
Abstract
Hematological parameters are widely used as integrative indicators of physiological condition in birds. The Cabanis’s Ground Sparrow (Melozone cabanisi), a Costa Rican endemic species commonly associated with coffee plantations and semi-modified landscapes, lacks baseline hematological information, limiting the interpretation of blood-based health assessments. In this study, we established descriptive hematologic intervals for M. cabanisi and evaluated the influence of intrinsic biological factors—pectoral muscle condition and molt stage—on hematological variation. Free-ranging individuals were captured using mist nets and processed following standardized protocols. We quantified erythrocyte, leukocyte, and thrombocyte parameters, plasma protein concentration, and erythrocyte indices. Mean corpuscular hemoglobin concentration (MCHC) was selected as a focal parameter to assess physiological condition. Effects of muscle condition and molt stage on MCHC were evaluated using ART ANOVA and complementary bootstrapped linear models. Hematological values for M. cabanisi were broadly consistent with those reported for other small passerines, indicating conserved erythrocytic and immunological profiles. Pectoral muscle condition emerged as the strongest predictor of MCHC, with individuals in better somatic condition exhibiting significantly higher values. In contrast, molt stage did not show a significant main effect on MCHC in conservative analyses, and no interaction between muscle condition and molt stage was detected. These results provide the first hematological reference intervals for M. cabanisi and highlight body condition as a key intrinsic driver of hematological variation. The reference values presented here offer a robust physiological baseline for future ecological, conservation, and wildlife health studies involving this endemic Neotropical passerine.
Keywords
Introduction
Hematological parameters are widely used as integrative biomarkers of physiological condition in birds, reflecting the combined effects of nutrition, immune status, metabolic demand, and environmental pressures. In passerines, indices such as hemoglobin concentration, erythrocyte counts, and leukocyte profiles provide insight into individual health and allostatic balance, often revealing physiological trade-offs not apparent from behavioral or demographic observations alone1–4.
Despite their broad utility, the interpretation of hematological values in free-ranging birds critically depends on the availability of species-specific reference intervals. Without appropriate baselines, it is difficult to distinguish normal physiological variation from deviations associated with stress, poor body condition, disease, or environmental disturbance5,6. This limitation is especially pronounced for Neotropical passerines, which remain substantially underrepresented in hematological and conservation physiology literature compared with temperate taxa 7 .
Body condition is one of the most important intrinsic factors influencing hematological variation in birds8,9. Metrics such as pectoral muscle development and fat reserves reflect recent energetic balance and are closely linked to erythropoietic activity and oxygen-carrying capacity 6, 10. Birds in superior somatic condition frequently exhibit higher hemoglobin-related indices, whereas individuals in poorer condition may show hemodilution or reduced erythrocyte parameters, particularly under nutritional or environmental constraints. These relationships have been reported across a range of passerine species inhabiting both natural and anthropogenically altered environments2,10.
Environmental context, particularly urbanization and habitat modification, can further modulate hematological and immune profiles. Studies have shown that exposure to urban stressors, pollution, and altered resource availability can influence leukocyte distributions, oxidative stress markers, and erythrocyte indices in free-living birds10–12. However, the magnitude and direction of these effects often depend on species-specific ecology and individual condition, reinforcing the importance of robust baseline reference values.
Molt represents another biologically relevant process with potential effects on hematology. Feather replacement is energetically costly and involves substantial protein reallocation, which may influence erythrocyte indices and immune parameters 13 . Nevertheless, empirical evidence remains mixed: some studies report detectable hematological shifts during molt, whereas others suggest that key blood parameters are tightly regulated to maintain physiological stability during this demanding life-history stage14,15.
The Cabanis’s Ground Sparrow is a resident Neotropical passerine endemic to Costa Rica, with a restricted distribution in the Central Valley, Turrialba Valley, and the western Monteverde region, occurring between 500 and 1700 m elevation. The species is primarily associated with early successional habitats, including thickets, shade coffee plantations, and young secondary forest, and is commonly found in semi-modified landscapes. 16 . Although the species frequently inhabits human-altered environments where physiological stressors such as agrochemical exposure and altered resource availability may be present, no hematological reference intervals have been previously established. This gap limits the application of blood-based biomarkers for assessing individual condition and population health in ecological and conservation contexts.
In this study, we (i) establish hematological reference intervals for M. cabanisi and (ii) evaluate the relationships between selected hematological parameters and key biological variables, including pectoral muscle condition and molt stage. By focusing on intrinsic drivers of hematological variation, this work provides a physiological baseline for future ecological, health, and conservation studies involving Neotropical passerines.
Materials and methods
Field capture and handling
During July–December 2024 and March–April 2025, individuals of M. cabanisi were captured in coffee plantations located in three provinces (Heredia, San José, and Cartago) within the Central Valley of Costa Rica using standard 12-m mist nets with a 36-mm mesh size. Sampling was conducted across three independent coffee plantation landscapes spanning the species’ known distribution in the Central Valley to better represent the geographic and habitat variability within the study population. The geographic location of the study sites is shown in Figure 1. To maximize capture efficiency while minimizing disturbance, mist nets were operated exclusively during the early morning hours (approximately 06:00–10:00 h), coinciding with the species’ peak foraging activity. Geographic location of the study sites where Melozone cabanisi individuals were captured in coffee plantations of Costa Rica. The upper right panel shows the national context with provincial administrative boundaries. The remaining panels depict high-resolution satellite imagery of the three study sites, delineated by colored polygons according to province: Heredia (orange), San José (blue), and Cartago (red). Capture sites were located within semi-modified landscapes dominated by shade coffee plantations and associated vegetation. All maps are shown in the CRTM05 projection; background imagery corresponds to Bing Satellite.
To ensure the safety and welfare of the birds, a continuous monitoring protocol was implemented, with the nets inspected at intervals not exceeding 20 minutes. Extraction and handling of the birds was conducted solely by trained personnel experienced in the manipulation of wild birds, in accordance with the Guidelines to the Use of Wild Birds in Research. The entire procedure was designed to minimize handling time—estimated at under 10 minutes per bird—and to reduce stress. Following sample collection and data recording, all individuals were fitted with a unique alphanumeric metal leg band and released immediately at the capture site. All fieldwork was conducted under research permits issued by the Sistema Nacional de Áreas de Conservación (SINAC-ACC-PI-RE-042-2024), Costa Rica.
Processing, sampling, and data recording
Immediately after extraction from the mist net, each bird was processed by a multidisciplinary team comprising biologists and a wildlife veterinarian. A set of standardized morphometric data was recorded for all individuals, including body mass (to the nearest 0.1 g using a precision balance), wing length, age, and sex (when determinable), as well as body condition (assessed through subcutaneous fat and pectoral muscle scoring) and molt status. Pectoral muscle condition was assessed by visual examination of the keel by blowing aside the feathers to expose the breast and following a four-point scale (0–3): 0 = keel prominent, muscle clearly depressed below the sternal edge; 1 = keel easily distinguishable, muscle level with the sternum; 2 = keel still discernible, muscle slightly rounded above the sternal edge; 3 = keel difficult to distinguish, muscle fully rounded 18 . Age was determined by assessing molt, which was scored using the Wolfe-Ryder-Pyle (WRP) system, which classifies birds into discrete plumage-based groups according to the number of molt cycles completed since hatching 19 . The geographic coordinates of the capture site, date, and any notable behavioral observations were also documented.
Each bird was manually restrained, and 0.1 to 0.2 ml of blood was collected from the jugular vein using a 25-gauge needle attached to a 1.0 ml syringe. Blood smears were made immediately on clean microscope slides. The remaining blood was placed into 1 ml VACUETTE® Heparin tubes and transported to the laboratory for same-day processing.
Hematological analysis
Hematological analyses were performed following standard procedures for avian hematology 20 . Total erythrocyte (TRBC), leukocyte (TWBC), and thrombocyte counts were determined manually using Natt and Herrick’s solution (1:200 dilution), prepared by a certified analytical chemistry laboratory (Quimflox S.A.) according to the original formulation. Cell counts were performed using an improved Neubauer hemocytometer. Erythrocytes were counted in the five erythrocyte counting squares (one central and four corner squares) of the central grid, corresponding to a total counting area of 0.20 mm2 and a counting volume of 0.02 μL. TRBC was calculated as the number of erythrocytes counted × 200/0.02 and expressed as ×109/L. Leukocytes and thrombocytes were counted in the nine large squares of the chamber, and concentrations were calculated using the formula (cells counted × 1.1 × 0.2).
Packed cell volume (PCV) was determined by the microhematocrit method after centrifugation of blood-filled Vitrex® non-heparinized capillary tubes at 15,000 rpm for 5 min. Hemoglobin concentration (Hb) was measured using a HemoCue® Hb 201+ analyzer based on the azide methemoglobin method (570 and 880 nm wavelengths). Mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), and mean corpuscular hemoglobin concentration (MCHC) were calculated using standard hematological equations. Differential leukocyte counts were performed on Wright-stained peripheral blood smears by counting 100 leukocytes under oil immersion (1000×) using a Delphi-X Observer™ microscope (Euromex; 10×25 mm eyepieces). Total plasma protein (TPP) was determined by refractometry using a Kalstein YR05915 ATC clinical refractometer and plasma obtained from the microhematocrit tube.
Statistical methods
Robust distribution hematology intervals (I) and descriptive statistics for hematological parameters in the study population (n = 50).
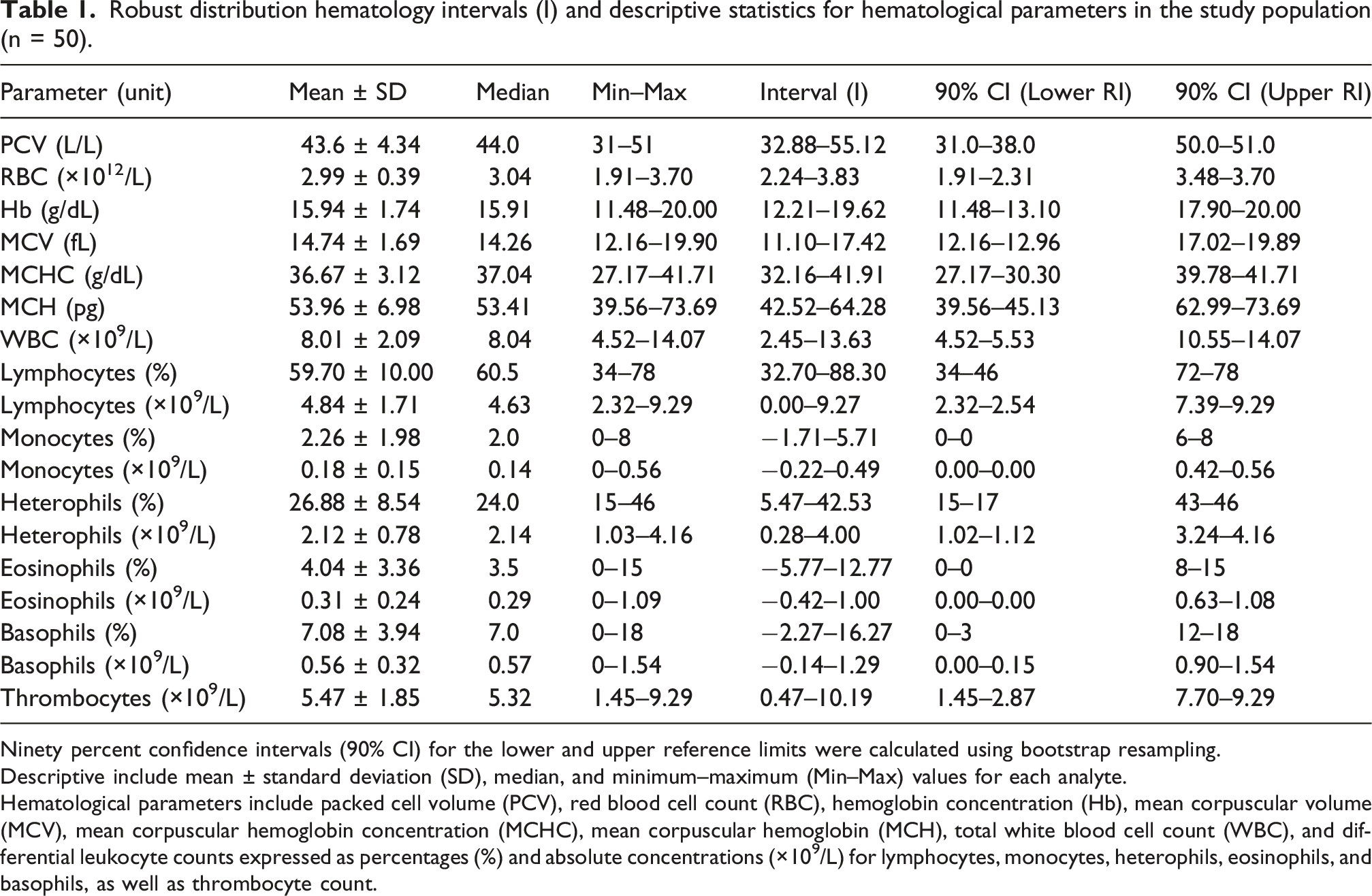
Ninety percent confidence intervals (90% CI) for the lower and upper reference limits were calculated using bootstrap resampling.
Descriptive include mean ± standard deviation (SD), median, and minimum–maximum (Min–Max) values for each analyte.
Hematological parameters include packed cell volume (PCV), red blood cell count (RBC), hemoglobin concentration (Hb), mean corpuscular volume (MCV), mean corpuscular hemoglobin concentration (MCHC), mean corpuscular hemoglobin (MCH), total white blood cell count (WBC), and differential leukocyte counts expressed as percentages (%) and absolute concentrations (×109/L) for lymphocytes, monocytes, heterophils, eosinophils, and basophils, as well as thrombocyte count.
The statistical analysis was performed in three sequential steps to evaluate the relationship between intrinsic biological variables and hematological parameters in Melozone cabanisi. Among all biological and morphometric variables evaluated, only pectoral muscle condition and molt stage were significantly associated with one or more hematological parameters; therefore, only these variables were retained for subsequent inferential analyses.
First, the distribution of each of the 18 hematological variables was assessed using the Shapiro–Wilk test for normality. Because all variables significantly deviated from normality (P < 0.05), and logarithmic transformation did not adequately normalize most distributions, parametric factorial analyses were considered inappropriate.
Second, the effects of pectoral muscle condition and molt stage were evaluated using Aligned Rank Transformation analysis of variance (ART ANOVA), a non-parametric factorial approach that allows testing of main effects and interactions while accommodating non-normal data and unbalanced experimental designs. This method was selected because it preserves the factorial structure of the analysis, unlike separate non-parametric tests applied independently to each predictor.
Results of Aligned Rank Transformation analysis of variance (ART ANOVA) evaluating the effects of pectoral muscle condition, molt stage, and their interaction on 18 hematological parameters in free-ranging Melozone cabanisi. F statistics, degrees of freedom (df), and corresponding P values are presented for each effect. Significant results are indicated by asterisks (P < 0.05, *P < 0.01, **P < 0.001), whereas marginally significant effects are denoted by a dot (P < 0.10). Muscle score was classified into two categories (Score 2 and Score 3), and molt stage followed the Wolfe–Ryder–Pyle (WRP) classification system.
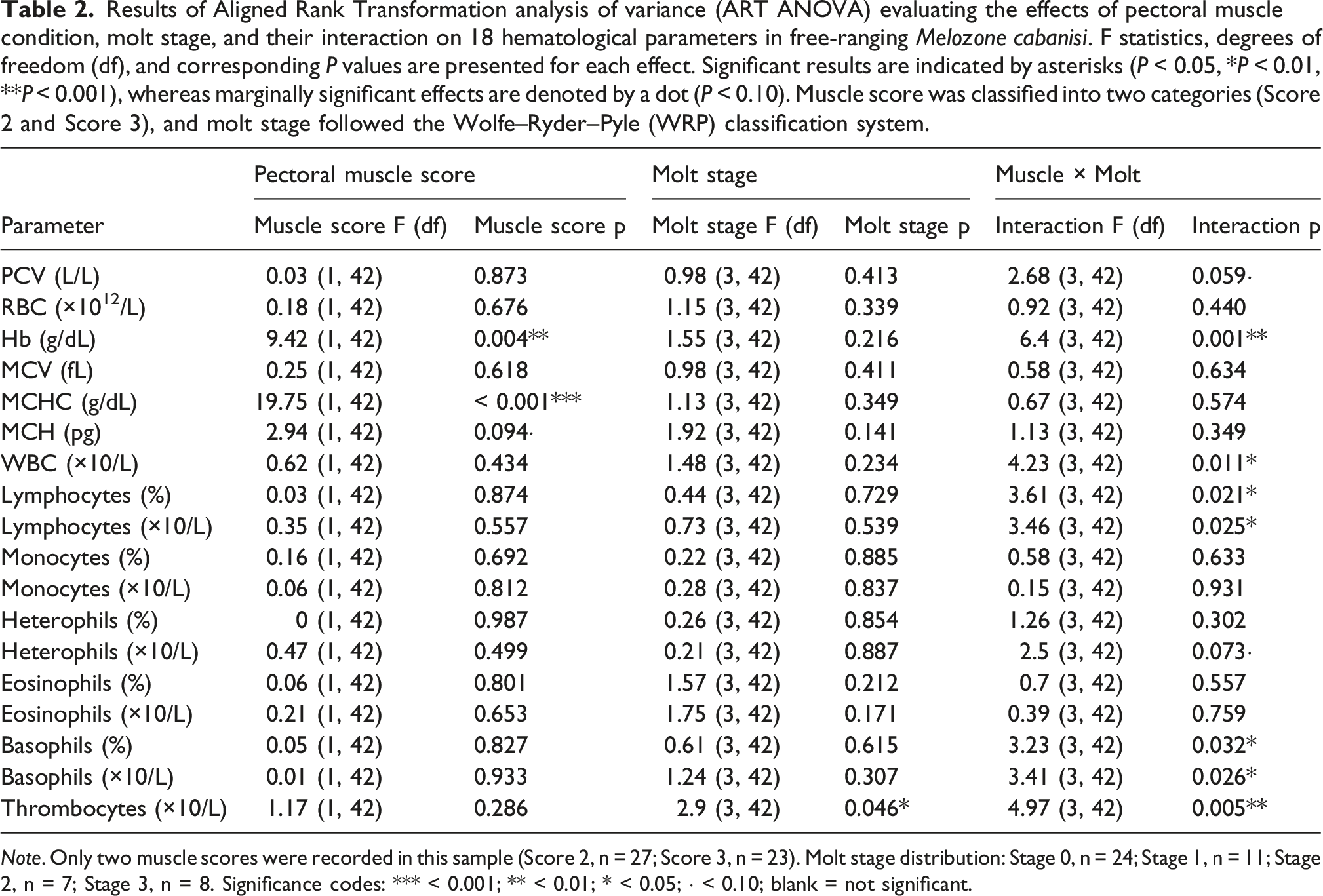
Note. Only two muscle scores were recorded in this sample (Score 2, n = 27; Score 3, n = 23). Molt stage distribution: Stage 0, n = 24; Stage 1, n = 11; Stage 2, n = 7; Stage 3, n = 8. Significance codes: *** < 0.001; ** < 0.01; * < 0.05; · < 0.10; blank = not significant.
Results
The final dataset comprised 51 free-ranging individuals representing multiple intrinsic biological categories, including 21 males, 13 females, and 17 individuals whose sex could not be determined based on plumage characteristics. Pectoral muscle condition included 27 individuals with muscle score 2 and 24 with muscle score 3, whereas molt stage was distributed as follows: stage 0 (n = 24), stage 1 (n = 12), stage 2 (n = 7), and stage 3 (n = 8) (Supplemental Table S1). Among all biological and morphometric variables recorded during processing, including body mass, wing length, age, sex, and fat score, only pectoral muscle condition and molt stage showed significant associations with one or more hematological parameters and were therefore retained for subsequent inferential analyses.
Descriptive hematologic intervals
Descriptive hematologic intervals for M. cabanisi are presented in Table 1. Summary statistics are provided for erythrocyte parameters, leukocyte profiles, thrombocyte counts, and plasma protein concentration. The dataset included individuals representing multiple molt stages and pectoral muscle condition categories, allowing for the estimation of values across naturally occurring physiological variation within the sampled population.
Effects of molt stage and pectoral muscle condition on MCHC
Shapiro–Wilk tests confirmed that all 18 hematological parameters deviated from normality (all p < 0.05). Log-transformation failed to correct this in most cases, ruling out parametric ANOVA.
Results from both models are reported in Table 2. Of the 18 parameters, three showed significant effects of pectoral muscle condition in the separate models. MCHC produced the strongest signal (F = 24.06, df = 1, 48, p < 0.001), followed by hemoglobin concentration (Hb; F = 12.62, p < 0.001) and mean corpuscular hemoglobin (MCH; F = 5.41, p = 0.024). Molt stage had no significant effect on MCHC (F = 1.22, df = 3, 46, p = 0.314), and the full factorial model confirmed no interaction for this parameter (F = 0.67, df = 3, 42, p = 0.574), meaning the muscle effect held across all molt categories. Hb was excluded as a focal parameter because its full factorial model returned a significant interaction (p = 0.001), making its muscle effect contingent on molt stage. MCH was set aside given its borderline significance and physiological coupling to MCHC — both reflect hemoglobin content per erythrocyte.
MCHC was selected as the primary focal parameter. It combined the strongest statistical effect, a clean absence of interaction, and established biological relevance as a body condition indicator in passerines (Minias, 2015; Johnstone et al., 2017). Molt stage showed a significant effect on thrombocyte count (F = 3.71, p = 0.018) and a borderline effect on eosinophil count (F = 2.81, p = 0.050); these are reported as secondary findings in Table 2
Discussion
The distribution hematologic intervals established for M. cabanisi fill a genuine gap—no prior blood parameter data existed for this Neotropical passerine. These values give researchers a physiological baseline for interpreting individual health status and support future ecological and conservation work on this and closely related taxa 18 . The overall profile is consistent with values reported for other small passerines, suggesting broadly conserved erythrocytic and immunological characteristics across related species6,19–22. Across all 18 parameters, the majority showed no significant association with either pectoral muscle condition or molt stage, which supports the interpretation that most blood parameters reflect stable physiological baselines rather than acute responses to intrinsic biological variation under the conditions sampled.
Leukocyte distributions are known to vary with infection status, latitude, and environmental context, including urbanization gradients23–25. Urban and human-modified habitats can impose physiological challenges, including increased oxidative stress and shifts in immune investment11,26. Environmental variables were outside the scope of this study, but the descriptive hematologic intervals established here provide a benchmark against which future deviations — whether linked to habitat quality, disease, or agrochemical exposure from coffee cultivation — can be evaluated.
Body condition and erythrocyte indices
Pectoral muscle condition was the strongest predictor of hematological variation in M. cabanisi. MCHC showed the most robust association (F = 24.06, df = 1, 48, p < 0.001), with birds in better condition (Score 3) consistently showing higher values than those in moderate condition (Score 2). Hemoglobin concentration (Hb) followed a similar pattern (F = 12.62, df = 1, 48, p < 0.001), with better-conditioned birds carrying higher Hb. However, the full factorial model returned a significant interaction between muscle condition and molt stage for Hb (F = 6.40, df = 3, 42, p = 0.001), meaning the muscle effect on Hb shifts depending on molt category—which limits its usefulness as an unconditional biomarker across life-history stages. MCH showed a borderline significant muscle effect (F = 5.41, df = 1, 48, p = 0.024), corroborating the MCHC result given that both indices reflect hemoglobin content relative to erythrocyte size3,27.
Muscle condition integrates recent energetic balance and nutritional status. Its association with erythrocyte hemoglobin parameters likely reflects variation in erythropoietic regulation or plasma volume dynamics rather than pathology. Birds in better condition may sustain more optimal hemoglobin packing within erythrocytes, while those with reduced muscle mass may show lower MCHC consistent with constrained energetic or protein reserves or with compensatory erythropoietic responses under chronic physiological stress2,10,12,28.
That said, the relationship between erythrocyte indices and body condition is not universal. Powell et al. found no significant MCHC–condition association in Noisy Miners despite marked chronic stress differences between urban and rural populations 29 . Lill et al. similarly found that MCHC did not reliably track condition in Welcome Swallow nestlings 30 . These contrasts underscore that the strength and direction of hematological-condition associations depend on how the condition is measured, the life-history stage examined, and the prevailing physiological context.
The present results should not be read as implying direct causality—they document a robust covariation. MCHC was selected as the primary focal parameter because it produced the strongest and cleanest statistical signal of all 18 parameters tested, with no confounding interaction with molt stage, and because it carries an established biological rationale as an indicator of erythrocyte physiology and individual condition in free-ranging passerines. It appears responsive to intrinsic condition variation while remaining stable enough to permit comparisons across individuals in different physiological states.
Molt and hematological stability
Molt stage had no significant main effect on MCHC (F = 1.22, df = 3, 46, p = 0.314), Hb (F = 0.71, df = 3, 46, p = 0.549), or any other erythrocyte index. Despite the energetic and protein demands of feather replacement, erythrocyte hemoglobin parameters remained conserved across molt stages—consistent with the strong regulatory control of hemoglobin concentration in birds, which is tightly maintained given its role in oxygen transport. Evidence from other taxa shows mixed patterns, with some blood parameters showing transient molt-related changes while others remain stable even during energetically demanding stages14,15,31,32.
The absence of a significant interaction between molt stage and muscle condition for MCHC is particularly useful. It confirms that the positive association between somatic condition and erythrocyte hemoglobin concentration holds consistently across the molt cycle—a meaningful practical advantage for ecological monitoring, where capture timing relative to molt cannot always be controlled.
Thrombocyte count was the only parameter with a significant main effect of molt stage (F = 3.71, df = 3, 46, p = 0.018), possibly reflecting tissue remodelling or minor inflammatory responses linked to feather follicle activity. However, the full factorial model revealed a significant interaction between molt stage and muscle condition for thrombocytes (F = 4.97, df = 3, 42, p = 0.005), so the molt effect is not uniform across condition categories. Eosinophil count showed a borderline molt effect (F = 2.81, df = 3, 46, p = 0.050), possibly indicating mild immune modulation during feather replacement. Both are noted as secondary observations needing larger, more balanced samples.
The remaining 13 parameters—including PCV, RBC, MCV, WBC, and all differential leukocyte counts—showed no significant associations with muscle condition or molt stage (all p > 0.10). Leukocyte profiles were within ranges reported for healthy passerines, further supporting the view that sampled individuals were in broadly normal physiological condition.
Implications for physiological assessment and health monitoring
MCHC is the most practical single hematological parameter for assessing the physiological condition of free-ranging M. cabanisi. The bootstrapped linear model confirmed that birds with the highest muscle score had, on average, 4.82 g/dL higher MCHC (95% CI: 2.81–6.83) compared with birds in moderate condition (intercept: 33.30 g/dL, 95% CI: 31.90–34.60)—roughly a 13% difference relative to the mean. This is a biologically meaningful gap, not statistical noise. Future work integrating hematology with parasitological screening would substantially deepen understanding of immune-physiological interactions in this species, given that parasite load was not assessed here.
Conclusions and future directions
Hematological variation in M. cabanisi is driven primarily by intrinsic physiological conditions rather than molt stage. MCHC stands out as the most reliable condition-sensitive parameter (F = 24.06, p < 0.001), with Hb and MCH providing corroborating evidence, and thrombocyte count showing a secondary molt association. The remaining 13 parameters were stable across both predictors, reinforcing the robustness of the descriptive hematologic intervals established here.
These findings carry conservation relevance for a species tied to shade coffee plantations and early successional habitats increasingly threatened by urban expansion in Costa Rica. Future studies should expand sampling across seasons, age classes, and habitat types, with larger balanced designs to resolve the interaction effects detected for Hb, thrombocytes, and leukocyte subsets
Supplemental material
Supplemental material - Descriptive hematologic intervals and intrinsic drivers of variation in the costa rican endemic cabanis’s ground sparrow (Melozone cabanisi)
Supplemental material for Descriptive hematologic intervals and intrinsic drivers of variation in the costa rican endemic cabanis’s ground sparrow (Melozone cabanisi) by Randall Arguedas, Lizbeth Ovares, María de la Paz Angulo-Irola, Ernesto Carman, Mario Baldi in Avian Biology Research.
Footnotes
Acknowledgements
The authors sincerely thank Oscar Zúñiga, Julio Madriz, José Pablo Castillo, Tamara Rojas, Diego Rodríguez, and Irvin Selles for their help during fieldwork, and Eduardo Vargas for designing the location maps.
Ethical considerations
All fieldwork was conducted in accordance with national regulations and ethical standards for wildlife research. Research permits were issued by the Sistema Nacional de Áreas de Conservación (SINAC), Costa Rica (permit number: SINAC-ACC-PI-RE-042-2024). All efforts were made to minimize stress and disturbance to animals during capture, handling, and sampling procedures
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.