
Abstract
DNA-binding zinc finger (Dof) proteins are plant-specific transcription factors involved in growth, stress responses, and hormone signaling. Although genome-wide analyses of Dof genes have been reported in various plant species, the hormone-responsive expression patterns of Dof genes in blueberries have not yet been explored. In this study, we conducted the first genome-wide identification of the Dof gene family in blueberries, identifying 32 Dof genes from the Vaccinium darrowii genome. Notably, a comparative analysis of expression responses to five hormonal treatments (IAA, GA, SA, ABA, and MeJA) revealed significant differences in their temporal induction patterns: IAA and SA triggered strong upregulation of responsive Dof genes in the early phase, whereas GA-induced upregulation occurred primarily in the late phase. This hormone-specific temporal pattern has not been highlighted in previous studies of the Dof family in other species. Promoter analysis further supported these findings, revealing an enrichment of IAA- and SA-responsive cis-regulatory elements. Based on comprehensive expression profiling, we identified six Dof candidate genes potentially involved in the upregulation of IAA and GA biosynthesis, as well as five Dof candidate genes associated with ABA, SA, and MeJA biosynthesis. These candidate genes provide specific targets for future functional studies. Overall, this study not only provides a comprehensive catalog of blueberry Dof genes but also uncovers distinct hormone-specific temporal expression patterns, offering a foundation for understanding their potential roles in hormone signaling.
Introduction
The DNA-binding one zinc fingers (Dofs) family of transcription factors is widely involved in plant-specific biological processes, playing key roles in plant growth and development, 1 stress tolerance, 2 and hormone signaling. 3 Dof proteins generally possess a highly conserved N-terminus DNA-binding domain that specifically recognize cis-elements to activate or repress target gene transcription, and a non-conserved C-terminal transcriptional regulatory region. 4
With the advancement of plant genome sequencing, the Dof gene family has been systematically identified in numerous species. Higher plants generally encode more Dof genes than lower plants 5 ; for example, rice, Arabidopsis, maize, and wheat encode 30, 37, 46, and 31 Dof genes, respectively.6–9 Dof gene expression patterns are diverse and often tissue-specific. Maize ZmDof genes are predominantly expressed in male flowers, 10 while soybean GmDof11 and GmDof4 are involved in lipid biosynthesis. 11 Members of the Dof family are differentially expressed across tissues and developmental stages, suggesting functional specialization.
Dof genes are also regulated by abiotic stresses and participate extensively in phytohormone signaling. Most ZmDof genes are up-regulated under salt and drought stress. 12 In hormone pathways, rice OsDof3 is involved in gibberellin-regulated gene expression 13 ; Arabidopsis OBP1 regulates salicylic acid (SA) and auxin-responsive genes 14 ; and tomato SlDof14 participates in ethylene-regulated fruit ripening. 15 Cold stress and exogenous ABA significantly affect Dof gene expression in Dendrobium huoshanense 16 ; and abscisic acid-responsive Dof genes have been linked to fruit storage in dwarf cherry (Wei). 17
Blueberry (Vaccinium spp.) is a small berry fruit of the family Ericaceae with high nutritional and healthcare value. Its fruits are rich in anthocyanins, phenolic acids, and flavonoids, possessing antioxidant, vision-protecting, and antitumor functions.18–20 The publication of multiple blueberry genomes, including V. macrocarpon, V. darrowii, V. myrtillus, and V. corymbosum,21–25 has facilitated gene family studies in this crop. Recent research has characterized several gene families involved in blueberry development and stress responses, such as the B-box family in fruit development,26,27 VcNCED3 in postharvest fruit softening, 28 and the VdGST family in anthocyanin metabolism. 29
Regarding the Dof gene family in blueberry, a genome-wide analysis has been reported in Vaccinium corymbosum, focusing on phylogenetic relationships, gene structure, and expression patterns across tissues and under abiotic stress. 30 However, the systematic response of blueberry Dof genes to multiple hormone signals—and whether they exhibit distinct temporal expression dynamics under different hormone treatments—remains unexplored. Hormones play crucial roles in regulating blueberry development, metabolism, and fruit quality (Ghosh et al., 2015). Given the extensive involvement of Dof proteins in hormone signaling across plant species, blueberry Dof genes are highly likely to influence growth and development through hormone response pathways.
In this study, we used the Vaccinium darrowii genome 22 to perform a genome-wide identification of the Dof gene family and systematically analyzed their expression patterns under five hormone treatments (IAA, GA, SA, ABA, and MeJA). The specific questions we aimed to address were: (1) How many Dof genes exist in the blueberry genome and how are they distributed across chromosomes? (2) What are the conserved structural and phylogenetic features of blueberry Dof proteins compared to those in other species? (3) Do blueberry Dof genes exhibit distinct temporal expression patterns under different hormone treatments, and can we identify specific candidate genes potentially involved in hormone biosynthesis regulation? By answering these questions, we sought to provide a foundation for understanding the regulatory roles of Dof transcription factors in blueberry hormone signaling.
Materials and methods
Plant materials and treatments
The experiment was conducted using the highbush blueberry cultivar Vaccinium ‘Legacy’ (a hybrid with a genetic background primarily from V. corymbosum and V. darrowi). The hormone concentration of 100 μM was selected based on a previous study in blueberry, 31 in which this concentration effectively elicited physiological and molecular responses. Similar concentrations have been widely applied in gene expression studies under hormone treatments in other fruit crops. Five treatments were applied: blueberry plants were irrigated with 100 μM MeJA, ABA, GA, IAA, or SA to simulate different hormonal stresses. Each treatment was conducted independently. Leaf samples were collected at 0 (control), 3, 6, 12, 24, and 48 h post-treatment. Three biological replicates were collected at each sampling time point and treatment condition. Following sampling, all specimens were rapidly frozen in liquid nitrogen and stored at −80°C for subsequent RNA extraction experiments.
Dof gene family identification, sequence analysis and chromosomal localization
The genome and annotation files for Vaccinium darrowii 22 were downloaded from Phytozome (https://phytozome-next.jgi.doe.gov/). The whole-genome sequences, including primary and secondary assemblies, are available at DDBJ/ENA/GenBank under accessions JAFMTH000000000 (BioProject PRJNA706655) and JAFMTI000000000 (BioProject PRJNA707651). To identify members of the Dof family, the HMM profile for the Dof domain (Pfam ID: PF02701) was downloaded from the Pfam database, and the blueberry proteome was aligned using the HMMER software with default parameters. To further enhance sensitivity, known Dof protein sequences from Arabidopsis thaliana were retrieved from Phytozome and used as query sequences for BLASTP alignment against the blueberry proteome, with an E-value threshold set at 1e−5. Candidate sequences overlapping in the HMMER and BLASTP searches were merged and subsequently submitted to the SMART database and the NCBI Conserved Domain Database (NCBI-CDD) to confirm the presence of a complete Dof domain. Additionally, redundant sequences and alternative splicing variants were excluded: for genes with multiple transcript isoforms, only the longest transcript was retained as the representative. The remaining genes were designated as blueberry Dof genes and named according to their chromosomal locations. All sequence extraction and processing steps were performed using the TBtools software.
After identifying the specific blueberry Dof family gene members, the physicochemical properties of the Dof sequence proteins were predicted using the ExPASy website (http://web.expasy.org/protparam/), and the protein secondary structure analysis was performed using the online website NPS@: Network Protein Sequence@nalysis (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html). For protein tertiary structure analysis of Dof family members using the swiss-model online website (https://swissmodel.expasy.org/). Subcellular localization was predicted in silico using the WoLF PSORT web server (https://wolfpsort.hgc.jp/).
Chromosomal localization and colinearity analysis of Dof transcription factors in blueberry
Using TBtools software to parse blueberry annotation files, we generated a physical map of blueberry Dof genes on chromosomes and visualized gene density bands. Dof gene duplication events were analyzed using MCscanX (Tang et al., 2008) and visualized within TBtools.
Analysis of blueberry Dof gene structure and conserved structural domains
The protein sequences of blueberry Dof family members were aligned and compared by MEGA software, and then the results were optimized by ESPript 3.0 online website (https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi). The analysis of conserved structural domains of blueberry Dof proteins was completed by the online software MEME (http://meme-suite.org/tools/meme). Blueberry Dof genes structure and conserved motifs were analyzed using TBtools software.
Multiple sequence alignment and evolutionary tree analysis
Cluster analysis was performed on 32 blueberry Dof family members, Arabidopsis thaliana, and Vitis vinifera. Arabidopsis Dofs gene information was sourced from the online database Tair (https://www.arabidopsis.org/), while grape Dofs gene information was obtained from the online database CRIBI Genomics (http://genomics.cribi.unipd.it). The 32 Dof protein sequences formed a data matrix. Sequence alignment was performed using the MAFFT online software's L-INS-i algorithm. A phylogenetic tree was constructed using the Maximum Likelihood (ML) method in MEGA software, with 1000 bootstrap repetitions. Tree visualization was performed using the online platform ChiPlot (https://www.chiplot.online/), 32 for subfamily classification and annotation of blueberry and Arabidopsis gene family members.
Promoter sequence binding element analysis
The 2000 bp preceding the CDS sequence of the blueberry Dof gene family was extracted (Table S1), then cis-acting element prediction was performed through the PlantCare (http://www.plantcare.co.uk/) website, and the results were visualized and plotted in TBtools.
RNA extraction and real-time fluorescence quantitative PCR validation
Total RNA was extracted from the leaves of blueberries under different treatments using RNA Simple Total RNA Kit (TIANGEN, Beijing, China), and RNA quality and concentration were determined by NanoDrop 2000 (Thermo Scientific, Massachusetts, USA), while 1% agarose gel electrophoresis was used to verify the extracted RNA integrity was verified. Total RNA was reverse transcribed into cDNA using M-MLV reverse transcriptase (Takara Bio Inc., Otsu, Japan). RT-qPCR reactions were carried out using TB Green Premix Taq II (Tli RNASEH Plus) (TaKaRa, Dalian, China) in the quantitative instrument Quant Studio 3 (ABI, Thermo Fisher Scientific, USA) for quantitative fluorescence analysis. Gene expression levels were calculated using the 2−ΔΔCt method. To validate the 2−ΔΔCt method, the consistency of amplification efficiencies between target and reference genes was verified. A pooled cDNA sample was serially diluted 4-fold, and the ΔCt values (Ct_target − Ct_GAPDH) were plotted against the log of the dilution factor. For all primer pairs, the absolute slope was <0.1, indicating approximately equal amplification efficiencies and thus satisfying the requirement for the 2−ΔΔCt method.The blueberry GAPDH gene was used as an internal control (Table S2); detailed primer information for the blueberry Dof gene is provided in Table S2.
Gene expression pattern analysis and data processing
Based on RNA-seq data, functional analysis of cis-acting elements responsive to five hormones (IAA, GA, SA, ABA, MeJA) identified 10 Dof candidate genes for gene expression pattern analysis. Hierarchical clustering heatmaps and visualization were performed using TBtools. One-way ANOVA followed by Dunnett's post hoc test was conducted using SPSS 26.0 software (IBM Corp., Armonk, NY, USA) to compare each treatment group against the control group at each time point. Statistical significance was indicated as ∗p ≤ 0.05, ∗∗p ≤ 0.01, and ∗∗∗p ≤ 0.001. Bar charts were generated using Origin software (OriginLab Corp., Northampton, Mass., USA).
Results and analysis
Screening of Dofs family members responsive to blueberry leaf hormones
By comparing online databases, this study identified a total of 32 blueberry Dofs transcription factors. Through comparative analysis with transcriptome sequencing results from blueberries subjected to five treatments (MeJA, ABA, GA, IAA, SA), 10 Dofs transcription factors responsive to hormonal treatments were ultimately identified.
Identification and chromosomal localization characteristics of Dof genes in blueberry
A total of 32 blueberry Dof transcription factors (named sequentially according to their positions on chromosomes) were identified by searching for Dof genes in the whole blueberry genome by HMM and calibrating their conserved structures (Fig. 1). 31 Dof genes were scattered on 12 chromosomes (Fig. 1), of which Chr6 had the highest distribution of Dof family member genes with 6 (Dof12-Dof17) or about 19.4%. The least distribution of Dof genes on Chr1 and Chr7 were both 1. The two Dof genes on Chr4, Chr5 and Chr12, and three Dof genes on Chr2, Chr3, Chr8, Chr9 and Chr11. In addition, the Dof32 gene, whose localization information was not present in the blueberry genome sequence and could not be displayed, was located on scaffold4.
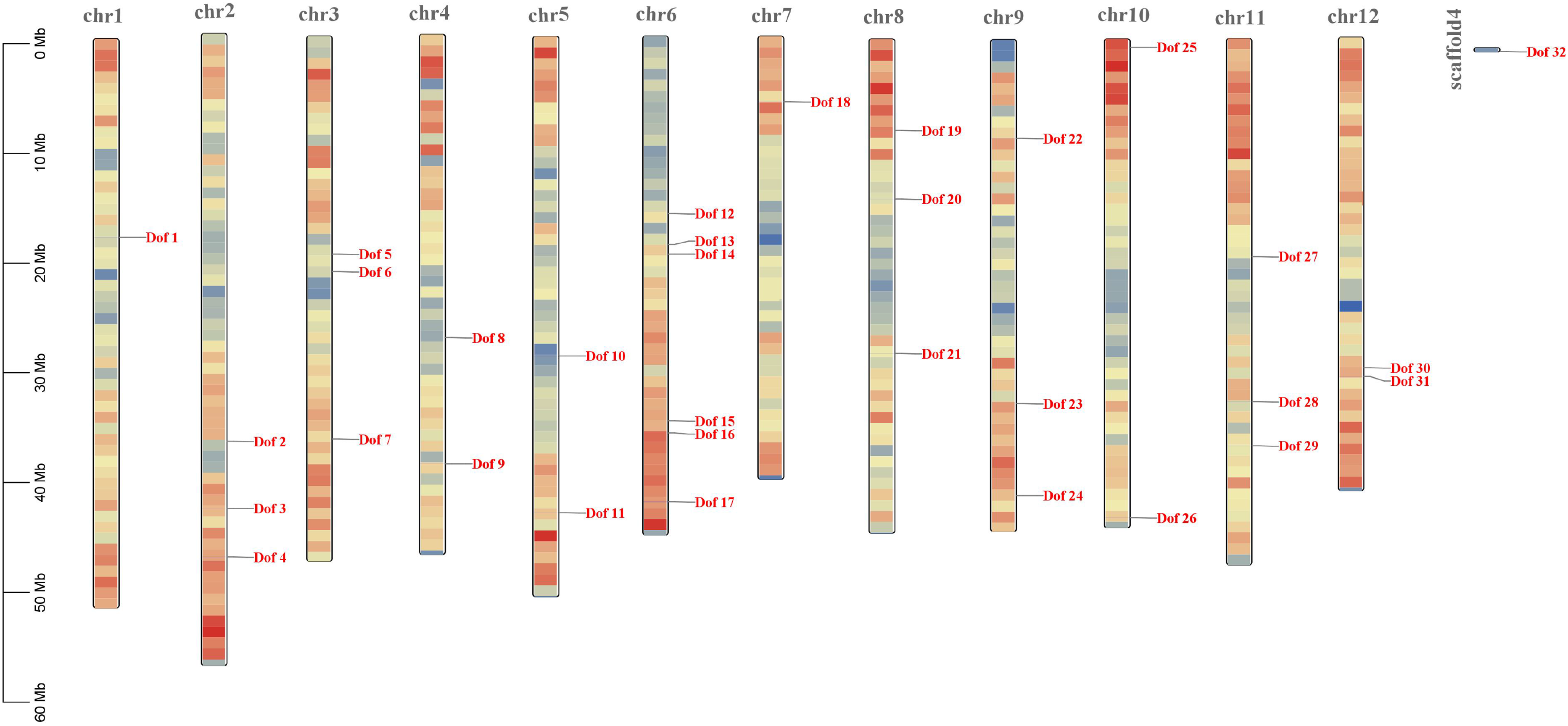
Chromosomal localization map of the blueberry Dof genes. The color of the chromosome bands shows the gene density, with the red region having a high number of genes and a high gene density, and the blue region having a low number of genes and a low gene density.
Colinearity analysis of blueberry Dof gene family members
Analysis of collinearity among Dof genes in blueberries (Figure 2) revealed 15 collinear gene pairs across 12 chromosome scaffolds, exhibiting uneven distribution. These paired genes share nearly identical gene structures and reside within the same subfamily. Chr1, Chr6, Chr9, Chr11, and Chr12 harbored the highest number of collinear genes, totaling 10 pairs. No collinear genes were identified on chromosomes Chr5, Chr7, or Chr3. Significant gene clusters exist on Chr3, Chr6, and Chr12. These four independent tandem repeat gene clusters (Dof5 and Dof6, Dof13 and Dof14, Dof15 and Dof16, Dof30 and Dof31) may represent gene duplication events.
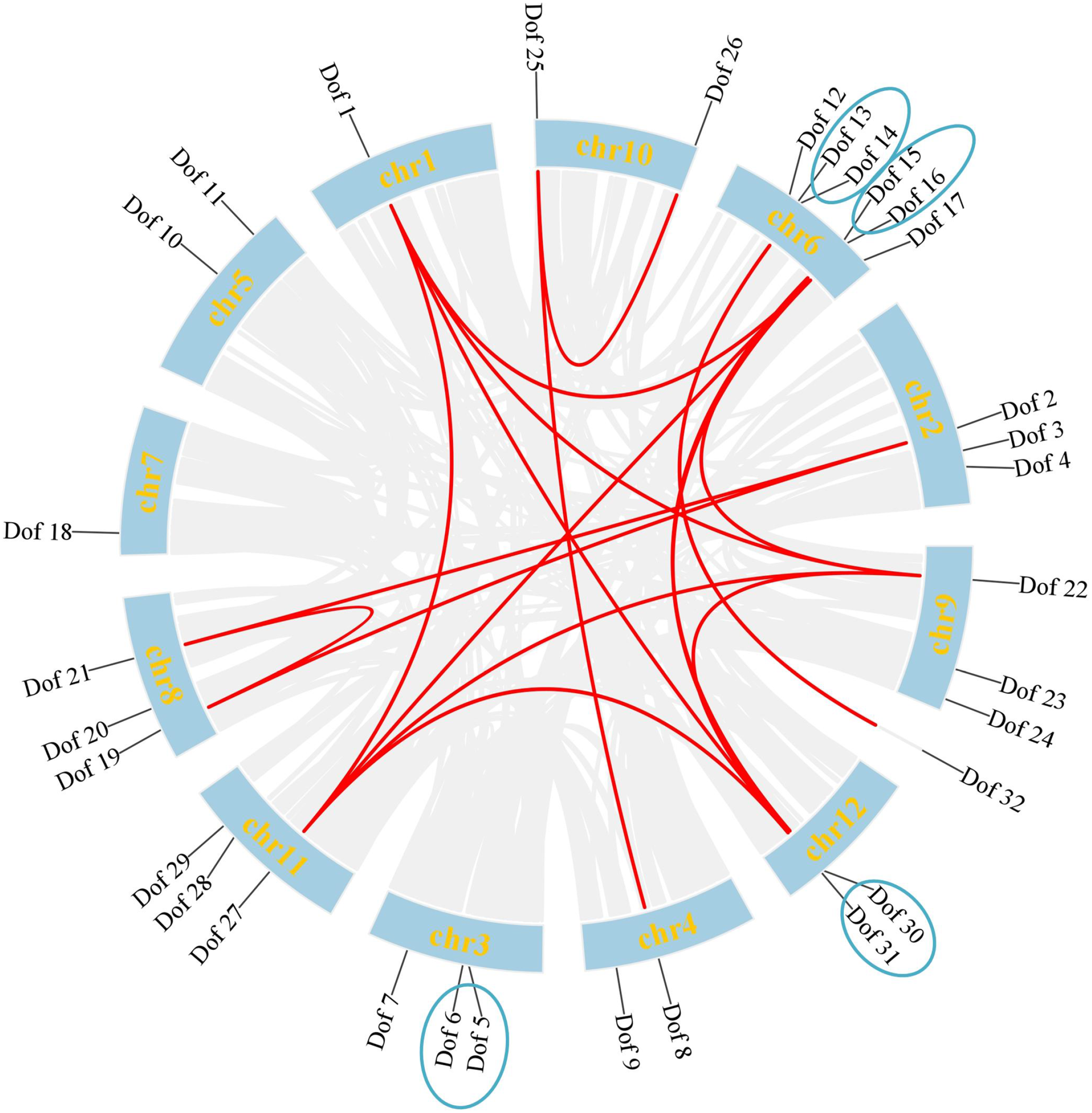
Collinearity analysis of Dof genes in blueberry. Gray lines represent all collinear blocks across the genome. Colored lines highlight collinear relationships between Dof gene pairs. Among these, gene pairs connected by red lines are segmentally duplicated, while those marked with blue circles are tandemly duplicated based on their close physical proximity on the same chromosome.
Physicochemical characterization of the Dof gene family in blueberry
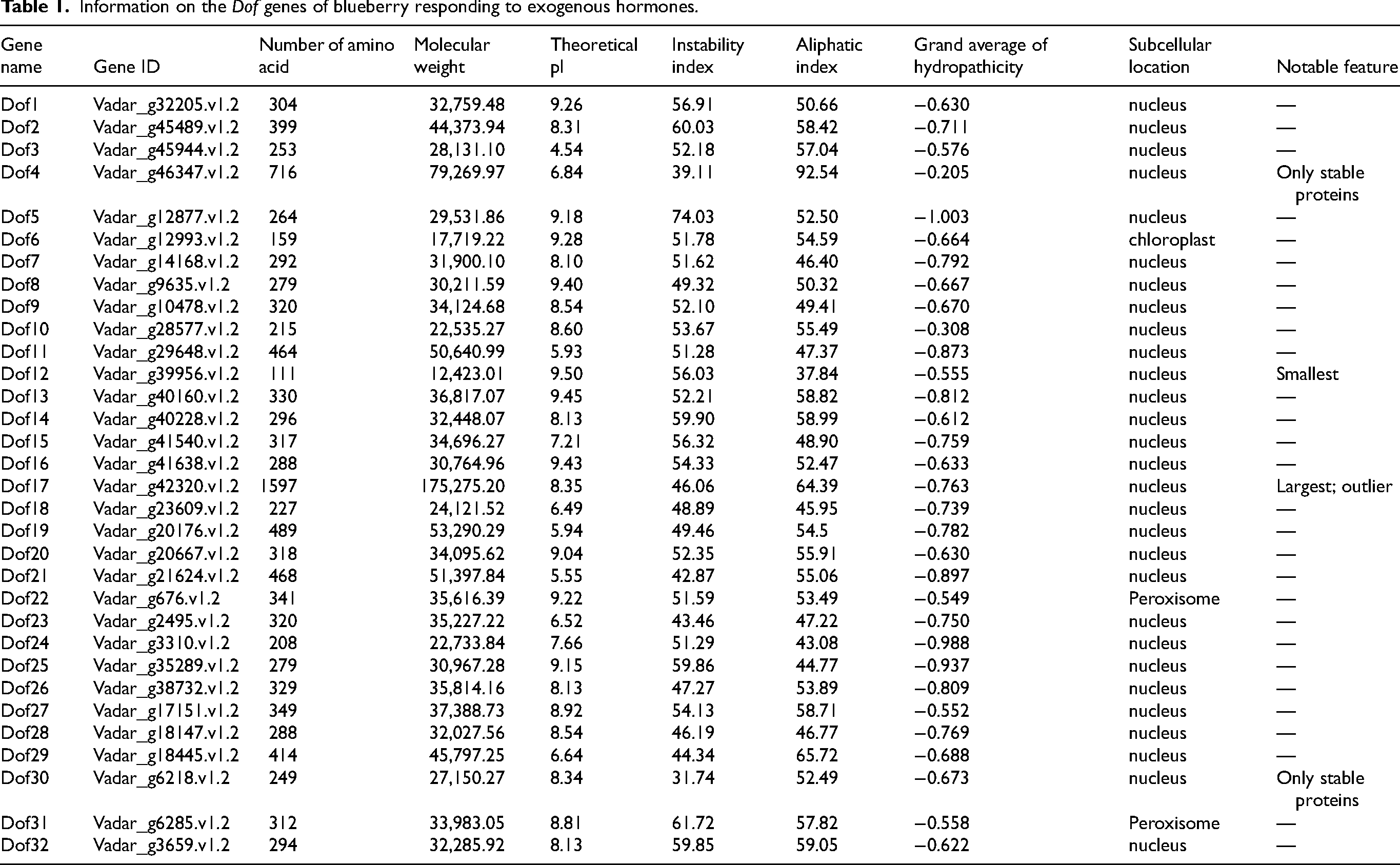
Physicochemical analysis of the 32 Dof gene family member proteins (Table 1) revealed that while most members exhibited typical characteristics, two significant outliers were present: Dof12 was the smallest protein in the family (12.42 kDa, 111 amino acid residues), whereas Dof17 was the largest (175.28 kDa, 1597 amino acid residues). The molecular weight of Dof17 exceeds that of Dof12 by more than 14-fold, indicating functional diversification, with larger Dof proteins potentially harboring additional domains for protein–protein interactions in hormone signaling complexes. The remaining proteins cluster within the conventional molecular weight range of 17.72–79.27 kDa. Theoretical isoelectric points (pI) ranged from acidic (4.54) to basic (9.50), with an average pI of 8.04, indicating an overall basic pI tendency among blueberry Dof family members. This basic pI tendency may facilitate electrostatic interactions with DNA during transcriptional regulation of hormone-responsive target genes. Regarding instability coefficients, only Dof4 and Dof30 were below 40, indicating relatively high amino acid hydrophilicity, while all other genes exceeded 40. All Dofs exhibited hydrophilicity coefficients below zero, confirming the hydrophilic nature of blueberry Dof proteins. Additionally, their lipophilicity coefficients were all below 100, further emphasizing their hydrophilicity. The general instability is consistent with the dynamic regulatory roles of transcription factors in hormone signaling, allowing rapid protein turnover in response to hormonal fluctuations. Furthermore, subcellular localization predictions indicate that most proteins localize to the nucleus. Notably, Dof6 localizes to chloroplasts, while Dof22 and Dof31 localize to peroxisomes. These atypical localizations suggest non-canonical functions, potentially linking Dof-mediated hormone responses to organelle-specific processes such as redox regulation and retrograde signaling.
Information on the Dof genes of blueberry responding to exogenous hormones.
Secondary and tertiary structure analysis of blueberry Dof family member proteins
There are regular repeats of proteins in protein secondary structure conformation, which can be derived from the distribution of protein secondary structure of blueberry Dof gene family members, which consists of alpha helix, extended strand, and random coil, but there is no beta turn, in which the random coil mainly serves to random coil mainly plays the role of connecting other structures. The prediction and analysis helped to understand the spatial structure of blueberry Dof proteins. The predicted results showed that the proportion of secondary structures of most Dof proteins ranged from random coil > extended strand > alpha helix, and the proportion of alpha helix ranged from 1.69% (Dof14) to 38.69% (Dof4), and the proportion of extended strand ranged from 3.72% (Dof27) to 10.81% (Dof12); and random coil ranged from 52.93% (Dof4) to 92.95% (Dof18) (Fig. 3(a)). Overall, a few alpha helix and extended strands were scattered throughout the amino acids and were connected by random coil.

Distribution of blueberry Dof proteins secondary structure (a) and tertiary structure analysis (b). In the protein secondary structure distribution diagram, blue indicates alpha helix, purple indicates extended strand, and orange indicates random coil. Different colors in the protein tertiary structure distribution diagram are used to distinguish different helices and folding regions.
Modeling and analysis of blueberry Dof protein members using SWISS-MODEL online retrieved the sequences F6M8H3.1.A and P68350.1.A with the highest similarity to the Dof5 protein sequence and the Dof12 protein sequence, with 60% and 63% similarity, respectively. The sequences with the highest similarity to Dof4, Dof5, Dof7, Dof10, Dof15, Dof18, Dof23, and Dof24 protein sequences were A0A834FZC0.1.A, A0A834LVZ1.1.A, A0A5B7B743.1.A, A0A7J7IA51.1.A, I1LR01.1.A, A0A834LMR2.1.A, I1KCF8.1.A, A0A6A4MS98.1.A, with a similarity range of 50% to 60%. Similarity to Dof1, Dof2, Dof8, Dof9, Dof11, Dof13, Dof14, Dof16, Dof19, Dof20, Dof21, Dof22, Dof25, Dof26, Dof28, Dof29, Dof30, and Dof32 protein sequences with the highest sequence similarity ranging from 40% to 50%. The sequences with the highest sequence similarity with Dof3 and Dof17 protein sequences were B9FPT7.1.A and 9evq.1.A. The protein tertiary structure coincided with the proportion of each structure of the secondary structure, and the protein tertiary construction model diagram is detailed in the Fig. 3(b).
Conserved structural domains and gene structure analysis of Dof transcription factors
Using the ESPript 3.0 online platform, we aligned the amino acid sequences of 32 blueberry Dof genes. The results revealed that the Dof domain exhibits high conservation (Figure 4), featuring a fixed C2-C2 structure. Amino acids in other domains also showed remarkable similarity, consistent with previous studies. 15 Based on the full sequences, a phylogenetic tree was constructed using MEGA7.0 software (Figure 4(a)) to analyze the phylogenetic relationships among blueberry Dof family members. Visualization of Dof gene structures using TBtools software (Figure 5(b)) revealed that all 32 Dof genes contained exons. Ten Dof genes lacked introns, 19 contained only one intron, Dof13 and Dof4 contained two introns, while Dof17 contained ten introns. This may be driven by the recent emergence of LTR retrotransposon-specific insertions into intronic regions of genes, accumulating and persisting under its specific evolutionary history (polyploidy, allopolyploidy, woody growth) and relatively weak purifying selection pressure. 33 Genes with closely clustered relationships exhibited similar structural patterns. Protein motif analysis via MEME (Figure 5(b) and (c)) revealed family members sharing similar motifs clustered within the same evolutionary branch. Based on phylogenetic relationships, gene structure, and protein motif distribution, Dof genes were clustered into six major groups (I-VI). Groups I, II, III, IV, V, and VI contained 2, 2, 12, 2, 8, and 5 Dof proteins, respectively. Due to its significant divergence, Dof17 was excluded from classification. Motif 1 represents the signature domain of Dofs, while the remaining motifs exhibit greater diversity and variation, yet remain conserved within their respective clades. Motif 4 is a domain specific to Cluster III. Motifs 2, 5, and 8 are unique to Cluster III. Motif 7 is present only in Clusters III and V, while Motif 10 is found exclusively in Clusters II and V.
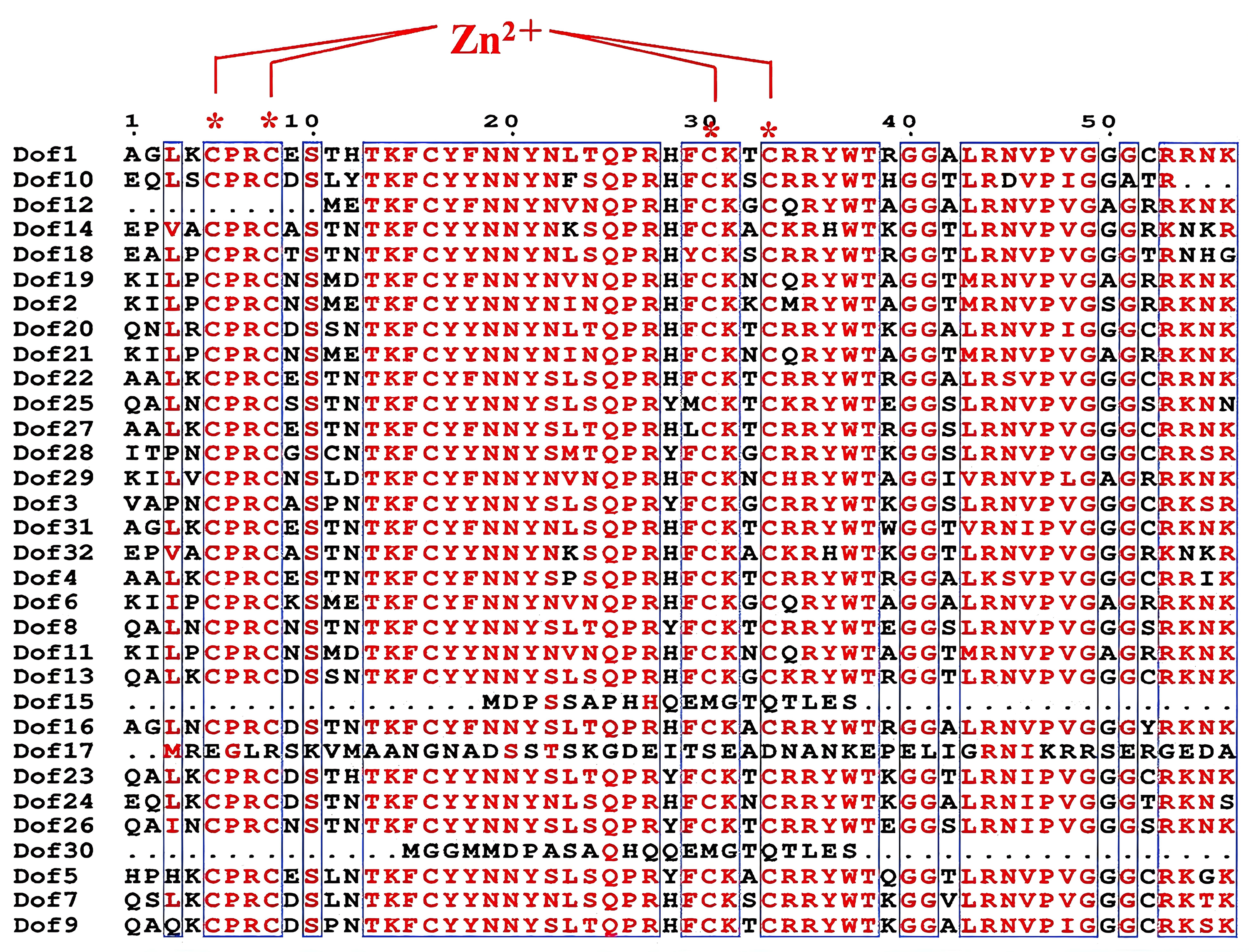
Sequence comparison of conserved structural domains of blueberry Dof proteins. Protein sequence sites identified in red are aligned, and boxes indicate that the sequences match exactly at the corresponding positions.
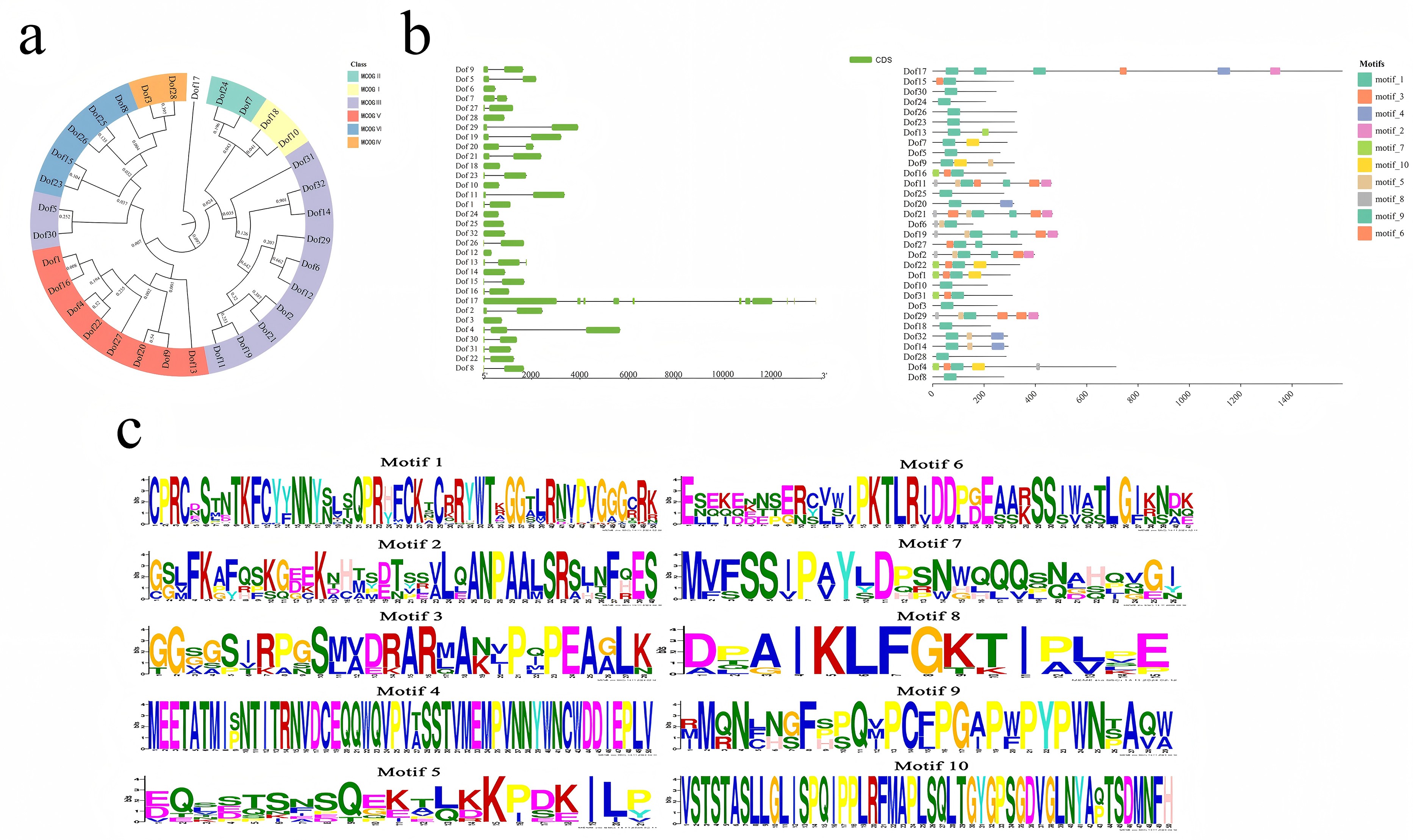
Analysis of 32 Dof protein conserved domains and gene structures note: a: phylogenetic relationships; b: gene structure and protein motif distribution diagram; c: analysis of conserved protein motifs (motif 1–10).
Evolutionary tree analysis of 32 Dof proteins
To investigate the molecular evolution and phylogenetic relationships of Dofs in blueberry, Arabidopsis, and grape, a mixed tree was constructed using the ML method (Figure 6). We integrated 32 blueberry Dof proteins, 34 Arabidopsis AtDof proteins, and 25 grape VviDofL proteins into a single data matrix. Following Da's classification method for grape VviDofL proteins (Da et al., 2016), blueberry Dof proteins in the phylogenetic tree were grouped into six major Dofs clusters (MCOG I, II, III, IV, V, VI). The clustering results showed that MCOG I contained 2 blueberry Dof proteins, MCOG II contained two blueberry Dof proteins, MCOG III contained twelve blueberry Dof proteins, MCOG IV contained two blueberry Dof proteins, MCOG V contained eight blueberry Dof proteins, and MCOG VI contained five blueberry Dof proteins. Dof transcription factors from the three species show no distinct separation in the phylogenetic tree, suggesting that Dof proteins from these three species have not undergone significant lineage-specific divergence, and may retain ancestral functions shared across species. Members within each clade are predicted to share functional similarity based on phylogenetic proximity, although experimental validation is required to confirm functional equivalence, and the clustering distribution results align with the internal clustering and structural analysis of blueberry Dof proteins shown in Figure 5. This phylogenetic clustering pattern provides a framework for hypothesizing functional conservation within subfamilies; however, functional divergence among closely related members cannot be ruled out without experimental evidence.
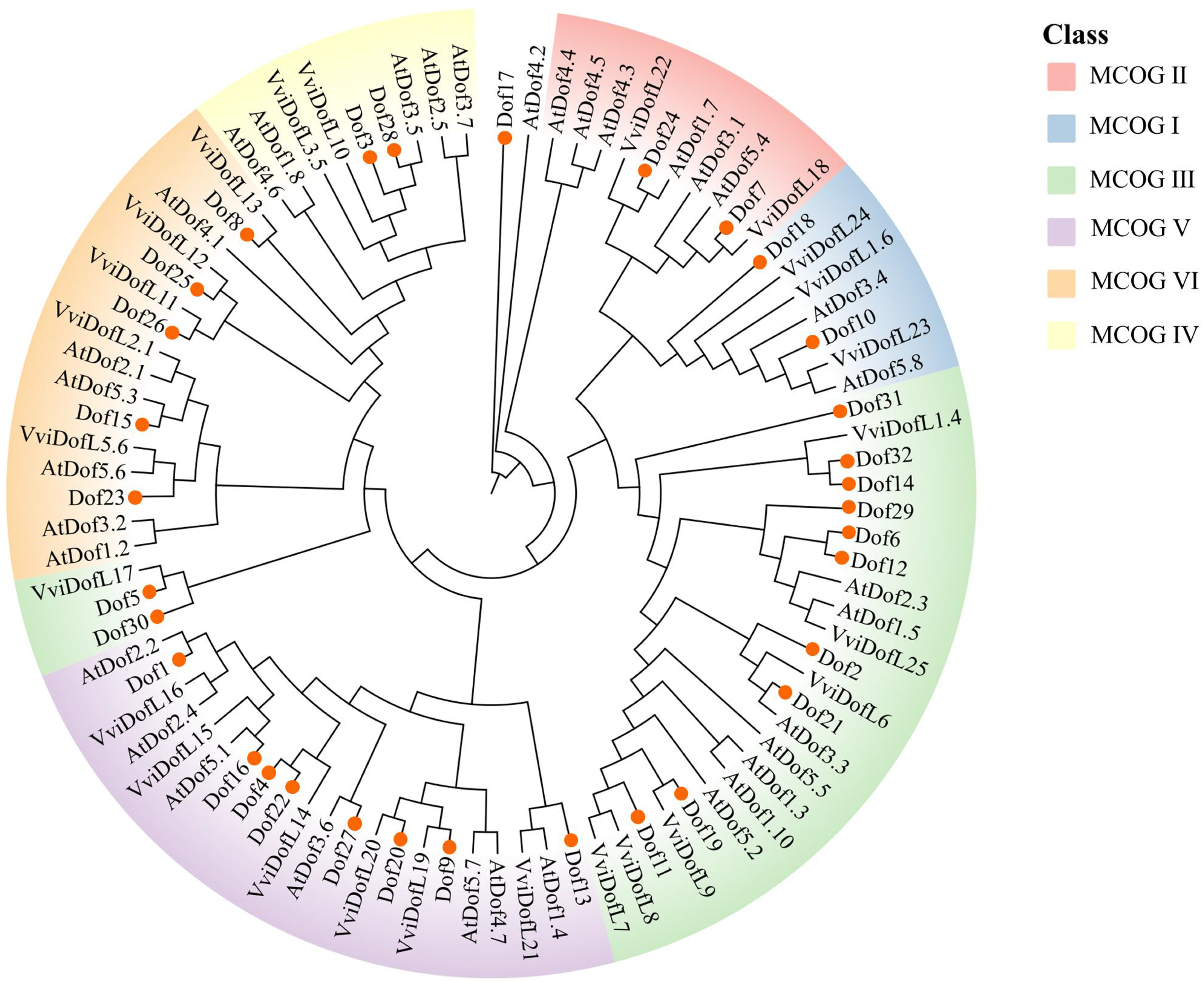
Phylogenetic tree of Dofs in blueberry, grape and Arabidopsis,. Different colors represent different subfamilies, with six colors denoting subfamilies (MCOGI-VI); red circles indicate blueberry Dof family protein members.
Cis-acting elements of blueberry Dof family members
Analysis of cis-acting elements in 32 blueberry Dof gene family members revealed multiple binding elements with variations among different genes, suggesting a complex expression regulation mechanism for Dof transcription factors (Figure 7). Most Dof family members were found to contain elements associated with plant hormones, such as GARE-motif (gibberellin), P-box, AuxRR-core, CARE, as-1, TGACG-motif, CGTCA-motif (methyl jasmonate-responsive), TGA-element (auxin), TCA-element (salicylic acid-responsive), ERE, ABRE (abscisic acid-responsive), and light-responsive cis-acting elements like G-box were also identified in Dof promoter sequences. Dof also possesses stress-related cis-acting elements, including DRE core (drought, salt, cold-responsive), TC-rich repeats (defense-responsive), WUE-motif, MYC, W box, STRE, MYB, MBS (drought-inducible), LTR (cold-responsive), and ARE (anaerobic-inducible). Furthermore, MYB binding sites can interact with MBS to regulate the expression of genes associated with the cytokinin signaling pathway. Among blueberry Dof family members, ABA response elements totaled 24 (26.7%); MeJA response elements numbered 16 (17.8%); GA response elements were 18 (20.0%); IAA response elements were 18 (20.0%); and SA response elements were 14 (15.6%) (Figure 7(a)). Based on function, all element promoters were classified into three categories: plant growth and development, plant hormone response, and plant stress defense response. Among elements related to plant stress defense response, MYB promoters exhibited the highest gene distribution in Dof, accounting for 30%. For plant hormone response elements, ABRE promoters were the most common (26%), representing ABA response elements. Among elements related to plant growth and development, G-box promoters were the most prevalent, accounting for 24%. The occurrence frequency of these three promoters was significantly higher than that of random distribution across the entire genomic background (p < 0.01), demonstrating that these three promoters may have functional associations with Dof genes rather than being merely coincidental. Furthermore, only Dof11 transcription factors responded to five types of hormone-responsive elements, while nine transcription factors—Dof1, Dof5, Dof7, Dof8, Dof17, Dof21, Dof26, Dof29, and Dof30 responded to four types of hormone-responsive elements. Interestingly, both the cis-acting elements responding to biotic and abiotic stresses in blueberries and those responding to exogenous hormones are abundant. All transcription factors contain MYB cis-acting elements, suggesting that Dof family members also play a significant role in blueberry drought tolerance.
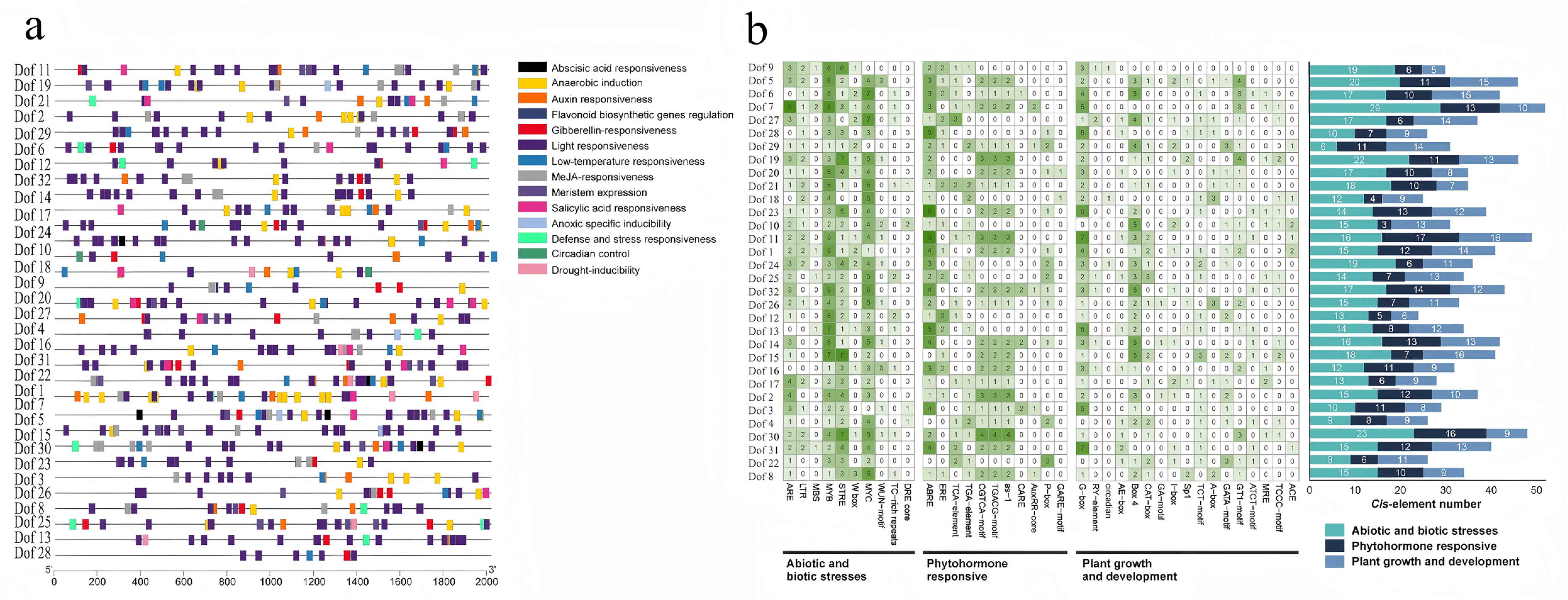
Analysis of Dof transcription factor gene promoter elements responding to five hormones. a: Distribution map of promoter response elements; b: Summary of the number of different promoter response elements distributed, with three colors representing the number of elements responding to abiotic stress, responding to hormonal stress, and responding to plant growth and development, respectively.
Expression patterns of blueberry Dof genes in response to exogenous hormones
Dof gene functions are directly or indirectly related to plant hormone regulation, with their expression patterns influenced by different categories of hormones. Simultaneously, hormones exert significant effects on blueberry growth and development, playing a crucial role in regulating flavor, nutrient content, high yields, and stable production. 34 Based on the above cis-acting element analysis, the promoter regions of the blueberry Dof gene family were found to be enriched in five types of hormone-related cis-acting elements (TGA-element, GARE-motif, ABRE, TCA-element, and CGTCA-motif), suggesting their extensive involvement in hormone signaling. Accordingly, 10 candidate Dof genes responsive to five hormones (IAA, GA, SA, ABA, and MeJA) were selected for RT-qPCR validation and expression pattern analysis (Figure 8), providing a foundation for future functional studies of Dof genes. Results revealed that the expression patterns of these 10 blueberry Dof genes were induced or suppressed to varying degrees following treatment with the five hormones, suggesting that these Dof genes may be broadly involved in blueberry hormone response pathways.
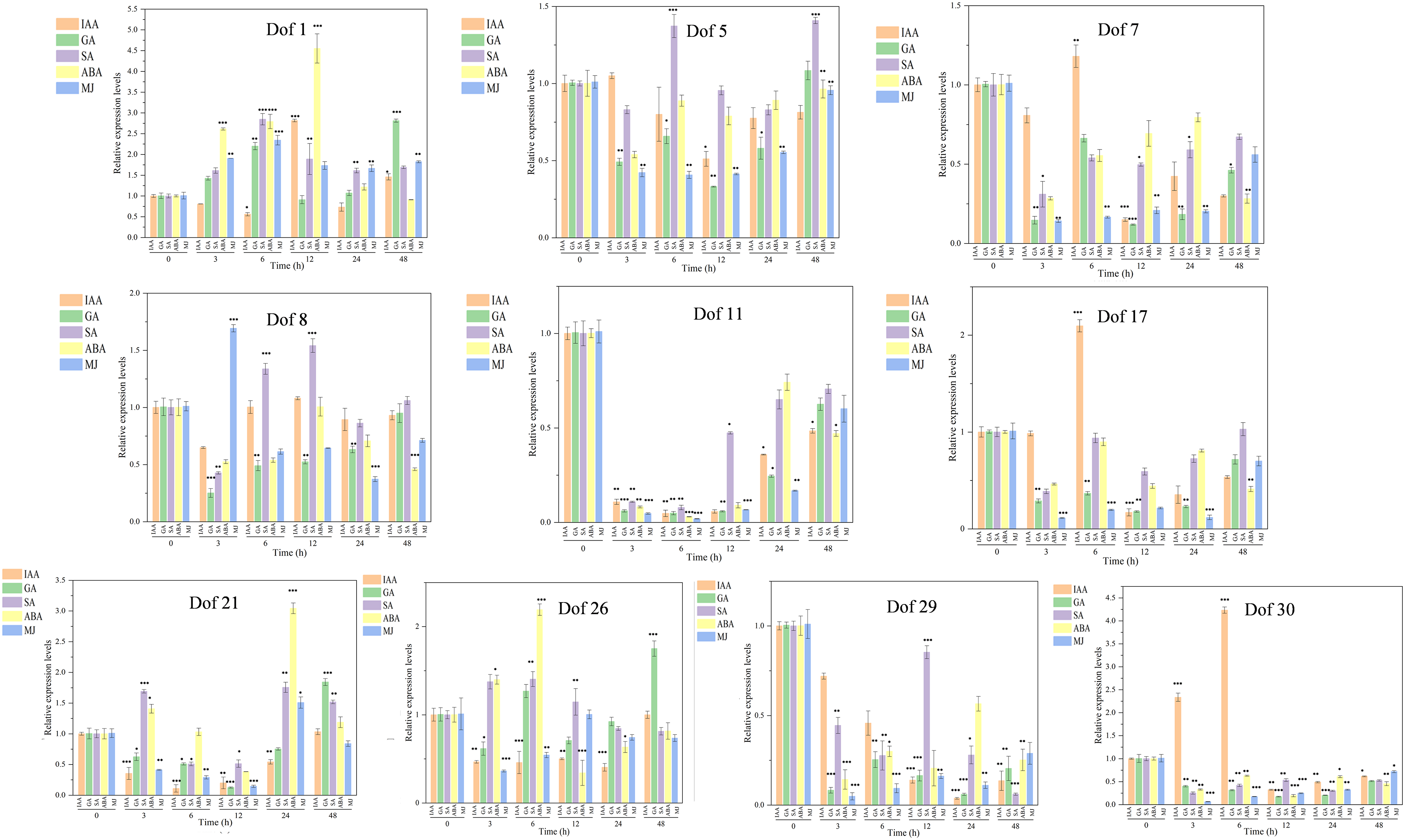
Expression analysis of 10 Dof genes in response to five hormone treatments. Error bars indicate standard deviation. ∗ denotes significant differences between control and treatment groups. ∗ p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001.
Overall, the 10 hormone-responsive Dof genes exhibited distinct expression patterns under the five hormone treatments, with IAA and SA eliciting the strongest and most rapid responses, while MeJA induced the weakest. A general trend emerged in which IAA and SA primarily triggered early upregulation, GA induced delayed responses, and ABA and MeJA tended to suppress Dof gene expression.
At 3 h of IAA hormone treatment, the Dof30 gene expression level was 2.3-fold higher than the control group, reaching its peak at 6 h (approximately 4.25-fold higher than the control group), with significant differences. Concurrently, the Dof7 and Dof17 genes also showed upregulation at 6 h, with expression levels 1.75-fold and 2.10-fold higher than the control group, respectively. These results indicate that Dof30, Dof7, and Dof17 are early-response genes to IAA treatment in blueberries, playing roles during the initial stages of IAA exposure. Furthermore, the four genes Dof11, Dof29, Dof21, and Dof26 showed significant suppression during IAA treatment. After 48 h of GA treatment, Dof5, Dof21, Dof1, Dof26 showed significant induction, with peak expression levels reached. Notably, Dof1 expression reached 2.75 times that of the control group. Dof8 expression exhibited an upward trend throughout GA treatment. Between 6 and 48 h, Dof5 expression showed a decrease followed by an increase after both IAA and GA treatments, indicating that IAA and GA share similar gain effects and produce synergistic interactions. Under SA treatment, five genes were upregulated and five were downregulated, with Dof26 exhibiting high expression during the initial 12 h. Under ABA treatment, three Dof genes (Dof21, Dof1, Dof26) showed more than double the expression levels compared to the control group, with Dof1 reaching 4.5-fold. Overall, gene expression levels following IAA and GA treatments were significantly lower than those after ABA treatment. This indicates ABA treatment more effectively modulates Dof gene expression than IAA or GA, with a greater number of genes showing up- or downregulation in the later stages compared to the early stages. Under MeJA treatment, seven Dof genes exhibited significant downregulation, with Dof8 being an early-response gene to MeJA. As MeJA treatment duration increased, the number of differentially expressed genes in blueberry leaves continued to rise, indicating that prolonged treatment primarily induced gene expression in response to MeJA. Overall, the 10 Dof genes exhibited substantial variation in response to MeJA, ABA, GA, IAA, and SA induction, though the response patterns and magnitudes differed among hormones. Among these, IAA treatment elicited the strongest response, followed by SA treatment, while MeJA induced the weakest response. Analysis of cis-acting specific recognition elements revealed that IAA-responsive elements were the most abundant, followed by SA-responsive elements. Furthermore, Dof gene responses under IAA and SA treatments were more pronounced than under other hormone treatments (Table 2). Therefore, it can be inferred that blueberries may play a crucial role in IAA- and SA-related pathways through Dof genes.
Hormone-responsive Dof genes in blueberry under five hormone treatments.

Values represent relative expression levels (2−ΔΔCt) compared to the control group (0 h). The time point of maximum significant response is indicated in parentheses (h). Asterisks denote statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001). – indicates no significant differential expression.
To assess whether the composition of cis-acting elements in promoters correlates with the observed expression patterns, we examined the relationship between hormone response elements and corresponding gene expression data. The results revealed that genes exhibiting strong responses to IAA treatment (Dof30, Dof7, Dof17) contain auxin response elements (TGA elements) in their promoters. Similarly, genes highly induced by SA treatment (such as Dof26, which exhibited sustained high expression during the first 12 h) contained SA-responsive TCA elements. In contrast, genes primarily repressed by MeJA and ABA (e.g., Dof8, Dof5) carried fewer JA- and ABA-responsive elements than IAA/SA-induced genes. Among the five types of hormone-associated elements, IAA- and SA-responsive elements were the most abundant, consistent with the stronger and more rapid expression responses observed under IAA and SA treatment compared to other hormones. These convergent results suggest that the structure of promoter cis-acting elements may contribute to the differences in hormone responsiveness among blueberry Dof genes.
Characteristics and patterns of blueberry dofs genes responding to five hormones
To investigate the response patterns of blueberry Dof transcription factors under exogenous hormone treatments (IAA, GA, SA, ABA, MeJA), this study analyzed their expression profiles using related transcriptome data and performed cluster analysis (Figure 9). Results revealed significant variations in gene expression levels across different hormone treatment durations. Across all hormone treatments, Dof1 and Dof5 exhibited consistently higher expression levels than other genes, with Dof1 reaching 4-fold expression after 12 h of ABA treatment. In MJ treatment, except for Dof1 and Dof8, other genes showed extremely low expression levels (<1-fold) during the initial treatment period (3–24 h), indicating a pronounced inhibitory effect. During ABA treatment, except for Dof1, Dof5, Dof7, and Dof8, other genes exhibited a pattern of initial decrease, followed by increase, and then decrease in expression levels. They were downregulated after 12 h of ABA treatment, reaching the lowest expression levels (<1-fold), while Dof21 showed upregulation after 24 h of ABA treatment (≥3-fold increase). Under SA treatment, except for Dof29 and Dof30, other genes were upregulated (expression ≥1-fold) during the late stage (24–48 h). Dof1 reached its highest expression (≥3-fold) after 48 h of SA treatment. In GA treatment, most genes were only upregulated (expression ≥1.5-fold) after 48 h of treatment. with Dof1 reaching 3-fold expression at this time. Conversely, Dof7, Dof29, and Dof30 showed significant downregulation (expression <0.5-fold) at 12–24 h post-treatment. Under IAA treatment, most genes (except Dof1 and Dof8) exhibited marked downregulation (<1-fold) after 12 h, while Dof1 expression increased approximately 3-fold. Most genes showed upregulation (≥2-fold) during the early phase (0–6 h) of IAA treatment, with Dof30 reaching 4.5-fold expression at 6 h.
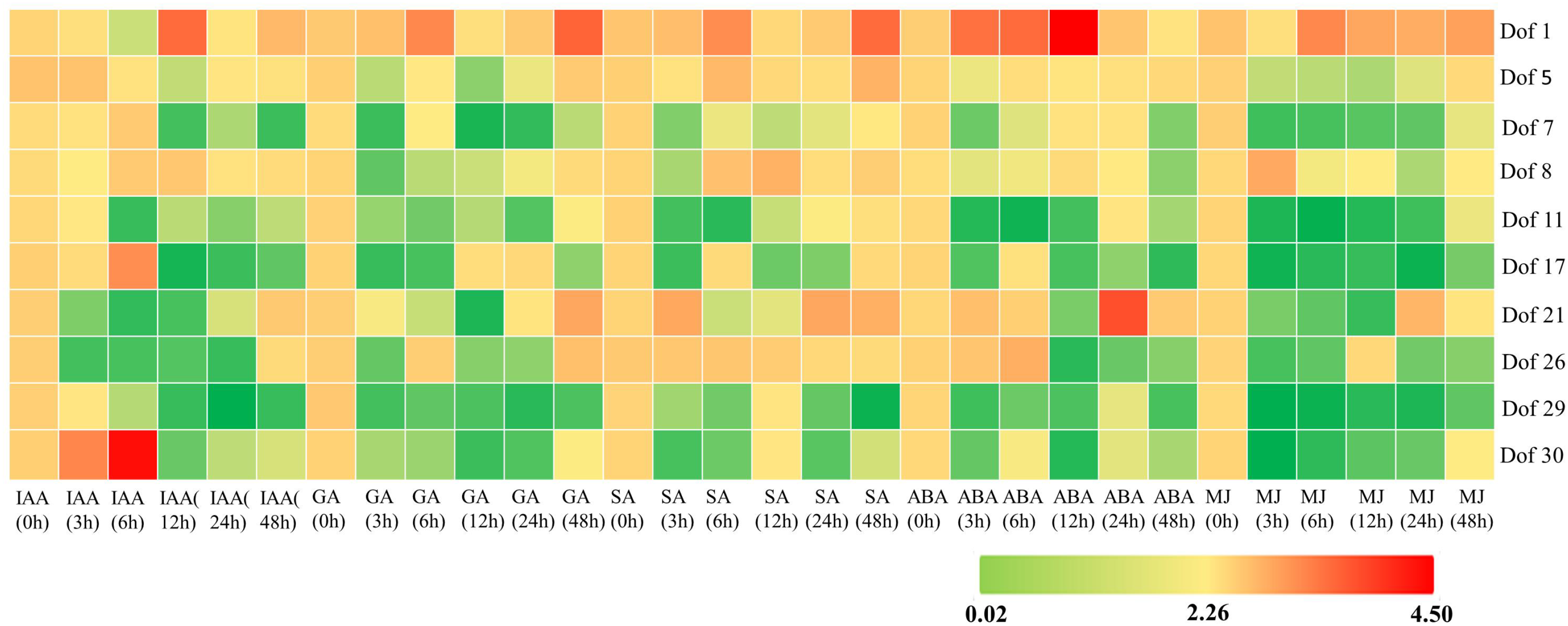
Expression analysis of blueberry Dof genes under hormone treatment. Red or green indicates higher or lower expression levels in each sample, respectively.
The expression heatmap indicates that blueberry leaves exhibited more upregulated genes after 0–6 h of IAA treatment than after 12–48 h treatment. Similarly, blueberry leaves showed more upregulated genes after 6–12 h of SA treatment than after 24–48 h treatment. Blueberry leaves exhibited more upregulated genes after 48 h of GA treatment than after 0–24 h of treatment. Blueberry leaves primarily respond to hormone treatments by inducing gene expression, indicating regulation during the early stages of exogenous IAA and SA treatments and the late stages of exogenous GA treatment, with distinct roles at different time points.
Discussion
Blueberry Dof family member analysis
Research on blueberry gene families has primarily focused on flower and fruit development and abiotic stress responses, but studies on hormone regulation within the blueberry Dof gene family remain unexplored. This study identified 32 Dof genes in the blueberry genome, fewer than those in camelina (103 CsDof genes, 35 and wheat (108 TaDof genes,, 36 both of which have undergone recent whole-genome duplication events that likely contributed to Dof family expansion. The number of Dof TFs in blueberry is comparable to that in rice (30 OsDof genes,, 37 tomato (34 SlDof genes,, 15 and pepper (33 CaDof genes,, 38 but higher than in durian (24 DzDof genes, 39 and eggplant (29 SmeDof genes,. 40 These numerical differences may reflect variations in genome size, ploidy level, and lineage-specific gene duplication or loss events. The relatively moderate number of Dof genes in blueberry is consistent with its diploid genome and the absence of recent large-scale duplication events, suggesting that the blueberry Dof family has remained relatively stable during evolution. Within the Vaccinium genus, a recent study identified 51 Dof genes in V. corymbosum, 30 substantially more than the 32 genes identified here in V. darrowii. This difference may be attributed to the fact that V. corymbosum is a tetraploid species, whereas V. darrowii is diploid (2n = 2x = 24), and polyploidization is often accompanied by gene family expansion. Additionally, Li et al. 30 reported that whole-genome duplication events contributed to the expansion of the VcDof gene family in V. corymbosum, a pattern also observed in other polyploid species such as wheat and camelina. The smaller Dof repertoire in V. darrowii is therefore consistent with its diploid genomic architecture and suggests a comparatively stable evolutionary history for this gene family in this species. The subcellular localization of blueberry Dof proteins is predominantly nuclear, with Dof6 localized to chloroplasts and Dof22 and Dof31 localized to peroxisomes. Protein localization correlates with function, as many transcription factors regulating gene expression localize to the nucleus. For example, Wang et al., 41 Wang et al. 42 found that green fluorescence was exclusively distributed in the nucleus upon transient transformation with the 35S::ThDof8-GFP plasmid, indicating that the ThDof8 protein from Tamarix hispida is localized to the nucleus. Overexpression of the ThDof8 gene reduces oxidative damage and increases levels of protective enzymes and proline, thereby enhancing the salt tolerance of Tamarix hispida. Ding and colleagues demonstrated that the wheat Dof family gene TaDof6 possesses transcriptional activation activity, with subcellular localization studies confirming its nuclear localization. Overexpression of this gene in wheat grains significantly increased amylopectin content while enlarging starch granule size and enhancing relative crystallinity. 43 Multiple sequence alignment results (Figure 4) reveal that all blueberry Dof transcription factors possess a fixed C2-C2 conserved domain. Gene structure and cis-acting element analyses indicate that blueberry Dof genes within the same subfamily exhibit similar exon/intron structures and cis-element sequences, demonstrating high conservation among blueberry Dof transcription factors. 15 Furthermore, the anomalous size and intron number of Dof17 may result from LTR retrotransposon insertion into its intron region, accumulated and preserved under specific evolutionary history and weaker purifying selection pressure. Trans-acting elements indicate that most blueberry Dof promoters contain hormone, MYB, MYC, Box 4, and secondary metabolism response elements, suggesting functional attributes related to hormones, stress, light, and metabolism for blueberry Dof family genes. Identification of the Dof gene family in wild olive (Olea europaea var. sylvestris v1.0) revealed the presence of promoter elements specific to light, circadian rhythm, auxin, and ABA responses, consistent with the present study. This suggests that Dof proteins may exhibit a degree of sequence conservation across different species, thereby maintaining their biological activities and functional roles. 44
Evolutionary development, duplication events, and colinearity analysis of Dof genes in the blueberry system
Using MegaX, a phylogenetic tree of Dof transcription factors in blueberry, Arabidopsis, and grape was constructed. The results indicate that all Dof transcription factors are grouped into six MCOG families (I, II, III, IV, V, VI), with blueberry Dof genes being underrepresented in the MCOG II subfamily. This aligns with Da et al.'s findings, who reported the lowest number of Dof genes in MCOG II, with only one grape VviDofL gene present (Da et al., 2016). This subfamily likely governs a highly conserved, non-redundant core regulatory function. The Arabidopsis AT1G29160.1 (ATDof1.5) gene participates in regulating light signaling pathways. However, since genes in Subfamily III containing light response elements are present in the phylogenetic tree, it is predicted that this gene may be involved in blueberry photoperiod and abiotic stress regulation processes (Corrales et al., 2014). The AT2G46590.1 (ATDof2.5) gene functions in seed germination. Blueberry Dof28 and Dof3 cluster with it, suggesting potential involvement in red light and seed development regulation. 45 Furthermore, Dof transcription factors from the three species did not show distinct separation in the phylogenetic tree, indicating that Dof transcription factors have not undergone significant differentiation and may share similar functions.
We utilized the MCScanX tool to analyze the homology relationships among members of this gene family based on whole-genome alignment results. We found that gene pairs located on chr3 (Dof5 and Dof6), chr6 (Dof13 and Dof14, Dof15 and Dof16), and chr12 (Dof30 and Dof31) share sequence similarity and reside within tandem repeat blocks, indicating they resulted from a single segmental duplication event.
To investigate the evolutionary conservation of this gene family, we constructed a collinearity map within the blueberry species. The results revealed significant collinearity among gene clusters located on chr1, chr6, chr9, chr11, and chr12, suggesting they did not originate independently but rather resulted from one or multiple genomic segment duplication events. These duplication events represent key evolutionary drivers of the Dof gene family's expansion and diversification. Despite undergoing chromosomal rearrangements and sequence divergence, the conservation of synteny among these genes strongly suggests that the internal structure of their gene clusters may be crucial for maintaining appropriate gene expression or synergistic functions. Future studies on the expression patterns and functions of each copy will help reveal how they promote species adaptability through subfunctionalization or neofunctionalization.
Analysis of Dof genes involved in hormone response
Analysis of gene expression patterns can provide insights into gene function. Hormone-responsive expression analysis in this study revealed that blueberry Dof1, Dof7, Dof17, Dof30, Dof21, and Dof26 exhibited significantly higher relative expression levels under short-term exogenous IAA and GA induction compared to the control group, showing an upward trend in expression levels during the early stage. However, the relative expression levels of blueberry Dof1, Dof26, Dof21, Dof8, and Dof5 were significantly lower than the control group under short-term induction by ABA, MeJA, and SA (Figure 10). This aligns with previous findings that the Brassica napus bnaaDof gene plays a potential role in regulating plant hormone responses, facilitating the transition from the vegetative to reproductive stage. 46 ABA treatment predominantly downregulated expression of BvDofs genes in sugar beet. 47 Exogenous application of GA and MeJA both induced downregulation of AktDof genes in trifoliate orange. 48 Apple MDof exhibits high sensitivity to abiotic stress, consistent with blueberry Vcdof1, Vcdof11, and Vcdof15.31,49 However, it should be noted that the inferred regulatory roles of these candidate Dof genes in hormone biosynthesis are based on correlative expression data and cis-element analysis. Functional validation through transgenic overexpression, gene silencing, or mutant analysis will be necessary to confirm their direct involvement in specific hormone biosynthetic pathways.
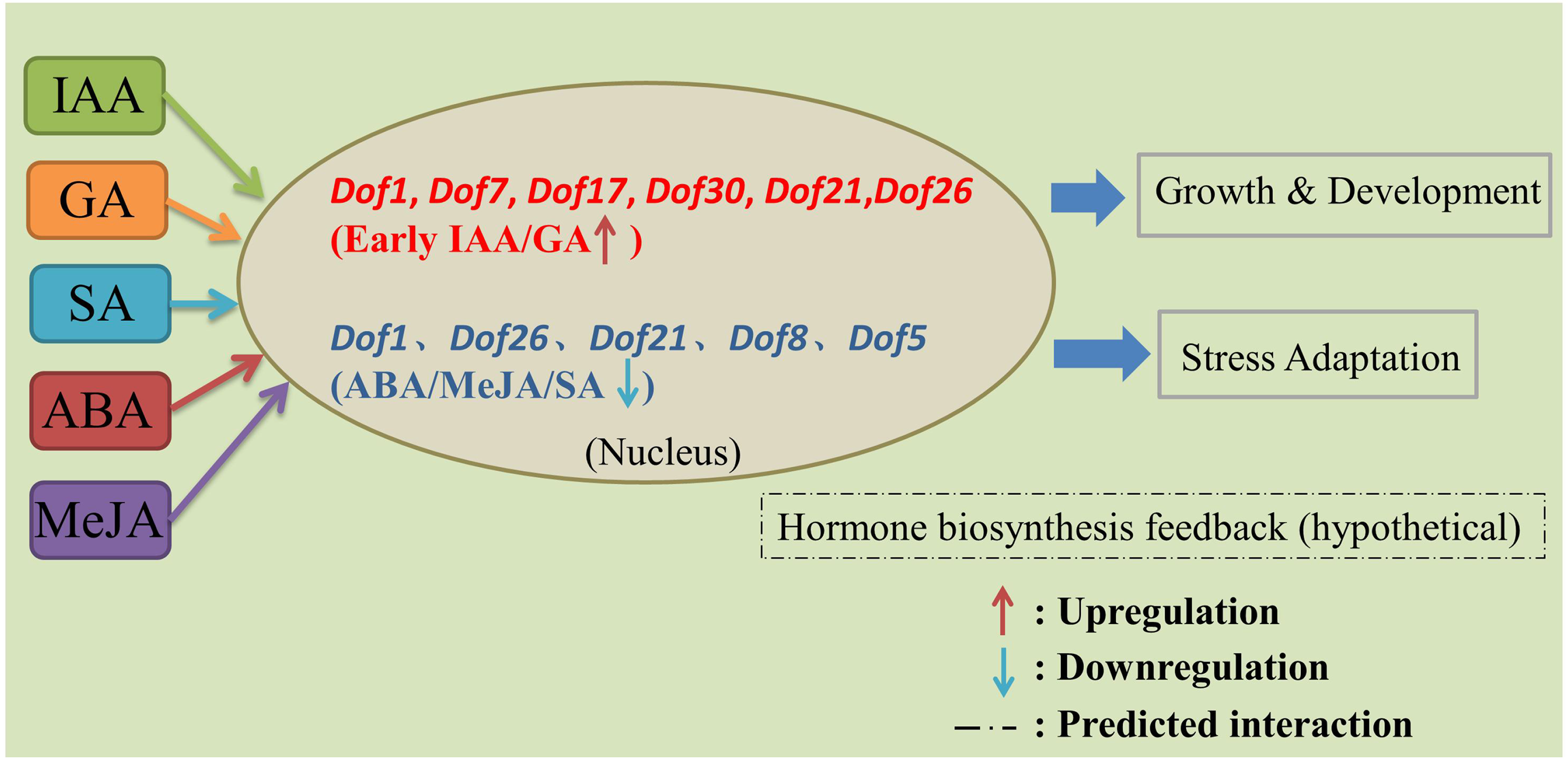
A proposed model of Dof-mediated hormone signaling in blueberry. IAA and GA induce early expression of six Dof genes (red), while ABA, MeJA, and SA suppress five Dof genes (blue). The Dof proteins are predicted to localize to the nucleus and may regulate hormone-responsive pathways. Dashed lines denote hypothetical interactions. This model is based on correlative data and requires functional validation.
Conclusion
This study identified 32 members of the blueberry Dofs gene family, with 31 distributed across 12 chromosomes. Based on physicochemical properties, blueberry Dof family members exhibit pI values leaning toward alkaline, protein hydrophilicity values below zero, and lipid coefficients below 100, further indicating strong hydrophilicity. Most Dof proteins are localized in the cell nucleus. Analysis of sequence alignment, phylogenetic relationships, protein motif distribution, and gene structure revealed that the blueberry Dof domain (Motif 1) is relatively conserved, featuring a fixed C2-C2 structure. Dof genes clustered into six major groups (I-VI), with closely related genes exhibiting similar protein motifs. Motif 1 is uniformly distributed as a typical Dof domain, while others are more dispersed and variable but conserved within clades. Motif 4 is a distinctive domain within Cluster III. Motifs 2, 5, and 8 are unique to Cluster III, while Motif 7 is present only in Clusters III and V, and Motif 10 is found exclusively in Clusters II and V. Further, analysis of cis-acting elements and response expression patterns revealed that most Dof promoter sequences contain hormone- and stress-related elements. Ten Dof promoters were identified as responsive to five hormones. RT-qPCR expression pattern analysis identified six Dof candidate genes positively regulating IAA and GA hormone synthesis and five positively regulating ABA, SA, and MeJA hormone synthesis. This provides a theoretical foundation and gene resources for studying the role of the Dof gene family in blueberry growth, development, and hormone responses.
Supplemental Material
sj-docx-1-ber-10.1177_18785093261466595 - Supplemental material for Genome-wide identification of the Dofs family and analysis of the expression pattern under different hormone treatments in blueberry
Supplemental material, sj-docx-1-ber-10.1177_18785093261466595 for Genome-wide identification of the Dofs family and analysis of the expression pattern under different hormone treatments in blueberry by Xiaochen Tu, Jilu Che, Yaqiong Wu, Zhengjin Huang, Wenlong Wu, Lianfei Lyu, Donglu Fang, Fuliang Cao, Weilin Li and Yicheng Wang in Journal of Berry Research
Footnotes
Author contributions
Yaqiong Wu and Donglu Fang designed the research. Xiaochen Tu, Jilu Che and Zhengjin Huang analyzed the data. Wenlong Wu and Lianfei Lyu provided the plant materials. Xiaochen Tu and Yaqiong Wu wrote the article. Fuliang Cao, Yicheng Wang and Weilin Li reviewed the article.
Funding
This research was supported by the “JBGS” Project of Seed Industry Revitalization in Jiangsu Province (JBGS[2021]021), and the Postgraduate Research and Practice Innovation Program of Jiangsu Province [SJCX24_0352 ] and Ruihua Nonprofit Foundation.
Role of the funding source
The funding bodies had no role in the design of the study; the collection, analysis, and interpretation of data; or the writing of the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The data supporting the conclusions of this study are included within the article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.