
Abstract
Avian hybridization has been predominantly studied in natural contexts, such as hybrid zones or the identification of hybrid individuals. However, captive settings offer a complementary framework for understanding the ecological and evolutionary mechanisms underlying hybridization. Artificial conditions might relax premating isolation barriers that typically prevent interspecific breeding in the wild, although certain mate recognition mechanisms could remain effective in captivity. Here, I present a comparative analysis of wild and captive hybrids in owls (order Strigiformes; families Tytonidae and Strigidae) to test two predictions: (1) captivity facilitates hybridization between morphologically more divergent species than observed under natural conditions, and (2) certain premating isolation mechanisms are still effective in captivity, with a focus on iris coloration as a visual cue in species recognition. In total, I compiled an overview of 26 reported cases of owl hybridization. Of these cases, 11 (42%) were supported by reliable documentation, involving 19 species (8% of 244 assessed species). Seven additional cases (27%) remain provisional, and eight (31%) were deemed unreliable. Comparative analyses of morphological traits indicated that captive hybrid pairs showed a broader range of interspecific differences in body mass, tarsus length, and wing length than natural hybrid pairs. No consistent differences were found between captive and natural hybrid pairs in terms of iris coloration, activity pattern, or trophic niche. Overall, hybridization in owls appears rare. The tendency for captive hybrids to involve morphologically divergent species suggests that artificial conditions relax certain constraints on interbreeding. However, the similarity in iris coloration among most hybridizing species supports a potential role for this trait in premating isolation. Future studies integrating genomic data, experimental approaches, and broader comparative datasets will be needed to clarify the impact of captivity on reproductive barriers.
Introduction
Hybridization in birds is a widespread phenomenon, with an estimated 10–20% of species reported to have hybridized in the wild 1 . However, this figure does not account for hybridization in captivity, where interspecific crosses might occur under conditions that differ markedly from natural settings2,3. To date, most research on avian hybridization has focused on natural systems, particularly the study of hybrid zones4–6 and the documentation of naturally occurring hybrid individuals7,8. While these studies have greatly advanced our understanding of reproductive isolation and introgression, captive crosses can also provide valuable insights into the ecological and evolutionary mechanisms underlying hybridization. For example, studies of wildfowl (Galliformes) and of pigeons and doves (Columbiformes) have used captive crosses to assess the fertility and viability of first-generation hybrids and subsequent backcrosses, shedding light on the evolution of postzygotic isolation9,10. Similarly, the description of captive hybrids between the Chiloé Wigeon (Mareca sibilatrix) and the Philippine Duck (Anas luzonica) has informed hypotheses about the evolution of sexual dichromatism in waterfowl 11 . Together, these examples highlight the added value of captive hybrids in understanding the processes that shape avian hybridization and speciation.
The artificial conditions of captivity can lead to the breakdown of premating isolation mechanisms that would normally prevent interspecific breeding in the wild 12 . In natural environments, assortative mate choice can be reinforced by behavioral displays 13 , acoustic signals 14 , and ecological preferences15,16. In captivity, however, individuals might be confronted with a limited pool of potential mates, sometimes including no conspecifics at all. Under these circumstances, heterospecific pairing might occur, either through relaxed mate discrimination or through active mate choice in the absence of alternatives17,18. In addition, captive hybridization may result from early-life imprinting or learned mate preferences on heterospecific individuals19,20. The relationship between imprinting and hybridization has been documented through cross-fostering experiments in wild populations of, among others, tits 21 , gulls 22 and geese 23 (but see this study 24 on flycatchers where cross-fostering did not impact mate choice). Similarly, captive cross-fostering experiments with European Kestrel (Falco tinnunculus) and American Kestrel (Falco sparverius) reported consistent mate preference for the foster parent. 25 These situations can give rise to hybrids between species that are geographically isolated in nature or that differ substantially in morphology, behavior, or ecology11,26. As a result, captivity can facilitate hybridization between more distantly related or phenotypically divergent species than would typically interbreed in the wild, such as Helmeted Guineafowl (Numida meleagris) x Chicken (Gallus gallus) or Helmeted Guineafowl x Indian Peafowl (Pavo cristatus), two species pairs that diverged more than 45 million years ago.27,28. Despite these striking examples, the prediction that captivity produces hybrids between morphologically more divergent species has not been formally tested in a comparative framework.
Nevertheless, premating isolation mechanisms could remain sufficiently robust to limit hybridization under captive conditions. Species-specific courtship displays, vocalizations, visual signals, and behavioral preferences often play a central role in mate recognition, and these mechanisms might persist in captivity 12 . Individuals might continue to preferentially select conspecific partners when available, or they might fail to respond to heterospecific courtship signals. For example, Baker 18 found that female Indigo Buntings (Passerina cyanea) and Lazuli Buntings (P. amoena) responded more to conspecific than to heterospecific male traits in captivity. In some cases, differences in body size, mating behavior, or reproductive timing may further impede successful pairing 29 . Consequently, although captivity can reduce ecological and geographic barriers, certain behavioral and recognition-based premating barriers could still constrain the formation of interspecific pairs. Determining the extent to which these barriers are weakened (or remain intact) in captivity can inform our understanding of premating isolation mechanisms.
In this study, I used hybridization in owls (order Strigiformes; families Tytonidae and Strigidae) to test two predictions. First, I examined whether captivity facilitates hybridization between morphologically more divergent species than occurs in nature. If premating barriers are relaxed under captive conditions, I would expect crosses between species that differ more strongly in morphological and ecological traits. Second, I tested whether specific premating isolation mechanisms are weakened in captivity. With respect to the second prediction, I focused on eye color as a potential cue in species recognition. In owls, iris coloration is closely associated with activity pattern and dominant lifestyle (e.g., diurnal versus nocturnal species), and might thus function as a conspicuous visual signal during mate choice 30 . In other avian groups, such as seabirds, similar visual traits have been proposed to contribute to premating isolation by facilitating assortative mating31,32. If eye color plays a role in species recognition in owls, natural hybridization should occur more frequently between species sharing similar iris coloration. In contrast, captive conditions may weaken the importance of such cues, allowing hybridization between species with different iris colors. Owls provide a suitable study system to explore these predictions as both natural and captive hybrids have been documented across multiple taxa.
Methods
I first compiled an evidence-based overview of hybridization in owls. Hybrid records were collected from three primary sources: the Handbook of Avian Hybrids of the World 33 , the online Serge Dumont Bird Hybrid Database (https://bird-hybrids.com/), and species accounts on the Birds of the World website 34 . For each reported hybrid combination, all primary references were retrieved and critically evaluated.
To assess the reliability of hybrid records, I applied the scoring scheme proposed by Ottenburghs 35 , which assigns different weights to particular types of evidence: observational reports (1 point), detailed morphological descriptions of museum specimens (2 points), and genetic confirmation (3 points). Because captive hybrids are often poorly documented in terms of morphology and genetics, I adapted this scoring scheme for captive records. Specifically, general reports or observational accounts of captive hybrids were assigned 1 point, whereas descriptions published in peer-reviewed journals were assigned 2 points. In addition, I did not discriminate between the circumstances of the captive birds (e.g., whether the parental birds were permanently housed together, were part of a breeding program or arrived at a rehabilitation center), because these details were not available for most cases. Only hybrid records with a minimum score of two points were retained for subsequent statistical analyses to ensure a conservative dataset.
Morphological (body mass, tarsus length, and wing length) and ecological data (trophic niche) for the parental species were obtained from the AVONET database
36
. Trophic niche included the relevant AVONET classifications (i.e. vertivore, invertivore, omnivore and aquatic predator). Eye color was classified based on species descriptions and color plates in Owls of the World
37
. Iris coloration was grouped into three main categories: yellow, orange, and brown (Figure 1). Previous work by Passarotto et al.
30
indicated that iris coloration is associated with activity pattern, with darker eyes being more common in strictly nocturnal species and brighter eyes more frequently observed in diurnal or crepuscular species. Hence, I also included the classification of nocturnal and diurnal owl species (following Passarotto et al.
30
). For each hybridizing species pair, I coded trophic niche, eye color and activity pattern as either matching (i.e. both species falling within the same category) or non-matching (i.e. species assigned to different categories). Together, all these traits were used to quantify morphological and ecological similarity between the parental species of the hybridizing species pairs. Overview of the three eye colors (yellow, orange and brown) used in the analyses, illustrated by representative owl species: Great Grey Owl (Strix nebulosa), Eurasian Eagle-Owl (Bubo bubo) and Barred Owl (Strix varia). Image credits can be found in the supplementary material.
All statistical analyses were conducted in R version 4.5.1. Morphological divergence between parental species was quantified as the absolute difference in body mass (grams), tarsus length (mm), and wing length (mm). All measurements in the AVONET database are species-level averages that include both male and female specimens. For each trait, morphological differences between captive and natural hybrid pairs were tested using non-parametric Kruskal–Wallis tests. To account for potential allometric scaling effects, the analyses were repeated using log-transformed trait values. For categorical variables (eye color, activity level and trophic niche), species pairs were classified as matching (1) or non-matching (0). The frequencies of matching versus non-matching species pairs were subsequently compared between captive and natural hybrid pairs using chi-square tests with simulated p-values.
Results
An evidence-based overview of hybrid owl records
Overview of hybrid owl records with supporting evidence. Detailed descriptions of all hybrid combinations can be found in the main text.
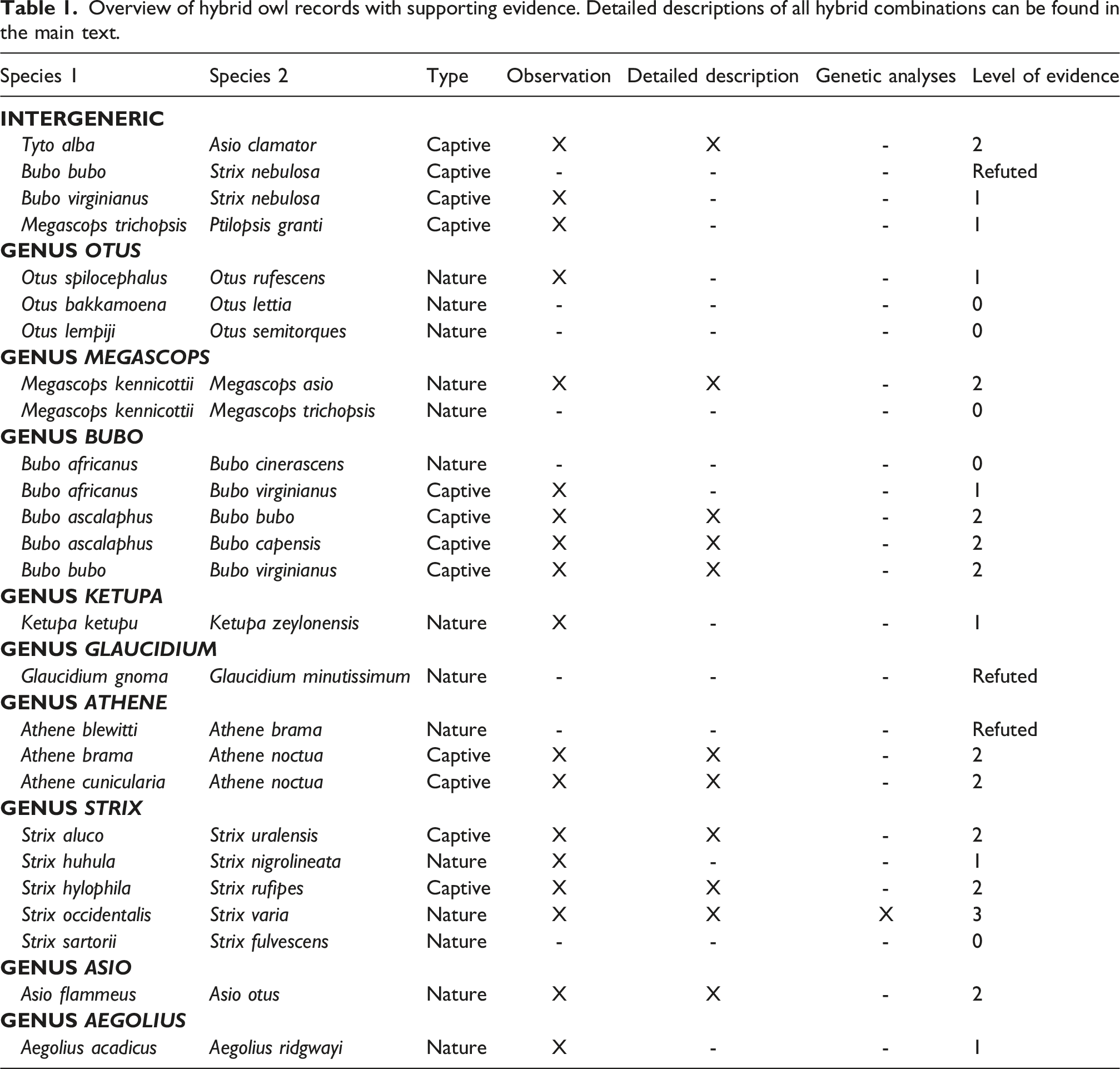
No interspecific hybridization was reported for the genera Phodilus, Psiloscops, Gymnasio, Lophostrix, Jubula, Pulsatrix, Scotopelia, Surnia, Taenioptynx, Micrathene, Xenoglaux, Ninox, and Uroglaux. In the following sections, I will provide a genus-level overview of all documented hybrid records, summarizing the supporting evidence and distinguishing between natural and captive cases. For the sake of completeness, I have also included hybrid records that are currently not supported by reliable evidence (i.e. reliability score of zero) or have been refuted.
Several hybrid records have been reported within the genus Otus, although the strength of evidence varies considerably. Two proposed species combinations – Indian Scops-Owl (Otus bakkamoena) x Collared Scops-Owl (Otus lettia) and Sunda Scops-Owl (Otus lempiji) x Japanese Scops-Owl (Otus semitorques) – are listed in the Serge Dumont Hybrid Database. However, the Birds of the World species accounts note that these taxa have been treated as conspecific 34 . In the absence of direct morphological or genetic evidence supporting hybridization, these records likely reflect taxonomic uncertainty rather than confirmed interspecific crosses. Another putative case concerns a specimen of the race ‘stresemanni’, which has been suggested to represent a hybrid between Mountain Scops-Owl (Otus spilocephalus) and Reddish Scops-Owl (Otus rufescens) on the basis that all measured characters were intermediate between the two taxa 38 . However, this claim does not appear to be supported by a peer-reviewed publication or detailed morphological assessment, and thus remains only marginally supported.
Within the genus Megascops, hybridization has been documented for Western Screech-Owl (Megascops kennicottii) x Eastern Screech-Owl (Megascops asio). Mixed pairs and putative hybrids have been reported from multiple locations, with several independent accounts in the literature39–41, providing consistent support for natural hybridization between these two species. In contrast, the proposed hybridization between Western Screech-Owl and Whiskered Screech-Owl (Megascops trichopsis) is unreliable. This species combination is mentioned in the Handbook of Avian Hybrids of the World 33 , which cites Duncan 42 as the source. However, the referenced work merely describes the altitudinal distributions of the two species, without presenting direct evidence for hybridization.
One putative wild hybrid combination has been suggested within the genus Bubo, although the strength of supporting evidence is questionable. For Spotted Eagle-Owl (Bubo africanus) x Grayish Eagle-Owl (Bubo cinerascens), Keith and Twomey 43 described several specimens of the subspecies B. a. tanae exhibiting intermediate morphological traits and suggested that intergradation between tanae and cinerascens might be expected in Kenya. However, Birds of the World notes that these species differ consistently in morphology and are not known to interbreed where their ranges overlap 44 . Consequently, there is no strong evidence for hybridization between these species. Several interspecific crosses within the genus Bubo have been reported under captive conditions. Hybrids between Pharaoh Eagle-Owl (Bubo ascalaphus) and Eurasian Eagle-Owl (Bubo bubo) were documented in Tel Aviv Zoo (Israel)45–49, and crosses between Pharaoh Eagle-Owl and Cape Eagle-Owl (Bubo capensis) were reported in Antwerp Zoo (Belgium)47,49,50. In addition, the successful rearing of a captive hybrid between Eurasian Eagle-Owl (Bubo bubo) and Great Horned Owl (Bubo virginianus) was described in an avicultural magazine 51 . Finally, the combination Spotted Eagle-Owl (Bubo africanus) x Great Horned Owl (Bubo virginianus) has been mentioned in an identification guide 52 , but detailed documentation on this hybrid combination is limited.
A potential case of hybridization has been reported between Buffy Fish-Owl (Ketupa ketupu) and Brown Fish-Owl (Ketupa zeylonensis). An observation on eBird documents a mixed pair accompanied by a putative hybrid juvenile (Macaulay Library asset 371348391). However, this hybrid record is solely based on field observations and photographic evidence, and no detailed morphological assessment or genetic confirmation is currently available. Additional documentation is therefore required to substantiate this case.
A possible case of hybridization between Northern Pygmy-Owl (Glaucidium gnoma) and Least Pygmy-Owl (Glaucidium minutissimum) was suggested by Griscom 53 , who proposed the occurrence of intergradation between the two taxa. However, this interpretation was later rejected by Monroe Jr. 54 , who stated that he could find no evidence of intergradation anywhere within the ranges of either species (p. 157). Consequently, there is currently no substantiated support for hybridization between these species.
Within the genus Athene, a possible case of natural hybridization has been reported between Forest Owlet (Athene blewitti) and Spotted Owlet (Athene brama). Pande et al. 55 described a putative hybrid individual from northern India. However, the scientific quality and validity of this record have been questioned by other authors, and the case remains controversial 56 . In contrast, two reliable cases of captive hybridization have been described. Crosses between Spotted Owlet and Little Owl (Athene noctua) were conducted experimentally by Stocks 57 , who reported that the resulting hybrids were infertile. The same author also noted a cross between Burrowing Owl (Athene cunicularia) and Little Owl, carried out at the Owl Center at Baytree Nursery by Jim Dean. These hybrids were also reported to be infertile.
Within the genus Strix, several hybrid combinations have been documented, with variable levels of support. In captivity, Tawny Owl (Strix aluco) x Ural Owl (Strix uralensis) hybrids have been reported 58 . Although the ranges of these species overlap in nature, there is currently no confirmed evidence of wild hybrids 59 . A captive hybrid between Rusty-barred Owl (Strix hylophila) and Rufous-legged Owl (Strix rufipes) was documented at the Berlin Zoo (Germany) 60 . Among natural cases, hybrids between Spotted Owl (Strix occidentalis) and Barred Owl (Strix varia) are well-documented61,62 and have been confirmed genetically63,64. A putative hybrid between Black-banded Owl (Strix huhula) and Black-and-white Owl (Strix nigrolineata) – also known as the San Isidro Mystery Owl – has been reported in southeastern Colombia. This putative hybrid combination was already mentioned by Meyer de Schauensee 65 who wrote that “Blake (l.c.) reports a bird from southeastern Colombia as intermediate with nigrolineata” (below the description of the Black-banded Owl, p. 145). However, hybridization has not been assessed morphologically or genetically. Similarly, although the apparent range replacement of Cinereous Owl (Strix sartorii) by Fulvous Owl (Strix fulvescens) in Oaxaca suggests the possibility of hybridization, no direct evidence exists 66 .
A hybrid specimens between Short-eared Owl (Asio flammeus) and Long-eared Owl (Asio otus) from the collections of the Canadian Museum of Nature has been described morphologically 67 . However, this specimen has not been confirmed using genetic analyses.
The taxon tacanensis has been suggested to be a hybrid between Northern Saw-whet Owl (Aegolius acadicus) and Unspotted Saw-whet Owl (Aegolius ridgwayi). König and Weick 37 recognize this form as a possible hybrid, but note that further field investigations are needed to confirm this interpretation.
Finally, a few intergeneric owl hybrids have been reported under captive conditions. At the St. Louis Zoological Garden, a male Western Barn Owl (Tyto alba) paired with a female Striped Owl (Asio clamator), producing four eggs in the first clutch (two fertile and two infertile) and two additional clutches over the following 15 months, although no further eggs were fertile 68 . A hybrid between Great Gray Owl (Strix nebulosa) and Eurasian Eagle-Owl (Bubo bubo) was erroneously reported in the Handbook of Avian Hybrids of the World. The original source 52 referred to separate captive hybrids of each species with Great Horned Owl (Bubo virginianus). McCarthy 33 misinterpreted this information to mean that Eagle Owl and Great Gray Owl have hybridized (Peter Pyle, personal communication). In contrast, captive hybridization between Great Gray Owl and Great Horned Owl has been reported 52 , although detailed documentation is limited. An unusual case of interspecific hybridization was recently brought to my attention (Calvin Becker, personal communication). According to information displayed at the Dullstroom Bird of Prey and Rehabilitation Centre in South Africa, a Whiskered Screech-Owl (Megascops trichopsis) paired with a Southern White-faced Owl (Ptilopsis granti) in captivity and reportedly produced two hybrid offspring. These species have non-overlapping natural distributions: the Whiskered Screech-Owl occurs in North and Central America, whereas the Southern White-faced Owl is native to sub-Saharan Africa. However, this case remains to be confirmed with detailed morphological or genetic analyses.
Morphological and ecological comparison of captive and wild owl hybrids
For the statistical analyses, only hybrid records with a minimum reliability score of two were retained (N = 11), comprising eight captive and three wild species combinations. Across all three morphological traits, parental species involved in captive hybridization exhibited a broader range of size differences than those involved in natural hybridization (Figure 2). Although these differences were not statistically significant, captive hybrid pairs tended to show greater divergence in body mass (Figure 2(a); Kruskal-Wallis test: χ2 = 1.05, p = 0.31), tarsus length (Figure 2(b); χ2 = 3.02, p = 0.08), and wing length (Figure 2(c); χ2 = 0.67, p = 0.41) compared to wild hybrid pairs. Repeating the analyses using log-transformed trait values to account for allometric scaling did not qualitatively alter the results. Likewise, including all hybrid records with a reliability score of at least one produced similar patterns (Table S1). However, when these hybrid records were included, differences in tarsus length and wing length between captive and natural hybrid pairs reached statistical significance (p = 0.01 and p = 0.03, respectively), whereas body was marginally non-significant (p = 0.052). Comparison of divergence in three morphological traits between the parental species of captive and wild owl hybrids: (a) Body mass, (b) Tarsus length, and (c) Wing length.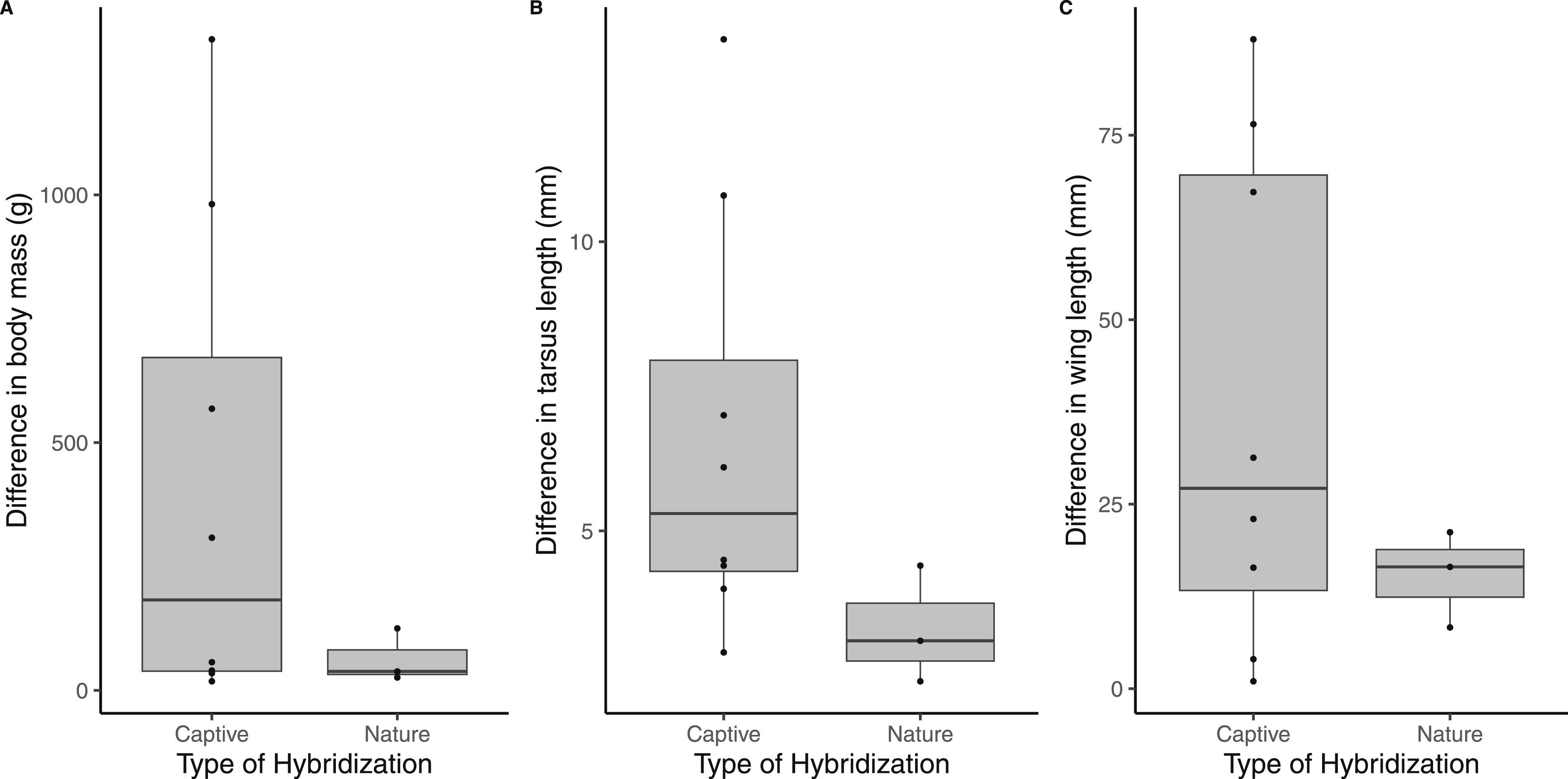
There were no clear differences between captive and natural hybrid pairs with respect to eye color (chi-square test with simulated p-value: χ2 = 0.41, p = 1.00), activity pattern (χ2 = 2.93, p = 0.27), or trophic niche (χ2 = 0.92, p = 0.56). Thus, hybridizing species did not differ systematically in these ecological or visual traits depending on whether hybridization occurred in captivity or in the wild. Notable exceptions included captive hybrids between Eurasian Eagle-Owl and Great Horned Owl, whose parental species differ in eye color (yellow versus orange), and natural hybrids between Spotted Owl and Barred Owl, whose parental species differ in activity pattern (diurnal versus nocturnal). However, these cases did not influence the overall patterns. Repeating the analyses using all hybrid records yielded similar results (Table S2).
Discussion
My survey indicates that hybridization in owls is relatively rare. Only 8% of the 244 assessed species were involved in hybrid combinations supported by reliable documentation. This proportion is comparable to estimates for parrots (7.5%) 2 , but markedly lower than estimates reported for woodpeckers (19%) 69 and terns (34%) 70 . Several factors might contribute to the apparent rarity of owl hybrids. First, many owl species have largely allopatric distributions, including numerous island endemics, limiting opportunities for secondary contact and interbreeding 37 . Second, owl hybrids may be difficult to detect morphologically, particularly in groups where plumage differences between species are subtle 71 . Third, genetic investigations of hybridization in owls remain scarce. To date, only a single hybrid case (Spotted Owl x Barred Owl) has been confirmed with molecular data63,64. Given the growing evidence that introgression – genetic exchange following hybridization – is widespread across birds72,73, it is plausible that genomic analyses will reveal additional, cryptic cases of hybridization in owls e.g.,74,75. Finally, owl hybridization could genuinely be rare due to strong prezygotic or postzygotic reproductive barriers 29 . Expanded behavioral, morphological and genetic sampling across contact zones is therefore needed to properly asses the incidence of hybridization in this bird order.
As predicted, the parental species involved in captive hybridization tended to be morphologically more divergent than those hybridizing in the wild. Some of the most extreme differences occurred within the genus Bubo, where captive hybrid pairs included species differing by approximately 500 to more than 1000 grams in body mass 51 . In contrast, the largest size difference among naturally hybridizing species was observed between Spotted and Barred Owl, which differ by roughly 125 grams. However, these comparisons are based on species-level averages derived from published datasets 36 . The actual body sizes of the individuals involved in the hybridization events could not be determined from the available references. It is therefore possible that some interspecific pairings involved unusually small or large individuals, potentially facilitating interbreeding. In addition, the sample size for the comparative analysis was limited (with only eight captive and three natural hybrid combinations), which reduces statistical power and might have obscured subtle patterns. Future studies should replicate this approach in other avian groups with numerous documented captive hybrids, such as waterfowl23,76 and wildfowl10,26, where larger datasets might allow for more robust tests of whether captivity promotes hybridization between morphologically more divergent species.
In contrast to the pattern observed for body size, I found no clear relationship between eye color similarity and the occurrence of hybridization. Most hybridizing species pairs (both in captivity and in the wild) shared similar iris coloration. One notable exception was a hybrid between Eurasian Eagle-Owl (orange eyes) and Great Horned Owl (yellow eyes) that was reared in captivity 51 . At first glance, the predominance of matching eye colors among hybridizing species pairs could suggest that iris coloration plays an important role in premating isolation in owls. However, this interpretation should be treated with caution. A critical limitation of the present analysis is the lack of an appropriate control for captive conditions. Ideally, one would compare hybridizing species pairs with other owl species that were housed together in captivity but did not interbreed. If eye color functions as a species-recognition cue, one would expect that species with different iris coloration would be less likely to hybridize, even when kept in close proximity. However, variation in captive conditions can also influence the probability of hybridization. Individuals housed together from an early age could imprint on heterospecific characteristics, increasing the likelihood of interspecific mating 19 . In contrast, individuals that encounter heterospecifics later in life (for example, in rehabilitation centers or following transfers between zoos) might be less likely to form pair bonds with heterospecific mates. Unfortunately, such comparative housing data were unavailable. Furthermore, closely related species tend to share similar eye colors, potentially confounding the analysis 30 . All confirmed natural hybrids occurred between species within the same genus, making it difficult to disentangle the effects of phylogenetic relatedness and eye color similarity. Nonetheless, some intergeneric captive hybrids involved species with similar iris coloration, such as Western Barn Owl and Striped Owl 68 . Taken together, these findings provide tentative support for a role of eye color in premating isolation, highlighting the need for more detailed behavioral and experimental data.
In conclusion, this study provides the first comparative assessment of natural and captive hybridization in owls and offers tentative insights into the role of morphological divergence and premating isolation. Hybridization in owls appears relatively rare, with only a small proportion of species (8%) involved in well-supported hybrid combinations. As predicted, captive hybrids tended to involve morphologically more divergent parental species than natural hybrids, suggesting that artificial conditions can relax certain constraints on interspecific mating. However, most hybridizing species shared similar iris coloration, providing some support for a role of this trait in premating isolation. Future studies incorporating genomic data, experimental approaches, and larger comparative datasets across different avian groups will be essential to clarify how captivity impacts the strength of reproductive barriers.
Supplemental material
Supplemental material - Comparing captive and wild owl hybrids reveals divergence in size of parental species, but not in iris coloration
Supplemental material for Comparing captive and wild owl hybrids reveals divergence in size of parental species, but not in iris coloration by Jente Ottenburghs in Avian Biology Research.
Supplemental material
Supplemental material - Comparing captive and wild owl hybrids reveals divergence in size of parental species, but not in iris coloration
Supplemental material for Comparing captive and wild owl hybrids reveals divergence in size of parental species, but not in iris coloration by Jente Ottenburghs in Avian Biology Research.
Footnotes
Acknowledgements
I would like to thank the anonymous reviewer for the constructive feedback and Peter Pyle for checking several owl hybrids in his identification guide to North American birds. And I am grateful to the librarians of the Natural History Museum London and the British Library for providing me with scanned copies of relevant papers.
Ethical considerations
Not applicable. All data is based on published information.
Funding
No funding is associated with the study.
Declaration of conflicting interests
The author declares no conflict of interest.
Data Availability Statement
All data associated with the study is available in the supplementary material.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.