
Abstract
Haemosporidians (Plasmodium, Haemoproteus, and Leucocytozoon) are blood parasites that infect birds and other vertebrates worldwide. Infections can cause disease that reduces survival and reproductive success, and may even lead to mortality. They are primarily assessed through prevalence, a population parameter that varies among species, regions, and seasons due to environmental and ecological factors. Although meta-analyses on this parameter exist, none globally synthesize data across the three haemosporidian genera and current detection methods (PCR and microscopy). The objective of this systematic review was to determine the overall prevalence of haemosporidian parasites in wild birds distributed across different zoogeographic regions. The review was conducted following the methodology proposed by the Joanna Briggs Institute (JBI) for scoping reviews and was reported according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses extension for Scoping Reviews (PRISMA-ScR) to estimate pooled prevalence and assess variation according to diagnostic method, continent, parasite genus, and bird family. The global pooled prevalence was 12.3%, with estimates of 12.1% by PCR and 13.1% by microscopy. Regardless of the diagnostic method, the continents with the highest prevalence were Africa (PMIC = 22.3%; PPCR = 13.5%) and Europe (PMIC = 28.6%; PPCR = 12.6%). The high heterogeneity among studies suggests that multiple biological, environmental, and methodological factors contribute to global variation in infection patterns. This study provides the first global reference of haemosporidian parasite prevalence in wild birds and highlights the need for standardized reporting of environmental and methodological variables to better understand the drivers of avian malaria dynamics.
Introduction
Haemosporidians are intracellular protist parasites belonging to the phylum Apicomplexa [1]. They infect a wide range of vertebrate hosts, including birds, mammals, and reptiles1,2. Their heteroxenous life cycle requires both a vertebrate host and a hematophagous dipteran vector, facilitating their broad global distribution1,3. These infections are of considerable ecological and epidemiological relevance, as they can cause anemia, reduced survival and reproductive success, and even high mortality in host populations1,4–7.
Prevalence, defined as the percentage of infected individuals relative to the total population at a given time 8 , is an important metric for assessing the impact and dynamics of these parasites. 9 It can be assessed at different biological levels, including within a single host species or across multiple species within a community 10 . Its magnitude can vary widely (0–100%) among species, regions, and seasons, depending on abiotic factors such as altitude and climatic conditions that favor transmission, as well as biotic factors such as vector availability and host exposure11–13. Additionally, migratory birds often exhibit higher prevalence and greater diversity of parasite lineages, highlighting the role of host movement in dispersing these parasites 14 .
Clark et al. (2014) highlighted that avian haemosporidians exhibit diversity patterns similar to those of their avian hosts, a key finding for understanding lineage diversity and host–parasite interactions in Plasmodium and Haemoproteus. Although this synthesis focused on parasite diversity, which is essential for elucidating evolutionary and phylogenetic patterns, it does not quantify infection prevalence in wild birds, a population parameter 8 required to understand the distribution of each parasite genus, identify the factors driving their transmission, and evaluate their implications for the conservation and health of wild bird populations15,16.
Quantitative syntheses of haemosporidian prevalence are comparatively limited. To date, only two systematic reviews with meta-analysis have been published. The first focused exclusively on Plasmodium infections in wild birds 17 . More recently, Swangneat et al. 18 expanded the analysis to include the three main haemosporidian genera; however, they restricted inclusion to PCR-based studies conducted in Southeast Asia, most of which involved domestic or captive birds. Consequently, pooled prevalence estimates integrating Plasmodium, Haemoproteus, and Leucocytozoon across wild bird populations at a global scale remain lacking, particularly those that explicitly account for differences between diagnostic approaches (microscopy and PCR).
Synthesizing the prevalence of haemosporidian parasite infections substantially contributes to the ecological and epidemiological understanding of avian malaria. Prevalence not only represents the proportion of infected individuals, but also serves as an indirect indicator of transmission intensity and host–parasite dynamics across different environmental and biogeographic contexts 19 . Integrating estimates at a global scale allows the identification of spatial gradients, the assessment of potential environmental and methodological effects, and the detection of heterogeneity patterns that are not evident in individual studies. Such syntheses also provide comparable baseline values across regions and taxa, facilitate evaluation of impacts on wild population health, and help identify geographic and taxonomic gaps that can guide future research and surveillance strategies.
Therefore, the aim of this study was to conduct a systematic review and meta-analysis of haemosporidian parasite prevalence in wild birds, assessing variation by diagnostic method, continent, parasite genus, and bird family, while providing a population level perspective.
Materials and methods
Inclusion criteria
Participants
The review considered studies conducted with blood samples from wild birds collected in the field or stored in blood banks. In studies that involved both wild and captive or domestic birds, they were considered only if the authors reported data for each group separately.
Condition
This review included studies on the prevalence of haemosporidian parasites, which are blood parasites that require a vector (dipteran) and a host (bird) to complete their life cycle, thereby affecting the health of infected individuals 20 .
Context
This review did not apply any geographical restrictions in the selection of studies.
Types of studies
The review included observational cross-sectional studies that assessed natural infections in wild birds that used molecular or microscopic techniques for the detection and identification of haemosporidian parasites. Experimental infection studies involving artificial inoculation under laboratory conditions were excluded. We also considered both studies published in peer-reviewed scientific journals and grey literature obtained through systematic searches in specialized databases or repositories.
The systematic review was conducted in accordance with the methodology proposed by the Joanna Briggs Institute (JBI) 21 and reported in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guidelines 22 .
Search strategy
The objective of the search strategy was to identify both published studies and grey literature in three steps. (1) A search was conducted in PUBMED, followed by an analysis of the keywords contained in the title and abstract, as well as the index terms used to describe the article. (2) A second search was performed using all identified keywords and index terms across all included databases. (3) We examined the reference lists of all articles included in the review to locate additional studies.
The resources consulted included PubMed, ScienceDirect, Web of Science, Scopus, ProQuest (ProQuest Central, Academic Complete, Academic Video Online, and ProQuest Dissertations & Theses), LILACS, and Google Scholar. The grey literature search included the repository La Referencia. The initial keywords used were: birds, wildlife, blood sample, and haemosporidian parasites. We considered only studies published in English, Spanish, and Portuguese for inclusion in this review. We did not apply any time restrictions to the searches. We detail the search equations used for each consulted resource in S2 Table. Below is the equation used in PubMed:
1. birds[MeSH Terms]
2. Aves[Title/Abstract] OR birds[Title/Abstract] OR avian[Title/Abstract].
3. #1 OR #2.
4. Animals, Wild[MeSH Terms].
5. (Wild[Title/Abstract] OR “Non-domesticated”[Title/Abstract] OR nondomesticated[Title/Abstract] OR undomesticated[Title/Abstract] OR feral[Title/Abstract] OR stray[Title/Abstract].
6. #4 OR #5.
7. Prevalence[Title/Abstract].
8. Haemosporida[MeSH Terms] OR malaria, avian[MeSH Terms].
9. Haemosporidia[Title/Abstract] OR Apicomplexa[Title/Abstract] OR Plasmodium[Title/Abstract] OR Leucocytozoon*[Title/Abstract] OR Haemoproteus[Title/Abstract] OR “Avian haematozoa”[Title/Abstract].
10. #8 OR #9.
11. #3 AND #6 AND #7 AND #10.
All searches were conducted independently by two reviewers to ensure the reproducibility and methodological rigor of the process.
Study selection
The search results were managed in Mendeley (version 2.77.0) in order to remove duplicate records. Subsequently, we carried out document screening using the Rayyan program
23
. Two reviewers (TBT and LJR) independently examined the titles and abstracts of the retrieved studies, based on the previously defined inclusion criteria, to determine their eligibility for full-text review. In cases where discrepancies arose between reviewers during the selection process, the third reviewer (FRG) resolved them. We present the results of the searches and the study selection process in a flow diagram, in accordance with the PRISMA guidelines (Figure 1). Flow diagram of the study design process.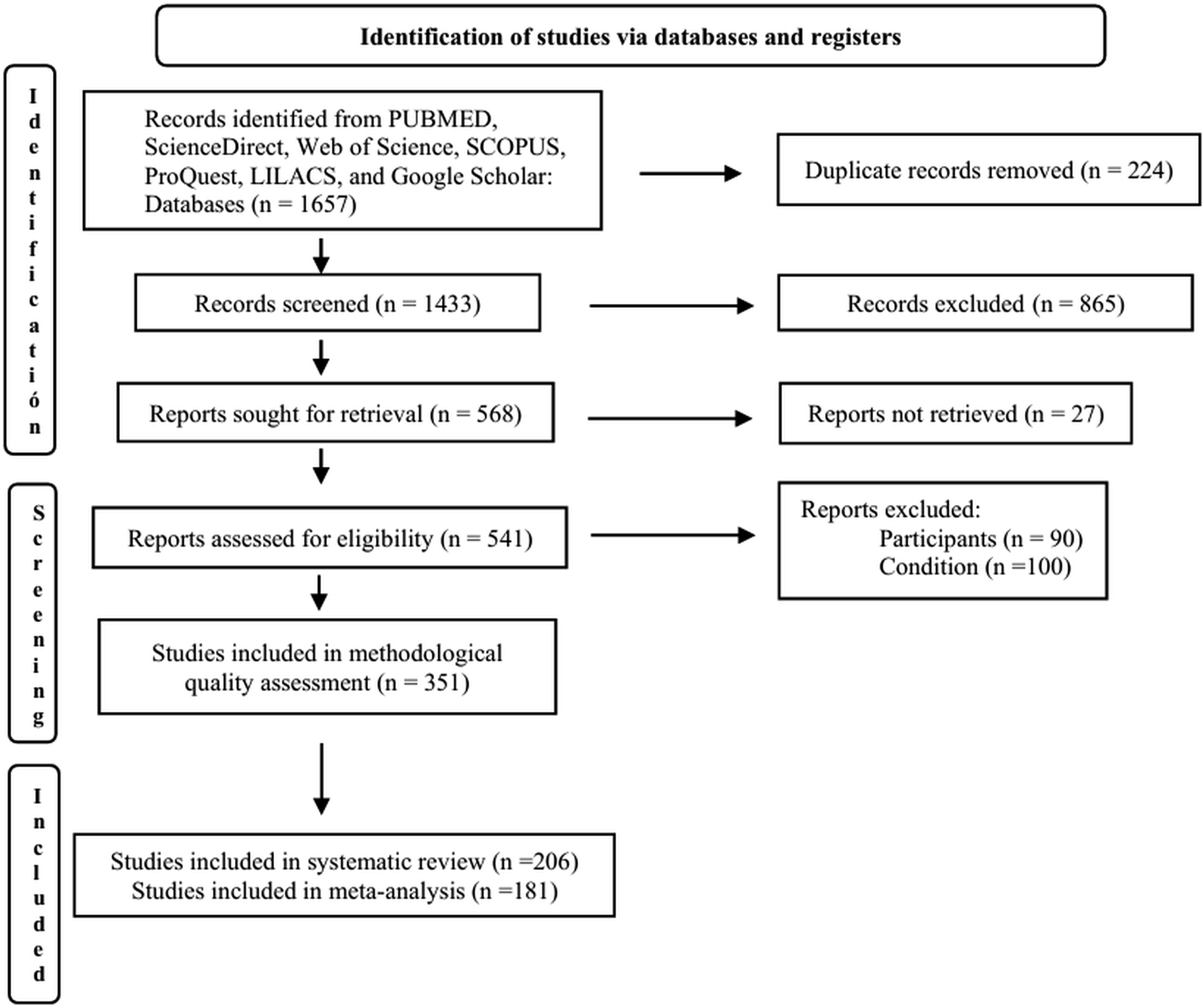
Methodological quality assessment
Two reviewers independently assessed the methodological quality of the studies that met the inclusion criteria. For this evaluation, the checklist for prevalence studies developed by the Joanna Briggs Institute (JBI) was adapted 22 . The questions considered in the evaluation were: Were the study participants sampled appropriately? Was the sample size adequate? Were the study subjects and setting described in detail? Were valid methods used to identify the condition? Was the condition measured in a standardized and reliable way for all participants? Was an appropriate statistical analysis conducted?
To ensure the methodological quality of the studies included in the review, a minimum criterion was established requiring each study to meet at least four of the six items adapted from the Joanna Briggs Institute (JBI) checklist for prevalence studies. This decision was based on the need to minimize the risk of bias and ensure that the estimates obtained came from studies with an acceptable level of scientific rigor. The authors assumed that including only studies that met at least four of the six evaluated criteria would allow the analyzed sample to be composed of research that, although potentially subject to certain limitations, met basic standards of validity in fundamental aspects such as sampling, sample size, validity of diagnostic methods, and statistical analysis.
Data extraction
Descriptive and relevant data from the studies included in the review were extracted using a standardized table created in an Excel spreadsheet S3 Data. One reviewer performed the extraction and subsequently verified it by a second reviewer to ensure the accuracy and consistency of the collected information.
Data analysis and presentation
The data extracted from the included studies were used to perform the meta-analysis using R software (v. 2025.05.1+513). The extracted variables included: author and year of publication, total sample size (n), number of infections of positive individuals, continent, diagnostic methodology (PCR or microscopy), host family, parasite species, and geographic and environmental variables (altitude, latitude, longitude, and temperature). When studies included multiple host species within a community, prevalence data were extracted and analyzed separately for each species, treating each as an independent record. These variables were used to calculate prevalence estimates and to explore potential sources of heterogeneity through subgroup analyses and meta-regression models.
For the meta-analysis, studies with extreme prevalence values (1% or 100%) and those with fewer than 50 individuals were excluded. This exclusion is based on the fact that small samples tend to generate unstable estimates with high variability, thereby increasing the risk of random error 24 . In the context of prevalence studies, small sample sizes may yield results that are not representative of the target population, thereby reducing the precision of the estimates. Studies with very small sample sizes often have low statistical power, which makes it difficult to detect true patterns and compare them with those of other studies included in the meta-analysis 25 . For these reasons, a minimum threshold of 50 individuals was established to improve the quality and consistency of the analyzed data. Heterogeneity among studies was statistically assessed using the I2 test, where estimates above 75% were considered indicative of significant heterogeneity. Likewise, the τ2 statistic was estimated to indicate the actual variance among study effects 26 . A random-effects model was employed to synthesize prevalence values using the R packages meta (version 8.2-1) 27 and metafor (version 4.8-0) 28 . Subgroup analyses were conducted to assess whether family, identification method, and biogeographic region influenced infection prevalence, using Cochran’s Q test to evaluate these differences 29 in the meta package.
Meta-regression analysis were conducted to evaluate the effect of publication year, temperature, altitude (meters above sea level), and latitude on prevalence estimates derived from PCR and microscopy. Environmental and spatial data were extracted directly from each article when available. In cases where it was not possible to explicitly retrieve geographic coordinates of latitude and longitude and the sampling location was identified, QGIS (version 3.34.12) 30 was used to obtain altitude (meters above sea level) and environmental temperature using the SRTM 30s Digital Elevation Model 31 and WorldClim 2.1 32 (BIO1, 30-second arc resolution), respectively. The resulting dataset was exported as a CSV file and incorporated into subsequent meta-regression analysis in R. Meta-regression models were fitted using the metafor package with restricted maximum likelihood estimation (REML). A leave-one-out sensitivity analysis 28 was also applied for each subgroup meta-analysis to assess the influence of individual studies on prevalence results using the metafor package.
Publication bias was not evaluated because conventional methods, such as funnel plots, may generate false asymmetry in proportion studies, particularly when small samples, extreme proportions, or zero events are present 33 . Moreover, it has been pointed out that applying these tests—originally developed for clinical trials—to prevalence studies is inappropriate and may lead to misleading conclusions 34 . Therefore, given the limited use of these tools in this type of research, all articles were included without individual evaluation of publication bias. The results are presented in tables and figures, accompanied by a narrative synthesis, to provide an adequate response to the review question.
Results
The search strategy identified a total of 1915 studies. After removing 482 duplicates, 1433 studies remained. Following the application of inclusion criteria based on title and abstract, 865 studies were excluded, leaving 568 for full-text review. However, the full text of 27 of these could not be retrieved. After assessment, 351 studies met the inclusion criteria of this review and were subjected to methodological quality evaluation (Figure 1).
Methodological quality assessment
A total of 206 studies achieved acceptable methodological quality; of these, 91 met all six established criteria, 109 met five criteria, and six met four criteria (S1 Data).
Characteristics of the included studies
The included studies were published between 1982 and 2024, in Spanish, English, and Portuguese, and reported the identification of haemosporidians using PCR, microscopy, or both methods. In total, the studies analyzed encompassed 135 bird families. Altogether, the included studies documented 36,401 specimens infected with haemosporidians, representing the total number of positive individuals reported in the selected publications.
The investigations were based on samples collected in 65 countries across all inhabited continents. The countries with the highest number of studies were: United States (38 studies), Brazil (22 studies), Spain (11), Germany (9), Mexico (9), Colombia (7), Ecuador (8), Bulgaria (7), New Zealand (6), Chile (4), and China (5) (Figure 2). At the continental level, the distribution of study samples was as follows: America (89 studies), Europe (65), Asia (23), Africa (26), and Oceania (13). Global prevalence of haemosporidian parasites in wild birds across different zoogeographic regions.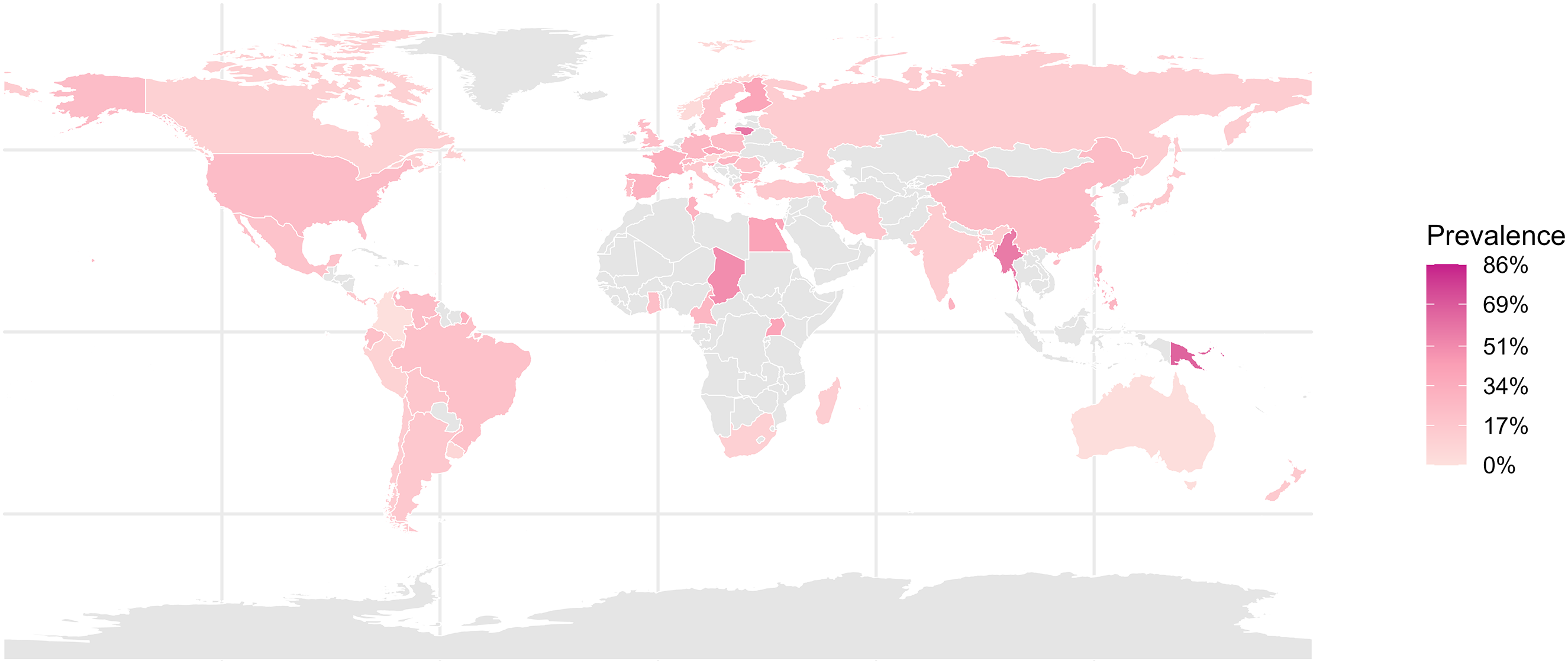
Pooled prevalence by diagnostic method
Overall
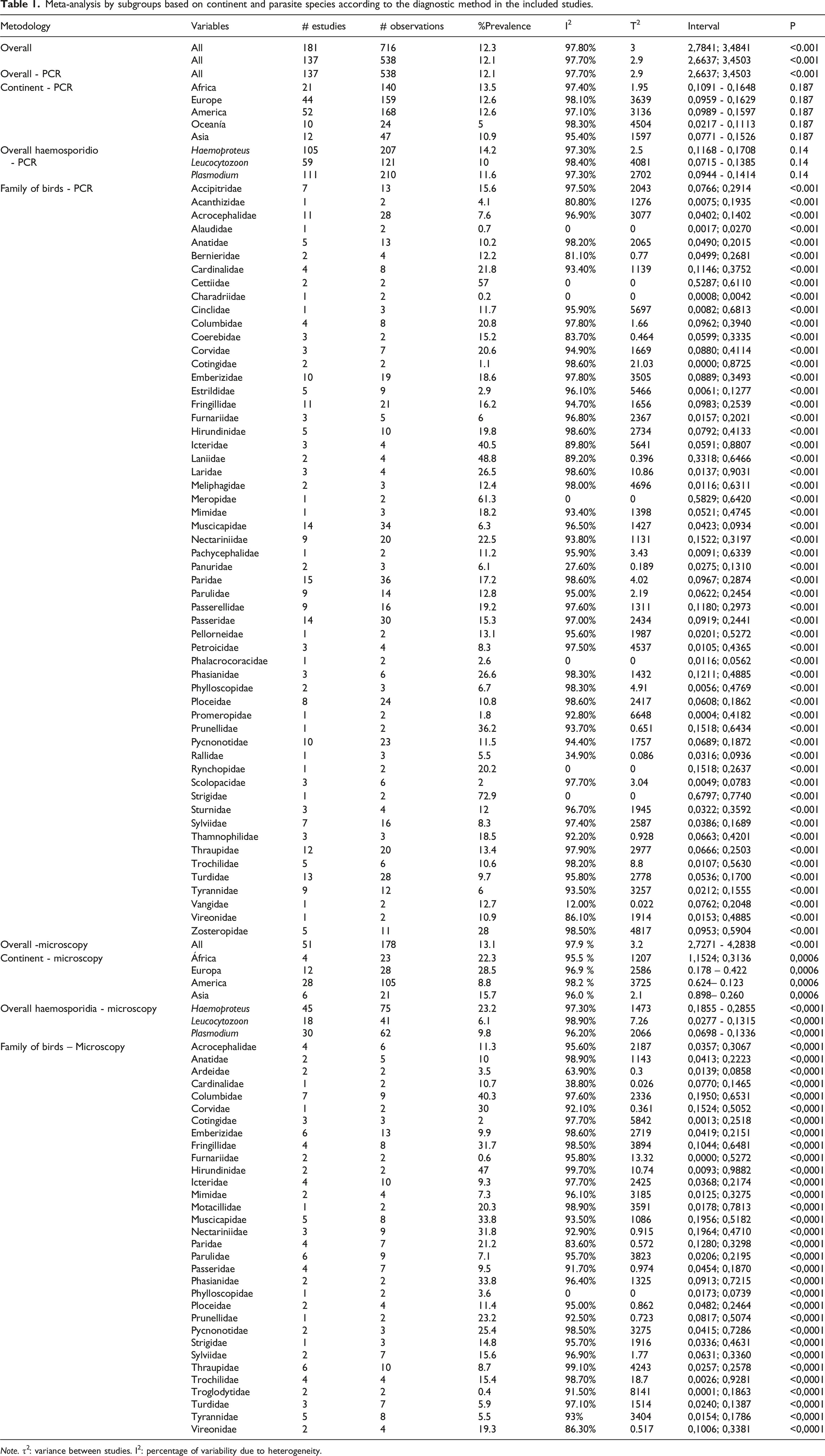
A total of 181 studies were included, showing high heterogeneity (I2 = 97.8%, τ2 = 3.00). Across these studies, 333,653 individuals were assessed, of which 32,290 were infected with haemosporidians, yielding a crude prevalence of P= 9.7%. The random-effects meta-analysis of proportions estimated the pooled prevalence at P= 12.3% (95% CI: 0.1096–0.1377).
PCR
Based on 137 studies using PCR, the estimated pooled prevalence was P= 12.1% (95% CI: 0.1063–0.1380). These studies also showed high heterogeneity (I2 = 97.7%, τ2 = 2.93).
PCR – pooled prevalence by continent
Meta-analysis by subgroups based on continent and parasite species according to the diagnostic method in the included studies.
Note. τ2: variance between studies. I2: percentage of variability due to heterogeneity.
PCR – pooled prevalence by parasite genus
The pooled prevalence estimates for the different parasite genera detected by PCR were: Haemoproteus: P= 14.2% (I2 = 97.3%, τ2 = 2.5006), Plasmodium: P= 11.6% (I2 = 97.3%, τ2 = 2.7022), and Leucocytozoon: P= 10.0% (I2 = 98.4%, τ2 = 4.0814). The test for subgroup differences showed no significant differences (Q = 3.93; df = 2; p = 0.1404) (Table 1).
PCR – pooled prevalence by bird family
The prevalence of haemosporidian infections varied widely among bird families. Some families showed notably high prevalence values, such as Monarchidae (monarch flycatchers; P = 81.0%), Strigidae (owls; P = 72.9%), and Meropidae (bee-eaters; P = 61.3%), while others, such as Charadriidae (plovers; P = 0.2%) and Spheniscidae (penguins; P = 0.6%), presented very low values. Heterogeneity was high in most cases (I2 > 90%). In contrast, families such as Alaudidae (larks and calandrias), Charadriidae, and Strigidae showed null or very low heterogeneity. Subgroup analysis revealed significant differences in infection prevalence among bird families (Q = 2135.58; df = 71; p < 0.001) (Table 1).
Microscopy
Fifty-one studies used microscopy, with an estimated pooled prevalence of P= 13.1% (95% CI: 0.1034–0.1615), showing high heterogeneity (I2 = 97.9%, τ2 = 3.24) (Table 1).
Microscopy – pooled prevalence by continent
The estimated prevalence by continent was P= 22.3% in Africa (I2 = 95.5%), P= 28.5% in Europe (I2 = 96.9%), P = 8.8% in the Americas (I2 = 98.2%), P= 15.7% in Asia (I2 = 96.0%), and P= 9.1% in Oceania (no heterogeneity estimated as only one study was available). These differences in point estimates were statistically significant (Q = 19.76; df = 4; p = 0.0006) (Table 1).
Microscopy – pooled prevalence by parasite genus
Regarding parasite genera detected through microscopy, the following prevalence estimates were observed: Haemoproteus P= 23.2% (I2 = 97.3%, τ2 = 1.473), Plasmodium P= 9.8% (I2 = 96.2%, τ2 = 2.066), and Leucocytozoon P= 6.2% (I2 = 98.9%, τ2 = 7.260). The test for subgroup differences was statistically significant (Q = 25.12; df = 2; p < 0.0001), indicating that prevalence estimates differed among parasite genera (Table 1).
Microscopy – pooled prevalence by bird family
The prevalence of haemosporidian infections varied widely among bird families evaluated by microscopy. Some families showed very high prevalence estimates, such as Laniidae (shrikes, P= 92.2%), Hirundinidae (swallows, P= 47.0%), and Passerellidae (new world sparrows, P= 47.5%), while others, such as Grallariidae (antpittas, P= 0.1%), Troglodytidae (wrens, P= 0.4%), and Furnariidae (ovenbirds, P= 0.6%), exhibited very low values. Heterogeneity across studies was high (I2 > 90%) in most families, such as Thraupidae (tanagers, 99.1%), Hirundinidae (99.7%), and Trochilidae (hummingbirds, 98.7%). However, some families showed low or null heterogeneity, such as Phylloscopidae (leaf warblers, I2 = 0%). Subgroup analysis revealed statistically significant differences in infection prevalence among bird families (Q = 410.28; df = 41; p < 0.0001).
Meta-regression
Of the 181 studies included in the meta-analysis, 84 contained sufficient information to be incorporated into the environmental meta-regression analysis. Of these, 64 studies reported explicit geographic coordinates, while 20 reported the sampling site as georeferenced to a clearly identifiable locality. Regarding environmental variables, 11 studies provided temperature data and 15 reported altitude or altitudinal ranges.
In the meta-regression analyses performed, no significant associations were observed between year of publication and estimated prevalence, for both PCR (β = −0.0049; p = 0.1467) and microscopy (β = −0.0016; p = 0.5963) (Figure 3(a) and (b), respectively). No significant associations were found between prevalence and environmental variables: altitude (β = 0.0003; p = 0.9584), latitude (β = −0.0772; p = 0.5387), and temperature (β = 0.0055; p = 0.6637) (Figures 4(a) and (b), and 5). Meta-regression of prevalence in relation to the year of publication of the studies, according to diagnostic method: (a) PCR and (b) microscopy. Meta-regression of prevalence in relation to geographic variables: (a) latitude and (b) altitude (m a.s.l.). Meta-regression of prevalence in relation to mean annual temperature.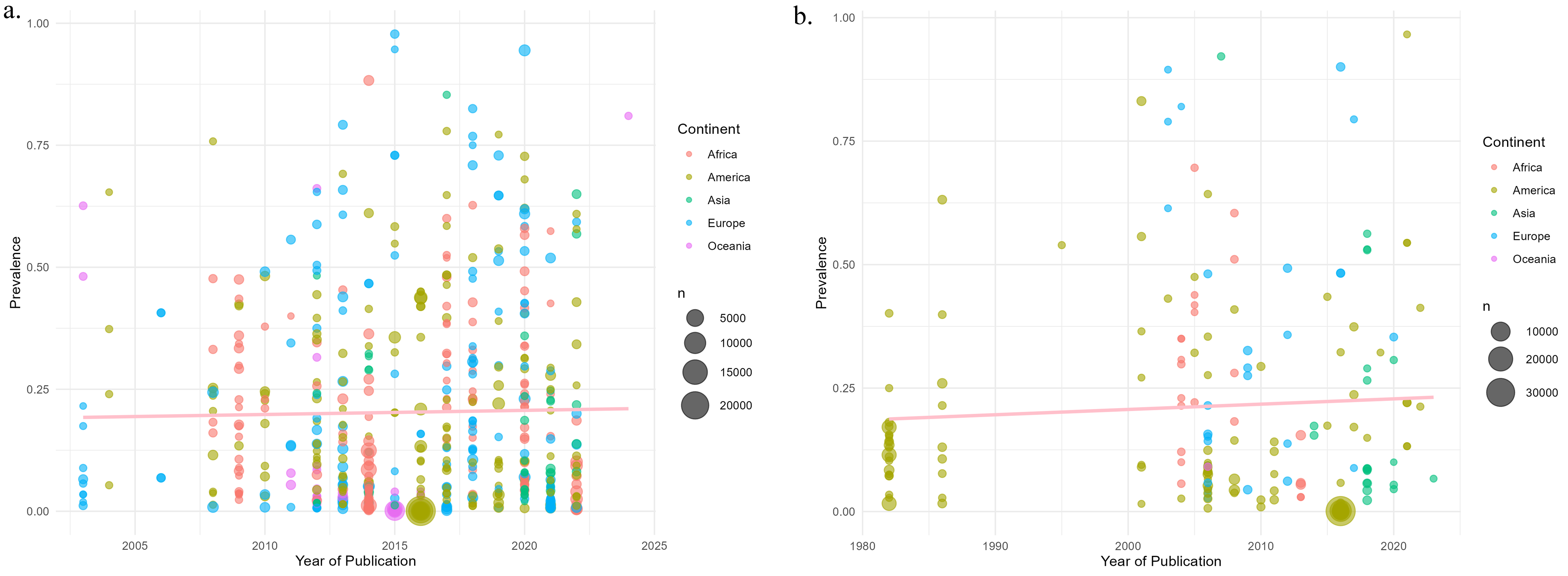
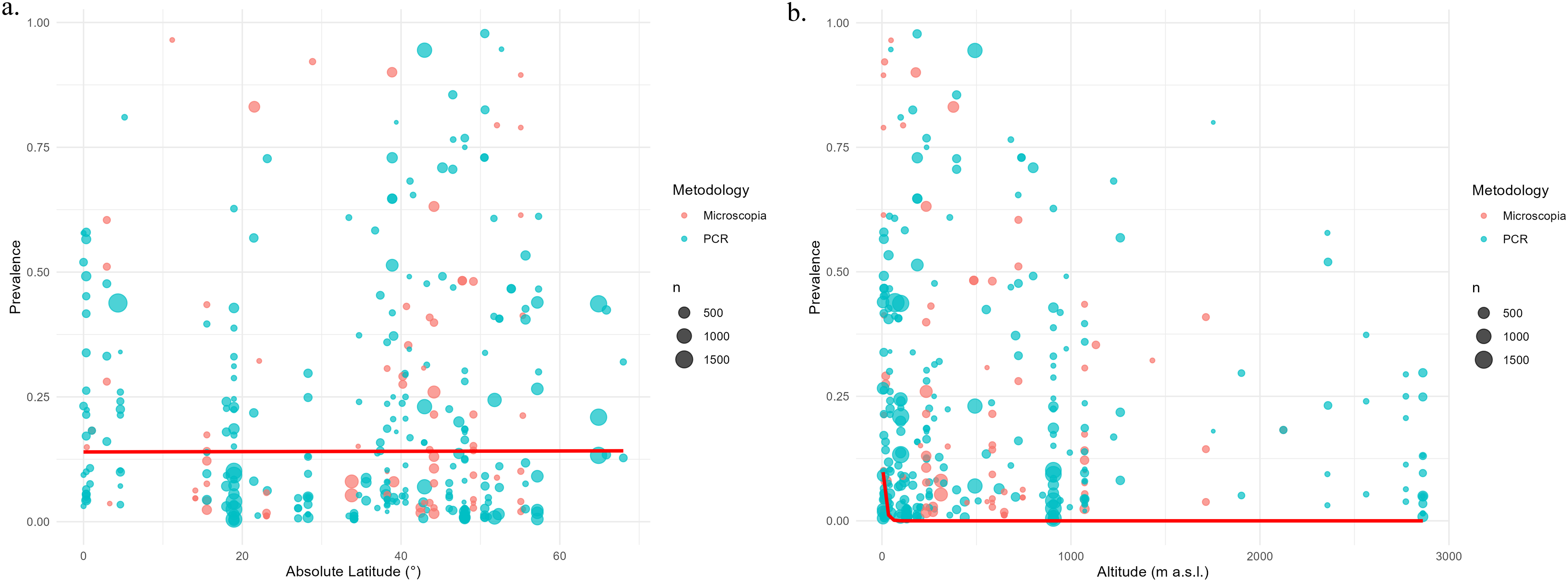
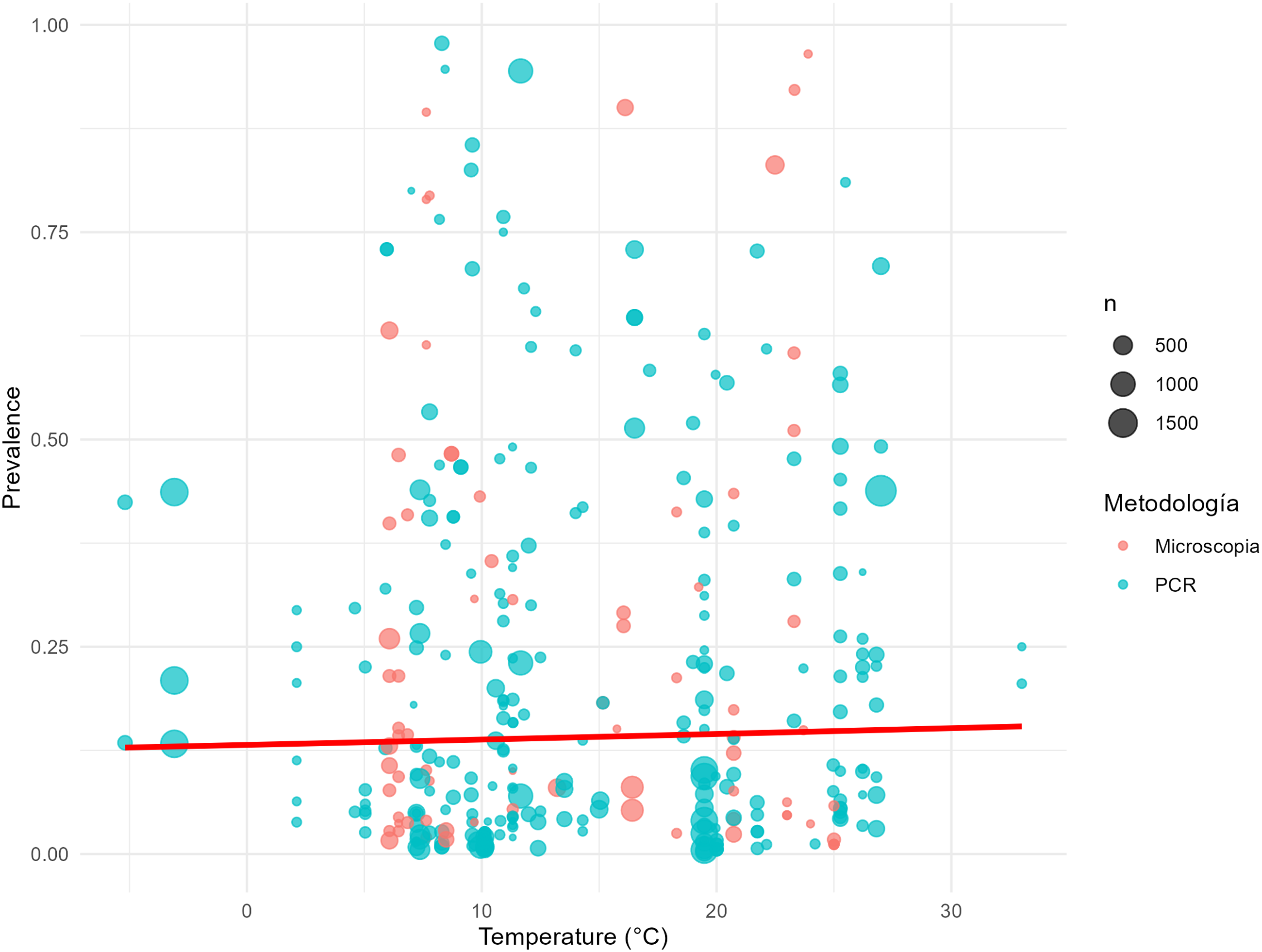
Sensitivity analysis
Upon removing the influential studies (61) identified in the sensitivity analysis, no significant changes were observed in the overall prevalence or in the prevalence obtained from the subgroup analysis. Additionally, no changes were observed in the heterogeneity of the studies included in these meta-analysis.
Discussion
This is the first systematic review and meta-analysis of the global prevalence of haemosporidian parasites in wild birds based on prevalence estimates obtained using microscopy and PCR. The systematic review included 206 studies that achieved acceptable methodological quality, from which the prevalence was synthesized. The included studies were published between 1982 and 2024, in Spanish, English, and Portuguese, and comprised data from 135 bird families. The meta-analysis showed that the global prevalence of infection was P=12% by PCR and P=13% by microscopy.
The results of the methodological quality assessment, based on an adaptation of the Joanna Briggs Institute criteria 22 , revealed frequent weaknesses in the included studies. One of these was the limited description of the sampling environment. Although most studies reported geographic coordinates or the general name of the sampling site, few provided detailed information about the habitat, ecological conditions (forest type, temperature, humidity, access to water sources, topography, among others), or characteristics of the bird community sampled, which limits the interpretation of environmental factors associated with prevalence. Statistical procedures were also often not clearly detailed; many studies reported prevalence values without explaining the formula used or providing the number of individuals sampled and positive, which limited traceability and reproducibility when manually calculating prevalence.
The concentration of studies with samples collected and developed in countries such as the United States and Brazil likely reflects not only institutional and logistical capacity to conduct research with wild birds but also access to natural populations and infrastructure for blood sample processing. In South America, Brazil stands out for its research tradition and technical capabilities, which have promoted scientific output in this field 35 . In addition to Brazil, Ecuador and Colombia are biodiversity hotspots, favoring the study of a wide range of host–parasite interactions6,36. However, although Colombia has the highest bird diversity in the world 37 and its ecosystems are suitable for investigating parasite ecological dynamics38,39, this review identified only eight studies with samples from this country, seven of which were processed locally. This suggests that research development depends not only on biological richness but also on factors such as research priorities, available scientific infrastructure, and the geographic distribution of birds.
The high heterogeneity observed in the meta-analysis indicates significant variability among the included studies. This heterogeneity can be attributed to geographic, taxonomic, and methodological factors. Ecologically and geographically, the studies encompassed a wide range of regions with distinct environmental conditions that influence the distribution and transmission of haemosporidians 40 . The taxonomic diversity of host birds also plays a role, as species differ in behavior, physiology, and evolutionary history, which can impact their susceptibility to infection1,41. Regarding methods, studies employed different diagnostic approaches (PCR, microscopy, or both), with variations in protocols, sample type and quality, and technical expertise, which significantly affected detection capacity and specificity 42 . For example43, demonstrated that poorly prepared or stained blood smears can lead to significant underestimations in microscopy, while PCR detection capacity may vary depending on sample volume, type, and extraction protocol 44 . Furthermore, prevalence is influenced by the time of year samples are collected, as parasitemia may fluctuate according to the birds’ reproductive cycle 1 .
In this review, Haemoproteus was the most prevalent genus in studies using both microscopy and PCR, consistent with previous research that has demonstrated its higher detectability compared to other genera. One possible explanation is that Haemoproteus often reaches higher parasitemias than Plasmodium, facilitating detection by both diagnostic methods even in chronic or low-intensity infections9,40,45,46. For example, Musa et al. 40 consistently observed high detection rates of Haemoproteus using microscopy, multiplex PCR, and nested PCR, thereby reinforcing the idea that this genus is easily detectable regardless of the method employed. Additionally, Haemoproteus exhibits notable host specificity3,41, with a higher prevalence in species characterized by particular ecological traits, such as insectivorous habits, forested habitats, and restricted distributions 41 . These factors suggest that both parasite and host characteristics influence higher prevalence values, which may also explain the high heterogeneity observed in family-level analysis.
Examining bird families with the highest estimated prevalence of haemosporidian parasites, no consistent patterns were observed. In PCR analysis, the families with the highest estimated prevalence included Nectariniidae (sunbirds and spider hunters, P= 22.5%, 10 studies), Hirundinidae (P= 19.8%, 10), Emberizidae (sparrows and buntings, P= 18.6%, 19), Passerellidae (old world sparrows, P= 19.2%, 16), and Paridae (chickadees, P= 17.2%, 36). For microscopy, the highest prevalence values were observed in Columbidae (pigeons, P= 40.3%, 9 studies) and Nectariniidae (P= 31.8%, 9 studies). Methodological studies suggest that for robust meta-analysis estimates, at least five studies are required, preferably more than nine, since including only the nine largest studies can reproduce up to 80% of the results of a complete meta-analysis47,48. Therefore, these families with ≥9 studies can be considered sufficiently represented to interpret prevalence estimates as robust. However, although these criteria improve statistical robustness, they may present ecological limitations, especially in highly diverse Neotropical communities, where many species are represented by small sample sizes 49 . The underrepresentation of species could obscure community-level processes, such as dilution or the dynamics of prevalence amplification50–53.
Prevalence values varied across continents depending on the diagnostic method employed. However, considering each method separately, Africa (PMIC = 22.3%; PPCR = 13.5%) and Europe (PMIC = 28.6%; PPCR = 12.6%) showed the highest estimates compared to other continents (Table 1). This result may be influenced by factors such as a long tradition in avian parasitology studies, which has promoted the use of standardized protocols, highly trained personnel, and high-quality blood smears, conditions that improve microscopy detection 43 . It is essential to note that not all studies employed both diagnostic methods; some used only microscopy, while others utilized both techniques. This methodological variability may affect the direct comparability between regions and lead to an underestimate or overestimate of the true prevalence when data are pooled.
The prevalence of haemosporidian parasite infection by genus estimated using PCR in this review was PPla=11.6% for Plasmodium, PHae= 14.2% for Haemoproteus, and PLeu= 10.0% for Leucocytozoon. These values differ from those reported by Swangneat et al. 17 , who reported PPla = 21%, PHae= 18%, and PLeu= 34%, respectively, in wild and domestic birds from Southeast Asia. The higher prevalences observed by Swangneat et al. 17 could be attributed to: (1) inclusion of only 15 studies, whereas this review analyzed 136 PCR-based studies; (2) focus on a single geographic region that may favor haemosporidian transmission due to ecological characteristics; and (3) the search for parasites was extended to domestic birds. This latter aspect is especially relevant for Leucocytozoon, as much of the reported prevalence is influenced by the study of Khumpim et al. 54 , documenting 81% prevalence in domestic birds. In comparison, Yan et al. 17 reported PPla= 12.8% in wild birds, slightly higher than the estimate obtained in this study (PPla= 11.58%). Differences may arise from inclusion criteria, as Yan et al. 17 considered only studies with sample sizes >400 individuals, which could affect general prevalence estimates.
Comparisons with captive wild birds show that the prevalences estimated here are similar or lower. Research in zoos and rehabilitation centers reports variable prevalence values (7%<P<30%), depending on parasite genus, diagnostic method, and host species. For example, in Brazilian zoo birds, P= 12.6% predominated by Plasmodium 55 ; in penguins from Japanese facilities, PPla= 7.5% and PHae= 1.7% 57; in rehabilitated raptors in Iran, PHae= 26.7% and PLeu= 10% 56 ; and in rescued birds in Portugal, overall prevalence reached P= 30.3% using PCR 57 . These data suggest that while captivity may facilitate parasite transmission due to host density and open facilities55,58, infection prevalence values are comparable to or exceed those observed in free-living wild bird populations.
This review has some limitations. First, only studies published in English, Spanish, and Portuguese were included, potentially excluding relevant research in other languages and limiting the comprehensiveness of the review. Environmental variables, such as temperature and altitude, were inconsistently reported across studies. When they were unavailable, geographic coordinates were used to extract altitude (SRTM) and mean annual temperature (WorldClim 2.1). While this approach allowed for data standardization, it may have introduced bias, as WorldClim provides long-term average climatic values rather than the actual temperature at the time of sampling. Consequently, this limitation could have influenced the observed relationships between altitude, temperature, and prevalence.
Finally, the MalAvi database was not considered as a source of information, even though 52.85% of the total references indexed in it were retrieved from the databases included in the review. The references not retrieved use laboratory tests or assays for parasite identification; others focus on studying vectors and transmission, as well as other relevant aspects different from those considered in the search strategy of this review.
Based on this review, we recommend that future studies on haemosporidian prevalence in birds include the precise geographic coordinates of sampling sites, the month or season of sampling, environmental variables such as temperature and altitude, a description of the habitat or ecosystem type, the total number of individuals sampled, the number of infected individuals, as well as overall prevalence and, when possible, prevalence by parasite species. This will facilitate reproducibility and comparability among primary studies, as well as the combining and synthesizing of their findings in future reviews of this nature.
Finally, the results of this systematic prevalence review provide a global estimate of population-level infection by haemosporidian parasites in wild birds, representing a valuable contribution to understanding the distribution and frequency of these parasites worldwide. Our findings identified bird taxonomic groups and continents with particularly high prevalence estimates, which can guide future research and conservation actions. Furthermore, the results highlight the importance of considering host, vector, and environmental characteristics in parasitic ecology studies, while also indicating gaps in some geographic regions, such as Paraguay, French Guiana, and post-Soviet states. This synthesis helps delineate monitoring priorities in studying host–parasite dynamics in wild birds.
Conclusion
This review provides a global overview of the prevalence distribution of haemosporidian parasites of the genera Plasmodium, Haemoproteus, and Leucocytozoon in wild birds, based on data obtained from blood samples. Plasmodium was the most prevalent genus, followed by Haemoproteus and Leucocytozoon. It is possible to estimate this parameter at different scales, including continent, country, bird family, and parasite genus. This approach enables the identification of regions, bird families, and parasite species with the highest prevalence, providing valuable information to prioritize future studies that guide avifaunal conservation efforts and epidemiological monitoring. Furthermore, publication year was not associated with prevalence estimates according to diagnostic method, and geographic (latitude and altitude) and environmental variables (temperature) were also not significantly associated with prevalence. No single variable was identified that could explain the observed variation. Instead, high heterogeneity among studies was observed, suggesting that multiple biological, environmental, and methodological factors contribute to the differences and should be considered in future analysis and comparisons.
Supplemental material
Supplemental material - Global prevalence of haemosporidian parasites in wild birds: Systematic review and meta-analysis
Supplemental material for Global prevalence of haemosporidian parasites in wild birds: Systematic review and meta-analysis by Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González in Avian Biology Research
Supplemental material
Supplemental material - Global prevalence of haemosporidian parasites in wild birds: Systematic review and meta-analysis
Supplemental material for Global prevalence of haemosporidian parasites in wild birds: Systematic review and meta-analysis by Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González in Avian Biology Research
Supplemental material
Supplemental material - Global prevalence of haemosporidian parasites in wild birds: Systematic review and meta-analysis
Supplemental material for Global prevalence of haemosporidian parasites in wild birds: Systematic review and meta-analysis by Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González in Avian Biology Research
Footnotes
Author contributions
Conceptualization: Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González.; methodology: Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González.; validation: Leidy Johanna Rueda Díaz; formal analysis: Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González.; investigation: Tiana Glemarjenni Beltran Toledo and Leidy Johanna Rueda Díaz.; resources: Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González.; data curation: Tiana Glemarjenni Beltran Toledo and Leidy Johanna Rueda.; writing—original draft preparation: Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz.; writing—review and editin: Tiana Glemarjenni Beltran Toledo, Leidy Johanna Rueda Díaz and Fernando Rondón González.; visualization: Tiana Glemarjenni Beltran Toledo and Leidy Johanna Rueda Díaz.; supervision: Fernando Rondón González.; project administration: Leidy Johanna Rueda Díaz.; funding acquisition: Fernando Rondón González. All authors have read and agreed to the published version of the manuscript. Please turn to the ![]() for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded by Vicerrectoría de Investigación y Extensión, Universidad Industrial de Santander; VIE-UIS 3707.
Declaration of conflicting interests
The authors declare no conflicts of interest.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.