
Abstract
Corvids such as magpies can cause economic losses and management conflicts in urban and agricultural landscapes, and repeated exposure can reduce the effectiveness of non-lethal deterrents through habituation. This study evaluated short-term behavioral responses and habituation patterns in a flock of magpies (Pica serica) that repeatedly foraged at the same site. A drone-only stimulus and a combined stimulus consisting of a drone carrying a taxidermized magpie with conspecific mobbing-call playback were applied sequentially six and five times, respectively, at two-day intervals. Each trial consisted of 10 min of stimulus exposure followed by 50 min of post-stimulus observation. Magpie numbers were recorded at 5-min intervals, and behaviors were classified as predator-related or predator non-related based on field observations. Audio recordings were also collected to interpret mobbing calls and behavioral context. Under the drone-only stimulus, predator-related behavior increased immediately after stimulus onset, but this response declined over repeated trials, and predator non-related behavior recovered even during stimulus exposure, suggesting rapid habituation. Under the combined stimulus, no magpies remained in the experimental area during stimulus exposure in any trial, and in early trials they did not return before the end of observation. However, in later trials, individuals showing non-predator-related behavior re-entered the area 20–25 min after stimulus removal, indicating that habituation was delayed but not fully suppressed. These results suggest that a species-specific combined stimulus can induce stronger short-term avoidance and delay habituation compared with drone-only stimulus, but additional strategies are required to maintain deterrent effectiveness during repeated applications.
Introduction
Conflicts between farmers and pest wildlife in rural areas have persisted for decades, and among them, corvid birds (Corvidae) are representative pest species that cause direct economic losses by reducing yield and marketability of specific crops during vulnerable periods. In sunflower, bird damage can account for approximately 14∼35% of revenue loss 1 , and in France, bird damage during emergence and the seedling stage has been identified as a major risk factor determining crop establishment; this can go beyond partial loss to increase the need for re-sowing and overall production uncertainty 2 . Damage is also evident in fruit orchards. In tankan (Citrus tankan) orchards in Okinawa, Japan, farmer surveys report that an average of 13.5% of fruits are damaged by wildlife, with the large-billed crow (Corvus macrorhynchos) accounting for 53∼56% of the damage 3 . In the Republic of Korea, magpies (Pica serica) primarily damage fruit crops such as apples and pears by pecking the fruit surface, thereby reducing their marketability. In 2018, crop damage caused by magpies was estimated at KRW 1.02 billion 4 . In this context, studies have examined the fruit-feeding behavior of magpies to better understand the types of damage inflicted on fruit crops 5 , and additional research has evaluated damage prevention methods using conditioned taste aversion 6 .
When damage by pest birds is substantial, demands for population reduction tend to intensify. However, lethal control targeting corvids often involves social controversy and questions of management legitimacy, and concerns have been raised that its damage-reduction benefits are not consistently verified in the field yet continue as routine practice 7 . Therefore, to achieve meaningful reductions in crop damage while ensuring social acceptability and sustainability, preventive management strategies centered on non-lethal deterrence should be prioritized over direct removal of wildlife.
Non-lethal deterrence can be broadly categorized into physical, chemical, and visual–acoustic stimulus-based approaches. Physical methods such as netting can be clearly effective by preventing access, but installation and maintenance costs increase substantially when applied at large farm scales 8 . Chemical deterrence uses substances that target species avoid; although some efficacy has been reported 9 , its performance can vary across species and environmental conditions, it can be costly, and effectiveness in real-world settings may be lower than in experimental contexts10,11. Visual and acoustic deterrents include predator decoys, lasers, ultraviolet devices, acoustic playback systems, and propane cannons, and they are relatively easy to deploy over wide areas 12 . However, stimulus-based deterrence may rapidly lose effectiveness with repeated use due to habituation13,14.
Animals can quickly adapt by modifying behavior through learning and habituation in response to repeated external stimuli. Habituation is defined as a reduction in behavioral responses to repeated stimuli without accompanying physiological changes, and deterrent stimuli that are initially effective may weaken or cease to be effective when applied repeatedly 13 . To suppress habituation and maximize deterrence, strategies have been proposed to elicit predator related behavior by audio visually recreating predator scenarios that birds are likely to perceive as genuine threats, rather than relying on simple visual or acoustic cues alone 15 . Birds also share risk information within groups via vocal signals, and alarm calls function as key cues that trigger collective defensive behavior 16 . Through such vocal signals, individuals can acquire risk information and adjust behavior even without direct predator encounters 17 .
Corvids are highly developed in social communication and threat recognition. Black billed magpies (Pica pica), in particular, have a complex signaling system in which mobbing and alarm calls vary according to predator context, and call syllable duration can influence approach or avoidance responses 18 . Beyond species-level recognition, magpies can discriminate threatening from harmless targets at the individual level 19 . In addition to predator recognition, corvids also exhibit advanced conspecific recognition. Crows can integrate visual and auditory information to identify conspecifics at the individual level 20 . Furthermore, based on their advanced cognition and sociality, corvids can acquire threat-related information through adult behavior and vocal signals, and they can continue to recognize novel objects or stimuli as threats through social learning even after reaching adulthood. Ravens (Corvus corax) rapidly learned to recognize individuals holding dead conspecifics as dangerous, and some individuals retained this learned information for up to four years 21 . Similarly, studies of American crows (Corvus brachyrhynchos) have shown that individuals can learn to recognize specific targets as dangerous by observing mobbing responses from conspecifics, even without direct threatening experience 22 . Therefore, combined stimuli that reinforces perceived predation risk by integrating visual threat cues with predator-associated acoustic signals may induce stronger antipredator responses and broader social learning than single cues, thereby delaying habituation.
The commercialization of UAVs (unmanned aerial vehicles), including drones, has enabled the field deployment of mobile stimuli, motivating research on UAV-based pest bird deterrence 23 . Early UAV approaches often relied on the UAV itself or UAV-generated noise; however, when threat cues are weak, UAVs may become habituated in the same manner as other visual–acoustic deterrents 24 . To address this limitation, attempts have been proposed to simulate predator scenarios by combining visual and acoustic cues on UAV platforms 25 , and studies have evaluated responses to drones equipped with acoustic playback devices or threat cues 26 . Wang et al. (2020) proposed a UAV carrying a bird taxidermy and a loud acoustic signal and quantified short-term outcomes such as the radius of influence, return time after stimulation, and changes in abundance before and after stimulus, demonstrating the potential of combined stimuli 27 . Nevertheless, despite the inevitability of repeated deployment in practical management, the rate at which predator-scenario-based combined stimuli become habituated during repeated applications remains insufficiently evaluated.
This study aims to quantify the behavioral responses of magpie (Pica serica) flock to a non-lethal deterrence method that integrates visual and acoustic predator cues on a drone, and to test both deterrence efficacy and habituation patterns under repeated exposure. We simulated a predator-attack scenario by combining a taxidermy-mounted drone with mobbing calls and recorded changes in abundance and predator related behavior during and after stimulus application. A drone-only stimulus was conducted under the same conditions to isolate the additional effects of the combined stimulus. We derived a recovery from time-series changes in abundance and behavior and inferred habituation by comparing recovery patterns across trials. This study provides an evidence base and analytical framework for quantitatively assessing the persistence of combined-stimulus non-lethal deterrence in urban pest bird management and will inform the design and deployment of stimuli to minimize habituation in long-term operations.
Materials and methods
Taxidermy-mounted drone flight and acoustic signal playback
Considering the seasonal pattern of magpie flock formation in Gwangju, Republic of Korea, we conducted field experiments over a 6-week period from December 2025 to January 2026. The experiments were implemented in two sequential phases according to stimulus condition. The first phase evaluated responses and habituation to the drone itself, without simulating a predator-attack scenario, whereas the second phase quantified responses and habituation to a drone operating under a predator-attack simulation. Each phase lasted 2 weeks, with trials attempted at 2-day intervals; six trials were conducted in the first phase and five trials in the second phase. The number of trials was determined to maximize standardized experimental repetitions within constraints imposed by the winter foraging period and field and equipment operation. Due to site conditions and operational constraints, the stimulus conditions could not be randomized or alternated; therefore, the two treatments were applied sequentially. After completion of the first phase, we conducted a 2-week monitoring period at 2-day intervals during the same time window to minimize carryover effects on the second phase and to assess any residual influence of the first phase before initiating the second phase.
During this monitoring period, abundance and behavioral categories were recorded using the same criteria as in the experimental trials to quantify baseline responses and their natural variability in the absence of stimuli. All trials were conducted between 15:00 and 16:00, consisting of 10 min of stimulus application followed by 50 min of observations of behavior and abundance within the study area. Preliminary observations showed that magpie flocks using the experimental area for foraging left the area between 17:00 and 18:00. This pattern was considered likely to reflect daily movement to roosting sites for resting after sunset 28 . To minimize the potential influence of this daily movement on the experimental results, all trials were conducted between 15:00 and 16:00. In addition, because the aim of this study was to quantitatively evaluate the degree of habituation of magpie flocks to the deterrent strategy, rather than to assess its long-term deterrent performance, the post-stimulus observation period was set to 50 min. For each trial, notable conditions (e.g., weather, raptor presence, and ambient noise) were documented on the datasheet and incorporated into subsequent interpretation of the results.
To simulate a predator-attack scenario, we used a taxidermy-mounted drone and mobbing call playback (Figure 1). To mimic a captured individual and maximize the visual cue, a taxidermized magpie prepared with wings spread was secured to the drone in an inverted, hanging posture (Figure 1(a)). To provide an acoustic cue of predation risk, magpie mobbing call was played throughout the drone flight period. The taxidermy and mobbing call used in this study were provided by the Animal Behavior and Ecology Laboratory, Department of Biological Sciences, Chonnam National University. The UAV and speaker were a DJI MAVIC 3 CLASSIC (DJI, China) and a PAS 800 (E&W, China). To securely mount the taxidermy on the drone and enable remote release, we used a release device (Release Device for MAVIC 3, ALIENTECH, China). To ensure stable flight and takeoff/landing while minimizing damage to the taxidermy, we added a wire frame to the taxidermy and connected it to the release device for use in the experiments (Figure 1(b) and (c)). Magpie taxidermy specimen and a drone equipped with the magpie specimen. (a) Magpie specimen and drone (scale bars: 10 cm). (b) Mounting frame for the magpie specimen and a remote-release device. (c) Schematic of the mounting frame and the remote-release device.
Study site and drone flight trajectory
The experiments were conducted at a rice paddy field on the campus of Chonnam National University, Gwangju, Republic of Korea (total area: 24,328.2 m2; latitude: 35.174° N, longitude: 126.898° E), where human access is limited and magpies forage during winter (Figure 2(a)). Preliminary observations confirmed that from November until the onset of the breeding season, flocks ranging from 10 to 70 individuals repeatedly formed and foraged between 14:00 and sunset. Based on these observations, the site was designated as an experimental area. The study area consists primarily of rice paddies and fields, with utility poles and trees along the perimeter that provide cover and vantage points for vigilance. Each trial was initiated only when ≥10 individuals were confirmed to be foraging as a flock within the experimental area. Study area and distribution of experimental structures. (a) Satellite image of the study area. Red dots indicate the locations of the observer and the recorder; yellow dots indicate the speaker’s location. The red dashed line denotes the boundary of the study area. (b) Schematic diagram of the study area and structure layout. The black dashed line indicates the drone flight path.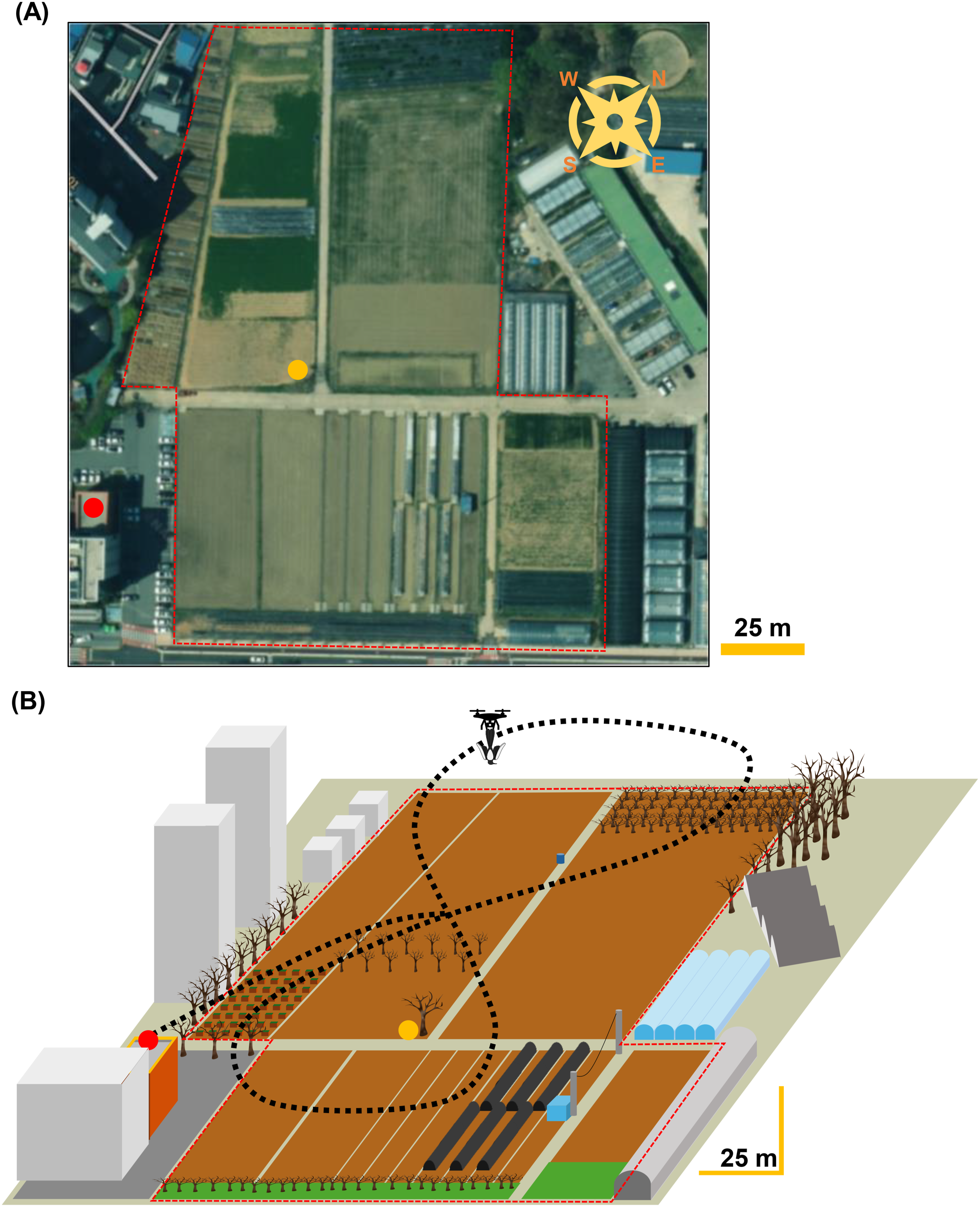
Observers and acoustic recordings were positioned on the rooftop of a nearby building adjacent to the rice paddy field, and the drone operator as well as the takeoff and landing point were set at the same location (Figure 2(b)). Drone flights began at 15:00 and continued for 10 min, following a figure-eight trajectory centered on a Zelkova tree located near the middle of the experimental area (altitude: 13∼15 m; speed: 8∼10 km/h). The speaker for mobbing call playback was placed beneath the central Zelkova tree and oriented toward the northern section of the field, where magpies primarily conducted foraging activities.
Quantification of habituation in magpies
Operational ethogram used to classify field-observed behavioral responses of magpies during drone-based stimulus trials.

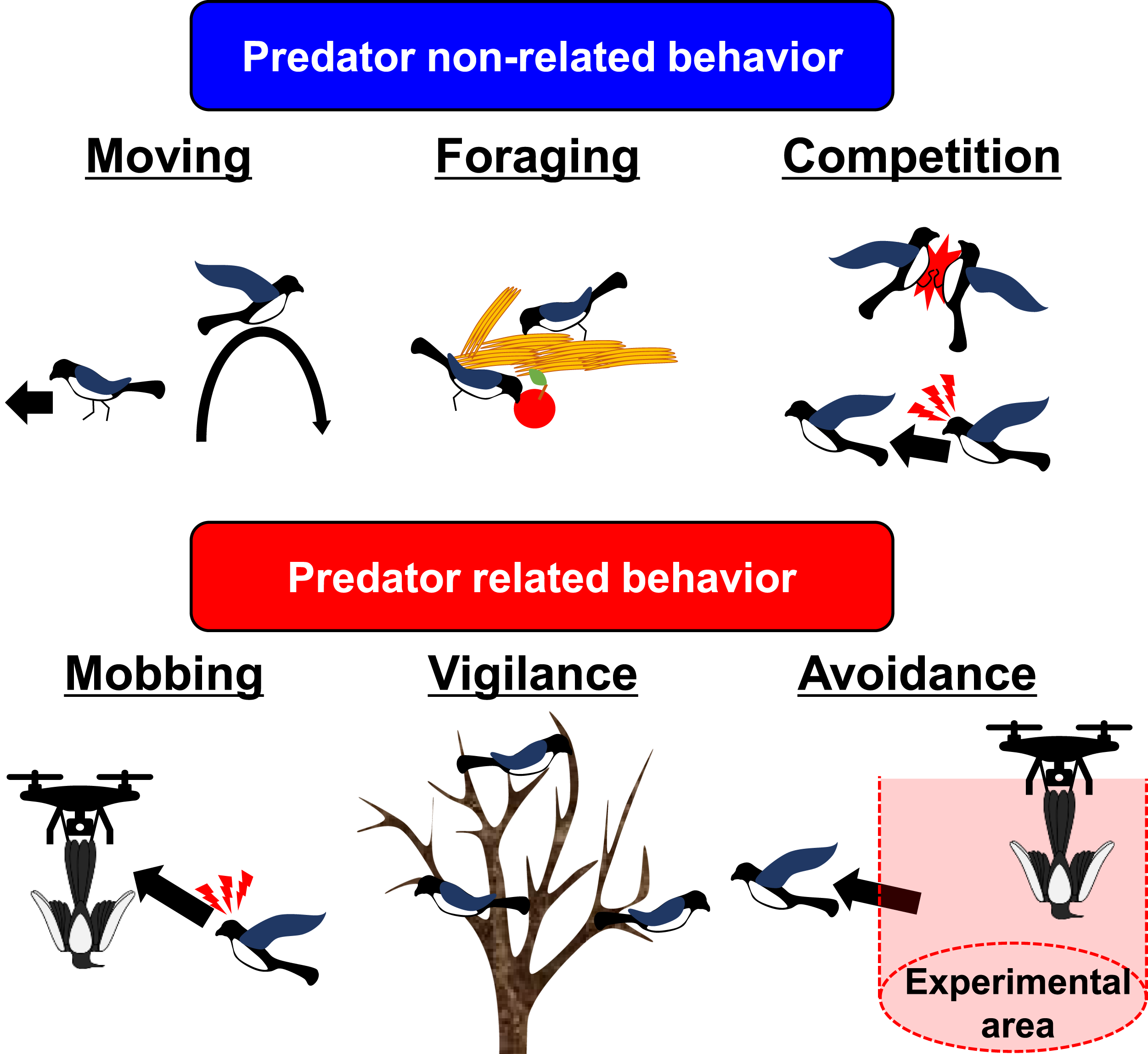
Schematic overview of the behavioral classification criteria used during field observations.
To reduce observer bias, one primary observer and two assistant observers conducted observations together from the same observation point, and behavioral classifications were recorded after cross-checking among observers according to the predefined criteria. At each observation point, the primary observer recorded the number of individuals and their behavioral categories, while the assistant observers simultaneously confirmed the position and movement direction of the same flock, whether individuals remained within the experimental area, and the apparent relationship between their behavior and the stimulus. When individual counts or behavioral classifications were unclear, the primary and assistant observers immediately cross-checked the situation at that observation point. Only the final consensus records, determined according to the predefined ethogram criteria, were used for subsequent analysis.
Mobbing responses were assessed using acoustic recordings obtained with a recorder (PMD661MKIII, Marantz Professional, China) and were quantified and interpreted based on mobbing call duration, note counts, and recording timestamps. Note counts were obtained using Raven Pro software, and each note was manually verified and counted by the analyst through direct auditory inspection of the recording. The primary outcome was recovery, defined as the return to no response after stimulus removal within each trial. Secondary outcomes were the number of individuals in each behavioral category (predator related behavior, predator non-related behavior), total number of individuals, and mobbing call metrics (notes/min and duration). The behavioral checklist and waveform information for recorded mobbing calls are provided in Figures S1∼S3. To distinguish playback mobbing calls from live magpie vocalizations in the combined-stimulus treatment, the spectrograms of the audio recordings were examined. The playback calls consisted of a short audio file that was repeatedly broadcast, and because the distance between the speaker and the recording device remained constant, they showed regular repeated intervals and similar waveform structures. Based on these characteristics, signals showing the same repeated interval and spectrogram pattern as the playback file were identified as playback calls and excluded from the analysis. Cases in which live vocalizations overlapped with playback calls and could not be clearly distinguished were also excluded from the analysis. Spectrogram information used to distinguish live mobbing calls from mobbing-call playback is presented in Figure S4.
Habituation to the taxidermy-mounted drone was quantified by normalizing the number of individuals showing predator non-related behavior within the experimental area to the total number of individuals showing predator non-related behavior at the start of each trial, expressed as a percentage (Figure 4). In addition, mobbing call duration was partitioned into three components—during stimulus, after stimulus, and total duration—to quantify the persistence of mobbing responses. Based on these measures, recovery was quantified. Recovery represents the duration for which responses persist after stimulus removal, and recovery is known to become faster as habituation progresses
29
. In this study, stimulus-induced responses were operationally defined as area departure, predator related behavior, and recovery as the rate of return to predator non-related behavior. Because individuals initially exhibiting predator non-related behavior constituted the primary exposure group for habituation assessment, normalization was based on the initial number of predator non-related behavior individuals. Recovery was calculated from the slope toward the 100% return point. If 100% was not reached by the end of observation, recovery was calculated using the maximum value attained; if no individuals showing predator non-related behavior were observed, recovery was recorded as 0. When 100% was reached during drone stimulus, recovery for that trial was not calculable and was indicated separately. Workflow for quantifying habituation and recovery responses of magpies to the drone. (a) Overview of the experimental design and schematic classification of magpie behaviors. (b) Procedure for population normalization and recovery calculation based on the experimental schematic.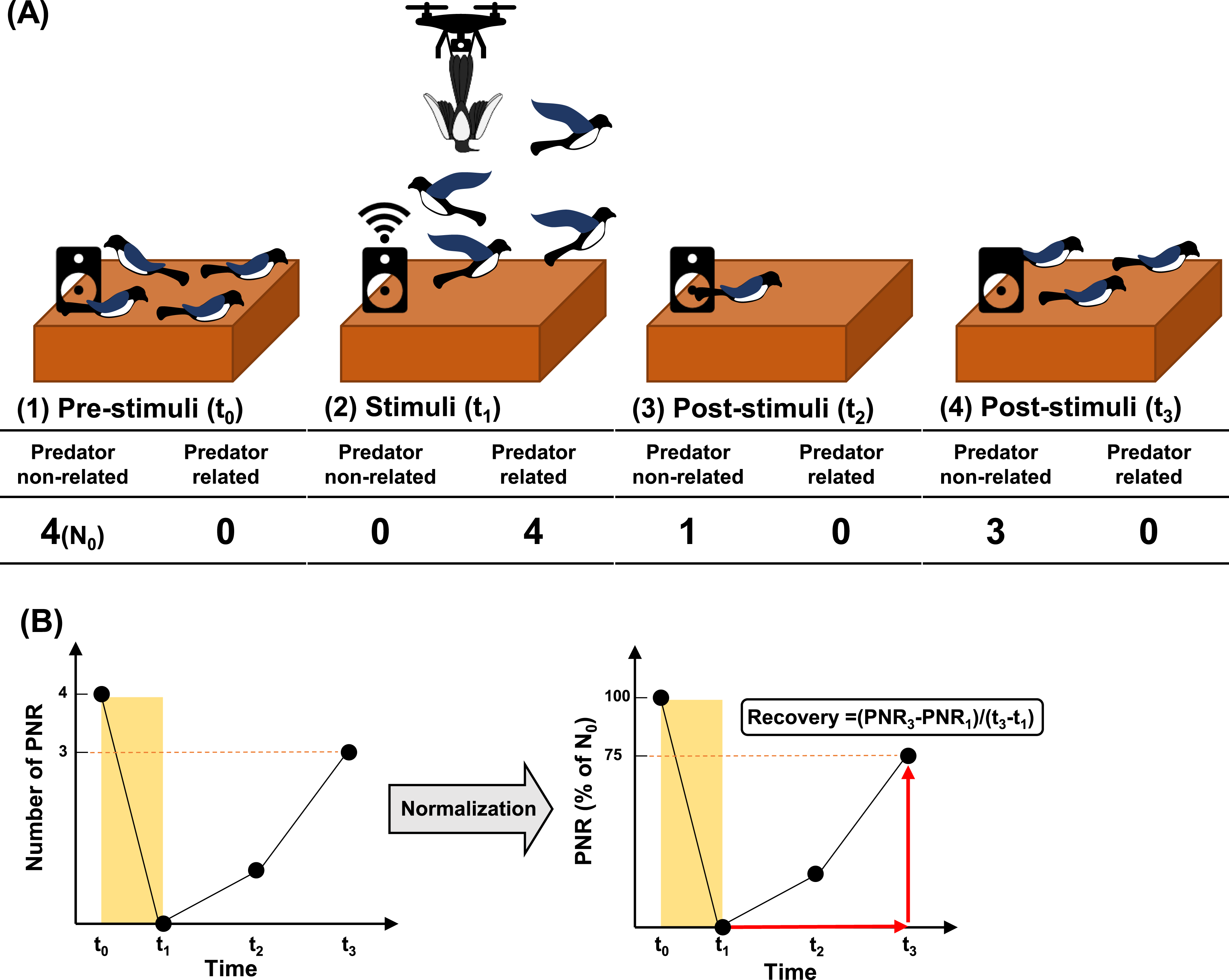
Each experiment was conducted on a single magpie flock, and the observational data consisted of behavioral-category counts recorded at 5-min intervals. Accordingly, individual 5-min intervals were not treated as independent samples; instead, recovery was calculated and compared at the trial level. The unit to which the treatment was applied was a single flock, and the unit for recovery calculation and analysis was each trial. Because this study aimed to quantify repeated-exposure responses within a single flock, generalization to the population level is limited. Observer blinding was not feasible because drone flight and acoustic playback were clearly perceptible in the field. As this study focused on quantifying repeated-exposure responses in a single flock, hypothesis-testing statistics for population inference were not applied. Instead, recovery and behavioral metrics were compared descriptively and visualized using each trial as the analytical unit.
Results and discussion
Behavioral and acoustic responses of magpie flock to stimuli
Under the drone-only stimulus, predator related behavior increased markedly during stimulus but declined across repeated trials, and the return of individuals showing predator non-related behavior accelerated after stimulus termination (Figure 5(a)). During trial 1, no individuals returned to predator non-related behavior during stimulus; however, from trial 2 onward, 21 individuals exhibited predator non-related behavior during the 10-minute stimulus period. Most individuals did not leave the experimental area but instead performed topping as part of predator related behavior, alternating between predator non-related behavior and topping depending on drone proximity. After stimulus termination, the number of individuals showing predator non-related behavior increased rapidly in all trials, whereas predator related behavior declined correspondingly. In trial 1, individuals resumed predator non-related behavior immediately after stimulus cessation, and within 5 minutes, 33 of 34 individuals returned to predator non-related behavior. Except for trial 1, increases in predator non-related behavior were observed during stimulus, and in trials 2 and 3, all individuals returned to predator non-related behavior during stimulus relative to the initial abundance. Raptor attacks were observed in trials 2 and 4; around these events, topping individuals increased and total abundance decreased slightly. The attack in trial 2 occurred at the end of the experiment, preventing further observation; in trial 4, however, a rapid increase in predator non-related behavior was observed 15 minutes after the attack. Although raptor attacks during the trials could have influenced magpie behavior, these events were included in the analysis because the number of individuals showing non-predator-related behavior used for recovery calculation had already reached 100% of the initial flock size at least 20 min before the attacks occurred. Therefore, we considered that these events were unlikely to have substantially affected the recovery data. Changes in magpie behavior, abundance, and mobbing calls in response to drone deployment. (a) Drone-only stimulus. Purple lines indicate the onset of raptor intrusion and attack on a pigeon flock spatially separated from the magpie flock within the experimental area. (b) Combined stimulus using a taxidermy-mounted drone with mobbing call playback.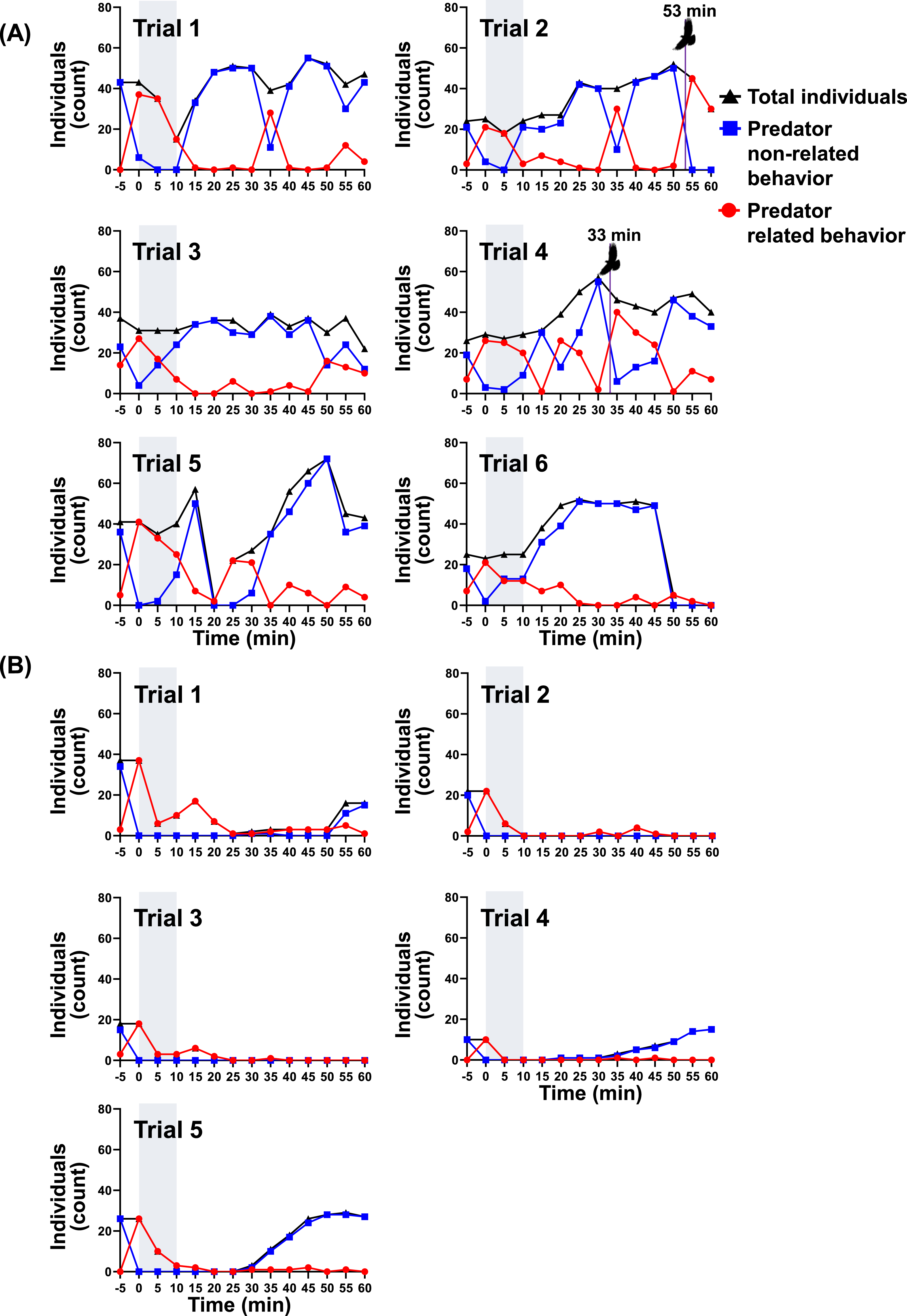
Under the taxidermy-mounted drone and mobbing call combined stimulus, most individuals immediately departed the experimental area, resulting in a substantial decline in total abundance (Figure 5(b)). No individuals exhibited predator non-related behavior during stimulus in any trial; except for 3∼10 individuals displaying topping behavior, all birds left the site. In trial 1, predator non-related behavior increased to 15 individuals 5 minutes before the end of observation. In contrast, in trials 2 and 3, no individuals were observed in the area until the experiment concluded. However, in trials 4 and 5, the first individuals showing predator non-related behavior reappeared 25 and 20 minutes after stimulus termination, respectively, and gradually increased to 15 and 27 individuals.
Comparison of mobbing call notes count per minute with the normalized predator non-related behavior index revealed stimulus-dependent patterns (Figure 6(a)). Under the drone-only stimulus, predator non-related behavior increased rapidly after stimulus termination, reaching 100% within 5∼10 minutes; notably, in trials 2 and 3, 100% was reached during stimulus. In all trials, values exceeded the initial baseline repeatedly, reaching up to 278%. Although irregular declines occurred after stimulus termination and increases in predator non-related behavior, recovery was generally rapid. When a raptor captured a pigeon, predator non-related behavior decreased to 0%; when the raptor merely pursued, it decreased to 31.5%. 15 minutes later, predator non-related behavior recovered to 242%, comparable to pre-attack levels. Mobbing call note counts remained generally low without a consistent pattern; however, in trial 2, note counts increased sharply to 60 notes/min immediately after a raptor attack on a pigeon. Normalized non-response and mobbing call following drone deployment. (a) Drone-only stimulus. Purple lines indicate the onset of raptor intrusion and attack on a pigeon flock spatially separated from the magpie flock within the experimental area. (b) Combined stimulus using a taxidermy-mounted drone with mobbing call playback.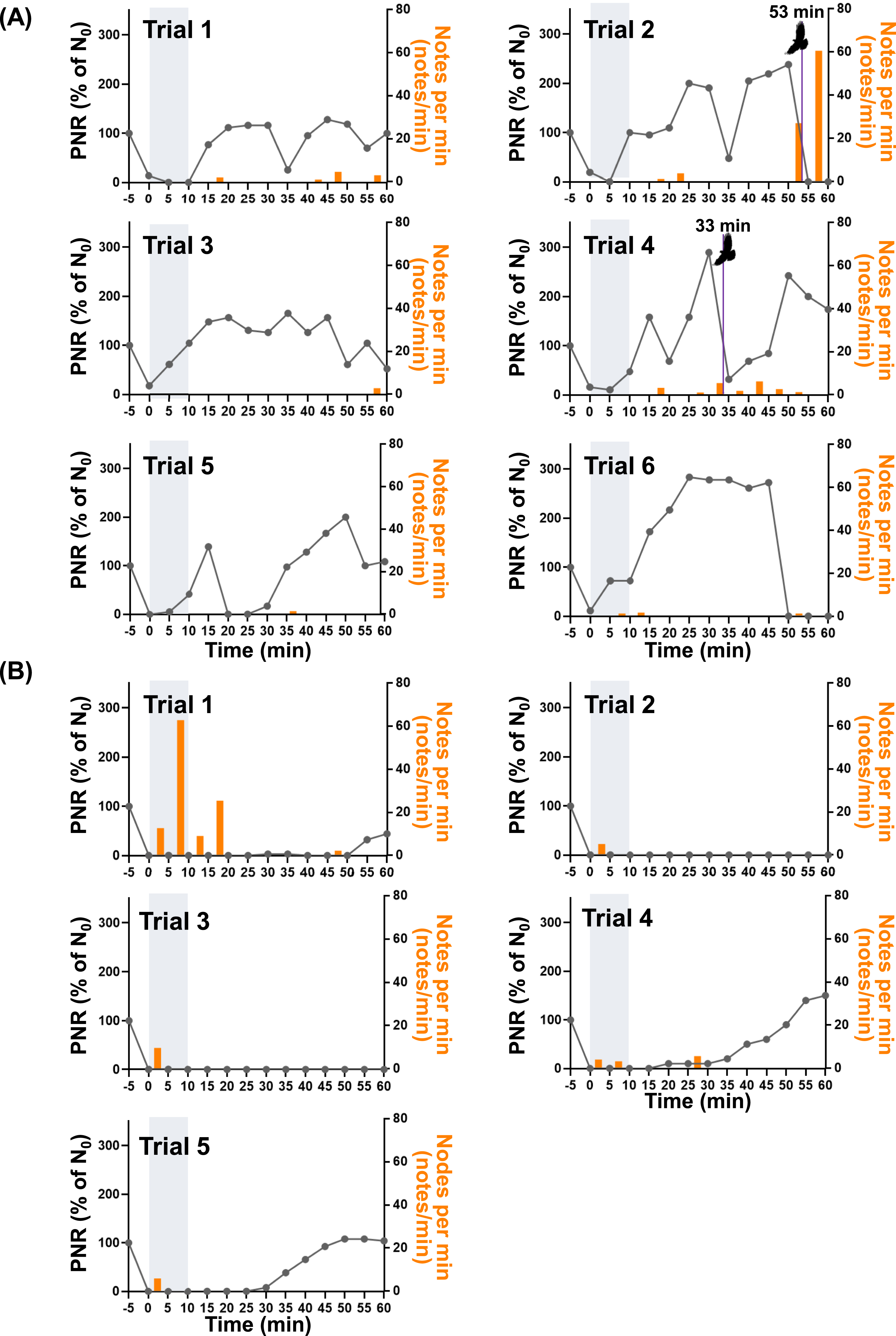
Under the combined stimulus, predator non-related behavior remained at 0 during stimulus application in all trials and persisted until experiment termination in trials 2 and 3 (Figure 6(b)). In trial 1, predator non-related behavior increased to 32.4% at 35 minutes after stimulus onset and to 44.1% by the end of observation. In trial 4, predator non-related behavior increased to 10% at 20 minutes after stimulus termination and gradually rose to 150% by the end of observation. In trial 5, predator non-related behavior increased to 7.7% at 25 minutes after stimulus termination and gradually reached 103.8%. Note analysis showed that in trial 1, mobbing call rates peaked at 62.8 notes/min during and up to 10 minutes after stimulus, but such high rates were not observed subsequently. In all trials except trial 1, note rates during stimulus application were low (maximum 9.6 notes/min), and after stimulus termination, note counts were either absent or remained low until the end of observation.
Changes in recovery and mobbing call duration across repeated trials
Recovery after stimulus termination and mobbing call duration showed distinct patterns depending on stimulus condition and trial (Figure 7). In trials 2 and 3 under the drone-only stimulus, predator non-related behavior reached 100% before stimulus termination; therefore, recovery could not be calculated. Comparison among trials 1, 4, 5, and 6 showed that recovery in trial 1 (12.0 %/min) was lower than in trials 4, 5, and 6 (21.9, 19.4, and 20 %/min, respectively) (Figure 7(a)). In addition, mobbing calls were rarely detected during stimulus in all trials. Extended mobbing call durations were observed only in trials 2 and 4, when raptor attacks occurred, with durations of 16.3 and 22.3 seconds after stimulus termination. Recovery of magpies and duration of the mobbing call. (a) Recovery and duration of mobbing call across trials. The left panel shows results for the drone-only stimulus, and the right panel shows results for the combined stimulus using a taxidermy-mounted drone with mobbing call playback. (b) In-stimulus recovery during drone-only stimulus.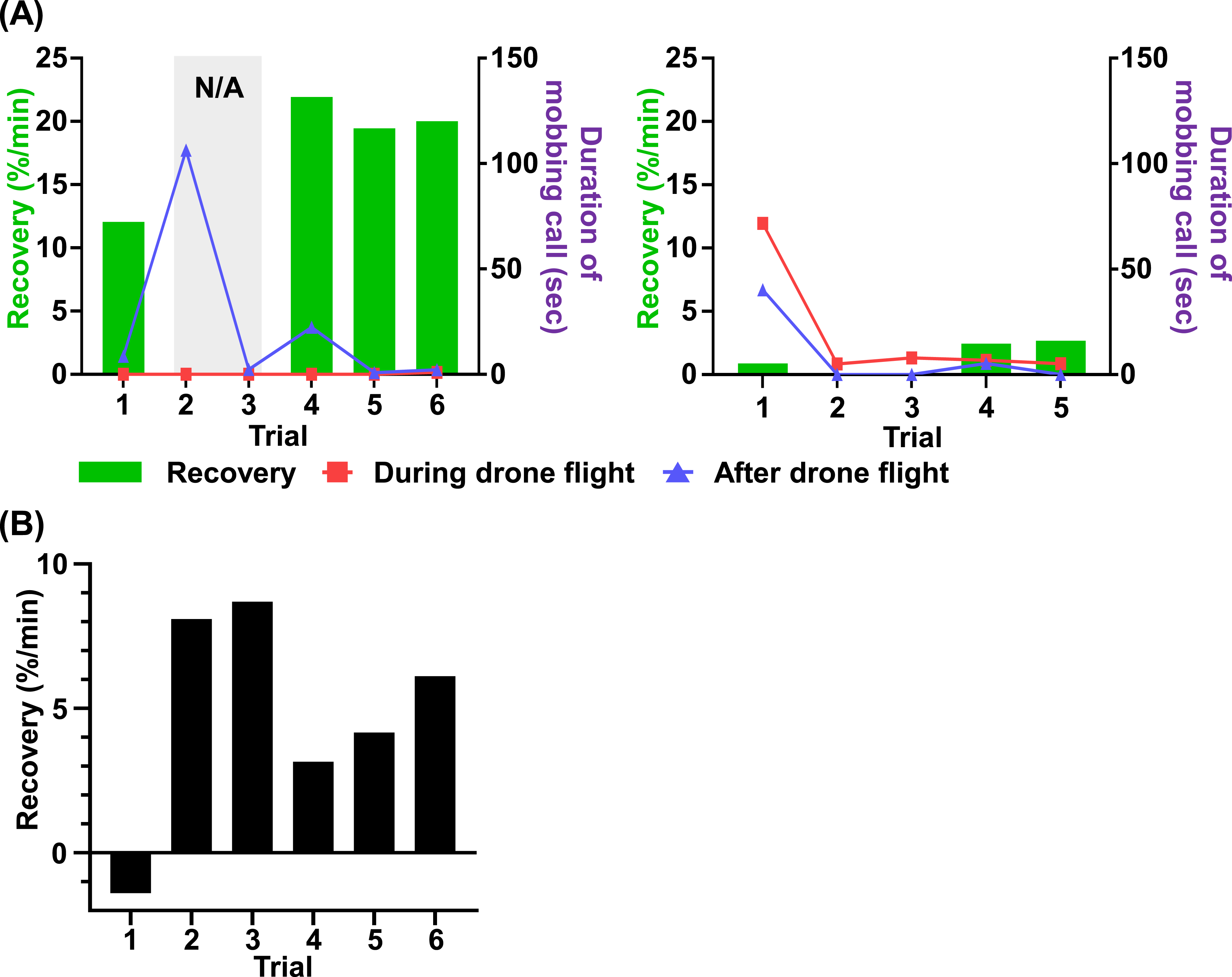
Under the taxidermy-mounted drone and mobbing call combined stimulus, recovery was not detected in trials 2 and 3. In trials 4 and 5, recovery increased to 2.4 and 2.6 %/min (Figure 7(a)), but these values remained substantially lower than those under the drone-only stimulus. In trial 1, mobbing calls were detected both during and after stimulus, with longer durations during stimulus (71.7 s). In contrast, except for a brief 5-second period during stimulus in trial 2, mobbing calls were not detected throughout the remainder of the observation period.
To complement recovery assessment under the drone-only stimulus, recovery during stimulus was additionally quantified (Figure 7(b)). In trial 1, recovery during stimulus showed a negative value (−1.4 %/min). In trials 2 and 3, where 100% predator non-related behavior was reached during stimulus, recovery values were markedly high (8.1 and 8.7 %/min). While no consistent trend in post-stimulus recovery was observed during the latter trials (4, 5, and 6), recovery rates during the stimulus application exhibited a progressive increase with each repetition (3.2 to 4.2 to 6.1 %/min).
Assessment of habituation across repeated stimulus exposure
Under the drone-only stimulus, all trials showed a consistent pattern of increased predator related behavior and decreased predator non-related behavior immediately after stimulus initiation; however, from trial 2 onward, individuals showing predator non-related behavior reappeared even during stimulus, and in trials 2 and 3, predator non-related behavior reached 100% during stimulus. These results suggest that the drone-only stimulus was not perceived as a meaningful predation threat by foraging magpie flock and that habituation progressed rapidly to the extent that foraging activity was maintained even while the stimulus was ongoing. Previous findings showing that drones could approach birds to within 4 m also support the rapid habituation and recovery observed in magpies 23 .
In contrast, under the combined stimulus using a taxidermy-mounted drone and mobbing call playback, predator non-related behavior remained at 0 during stimulus application and flock departed the area; in trials 2 and 3, the site remained unoccupied until the end of observation. This pattern indicates that the combined stimulus increased perceived predation risk relative to the drone-only stimulus and, consequently, delayed habituation. Nevertheless, the re-entry of individuals showing predator non-related behavior and their sustained increase 20∼25 minutes after stimulus termination in trials 4 and 5 can be interpreted as emerging signals of habituation even under the combined condition. In this study, the number of repeated exposures was limited because flocking ceased with the onset of the breeding season; however, with further repeated exposure, faster recovery driven by habituation is likely.
Differences in behavioral responses to the drone-only stimulus versus the taxidermy-mounted drone and mobbing call combined stimulus
Marked differences were observed in flock behavior during stimulation and after stimulus removal between the drone-only stimulus and the combined stimulus. Under the drone-only stimulus, the initial approach elicited predator related behavior; however, most individuals primarily shifted to nearby trees and engaged in topping (vigilance/observation). With repetition, this response weakened, accompanied by an increase in individuals showing predator non-related behavior during stimulus. In contrast, under the combined stimulus (taxidermy-mounted drone and mobbing call), all trials elicited strong predator related behavior characterized by departure from the experimental area, resulting in a sharp reduction in total individuals, and in some cases complete abandonment of the area. Taken together, these differences in behavioral responses and recovery suggest that constructing a predator-attack context using a conspecific carcass-associated drone and mobbing call playback can elevate a previously non-threatening stimulus to one perceived as comparable to a predator threat. This finding further indicates that even without specialized equipment, conventional devices may be leveraged for deterring cognitively advanced birds if an appropriate threat context is established.
Interpreting whether the enhanced effect of the combined stimulus reflects additive contributions of visual and acoustic cues, or a synergistic interaction that amplifies threat perception, is central to designing long-term deterrence strategies. Previous studies suggest that combined cues can strengthen risk assessment relative to single cues, and that integrating visual and acoustic stimuli may prolong or reinforce avoidance responses, thereby maintaining deterrence under repeated exposure 30 . Social risk signals such as alarm calls can also shift receivers’ attention allocation and predator-search behavior, increasing the perceived threat value of the same visual cue; thus, combined stimuli may generate stronger threat perception than single stimuli and potentially delay habituation 31 . However, combined stimuli do not always evoke stronger responses than single cues. Depending on the relative informational value of each cue, combined presentation may elicit responses similar to, or even weaker than, those induced by a single cue 32 . Therefore, when aiming to achieve amplified effects, the types and combinations of stimuli should be carefully designed with local field conditions in mind.
Comparison with behavioral responses to natural raptor attacks and approaches
Three raptor-related events observed during the study period (one pigeon predation event, one pigeon chase event, and one approach without hunting behavior) were characterized by pronounced topping-centered predator related behavior without area abandonment by the magpie flock. In the non-hunting approach event, predator non-related behavior recovered rapidly within approximately 5∼10 minutes. Even when raptors captured or chased pigeons, the target was not a conspecific, the direction of raptor attention and attack was relatively clear, and the flock may have judged that information acquisition and risk management via topping were feasible; under such conditions, vigilance through topping may have been more advantageous than leaving the area. Reports that magpies adjust their responses depending on predator context support this context-dependent risk assessment 18 .
In contrast, the taxidermy-mounted drone and mobbing call combined stimulus consistently induced area abandonment and low recovery compared with natural raptor events. This difference suggests that the flock may have evaluated the combined stimulus as a threat that could not be resolved quickly, even with mobbing responses. Mobbing is a high-cost behavior involving energetic and time expenditure and increased risk exposure; as threat persistence increases, abandoning the area may become more advantageous than continued responding 33 . Under the combined stimulus, both (i) mobbing call playback originating from an unknown group and (ii) the perceived threat within the area (the taxidermy-mounted drone) were not removed by the flock’s immediate response. Because this threat was repeated in trials 2 and 3, the flock may have assessed that the costs of sustained mobbing outweighed the benefits and therefore selected area abandonment. Consistent with this interpretation, corvids have been reported to learn high-risk foraging sites and subsequently reduce access to those sites 32 .
The recovery patterns observed in trials 4 and 5 may be explained by seasonal and ecological constraints: during winter, the value of food resources is high, and competitive pressure may increase as magpies share resources within the same area with azure-winged magpies and large-billed crows, making prolonged avoidance difficult. In trial 4, when large-billed crows approached the area and the likelihood of losing access to resources increased, heightened foraging pressure may have promoted area return and resumption of foraging. Under these resource pressures, habituation driven by repeated exposure may also have progressed, leading to observable recovery even under the combined stimulus.
Habituation-mitigation strategies for long-term behavioral control
Our results indicate that the combined stimulus elicited stronger predator related behavior (i.e., greater deterrence) than the drone-only stimulus, but the increasing recovery across repeated exposures suggests a potential decline in deterrence effectiveness due to habituation. To mitigate this limitation and achieve long-term behavioral control, we propose the following strategies.
First, designs that go beyond simple approach and repeated flight paths may be more effective, including simulating predator-like pursuit, rapid approaches, and abrupt directional changes that resemble hunting behavior. Drone morphology may also matter. For example, predator-like shapes could further suppress habituation and maximize deterrence. A robotic predator mimicking raptor flight (Robot Falcon) reportedly produced stronger deterrence than conventional drones and existing measures, with limited evidence of habituation during the deployment period 34 .
Second, because repetition of identical stimuli accelerates habituation in long-term operations, deliberate variation across time, space, and form is required. Specifically, the interval between trials, stimulus duration, and onset timing should be randomized; drone approach direction, altitude, speed, and flight trajectories should be varied across trials to reduce predictability 13 . For acoustic stimuli, rotating calls from different individuals and varying playback structure—rather than repeatedly using the same file—may reduce repetitiveness and mitigate habituation 31 . Finally, alternating and mixing multiple deterrence tools (e.g., drones, visual cues, and acoustic cues), rather than relying on a single device, may delay learning of any one stimulus and improve long-term effectiveness.
Need for future evaluation across multiple magpie flocks and long-term field validation
This study quantified deterrence effects and habituation dynamics by repeatedly applying predator-related stimuli to a single magpie flock; however, the use of a single flock limits the ability to reflect regional variation. Moreover, because the study targeted one local population, it is not possible to claim generalized species-level responses. Future work should conduct experiments across multiple populations and regions and interpret results using statistical analyses to obtain more robust and generalizable evidence. In addition, because the study involved free-ranging flocks that naturally aggregated in the field, it cannot be confirmed that the same individuals participated across trials, and confounding factors such as raptor presence, interspecific competition among corvids, and abiotic conditions (e.g., weather) could not be fully controlled. Individual identification via capture or marking would help address the first limitation. Conducting experiments in a controlled environment with sufficient space would also enable stronger causal inference by reducing environmental confounding.
Because the present study evaluated short-term behavioral responses and recovery patterns during the 50-min post-stimulus observation period, it cannot determine whether the observed avoidance responses would lead to actual crop damage reduction or sustained deterrent effectiveness. Future studies should therefore be conducted in orchards or other agricultural fields where magpie damage actually occurs, and should include quantitative measurements of fruit damage, such as the proportion of damaged or missing fruits based on field sampling of orchard blocks, as well as longer-term monitoring over several hours after stimulus application 35 . Such field-based assessments would be necessary to determine whether the combined stimulus can provide practical and sustained protection against magpie damage under real agricultural conditions.
Conclusion
This study quantitatively evaluated habituation dynamics in a single flock of magpies foraging during winter by repeatedly applying a drone-only stimulus and a combined stimulus consisting of a taxidermy-mounted drone and mobbing call playback, and by comparing behavioral responses and recovery. Under the drone-only stimulus, predator related behavior was elicited, but individuals rapidly returned to predator non-related behavior after stimulus removal, resulting in high recovery. Habituation also progressed quickly, as indicated by an increasing number of individuals showing predator non-related behavior even during stimulus with repeated trials. In contrast, the combined stimulus produced a strong behavioral control effect, with complete area abandonment and no return to predator non-related behavior. However, recovery increased across repeated trials, indicating a decline in control efficacy due to habituation. These findings suggest that long-term deployment of UAV-based deterrence incorporating predator cues will require additional habituation-mitigation strategies, such as varying stimulus composition. By establishing quantitative criteria for long-term evaluation of visual and acoustic deterrence and by quantifying both efficacy and habituation potential of cognition- and behavior-based pest bird management, this study provides evidence for practical applicability and identifies directions for improving long-term implementation.
Supplemental material
Supplemental material - Habituation of magpies (Pica serica) to taxidermy-mounted drone and mobbing call for flock control
Supplemental material for Habituation of magpies (Pica serica) to taxidermy-mounted drone and mobbing call for flock control by Moon-Su Kim, Jiyoung Yun, Soo-Yeon Lee, Hyunwoo Lim, Seong Young Ko, Ha-Cheol Sung and Eung-Sam Kim in Avian Biology Research
Footnotes
Acknowledgements
We gratefully acknowledge the Agricultural Practice Education Center at Chonnam National University for their permission and support in using the experimental paddy fields.
Ethical considerations
This study consisted of non-invasive field observations and deterrent stimuli without capture, marking, or sampling of wild birds and was conducted in accordance with relevant guidelines and regulations. The study protocol was reviewed and approved by the Chonnam National University Institutional Animal Care and Use Committee (approval no: CNU IACUC-YB-2025-87).
Funding
This work was supported by the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT (grant No. RS-2026-25478676). This research was also supported by Global-Learning & Academic research institution for Master, PhD students, and Postdocs (LAMP) Program of the NRF funded by the Ministry of Education (grant no.: RS-2024-00442775) and by the Regional Innovation System & Education (RISE) through the Gwangju RISE Center, funded by the Ministry of Education and the Gwangju Metropolitan Government, Republic of Korea (grant no.: 2026-RISE-05-011).
Declaration of conflicting interests
The authors declare no conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data associated with this research are provided in the manuscript, the supplementary information.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.