
Abstract
Both wild birds and commercial poultry species have the typical sex ratio at birth, which means that about half of the males are unable to produce eggs. Relationship between embryo sex and egg geometrical and physical properties is one of the most reexamined assumptions. The aim of this study was to identify the relationship between the geometric and physical parameters of Japanese quail (Coturnix japonica) eggs and the sex of the embryos contained within them. The various parameters tested included egg shape, asymmetry and conicity indices, volume, surface area, weight, densities of the whole egg and its contents, and several parameter combinations based on these variables. Of these, only heat transfer index showed significant differences between the groups of male and female eggs on Day 5 during incubation. It was hypothesized that the value of this coefficient is related to the stage of embryo development and the intensification of metabolic rate. This may differ significantly between male and female embryos. Differences in heat transfer values provided accurate information about the potential culling of approximately 18% of the total number of eggs containing male embryos. Suggestions were made for improving the conditions for similar studies aimed at increasing the efficiency of egg sexing.
Introduction
Birds, including poultry species used for commercial egg production, tend to be born with the usual sex ratio, meaning that around 50% (males) cannot be used to lay eggs of their own.1–4 Raising these males for meat production is ineffective, as egg-laying breeds have not been selected for optimal feed conversion for meat production.5–10 This problem is particularly acute in chicken egg production11–14 as chicken eggs constitute the vast majority consumed worldwide. It is similarly relevant, however, in duck egg farming,15,16 as well as in the production of edible Japanese quail (Coturnix japonica) eggs.17–19
Historically, interest in the issues of sex ratio and its potential shift in poultry has existed for a long time.20–28 To resolve these, one of the most reexamined hypotheses is the connection between the geometrical and physical characteristics of the egg and the sex of the embryo.29–32 Modern technological innovations in the field of spectral-based techniques have brought the possibility of egg sexing during incubation closer (for examples, see the most recent reviews33–35). Critical refinements will make the predictive capability more accurate, and the incubation stages at which such identification is possible is a key criterion. A limiting factor, however, is the cost and complexity of industrial interpretation of these technological solutions. While we can assume that large hatcheries examining thousands of eggs daily will be able to afford such costs, smaller hatcheries, often farm-based, will require more cost-effective solutions, including straightforward techniques for determining oomorphological indicators.36–39
Geometrical and physical parameters vs egg sexing
One of the most common assumptions, periodically emerging and re-examined by various scientific teams, is the relationship between embryo sex and egg shape.34,40 The concept of shape most often refers to the shape index, i.e., the ratio of the maximum breadth of the egg to its length (B/L) proposed over 100 years ago. 41 Published results on chicken eggs can be contradictory.42,43 Each new publication demonstrating a significant correlation between B/L and embryo sex, 44 however, encourages further authors to continue their work in this direction.
The question of the relationship between embryo sex and egg geometry has been applied to Japanese quail eggs and, again, results are not clear-cut. For example, Idahor et al., 17 having found some female prevalence over males in round and oval eggs, nevertheless concluded that “it may probably not be achievable to predetermine quail chick sex at the pre-hatching stage using egg indices.” In later studies, these authors 18 found that, in addition to round and oval eggs, spherical and elliptical eggs also had different male-to-female ratios. They were, however, unable to develop an effective mechanism for identifying embryo sex in any one particular egg. As a result, the conclusion of these studies 18 remained unchanged from the previous one: “Japanese quail egg shape may not influence chick sex”. Before this, Dioses et al. 19 had examined a range of quail egg parameters that could provide a comprehensive solution to their sex classification. They concluded that “morphological features of the Japanese quail egg, such as eccentricity and shape index, can be used as significant factors in classifying its sexes”. In addition to geometric parameters, the authors 19 considered shell color and the main physical parameter of the egg, its weight (W).
The use of physical parameters formed the basis for similar studies of chicken eggs from Rhode Island and White Leghorns. 45 This study attempted to assess differences in chicken eggs by the sex of their embryos using the density of the contents (D i ) and the whole egg (D). D i was calculated using an empirical relationship that included egg weight (W), volume (V), and length (L). Despite the positive results, similar studies have not been continued.
Prospects for using the thermal conductivity of an egg
One promising avenue for exploring possible differences between male and female embryos may be the egg’s thermoregulation,46–48 specifically its thermal conductivity index. Thermal conductivity measures how quickly an object absorbs or releases heat when heated or cooled.49–51 We considered this temperature parameter to be promising because: (1) it is relatively easy to assess, especially during the incubation process, by removing the eggs from the incubator and measuring their temperature changes during natural cooling; (2) our studies on goose eggs 52 demonstrated significant differences in the cooling rates of fertilized eggs, specifically differences between those with dead and viable embryos. Furthermore, the cooling rate was also predictive of some egg morphological parameters. Despite the popularity of the thermal conductivity parameter of eggs, research has been limited by the requirements of the food industry, such as predicting the cooling period after cooking, 53 and incubation theory.54,55
The real prerequisites for the use of studies of thermal conductivity for egg sexing have existed for some time. For example, Tagirov and Rutkowska 56 demonstrated that after just 36 hours of incubation, male zebra finch embryos grow faster than females. The authors attributed this to the fact that males have higher expression of the growth hormone receptor (GHR) gene, as well as certain genes responsible for cell cycle regulation and metabolism. Given that embryonic activity is associated with heat production,57–59 it is possible that assessing the heat production conditions of eggs will allow us to determine the sex of the embryo already in the early stages of incubation. Differences in metabolic rate between female and male embryos were also found in a study of the eggs of the Eurasian kestrel, Falco tinnunculus. 60 In this species, female embryos have a shorter embryonic period than males, allowing females to hatch earlier and occupy a higher rank than males in the size hierarchy within the brood.
Since heart rate can also indirectly indicate embryonic metabolic rate, Glahn et al. 61 demonstrated different heart rate values in White Leghorn chicken embryos. Importantly, the authors found that sex-related differences in heart rate differed between female and male embryos. Thus, based on the described results, it can be assumed that the metabolism of male and female embryos may differ during incubation. 62 Under certain environmental conditions, such as cooling, there is evidence that embryos of a particular sex respond differently to such thermal stress.
Some notes from classic theory
The classical theory of heat transfer helped determine a physical indicator that quantitatively reflects the magnitude of thermal conductivity.
As follows from Newton’s law of cooling,
63

Winterton
63
provided a formula for determining the constant β, according to which it depends on the heat transfer coefficient, area for heat flow, volume of an object, its density, and specific heat. Considering that several parameters that affect the heat transfer coefficient and specific heat, such as air flow, measurement characteristics, and object location, would remain constant in our case, we assumed the use of an integral index h that combines the heat transfer coefficient and specific heat values. To demonstrate the physical meaning of this index and its similarity to the heat transfer coefficient, we decided to call it the heat transfer index. Then, according to the theoretical findings of Winterton
63
and our assumptions, the constant β in the case where the cooling object is the egg can be expressed, after some of the corresponding mathematical transformations, as follows:

Considering equations (1) and (2), the heat transfer index (h) can be expressed as follows:
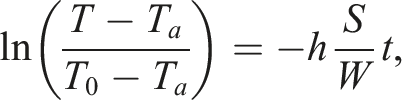
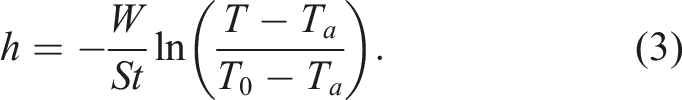
If we consider only the inner part of the egg (W
i
), which weight is the difference between the weights of the entire egg (W) and its shell (W
s
), i.e., W
i
= W – W
s
, then
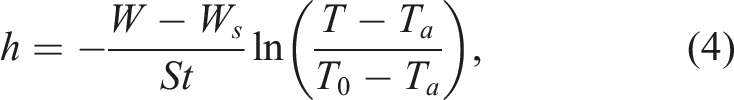
Study aim formulation and choice of influencing parameters
The results of shifting the sex ratio in eggs with different geometric and/or physical parameters are intriguing, but their effective practical application is only possible when we master the technique of specifically influencing the hen’s body to produce eggs of a specific design deliberately. However, even in this case, it is highly doubtful that a mother bird laying, say, only oval eggs, would maintain a constant male-to-female ratio of 15:23, similar to that obtained, for example, by Idahor et al. 17 Rejecting all oval eggs from the incubation process, hoping to eliminate a certain, even predominant, number of “unnecessary” males, while simultaneously sacrificing a certain number of “valuable” females, is clearly economically unviable. Separating eggs by sex can be economically feasible if one or a set of parameters can be identified that demonstrate a clear and consistent boundary between the distribution of eggs with male and female embryos. Therefore, the more initial parameters involved in an experiment, the higher the likelihood of identifying such a boundary. Consequently, we decided to focus on what we believe to be the most promising parameters, the study of which could allow us to determine a further trend in the in-depth study and testing of scientific approaches to solving this problem.
Considering the availability of a number of publications on the possibility of using certain geometric and/or physical features to assess the potential difference between “male” and “female” eggs, it was decided to analyze these parameters in addition to the h value, in which we included the most promising, from our point of view,
64
indices: - The shape index, reflecting the ratio of the maximum breadth to the length of the egg (B/L); - An asymmetry index, reflecting the ratio of the displacement w of the B-axis from the egg’s center to its length (w/L); - A conicity index, reflecting the ratio of the egg’s diameter D
p
at a point shifted by L/4 from the sharp end to its maximum breadth (D
p
/B); - An integral geometric index that we called egg geometrical index (EggGI),
65
which includes all three of the above indices: - Metabolic index, reflecting the ratio of the surface area of the egg S to its volume V; - Egg density, D, which is the ratio of its weight W to its volume V; - Egg content density, D
i
, determined by the ratio of the weight of egg content W
i
to its volume V
i
.
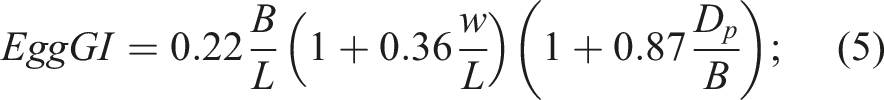
Since the D i value is predictive, its calculation, as well as the calculation of W s , became possible due to previously conducted theoretical66,67 and experimental studies 68 to determine the relationship between the morphometric parameters of eggs.
As a result, the aim of our research was to find possible differences in the thermal conductivity of Japanese quail eggs with male (M) and female (F) embryos at different stages of incubation, the value of which, expressed by the heat transfer index (h), taken separately or in combination with other geometric and/or physical parameters, will demonstrate a clear boundary between M and F.
Material and methods
Ethics statement
The study was conducted according to the guidelines of the Declaration of Helsinki and the L. K. Ernst Federal Research Center for Animal Husbandry (LKEFRCAH) ethical guidelines. The respective Protocol No. 4 was approved by the LKEFRCAH Commission on the BioEthics of Animal Experiments on 13 June 2024.
Experimental birds
We used Japanese quails of the F2 resource population previously developed as described elsewhere.69–71 The age of quails was 4–5 months. The male to female ratio was set at 1:5. The conditions for growing, keeping and feeding the experimental flock of quails met generally accepted requirements.72–74 In particular, the quail housing facilities were maintained at a temperature of 20–25°C and humidity of 55–65%. The birds had free access to feed and clean water. Commercially prepared feeds were used for feeding.
Traits measured and calculated
A total of 295 eggs were selected for analysis. The eggs were photographed in a horizontal position and weighed (W) within an accuracy of 0.01 g. Using a caliper, the length (L) and maximum width (B) of the eggs were determined within an accuracy of 0.1 mm. Other geometric dimensions were estimated from the egg images using Microsoft Picture Manager software. The L and B values, as well as the distance (w) of the displacement of the B axis from the egg center 75 and the egg diameter (D p ) at a point located at a distance of L/4 from the pointed end,76,77 were measured within an accuracy of 1 pixel. Knowing the L size in mm and pixels, respectively, we converted the geometric parameters w and D p into the metric system.
The volume (V) of eggs and the surface area (S) were calculated using previously derived universal formulae,
78
the adequacy of which was assessed, among other avian eggs, on quail eggs, as follows:
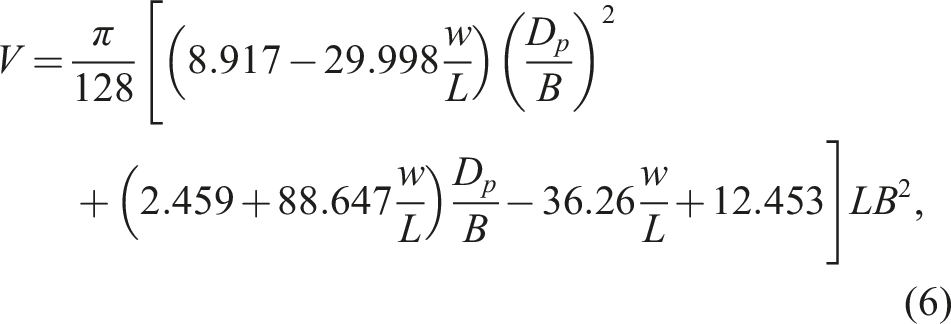
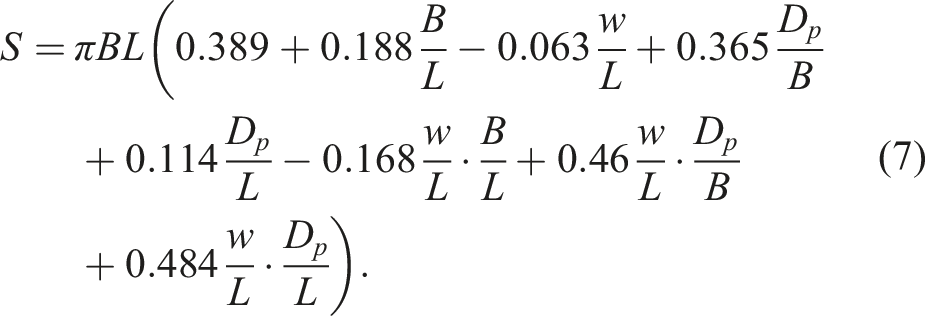
The density (D) of eggs was calculated as the ratio of W to V.
The weight of the shell (W
s
) was calculated using previously derived formula from Narushin et al.:
68
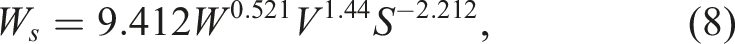
Using the experimental data from our previous studies on the quail eggs of F2 resource population,
68
an empirical formula for calculating the density of egg interior (D
i
) was deduced. The formula similar to equation (9) is based upon the values of a set of egg basic parameters: W, V and S. The obtained correlation between the measured and calculated values of D
i
was 0.938.
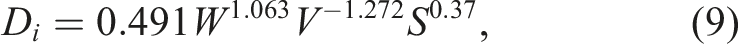
After all measurements were taken, the eggs were placed in an Rcom Maru 190 Deluxe MAX incubator (Autoelex Co., Gyeongsangnam-do, Korea) incubator with individual cells for each egg and incubated for 17 days. Initially, the eggs were set in the incubator for a period of 2 hours. After that, they were removed and a shell surface temperature (T) of each egg was measured with an infrared thermometer at an upper middle point for four times: immediately after removing the eggs from the incubator, and then three times with 0.5–hour interval each. An ambient temperature (T a ) was also measured on a daily basis. Such a procedure was repeatedly undertaken over each of 17 incubation days. Each egg was weighed at the same frequency being indicated as W1, W2, …, W17 respectively).
The values of the heat transfer indices (h) were calculated according to equation (5).
After hatching, the sex of the day-old quails was determined by cloacal examination as described elsewhere.79–84
Statistical data processing
A number of mathematical and statistical procedures found in the STATISTICA 5.5 program (StatSoft, Inc./TIBCO, Palo Alto, CA, USA) and Microsoft Excel applications were utilized to process the data. Accordingly, the Pearson correlation coefficient (R) value was used to evaluate the validity of the associations that were found, and their significance was confirmed with t-test at the p < 0.05 level.
To visualize the obtained results, which have score values, a graphical representation technique was used on a coordinate grid with equal values of datasets of the estimating parameters along the abscissa and ordinate axes.
Results and discussion
Raw data production and processing
Of the 295 eggs used in the experiment and suitable for further analysis (i.e., excluding unfertilized, damaged, and those containing embryos that died at various stages), 110 eggs were successfully hatched, 67 of which contained males, conventionally designated as M, and 43 females, designated as F, respectively. For further statistical analysis and mathematical interpretation of the relationships between Japanese quail sex and the measured/calculated egg parameters, hatched males were assigned number 1, and females 2. Thus, a mathematical function containing 67 ones and 43 twos was obtained. Despite the scoring of this function, the use of the Pearson correlation coefficient was considered preferable than the Spearman correlation for a number of reasons: the amount of data obtained is large enough; independent indicators have very wide ranges of variation, and therefore there is no reason to assume any nonlinear relationship between dependent and independent parameters; and the approximation of the scoring function will make it possible to evaluate the obtained values not as an unambiguous hit in the numerical series “1” or “2”, but as a more or less reliable probability of making a decision whether to attribute this result to the presence of a male (1) or a female (2) in the egg.
Embryo sex and egg geometric parameters
A comparative assessment of M and F eggs based on their geometric conformity revealed no significant differences. For example, the correlation between the shape index (B/L) and embryo sex was 0.130. For the asymmetry index (w/L), R = −0.087, and the conicity index (D p /B) demonstrated this relationship at a level of 0.021. Using the integral geometric index EggGI (equation (6)) also failed to increase the probability of a correct prediction, demonstrating R = 0.114. As expected, neither the differences between the M and F groups based on these geometric characteristics nor the resulting correlation coefficients were significant.
To visualize this fact, we selected images of geometrically similar eggs containing embryos of opposite sexes (Figure 1). Images of eggs containing male (A) and (B) and female (C) and (D) embryos.
Considering that the highest correlation coefficients with the sex of the embryo were demonstrated by B/L (0.130) and EggGI (0.114), we visualized these relationships using the graphical functions presented in Figure 2. Visualization of the results of the shape index, B/L (A) and EggGI (B) values from a group of eggs with male (M) and female (f) embryos.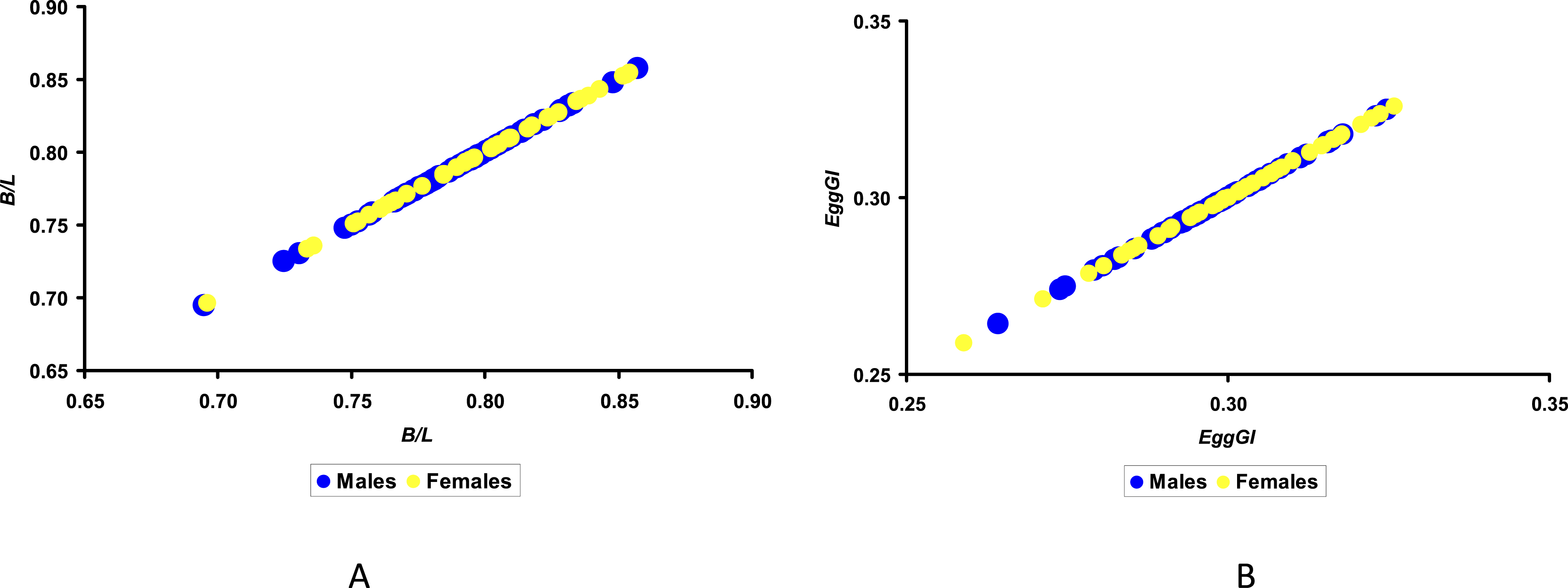
The only thing that can be concluded from analyzing the resultant graphical dependences is that the lowest EggGI was demonstrated by the egg containing a female embryo (Figure 2(B)). We have shown an image of this egg in Figure 1(D). However, to say unequivocally that breeding quail eggs with such ratios of the geometric parameters (EggGI = 0.259) can guarantee the hatching of only females would be completely illogical, given that a similar egg (EggGI = 0.264) corresponds to a male (Figure 1(B)).
Following the logic found in other similar studies17,18 and arbitrarily dividing the resulting diversity of forms into certain broad categories, we can conclude that males prevail in the elongated eggs (with lower B/L values), while females occur more often in the round eggs (with higher B/L values). To confirm or refute this fact statistically, we decided to use the chi-squared (χ2) test.85,86 For this purpose, we assigned the group of eggs with B/L < 0.78 an index of 1, that with B/L = 0.78–0.8 an index of 2, and that with B/L > 0.8 an index of 3. The M:F sex ratio for each B/L group is presented in Figure 3. The number of eggs with male and female embryos, divided into groups according to the shape index (B/L): 1. B/L < 0.78; 2. B/L = 0.78-0.8; 3. B/L > 0.8.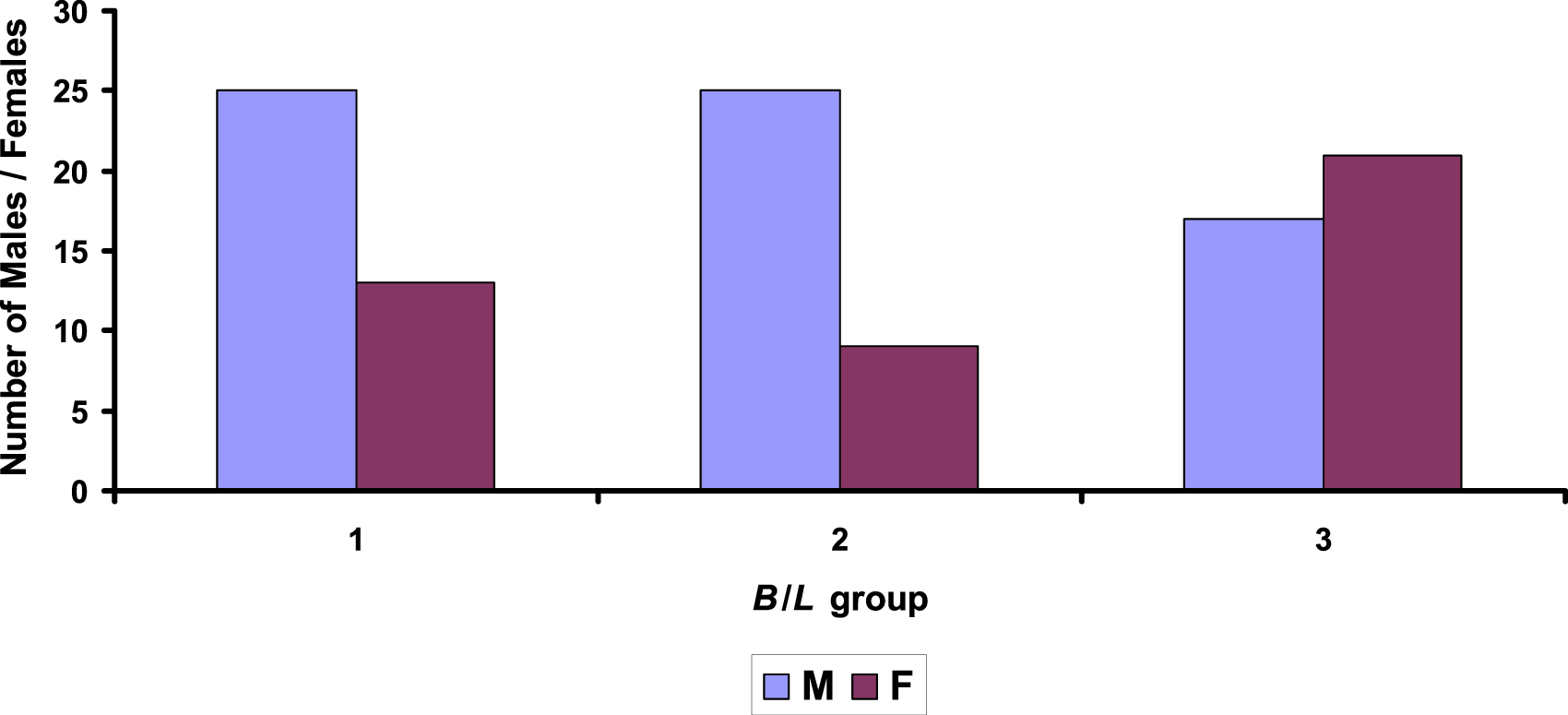
The χ2 test confirmed the statistical significance of differences in the M:F ratios across egg groups with different shape index values. The calculated χ2 value was 6.83, while a table value was 5.99 (for 2 degrees of freedom and p < 0.05). Despite the statistically confirmed differences, such observations do not provide a practical solution to the problem of egg sexing. It would be completely pointless to incubate only round eggs that would still produce about 45% males. This would also result in a loss of about 30% of the females that would hatch from the more elongated eggs of other B/L groups. Nevertheless, the difference in sex ratios based on the distribution of eggs among B/L groups was statistically significant.
Derived egg parameters (V and S), calculated under geometric measurements (equations (7) and (8)), also failed to provide any hope of using them as reliable markers of egg sex, either individually or as the S/V ratio, which we termed the metabolic index. Differences between M and F groups for these parameters were insignificant, as were the R values, which demonstrated extremely low results: 0.026 for V, 0.019 for S, and −0.032 for S/V.
Embryo sex and egg physical parameters
We included egg weight (W) and density values (D, D
i
and D
i
/D) in the group of physical parameters. Similar to the geometric parameters, eggs containing male and female embryos showed no significant differences with any of the physical parameters. Furthermore, no correlation was observed throughout the entire incubation period. The correlation between the M and F groups with W did not exceed 0.021 on Day 15 of incubation (Figure 4). Visualization of the results of egg weight, W, on Days 1 (A), 5 (B) and 15 (C) of incubation for the groups of eggs with male (M) and female (F) embryos.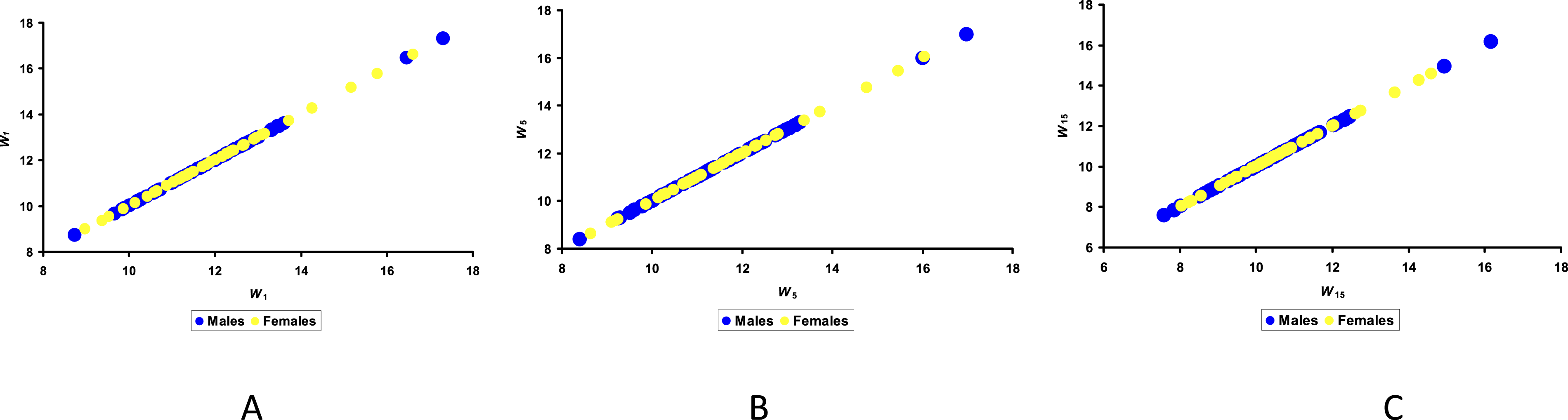
The major thing that can be concluded from analyzing the graphical dependences (Figure 4) is that the variations in weight values are somewhat greater for male eggs than for female ones. Moreover, these values decrease slightly for female eggs during incubation, indicating that eggs containing female embryos lose weight somewhat more quickly than those in the M group. At the same time, analysis by groups based on the rate of decrease in W during incubation also revealed no significant differences.
A similar situation was observed when analyzing egg density (D). The correlation between its values and the distribution across groups M and F, while somewhat higher than for W, was clearly insufficient and insignificant for the entire incubation period, reaching its maximum at −0.058 (Figure 5). Visualization of the results of egg density, D, on Days 1 (A), 5 (B) and 15 (C) of incubation for.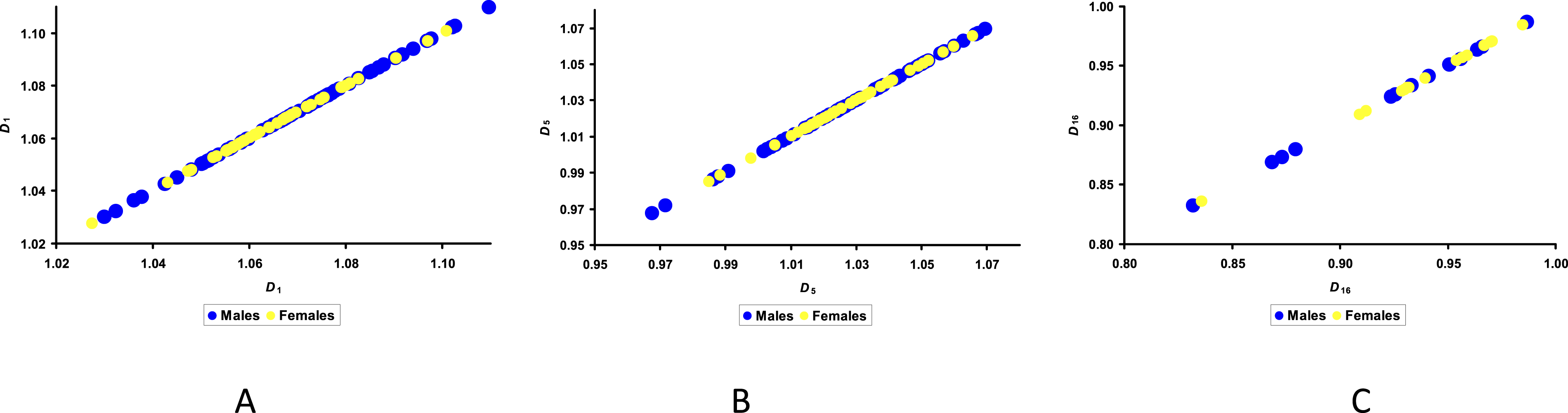
Unfortunately, neither the obtained D values nor the dynamics of their changes during the incubation process allow to make any adequate prediction that could bring us closer to solving the problem of separating eggs according to the sex of their embryos.
The resultant series of the values of the densities of egg interior (D
i
) demonstrated the highest R value of −0.137 compared to other physical parameters. However, neither its magnitude nor the differences in this parameter between groups M and F were significant (Figure 6). Visualization of the results of the density of egg interior, D
i
for the groups of eggs with male (M) and female (F) embryos.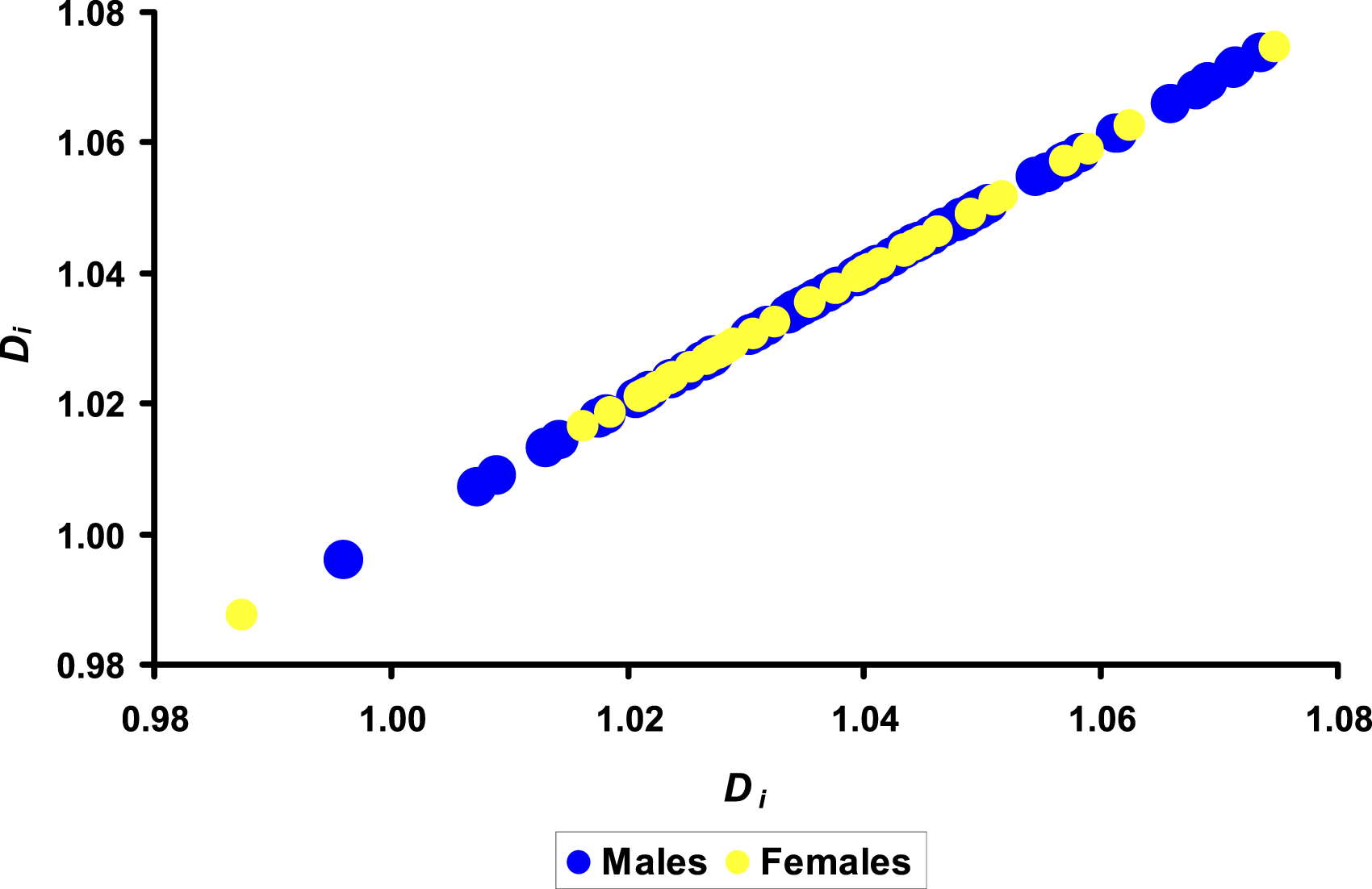
Although the eggs with minimum and maximum D i values correspond to female embryos, analysis of Figure 6 suggests that the boundary values of the density of egg interior are more characteristic of male eggs.
In contrast to our earlier experiments on chicken eggs, 45 in which the D i /D ratio showed significant differences, such effect was not observed in the present experiment.
Relationship between the heat transfer index of an egg and the sex of its embryo
The results of logical reasoning and theoretical assumptions demonstrated the feasibility of using the heat transfer index of the egg contents, calculated according to equation (5), in our analysis. However, we also considered the coefficient that accounts for the heat transfer of the whole egg, including its shell (equation (4)). To avoid confusion, we designated the heat transfer index of the egg interior as h i (equation (5)), and that of the whole egg as h e (equation (4)).
We first examined the characters of cooling curves for the eggs containing embryos of different sexes. We also attempted to determine the nature of possible changes in these curves across the days of incubation. To analyze this data, we used the average meanings of the measured temperatures for groups M and F (Figure 7). Cooling curves as averages for groups of eggs with male (M) and female (F) embryos on Days 1 (A), 5 (B) and 15 (C) of incubation.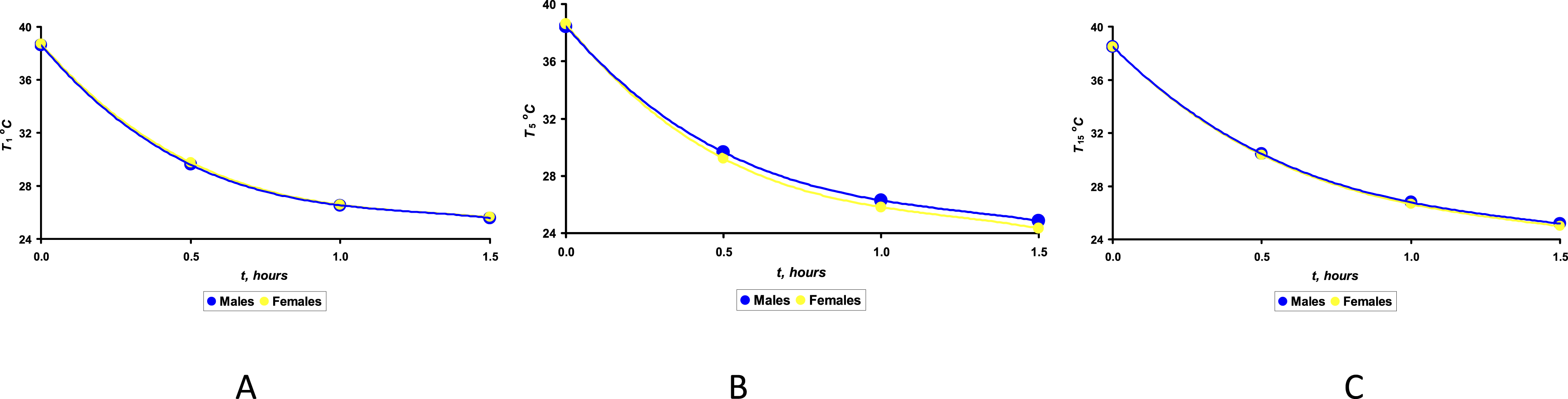
Analysis of the graphical dependences (Figure 7), firstly, confirmed their full compliance with the theoretical cooling curves of physical bodies presented in the classical literature on this topic.
63
This fact indirectly confirms the compliance of such a complex structure as a bird’s egg with the laws of the theory of cooling being elaborated for classical solids. Thus, we confirmed the validity of further analysis aimed at comparing the heat transfer indices of different eggs. Secondly, on Day 5 of incubation, we recorded the fact of significant differences (p < 0.05) in the cooling rate of eggs with female (yellow line) and male (blue line) embryos (Figure 7(B)). Within the same time (1.5 hours) and under the same environmental conditions (the ambient temperature was 24°C), the eggs of group F reached an average temperature of 24.1°C, while for group M this temperature was 24.8°C. No significant differences in cooling temperatures were observed for any other day of the incubation period. Therefore, our focus was on analyzing heat transfer indices, specifically on Day 5 of incubation. The h
i
and h
e
values were calculated for each egg, and a visualization of the resultant data, compiled for each M and F groups, is presented in Figure 8. Visualization of the results of the heat transfer indices, h
i
(A) and h
e
(B) for the groups of eggs with male (M) and female (F) embryos on Day 5 of incubation.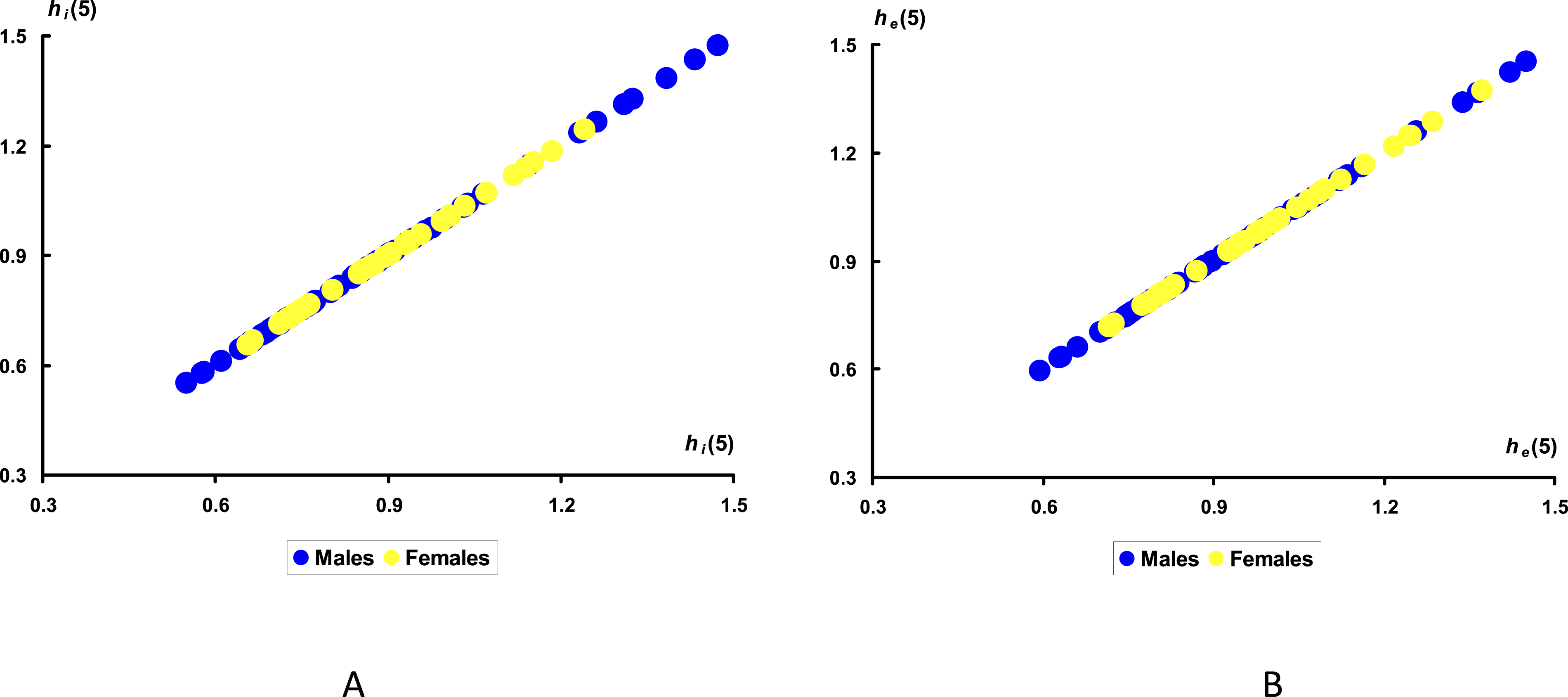
The obtained results demonstrated the clear success and absolute correctness of our assumption regarding the exclusion of the W s values from the calculation of the heat transfer indices as the key parameter for determining the sex of a particular egg. That is, the h i values (Figure 8(A)) encompassed a larger proportion of M eggs (blue dots), which extended beyond the F eggs (yellow dots). While h i could help to separate 12 eggs, representing approximately 18% of the total number of M group, using h e assisted in separation of only 5, or just over 7% of eggs with male embryos. A major success also lay in the clear delineation of the h i data area belonging to group M and extending beyond the common (central) zone characteristic of both groups M and F. The main advantage of the resultant distribution between the groups was that it was possible to isolate only the eggs of group M. In industrial poultry farming, males bear a lower industrial burden, as they are not involved in laying table eggs. Therefore, eggs containing males are subject to, if not 100%, then at least partial culling. The value of the heat transfer indices of the egg interior (h i ) proved particularly useful for this type of separation.
A logical question arises: how to calculate the regions corresponding to eggs of the group M? It is too early to give a definitive answer. Several options are possible. The first, and the simplest one is that the eggs with h i values being > 1.24 and < 0.65 contain male embryos and are therefore subject to rejection. A second option is that any sample of eggs may contain regions of minimum and maximum h i values characterizing the area of male embryos, like our results in Figure 8(A). Given the observed h i variations for the entire sample of eggs used in the experiment, at a level of 0.872 ± 0.189, regions containing male embryos correspond to those in which h i is above its mean value of +2σ and below its mean of –σ. A more accurate calculation can only be determined after conducting a series of additional experiments.
Another question was: Why did these differences arise on Day 5 of incubation? After all, no significant differences in the h
i
values between groups M and F were observed either earlier or later. To confirm this, Figure 9 shows the h
i
values for Days 4 and 6 of incubation. Visualization of the results of the heat transfer indices, h
i
for the groups of eggs with male (M) and female (F) embryos on Days 4 (A) and 6 (B) of incubation.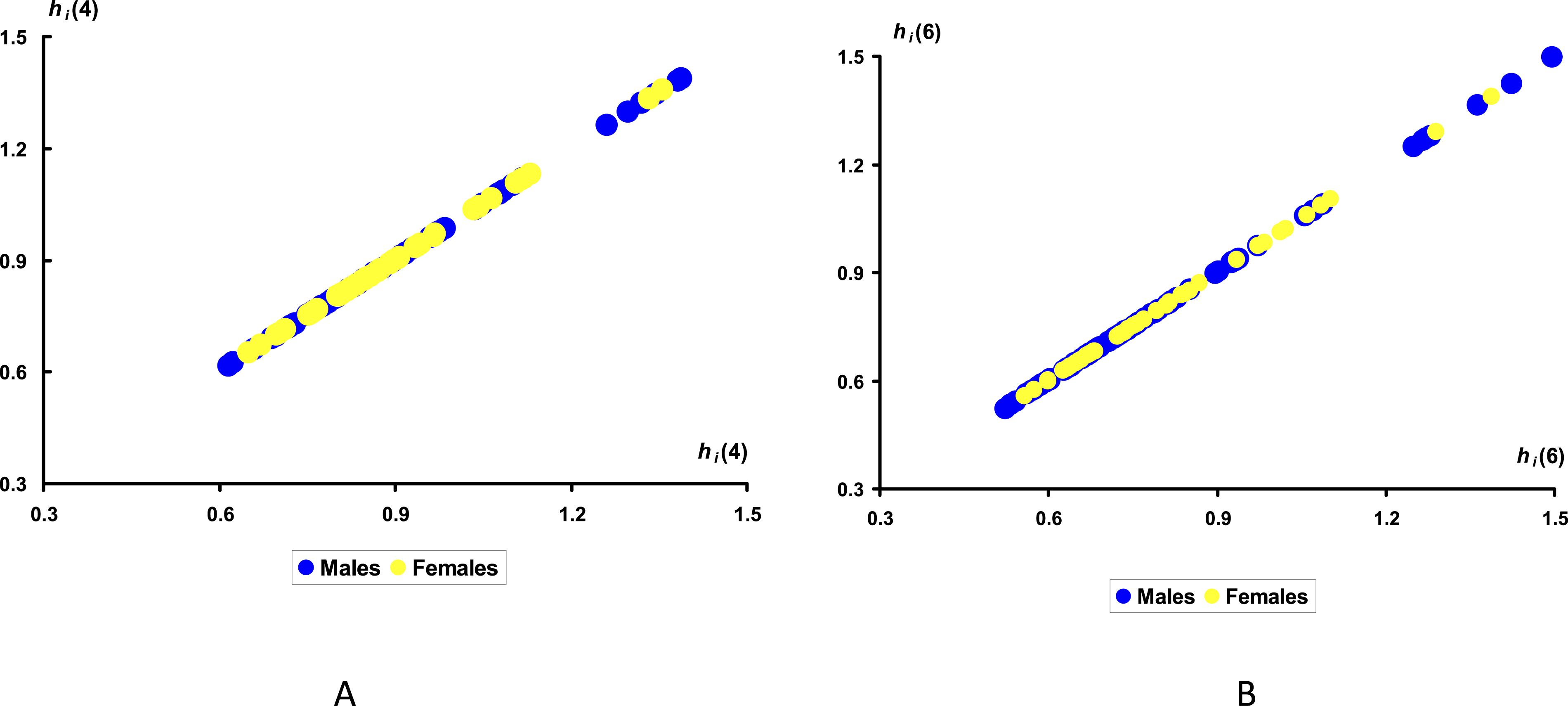
As shown by an analysis of studies of morphophysiological changes,87–90 the Japanese quail transitions from the early embryonic stage to the stage of intensive organogenesis on Day 5 of incubation. This can be one of the most “sudden” days of development. The main key changes occurring during this period include a sharp increase in the size of the embryo, intensified development of the cardiovascular system, the beginning of the formation of the respiratory system, and, importantly, the onset of differentiation of the sex gonads. The gonads are still the same in both sexes (indifferent), but it is during this period that the architecture is laid, along which the division into the ovary and testis later begins. Thus, Day 5 of incubation can be considered as the period of the most rapid morphogenesis. Apparently, this surge in embryonic changes also affects the temperature balance of the egg, leading to some differences in their heat transfer process that we were able to record between male and female embryos.
Conclusion and further steps
The results of the study confirmed the doubts of the skeptical majority, who believed that the secret of an embryo’s sex is somehow encoded in the geometric shape of its egg. Neither the shape of the egg’s contours nor the values of its physical parameters allowed for even a partial establishment of a significant correlation with sex. Only the logically and scientifically substantiated hypothesis of a difference in thermal conductivity at a certain point in embryonic development offers any hope of resolving this problem. The main achievement of this study is the discovery of a clear boundary between male and female embryos in the heat transfer indices of Japanese quail egg contents on Day 5 of incubation. This boundary, however, allowed for the exclusion of only about 18% of males, the mere discovery of such differences can be considered as an achievement. The obtained results are extremely promising and open a whole new field of research, experiments, and analytical perspectives. What can be improved, and where to go further?
First, discrete data of the temperature collection should be replaced with continuous monitoring of the egg cooling process. It is possible that the temperature curve itself could provide additional information on the embryonic characteristics of the eggs.
Second, it may be more promising to increase the number of temperature measurement points along the shell surface. This is due to both the shell’s variable thickness from the sharp end to the blunt end, and the potential location of the embryo inside the egg.
Third, the accuracy of shell weight calculations should be improved, going beyond the empirical relationship. It is possible that several instruments, such as non-destructive elastic deformation91,92 and/or an ultrasonic caliper, 93 could prove invaluable in the non-destructive determination of W s .
The relevance and demand for the process of embryo sex determination in quail eggs is inferior to that in chicken eggs. This is due to both the incubation volumes and the ethical issues of culling day-old male chicks.12–14 Nevertheless, the fact that quail can be considered a model species in research relevant to industrial poultry farming has been undeniably recognized,94–96 including for the study of embryonic changes and the development of incubation techniques. 88 Thus, it can be safely stated that the relationship between the heat transfer index of the egg interior can be a potential solution to the problem of sex determination in the early stages of incubation for a wide range of poultry species.
Footnotes
Author contributions
Valeriy G. Narushin: conceptualization, data curation, formal analysis, investigation, methodology, resources, software, visualization, writing – original draft, writing – review & editing. Natalia A. Volkova: data curation, funding acquisition, investigation, methodology, project administration, supervision, validation, writing – review & editing. Danila A. Sotnikov: investigation, validation. Anastasia N. Vetokh: investigation, validation. Ludmila A. Volkova: investigation, validation. Darren K. Griffin: supervision, writing – review & editing. Michael N. Romanov: project administration, validation, visualization, writing – original draft, writing – review & editing. Natalia A. Zinovieva: funding acquisition, project administration, supervision, writing – review & editing.
Funding
This research work was supported by the Russian Science Foundation (Grant No. 24-16-00294).
Declaration of conflicting interests
The authors declare no conflicts of interest related to this work. The funders had no role in the design of the study; in the collection, analyses or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
Author statement
All procedures in this trial adhered to standard husbandry and experimentation practices as also reviewed and approved by the responsible LKEFRCAH ethics committee (see details in Ethics statement section). Research data is available upon request. To request the data, contact the corresponding author of the article. During the preparation of this work, the authors did not use IA tools.