measuring glucocorticoid metabolites in common waxbill excreta. This enables future studies of behavior in this bird species with minimum disturbance or handling bias and ensuring animal welfare requirements.
Keywords
Introduction
The common waxbill (Estrilda astrild) is a small estrildid finch native to sub-Saharan Africa that has become invasive and widely distributed in many parts of the world.1,2 This species has been successfully used as a model in diverse behavioral and physiological studies over the recent years,3–7 and the non-invasive monitoring of steroid hormone patterns will enable the study of new research questions in this species.
Non-invasive hormone measurement is a valuable alternative to traditional blood sampling in behavioural endocrinology research (mammals, 8 birds, 9 review 10 ). Measuring hormone concentrations in plasma can pose several challenges, which include potential confounding effects from handling and sampling stress, as well as health risks. In small animals, the limited volume of blood that can be sampled may limit analyses and still risk jeopardizing the health of animals.11–13 Faecal analysis has emerged as an effective alternative to overcome these issues, offering a non-invasive method of hormone measurement that has become more popular over the past years. It allows for long-term monitoring without influencing the behavioural or endocrine state of individuals, as droppings can be repeatedly collected from the same individual.8,14,15 Additionally, non-invasive methods allow tracking short-term hormone responses to social or environmental challenges, or to experimental exogenous treatments.16,17 Excreted hormone metabolites reflect cumulative hormonal secretion over several hours. 11 This may downgrade episodic hormone fluctuations compared to blood sampling, which might be influenced by the sampling procedure itself.13,17,18
Despite the numerous advantages of measuring excreted metabolites, one significant drawback is the need for prior validation to accurately assess changes in metabolite levels in any given species.17–19 Since excreted hormone metabolites consist of various metabolized compounds, in this study, initial validations focused on analytical processes to determine the most suitable extraction and quantification methods. Furthermore, faecal metabolites may differ across species and sexes, so physiological or biological validations are also essential. In particular, physiological challenges with releaser hormones are recommended to evaluate changes in excreted metabolite levels relative to baseline values. 17 This approach ensures the suitability of the assay employed for measuring endocrine activity.
Circulating steroid hormones are a diverse class of organic compounds composed of 17 carbon atoms arranged in a four-ring structure. 20 In response to a stressful event, vertebrates trigger a series of highly conserved neuroendocrine actions involving the hypothalamic–pituitary–adrenal (HPA) axis, which is regulated by adrenocorticotropic hormone (ACTH), and activates the release of glucocorticoid (GC) hormones. In avian species, corticosterone is the predominant GC hormone released along the HPA axis.17,21 Another key hormonal axis is regulated by luteinizing hormone-releasing hormone (LHRH) which activates the hypothalamic-pituitary-gonad (HPG) axis and regulates reproduction and social behaviour in all vertebrates. Activation of the HPG-axis leads to the release of progesterone, androgens (testosterone) and oestrogens (oestradiol) predominantly from the gonads into the blood. 22 Steroid hormones have low water solubility and generally bind to water-soluble proteins to facilitate their transport through the bloodstream to target tissues. 21 These hormones are primarily metabolized and conjugated in the liver and then excreted via urine and faeces,18,23,24 either in conjugated or non-conjugated forms, with variations depending on the species.25,26 Given potential differences between species in metabolism and excretion of steroid metabolites, it is crucial to validate the measured metabolites and techniques for each model species.
Here we test a non-invasive method for assessing hormonal activity by using liquid chromatography mass spectrometry (LC-MS) to measure steroid metabolites in common waxbills excreta. Although LC-MS has been widely used to study hormone profiles in primates and humans in clinical contexts,27,28 its application to study hormones in excreta is less common. Traditionally, steroid analysis from droppings has relied on group-specific immunoassays (in various vertebrate species, including birds 17 ) due to the group-specific antibody binding to functional groups of a metabolite (e. g. 11,17-OH) rather than a single steroid itself, 13 lower cost compared to LC-MS, 28 and ease of use when targeting a single (known) steroid. 29 However, immunoassays are limited to measuring only one hormone per sample, and may have high cross-reactivity (i.e., extension of the reaction of an antibody to others than its corresponding antigen 30 ), which may be a disadvantage for analytical precision. In contrast, LC-MS offers superior differentiation between similar compounds,27,31,32 effectively reducing cross-reactivity issues inherent to immunoassays. 33 It may also be a valuable tool for further characterizing the steroid metabolites detected in excreta by immunoassays. 13 Moreover, LC-MS has the advantage to allow simultaneous measures of multiple steroids from both the HPA and HPG axes in a single analysis, including various metabolites. 20 Given these benefits, we opted for the LC-MS method to identify and quantify steroid hormone metabolites in excreta.
Our common waxbills’ faecal steroid measurements comprised four steps, with the first three focused on analytical validation:
1. Identification of main steroid metabolites and physiological ranges in common waxbill excreta: this initial step involved scanning preliminary non-hydrolyzed excreta samples to identify key steroid metabolites and establish their concentration ranges. A standard commercial assay was used to scan for immunoreactive evidence of 23 common steroid hormones (Thermo Fisher Scientific) in common waxbill excreta.
2. Assessment of the lyophilization process: this step evaluated the effectiveness of freezing methods for lyophilization, by determining recovery rates of steroid metabolites in common waxbills excreta.
3. Examination of conjugation patterns via enzymatic hydrolysis: given that species differ in how steroids are excreted, this step is crucial for measuring biologically relevant metabolites in excreta. Enzymatic hydrolysis was used to deconjugate steroid metabolites, and then steroid recovery was compared between non-hydrolyzed and hydrolyzed samples.
4. Biological relevance: If the metabolites measured in common waxbill excreta are biologically relevant, significant changes of GC-metabolites should be detected when inducing hormonal release through an ACTH challenge, 24 and changes in sex steroid metabolites should be detected when inducing hormonal release through a LHRH challenge, in both sexes. 34
Methods
The analytical and biological tests were conducted using 24 common waxbills, 12 males and 12 females, captured in February 2017 from agricultural areas near Póvoa de Varzim, in northern Portugal. The birds were housed in same-sex groups of four individuals across three large bird cages (88.5 cm × 30 cm × 40 cm) located in an outdoor aviary. The aviary had natural temperature and ventilation, was protected from direct sunlight but had natural light supplemented with ceiling lamps in a natural light-dark cycle. The birds were given ad libitum access to food (commercial seed mixture for exotic birds, Tropical Finches, Australian Finches, Versele-Laga) and drinking water. Additional water containers provided bathing two or three days per week. Prior to the start of the experiments, the birds underwent a four-month acclimation period in the cages. Capture of birds was authorized by permit number 57/2017/CAPT from the Instituto para a Conservação da Natureza e das Florestas, and animal procedures were approved by Orbea committee at CIBIO-InBIO (reference no. 2017-01).
For analytical steps 2 and 3, faecal samples were collected from the 12 birds, six males and six females. Each bird was temporarily isolated in its own cage, which was divided into two sections using a plastic partition. We lined the floor of the cage with a clean sheet of aluminum foil placed below the cage floor grid. We sampled 30-minute periods during the morning, until obtaining droppings from a total of 48 30-minute periods (24 for males and 24 for females). We pooled all droppings from the same bird in the same 30-minute period, and hereafter refer to this as a faecal sample.
To test the biological relevance of the measurements of excreted steroid metabolites (step 4), we performed two experimental manipulations: one targeting adrenocortical activity (HPA axis) through the administration of ACTH, and the other targeting gonadal activity (HPG axis) via the administration of LHRH. The ACTH and LHRH challenges are well-established methods for validating the non-invasive measurement of excreted steroid metabolites. Our primary objective was to determine which most common steroids and metabolites are excreted, in either sex, and establish a reliable method for monitoring the species across different contexts.
Analytical measurement: Step 1
As an initial screening step intended to identify steroid metabolites, faecal samples of common waxbills were extracted and purified as described below, and then analysed by LC-MS. We then performed a LC-MS for the analysis of androgenic and estrogenic steroids, which followed a Thermoscientific validated method with some modifications. 35 Samples were injected in a Liquid Chromatograph Thermo Finnigan Surveyor HPLC System (Thermo Scientific, MA, USA), coupled with a Mass Spectrometry LCQ Fleet™ Ion Trap Mass Spectrometer (Thermo Scientific, MA, USA). Data acquisition and processing was made with XcaliburTM version 2 software (Thermo Fisher Scientific, MA, USA). Mass Spectrometer Tune Method parameters optimization was performed through direct injection of a standard mixture of 18-OH-Corticosterone, Cortisone, 11-Deoxycorticosterone, Testosterone, and Androsterone (Tables S1 and S2) solution of 1 ppm in LCMS methanol. Mass Spectrometer operated in electrospray positive polarity mode using Collision Ionisation Dissociation (CID) corresponding to the [M+H]+ molecules ion precursors and respective diagnostic fragments. The spray voltage maintained at 3.5 kV; Capillary temperature at 275°C; Capillary voltage and tube lens maintained at 20 and 68 kV, respectively. Nitrogen was used as sheath gas at 30 and as auxiliary gas at 10 (arbitrary units). Helium was used as buffer gas. When in Colission Induced Dissociation (CID) Mode, collision energy was at 16 to 25 eV. Separation was achieved on a Acccore C18 (100 × 2.1 mm I.D., 2.6 μm, Batch: 15157, Thermoscientific) kept at 30°C, with a flow rate of 0.3 mL/min. and injected volume was 10 μL in a no waste mode. The eluents used were methanol (A) and water (B) both acidified with formic acid at 0.1% (v/v). The gradient program started at 30% A, increasing to 40% A in 10 min, and to 90% in 15 min turning back to 20% in 1 min and to the initial conditions in 2 min, equilibrating more 8 min with 20% A. For retention time and mass parameter information please see Tables S1 and S2.
We used a standard commercial 23-steroids analysis kit (Thermo Fisher Scientific), which is designed to analyse the most common and best candidate steroids and metabolites for research purposes. While we acknowledge that some biologically relevant steroids and metabolites may not be included in this panel, the kit was selected for its practicability and broad applicability. Similar analyses were also performed in killer whales, Orcinus orca, faecal samples. 36 The analysed steroids were: Aldosterone, Allopregnanolone, Androstenedione, Androsterone, Corticosterone, Cortisol, Cortisone, 11-Deoxycorticosterone, 11-Deoxycortisol, 21-Deoxycortisol, 18-Hydroxycorticosterone, Dehydroepiandrosterone, Dehydroepiandrosterone-Sulfate, Dihydrotestosterone, Etiocholanolone, Pregnenolone, 17-Hydroxy-Pregnenolone, Progesterone, 17-Hydroxy-Progesterone, Testosterone, Estradiol, Estriol and Estrone.
Following the analysis of the 48 samples obtained from both males and females, for the analytical step, we calculated the limit of detection (LD): the lowest concentration that can be detected with statistical significance; limit of quantification (LQ): the lowest amount of analyte in a sample which can be quantitatively determined with suitable precision and accuracy; and total mean area (Area) for each sample.
Analytical measurement: Step 2
Since bird excreta contains both faeces and urine, they can vary in consistency. To evaluate the lyophilization process, after sample collection, half of the samples (12 from males and 12 from females) were lyophilized, by freeze drying samples at −80°C, while the other half were frozen at −20°C as controls.
All samples, both lyophilized and non-lyophilized, were subjected to the extraction process, during which steroids were extracted using 60% methanol in double-distilled water. The samples were vortexed for 15 minutes and centrifuged at 1500 g for 10 minutes, after which the supernatant was collected. This extraction protocol is widely used for extracting steroid hormones from droppings.13,18,37
Analytical measurement: Step 3
Steroids are extensively metabolized in the liver and the gut and may be excreted either in non-conjugated form or conjugated to more polar substances, such as sulphates or glucuronides. 38 To assess if quantification of recovered excreta steroids increased, we performed enzymatic hydrolysis to cleave the conjugated steroids. Half of the samples (6 lyophilized and 6 non-lyophilized from males, 6 lyophilized and 6 non-lyophilized from females) underwent enzymatic hydrolysis using β-glucuronidase-arylsulfatase (from Helix pomatia, Roche), while the other half served as control. For enzymatic hydrolysis, 100 µl of enzymatic mixture (1:250) in sodium acetate buffer (0.1M, ph 4.8) was added to 100 µl of each sample. The tubes were then sealed and incubated overnight at 37°C to allow for optimal enzyme activity. This process ensures the conversion of conjugated steroids into non-conjugated forms, enabling a more accurate measurement of total steroid concentrations with LC-MS in the samples.
Biological relevance: Step 4
The biological relevance experiments were conducted between June 20 and July 17, 2017. For the experiment, each bird was isolated in one side of its home cage, divided by a plastic partition. The cage floor was covered with aluminium foil. To determine the baseline levels of steroid metabolites, droppings were collected from the aluminium foil between 7a.m. and 9a.m. (UTC) at 30-minute intervals. Each collection was performed by pulling the lower drawer of the cage without disturbing the birds, and the aluminum foil was replaced. At 9 a.m., each bird was briefly removed from its cage to receive an intramuscular injection of either ACTH or LHRH (details below). Handling time was two minutes. Following the injection, droppings were collected at 30-minute intervals over the next six hours and immediately frozen at −20°C. In the lab, these samples were lyophilized, extracted and all were enzymatically hydrolyzed to cleave conjugated steroids as described above.
For the ACTH challenge, six males and six females were injected intramuscularly with 1.8 IU of ACTH (Adrenocorticotropic Hormone from porcine pituitary powder, A6303-100, Sigma-Aldrich), dissolved in 20 µl of saline solution. The dosage was estimated based on the mean weight of common waxbills (ca. 9 g in captivity), following studies in other avian species (white-crowned sparrows, Zonotrichia leucophrys gambelii; 39 European stonechats, Saxicola torquata rubicola37,40). In passerine birds, a dosage of 0.2 IU/g of bird weight is widely accepted as effective in stimulating the pituitary system. 41
For the LHRH challenge, six males and six females were injected with 0.5 µg of LHRH (Luteinizing hormone releasing hormone, L0637, Sigma-Aldrich) dissolved in 20 µl of saline solution. This dosage, like the ACTH dosage, was estimated based on body mass, with reference to studies in other passerine species (northern cardinals Cardinalis cardinalis, 42 European stonechats,37,40 dark-eyed juncos Junco hyemalis 43 ).
Statistics
For the analytical validation, we ran a General Linear Mixed Model (GLMM), with a normal distribution, to evaluate the effectiveness of the lyophilization (Step 2) and deconjugation (Step 3) steps. The dependent variable was the percentage of mean area. The fixed factors included were sex (male, female), lyophilization process (lyophilized samples, non-lyophilized samples), and enzymatic hydrolysis (hydrolyzed samples; non-hydrolyzed samples). Mean Area was log transformed log10 (x + 1) to approach a normal distribution.
For the biological validation, we collected a total of 244 faecal samples (i.e., sets of droppings, per bird, over a 30-min period), 114 samples for the ACTH challenge and 130 for the LHRH challenge. Due to technical issues, we were unable to recover steroid metabolites from 22 samples in the ACTH challenge and 25 in the LHRH challenge. This resulted in a final dataset of 197 samples (92 for the ACTH challenge and 105 for the LHRH challenge). On average, we collected 12.3 ± 0.52 droppings per hour (mean ± SE), with rates of 11.5 ± 0.73 samples per hour for the ACTH challenge and 13.1 ± 0.62 samples per hour for the LHRH challenge, approximately one dropping per bird per hour. For statistical analysis, we defined each hour as a time period and grouped the two hours before the injection as a single time period also. In the ACTH treatment, one bird had only two measurements and was removed from the analyses, resulting in 11 birds (5 males and 6 females). Due to highly non-normal distribution of the variables, we ran non-parametric Friedman tests. These tests analyzed hormone measurements as dependent variable, grouping data by individual bird, and using time periods as repeated blocks (pre-injection, 0 to 1 hour post-injection, 1 to 2 hours post-injection, etc.). When there were more than one faecal sample available per bird per time period, we averaged those samples. All statistical analyses were performed using R Software v. 4.4.0 (R Core Team 2024).
Results
Analytical measurement
Following the analysis of the 48 samples obtained from both males and females, we were able to recover and identify 18 out of the 23 initial candidate steroids. Out of these, 11 steroids were reliably detected in all samples, with an Area/LQ > 2, and were thus confirmed as good candidates to be used in the biological measurement. This group of 11 steroids contained five GC steroids (Table S1) and six reproductive steroids (Table S2). Among these we chose five metabolites in the excreta to be analyzed based on reliability of measurements and biological relevance: for GC we chose 18-OH-corticosterone, cortisone and 11-deoxycorticosterone; and for reproductive steroids we chose testosterone and androsterone (Tables S1 and S2). Of the five target hormones, androsterone was underneath detection limits in the samples for biological validation. Therefore, the analyses were restricted to four hormones (18-OH-corticosterone, cortisone, 11-deoxycorticosterone and testosterone).
The mean area for total steroid metabolites recovered did not differ between samples that were or were not lyophilized prior to storage at −20°C (F1, 44 = 0.28, p = 0.60), nor between the sexes (F1,44 = 0.79, p = 0.38, Figure 1). In contrast, enzymatic hydrolysis significantly enhanced the recovery of steroid metabolites from common waxbill excreta, with hydrolyzed samples showing more than four-fold increases in the mean area of metabolites recovered than non-hydrolyzed samples (F1,44 = 84.23, p < 0.001, Figure 1). Percentage of mean areas for total steroid metabolite recovery, from common waxbills excreta, analysed by LC-MS. Bars show means ± standard errors. The data are grouped by type of sample processing (control; lyophilization; enzymatic hydrolysis; lyophilization plus enzymatic hydrolysis), and by sex. N = 24 samples per sex (6 per sex per treatment).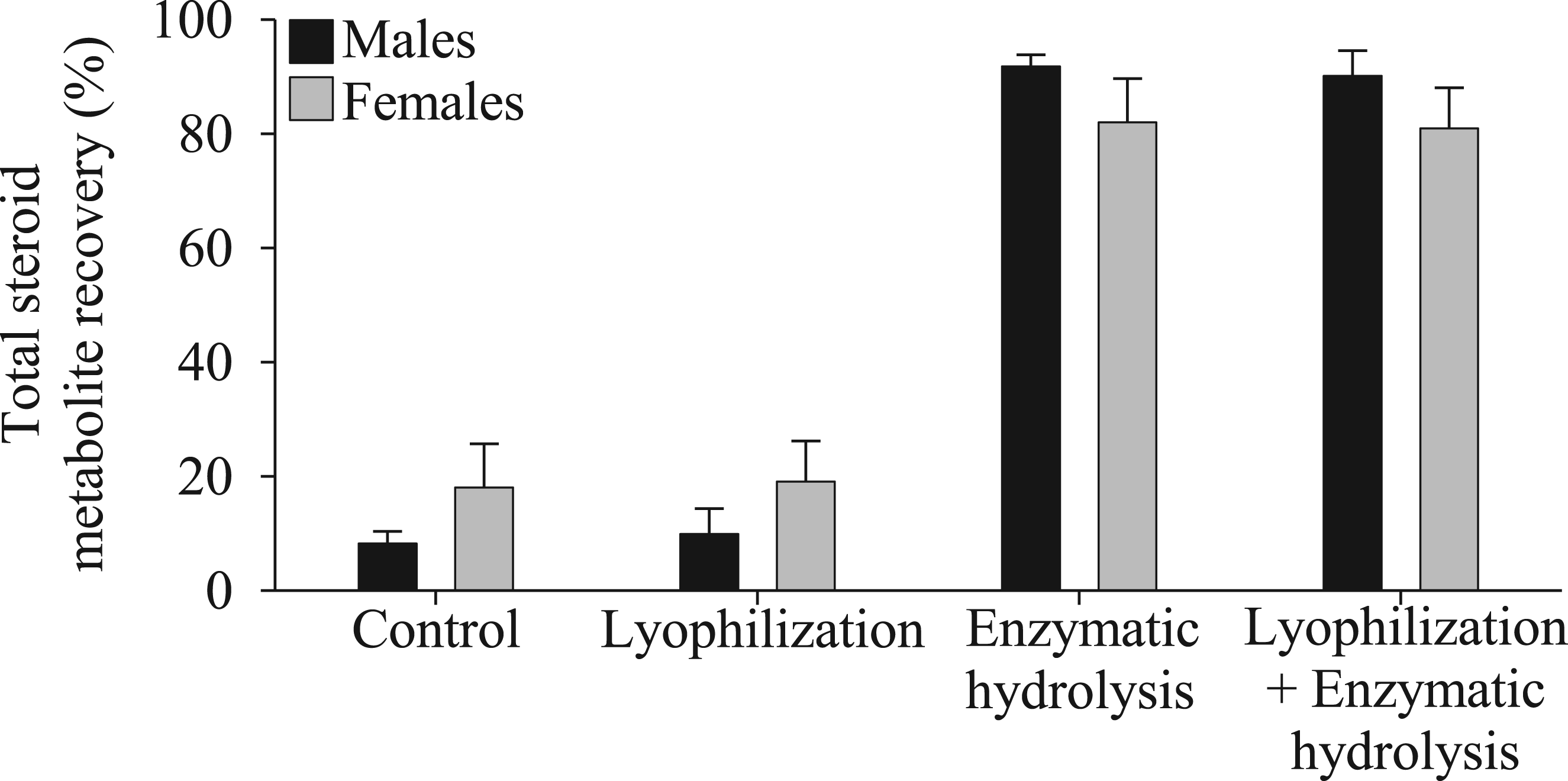
Biological relevance
The exogenous administration of ACTH led to an increase in some GC metabolites from hydrolyzed faecal samples. The peak concentration of ACTH-induced cortisone metabolites occurred between two and three hours after treatment (Figure 2(b)), with significant differences between sampling times (χ2 = 13.97, p = 0.030, d.f. = 6). For 18-OH-corticosterone, data also suggested a peak about two hours post-ACTH treatment (Figure 2(a)), but differences among sampling times were not statically significant (χ2 = 7.75, p = 0.257, d.f. = 6). ACTH treatment did not cause significant changes of the measured patterns of 11-deoxycorticosterone or testosterone (11-deoxycorticosterone: χ2 = 3.92, p = 0.687; testosterone: χ2 = 7.14, p = 0.304, d.f. = 6, Figure 2(c) and (d)). Profiles of excreted metabolite concentrations in common waxbills Estrilda astrild injected intramuscularly with ACTH (ACTH challenge). Dots represent values from individual birds, and bars represent medians (± interquartile ranges), from before injection (injection at time 0) until six hours post-injection.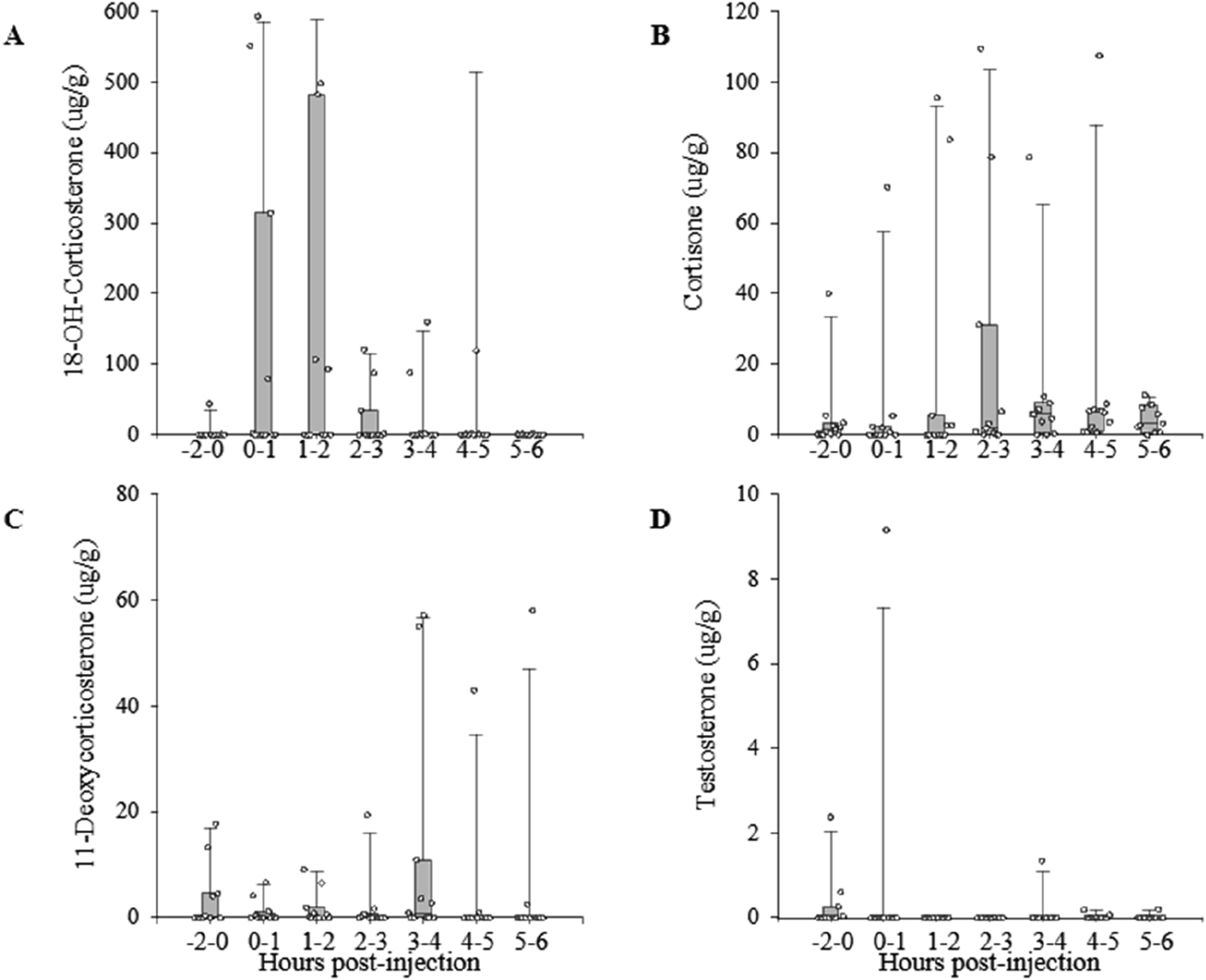
We did not detect significant changes of testosterone metabolites levels in common waxbills’ excreta within six hours after the LHRH challenge (χ2 = 2.10, p = 0.910, d.f. = 6, Figure 3(d)). LHRH treatment also did not result in significant differences of GC metabolite levels across sampling times (18-OH-corticosterone: χ2 = 2.81, p = 0.832; cortisone: χ2 = 7.00, p = 0.320; 11-deoxycorticosterone: χ2 = 8.07, p = 0.233, d.f. = 6; Figure 3(a)–(c)). Profile of excreted metabolite concentrations in common waxbills Estrilda astrild injected intramuscularly with LHRH (LHRH challenge). Dots represent values from individual birds, and bars represent medians (± interquartile ranges), from before injection (injection at time 0) until six hours post-injection.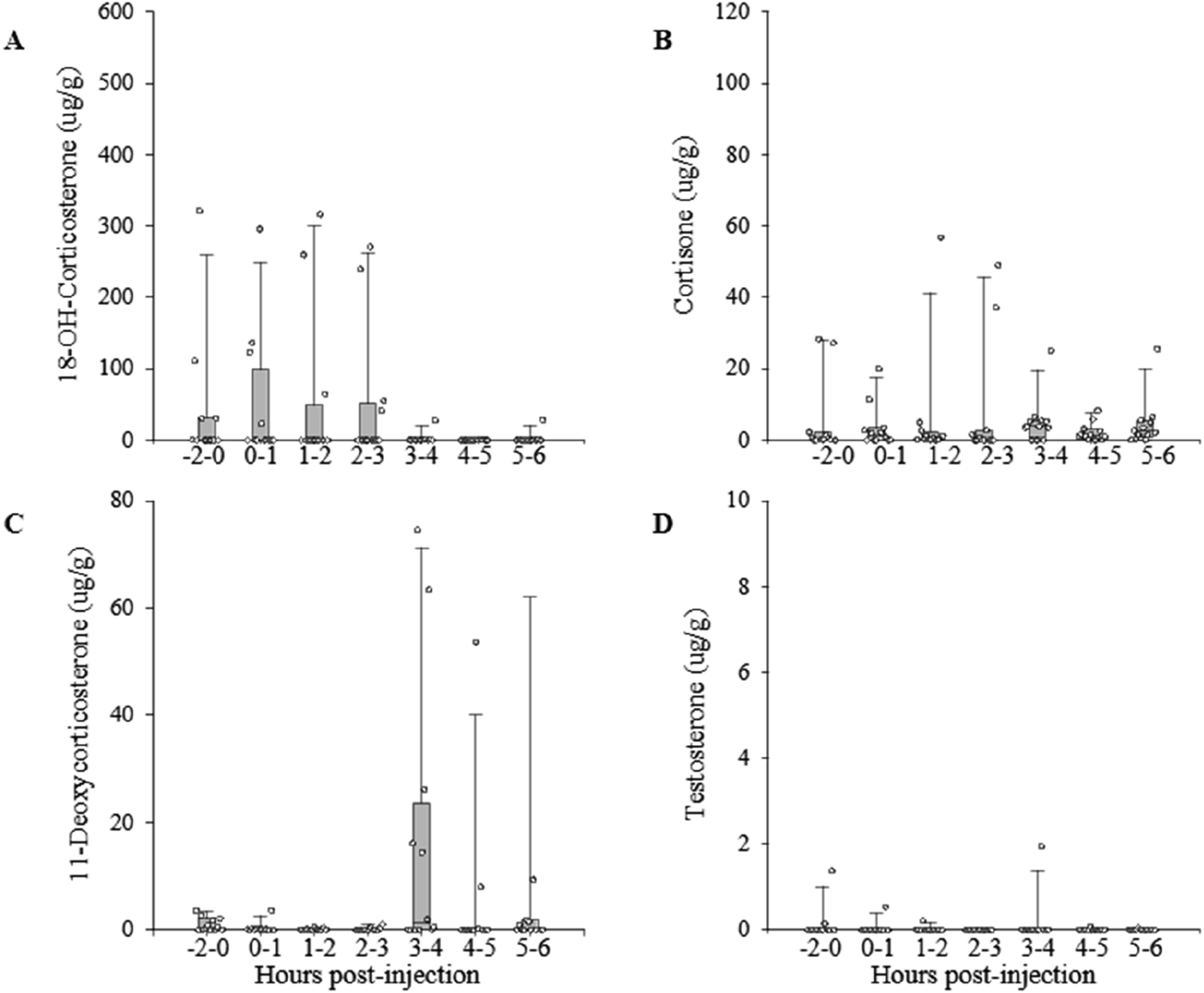
Discussion
The objective of this study was to test a new method for measuring steroid metabolites in droppings of common waxbills, Estrilda astrild. Given species-specific differences in the metabolism and excretion of GC, it is necessary to assess the suitability of the methods used for each individual species.17,44 We successfully validated a method for measuring GC metabolites in common waxbill excreta with LC-MS, and cortisone was identified as the primarily excreted GC metabolite following an ACTH challenge. We chose to use the LC-MS analysis to screen for specific steroids, as we were unaware of which GC were present in common waxbills excreta.
A cortisone enzyme immunoassay has been identified as providing a meaningful proxy of excreted GC metabolites in other species, including chicken, Gallus domesticus, 45 capercaillie, Tetrao urogallus, 46 and Japanese quail, Coturnix japonica 47 . However, these studies employed group-specific EIA for 3,11-dioxo GC metabolites (rather than cortisone itself), whereas the results of our study are based on LC-MS with a specific standard for cortisone. It is indeed unlikely that cortisone derives from circulating corticosterone in common waxbills’ blood, as it is a precursor and derivate of cortisol, not corticosterone. However, the observed response patterns clearly indicate that the LC-MS detected a GC-metabolite similar to cortisone. Therefore, we suggest that future assays based on cortisone standards may serve as a non-invasive and reliable approach for monitoring stress in common waxbills.
However, the LHRH challenge did not yield sufficient metabolite recovery to be used as a monitoring tool for testosterone or other steroids measured in this study of common waxbill excreta.
The analytical validation also demonstrated that enzymatic hydrolysation of samples prior to LC-MS analysis is a crucial step in recovering steroid metabolites from common waxbill excreta. Enzymatic hydrolysation significantly enhanced metabolite recovery, suggesting that GC metabolites in common waxbills are predominantly excreted as conjugated forms, which need to be cleaved before LC-MS can efficiently recover steroid metabolites. This methodological finding aligns with the results of similar studies on GC metabolites in bird droppings, including great tits, Parus major, 48 European stonechats, Saxicola torquata rubicola, 40 and Adélie penguins, Pygoscelis adeliae 49 .
We could not detect significant differences of the tested steroid hormones between sampling times in response to the LHRH challenge, and the recovery of testosterone was extremely low. This prevents, for the moment, using LC-MS for measuring excreted sex steroids in common waxbills. The lack of detectable amounts of sex steroid metabolites might be attributed to stress associated with the isolation, injection, and sample collection during the LHRH challenge (Figure 3(a)). Stress will activate the HPA-axis and may have caused negative feedback within the HPG axis, decreasing secretion of sex steroids from the gonads.50,51 As such, an increase in GC is often correlated with decreased sex steroids levels, as observed in amphibians, 52 birds 53 and mammals54,55. However, this was not observed in the pattern of excreted cortisone metabolites after LHRH (Figure 3(b)) and it also differed from cortisone after ACTH (Figure 2(b)). Another possible explanation could be seasonal or sex-specific differences in responsiveness to the LHRH challenge, which may depend on the presence of LHRH receptors in target organs. 56 Additionally, other authors reported variation of individual sensitivity to LHRH due to factors such as the social environment, 57 fitness or season16,43,57. For waxbills, this remains to be understood.
After the ACTH challenge, and after enzymatic cleavage of acid rests such as sulphates or glucuronides, the primary GC measured in common waxbill excreta was cortisone, which was also the only metabolite to show significant differences across sampling times. The excretion pattern of cortisone revealed a prominent peak, at two to three hours post-injection. The timing and duration of this peak is consistent with excretion rates reported from other bird species, with the onset of excretion being due to a first peak of steroids from renal origin, and its permanence over an hour or more due to a second peak of steroid metabolite excretion via the gut.24,45,46 Excretion of urine is expected to be faster than intestinal excretion, and both urine and faecal matter are excreted jointly through birds’ cloaca, which may explain the long peak of cortisone in our results. In addition to the physiological response to ACTH, stress may also have been induced by the procedure itself (i.e., isolation, handling and injection), despite of the fact that handling was brief. 58 Nonetheless, the absence of a statistically significant cortisone peak in the LHRH challenge, where birds were also isolated and handled, indicates that the ACTH challenge was at least in part responsible for producing a physiological stress response.
Our results indicate that it is feasible to develop a specific antibody-based immunoassay to measure GC-metabolites similar to cortisone in common waxbills’ excreta, as a means for monitoring stress responses non-invasively. This would offer a quicker, simpler, and more affordable assay method for stress assessment in this species. Either way, the method presented here enables future studies of individual stress patterns in a social context, such as personality, dominance, cognition and other behavioral traits of common waxbills.
Supplemental material
Supplemental material - Measurement of excreted steroid metabolites in common waxbills Estrilda astrild
Supplemental material for Measurement of excreted steroid metabolites in common waxbills Estrilda astrild by Sandra Trigo, Joana Azevedo, Pedro Reis, Ana Cristina R. Gomes, Gonçalo C. Cardoso, Katharina Hirschenhauser, Marta C. Soares in Avian Biology Research
Footnotes
Funding
This work was supported by Fundação para a Ciência e a Tecnologia (FCT) with project funds from Fundo Social Europeu: PTDC/BIA-COM/2644/2020 (doi: 10.54499/PTDC/BIA-COM/2644/2020) to ST, and PTDC/BIA-ECO/32210/2017 to GCC. ST was supported by FCT CEEC IND5ed (doi: 10.54499/2022.03732.ceecind/cp1730/ct0009); ACRG by PTDC/BIA-ECO/32210/2017; GCC by FCT CEEC IND5ed (doi: 10.54499/2022.07867.CEECIND/CP1730/CT0005); and MCS by CEEC Ind2021 (doi: 10.54499/2021.01458.CEECIND/CP1668/CT0003).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.