
Abstract
This study was undertaken to test the hypothesis that altered podocyte slit protein nephrin distribution is associated with disturbed polarity protein expressions in podocytes from preeclampsia (PE). We examined expressions and distributions of nephrin, podoplanin, polarity protein partitioning defective-3 (PARD-3), and PARD-6 in podocytes from PE. Podocyte cell line (AB 8/13 cells) was used as control. Podocytes were found in all severe PE cases. In contrast, no podocyte was found in the samples from normal pregnancies and mild PE. Compared to control cells, nephrin, PARD-3 and PARD-6 expressions were reduced or lost in podocytes from severe PE. Podoplanin was expressed in podocyte surface membrane on control cells but reduced in podocytes from PE. These findings indicate that loss of slit protein nephrin and polarity protein PARD-3 and PARD-6 on foot processes could explain for podocyte detachment from glomerular basement membrane and lead to podocyte shedding in PE.
Introduction
Preeclampsia is a unique human pregnancy complication characterized by elevated blood pressure and newly-onset proteinuria after 20 weeks of gestation. The presence of protein in the urine signifies kidney injury in preeclampsia. 1 Protein leakage into urine is a consequence of renal barrier dysfunction/loss of integrity. There are at least 3 functional components of the renal barrier: glomerular endothelial cells, glomerular basement membrane, and podocytes. In preeclampsia, the glomerular endothelial cells display swelling and hypertrophy, collectively referred to as endotheliosis. 1 The glomerular endothelial swelling and hypertrophy render the glomeruli ischemic. At present, little information is available as to how the characteristic endotheliosis might contribute to the renal barrier dysfunction in preeclampsia. Interestingly, recent clinical studies highlight the injury of podocytes in this pregnancy disorder. For example, Garovic et al reported that podocytes were present in the urine of virtually all preeclamptic women that was examined. 2 In accordance with this, subsequent studies demonstrated reduction of podocyte slit protein nephrin expression in autopsy and biopsy renal materials from women with preeclampsia. 3,4 Thus, it is evident that the epithelial part of the glomerulus, that is, the podocytes, also plays an indispensable role in the pathophysiology of kidney injury in preeclampsia.
Podocytes anchor to the glomerular basement membrane through their foot processes while the main cell body is floating within Bowman’s capsule. The foot processes interdigitate to form a tight meshwork called filtration slit that covers the outermost surface of the glomerular basement membrane. The filtration slit is bridged by a slit diaphragm. Podocytes are shed from the glomerular basement membrane due to dysfunction/loss of integrity of this anchorage machinery, as seen in focal segmental glomerulonephritis and lupus nephritis. 5 We and others have demonstrated that there is a broad spectrum of molecular alterations in podocytes from preeclampsia, including slit-associated protein nephrin, podocyte surface protein GLEPP-1, cytoskeleton marker synaptopodin, and actin-linker protein ezrin 3,4 ). However, little is known of the phenotypes of shed podocytes in preeclampsia. Podocytes are polarized cells and polarity protein expressions have never been studied in shed podocytes from preeclampsia. We proposed that altered podocyte slit protein nephrin distribution is associated with disturbed polarity protein expressions in podocytes from preeclampsia. To test this hypothesis, we extracted and cultured podocytes from urinary samples from women with preeclampsia and specifically examined expressions and distributions of slit protein nephrin, apical surface protein podoplanin, and polarity protein partitioning defective-3 (PARD-3) and PARD-6 in podocytes from preeclampsia. Podocyte cell line (AB 8/13 cells) was used as control.
Materials and methods
Urine sample collection
The study was approved by the Institutional Review Board (IRB) for human research at Louisiana State University Health and Sciences Center – Shreveport (LSUHSC-Sh), Louisiana, and conducted in the Department of Obstetrics and Gynecology, LSUHSC-Sh. Women with normal and preeclamptic pregnancies were enrolled in this study when they were admitted to the Labor and Delivery Unit. Normal pregnancy was defined as uneventful pregnancy with blood pressure <140/90 mm Hg and no proteinuria. Mild preeclampsia was defined as persistent elevated blood pressure over 140/90 mm Hg on 2 separate readings and/or proteinuria of 1+ or more on dipstick or urine protein ≥ 300 mg/24 hours. Severe preeclampsia was defined as persistent elevated blood pressure over 160/110 mm Hg on 2 separate readings and urine protein ≥2 g/24 hours. No patient had a sign of infection. Pregnant women with nephrotic syndrome were also recruited for the study. Nephrotic syndrome was defined as urine total protein excretion greater than 3.5 g/24 hours, low serum albumin level, high serum cholesterol level, and peripheral edema with maternal blood pressure ≤140/90 mm Hg. Urine specimen was collected in a strictly sterile manner after consent was obtained. Smokers and pregnancy complicated by either chronic hypertension or diabetes were excluded from the study. A total of 33 urine samples were used in this study. Demographic data from which the urine sample was used in the study are summarized in Table 1 .
Demographic Characteristics of Pregnant Women From Which Urine was Used in the Study a
a Data are expressed as mean ± SD.
Podocyte isolation and cultivation
Freshly obtained urine specimen, about 100 mL with 50 mL in each tube were centrifuged at 1500 rpm at 4°C for 5 minutes. Cell pellets were resuspended and washed with medium RPMI 1640 (GIBCO, Carlsbad, California). Cell pellets were obtained again by centrifugation. Cell pellets were resuspended by RPMI 1640 supplemented with 10% fetal bovine serum (FBS) and antibiotic-antimycotic solution (Sigma, St. Louis, Missouri) and ITS liquid media supplement containing insulin at 10 mg/mL, transferrin at 5.5 mg/mL, and selenium at 5 ng/mL (Sigma) as previously described. 6 Cell suspension from one 50-mL aliquot was seeded into a 24-well/plate on fibronectin-coated cover slips for cell morphology study. Cell suspension from the other 50 mL aliquot was seeded into a 6-well/plate for cell growth monitoring. Plates were then placed in a 37°C cell culture incubator with 5%CO2 and air. After overnight incubation, the plates were washed with phosphate buffered saline (PBS) twice and fresh RPMI 1640 was replaced. The medium was changed every other day. Cells grown on cover slips were used for immunofluorescent staining (see below). Cells seeded in a 6-well/plate were monitored for cell colony formation as an indicator of relative cell growth. This was done by counting cell colony number under a phase-contrast microscope (Olympus CK40, Japan) on day 3, day 6, and day 10 after seeding. Cell clone was defined as a cell aggregate containing at least 5 cells per clone and recorded for quantitative and statistical analysis.
Human podocyte cell line (AB 8/13) culture
AB 8/13 cell line is an immortalized human podocyte cell line. This conditionally immortalized human podocyte cell line was developed by transfection with the temperature-sensitive SV40-T gene and a telomerase gene into a primary isolated normal human podocytes. 6 Transfected cells proliferate at the “permissive” temperature of 33°C. After transfer to the “non-permissive” temperature of 37°C, they enter growth arrest and express markers of differentiated in vivo podocytes, including the novel podocyte proteins of nephrin, podocin, CD2AP, and synaptopodin, and other molecules known to be located at slit diaphragm. 6 Proliferation of these cells under 33°C is driven by temperature-sensitive SV-40 Large-T antigen, but under 37°C the antigen is inactivated. Cells stop proliferating and complete differentiating within 10 to 14 days. 6
AB 8/13 podocytes were cultured with RPMI-1640 medium supplemented with 10% FBS and ITS liquid media supplement as previously described. 6 Medium was changed every other day. Cells incubated at 37°C for 10 to 14 days were stained with antibodies against nephrin, podoplanin, PARD-3, and PARD-6, which served for comparison to podocytes from women with preeclampsia.
Protein expression of SV-40 Large-T antigen was also examined by Western blot in AB 8/13 cells that were cultured under 37°C to ensure SV-40 Large-T antigen inactivation. An aliquot of 10 µg total cellular protein was used to run sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) under reducing condition. A standard Western blot procedure was performed. Specific antibody against SV-40 Large-T antigen was purchased from Santa Cruz Technology, Inc. (San Diego, California).
Immunofluorescent Staining
For immunofluorescent staining, cells grown on cover slips were fixed with 2% paraformaldehyde in 4% sucrose for 8 minutes at room temperature followed by washing cover slips 3 times with 1% bovine serum albumin (BSA), 5 minutes each. After blocking with 1% BSA and 1% glycine in PBS for 1 hour at room temperature, cells were incubated with specific antibodies that against human nephrin (Abcam, Cambridge, MA), podoplanin, PARD-3, and PARD-6 at 4°C overnight. Antibodies against podoplanin, PARD-3, and PARD-6 were purchased from Santa Cruz. After washing with 1%BSA, the cover slips were incubated with Cy 3-conjugated (Jackson Immunotech Lab, Westgrove, Pennsylvania) or Alex488-conjugated (Molecular Probe/Invitrogen, Carlsbad, California) secondary antibody. After staining, coverslips were mounted on glass slides with Vectashield Mounting Medium with DAPI (Vector Lab Inc, Burlingame, California). The slides were reviewed under a fluorescent microscope (Olympus IX-71, Japan). Images were captured with a digital camera linked to a PC computer with imaging software PictureFrame (Uptronics Inc. Sunnyvale, California). For Z-stacking, images were captured by Apotome Observer (Carl Zeiss, Inc. Germany) and were reconstructed with Axiovision software (Carl Zeiss).
Nephrin Expression
Nephrin expression was examined by Western blot in shed podocytes from 2 nephrotic syndrome and 2 patients with severe preeclampsia. Total cellular protein was extracted with ice-cold protein lysis buffer containing 50 mmol/L Tris, 0.5% NP40, and 0.5% Triton X-100 with protease inhibitors of PMSF, DTT, leupeptein, and aprotini. An aliquot of total cellular protein (10 µg of each sample) was subject to electrophoresis (Bio-Rad, Hercules, California) and then transferred to nitrocellulose membranes. The membranes were then probed with nephrin antibody (R&D, Minneapolis, Minnesota). The bound antibody was visualized with an enhanced chemiluminescent (ECL) detection Kit (Amersham Corp, Arlington Heights, Illinois).
Statistical Analysis
Podocyte colony numbers were analyzed by the computer software program StatView (SAS Institute Inc., Cary, North Carolina). Data are presented as mean ± SE. A probability level of <.05 is considered statistically significant.
Results
Podocytes Are Shed From Pregnant Women Complicated With Severe Preeclampsia and Nephrotic Syndrome
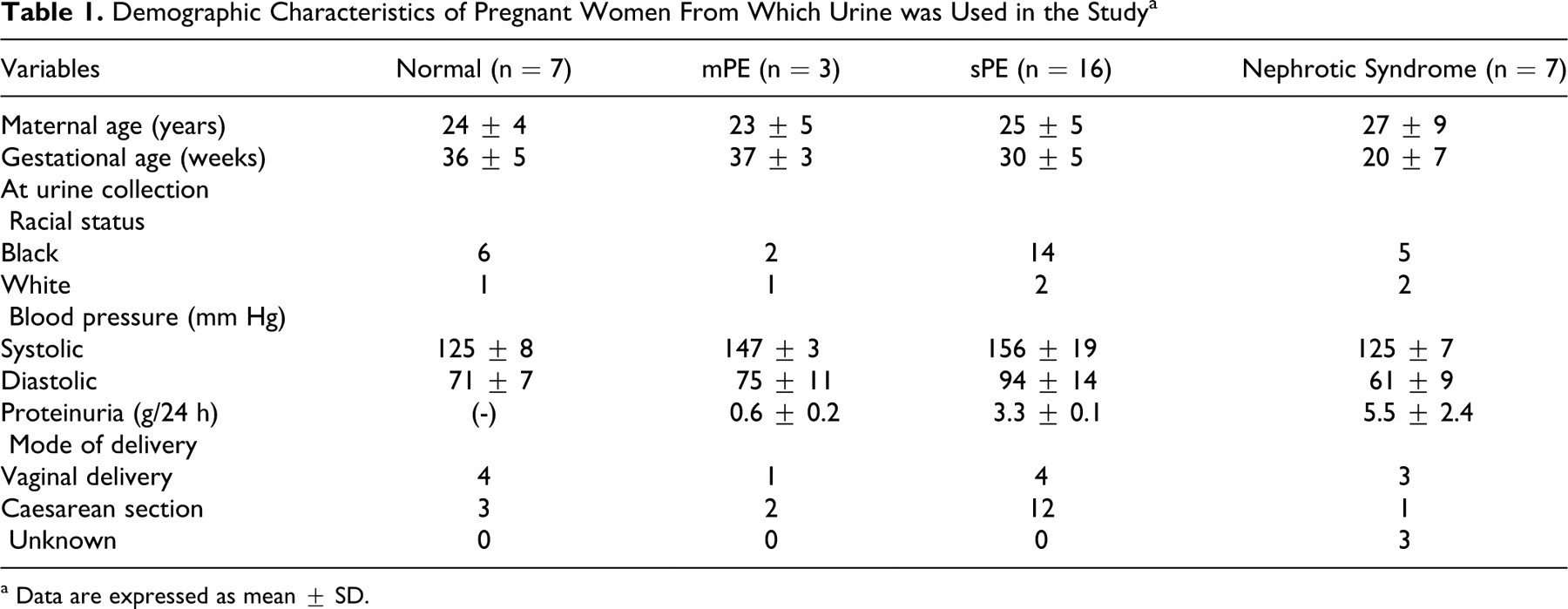
Podocytes were extracted from 33 urine specimens, 7 from normal term pregnant women, 3 from mild preeclampsia, 16 from severe preeclampsia, and 7 from nephrotic syndrome. Among these urine specimens, viable podocytes were found in all cases with severe preeclampsia and nephrotic syndrome. In contrast, podocytes were not seen in urine specimens from women with normal pregnancy or mild preeclampsia. These urine podocytes exhibited a similar morphology to AB 8/13 podocytes, that is, cobble-stone appearance as the proliferative phenotype and arborized morphology as the differentiated phenotype. 6 Both proliferative and differentiated podocytes were noticed in urine podocytes shed in severe preeclampsia and nephrotic syndrome. However, most of the shed podocytes were in the proliferative stage and presented as the cobblestone appearance. Cell clones were formed within a week after seeding as shown in Figure 1A . These cells were capable of proliferating in vitro and can be subcultured. After subculture, these cells gradually became differentiated phenotype revealed as arborized morphology, as shown in Figure 1B. Cells with arborized morphology were relatively larger with octopus or oval shapes. These cells were unable to proliferate. Shed podocytes, both proliferative and differentiated, expressed podocyte slit specific protein nephrin (see below).
Morphology of shed urine podocytes from preeclampsia in comparison to AB 8/13 podocytes. The shed urine podocytes present cobble-stone appearance when they were in active proliferative stage (<10 days) (A), which was similar to AB 8/13 podocytes cultured under 33°C (B). After prolonged culture (>20 days), the shed urine podocytes presented oval shape with the cell size (C) that was similar to the differentiated AB 8/13 podocytes cultured under 37°C (D). A and C: Bar = 200 micron; and B and D: bar = 50 micron, respectively.
Figure 1 shows the cell morphology comparison between the shed podocytes and AB 8/13 control podocytes. Under the permissive temperature (33°C), AB 8/13 podocytes present cobblestone morphology (Figure 1C). After switching to the nonpermissive temperature (37°C), these cells become enlarged and present arborized morphology, losing proliferative potential. Figure 1B and D show a representative higher magnification of differential morphology of shed podocytes and AB 8/13 podocytes, respectively.
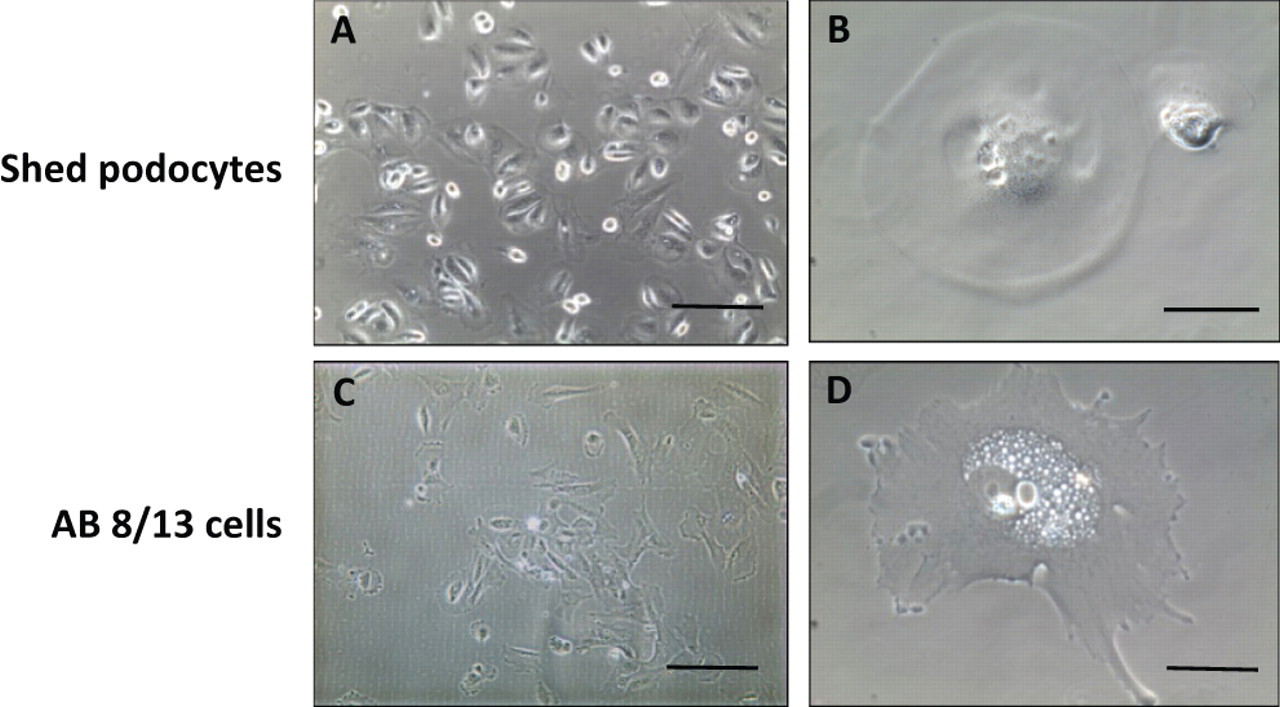
To observe shed podocyte growth, podocyte clone formation was monitored and clone number was counted on days 3, 6, and 10 under a phase-contrast microscope. Figure 2 upper panel shows an example of shed podocytes grown in culture on days 3, 6, and 10. Figure 2 lower panel shows clone numbers of shed podocytes on days 3, 6 and 10 from each group. There were no podocytes in urine samples from normal pregnancies and from mild preeclampsia, but podocytes were found in urine specimens from all severe preeclampsia cases and nephrotic syndrome cases. Interestingly, more podocytes were shed from patients with nephrotic syndrome than from severe preeclampsia, supporting the idea that the degree of podocyte injury is even worse in pregnant women complicated with nephrotic syndrome. In some cases, shed podocytes were subcultured up to 3 passages in vitro in our culture conditions.
Shed urine podocytes grown in vitro. Upper panel shows a representative image series of one clone of shed urine podocytes from severe preeclampsia when cultured in vitro. On day 3, cell clone formation was evident and counted (A). More cells were grown within the same cell clone on day 6 (B) and on day 10 (C), indicating proliferative potential of shed podocytes in vitro. Lower panel shows the number of podocyte clone formation counted on days 3, 6, and 10 after seeding. Cell pellet from 50 mL urinary specimen were seeded into a 6-well/plate. No podocyte clone formation was found in urinary samples from normal and mild preeclamptic pregnant women. In contrast, more podocyte clone formation was found in shed podocytes from pregnant patients complicated with nephrotic syndrome than from severe preeclampsia.
Nephrin Is Strongly Expressed in Differentiated Podocytes
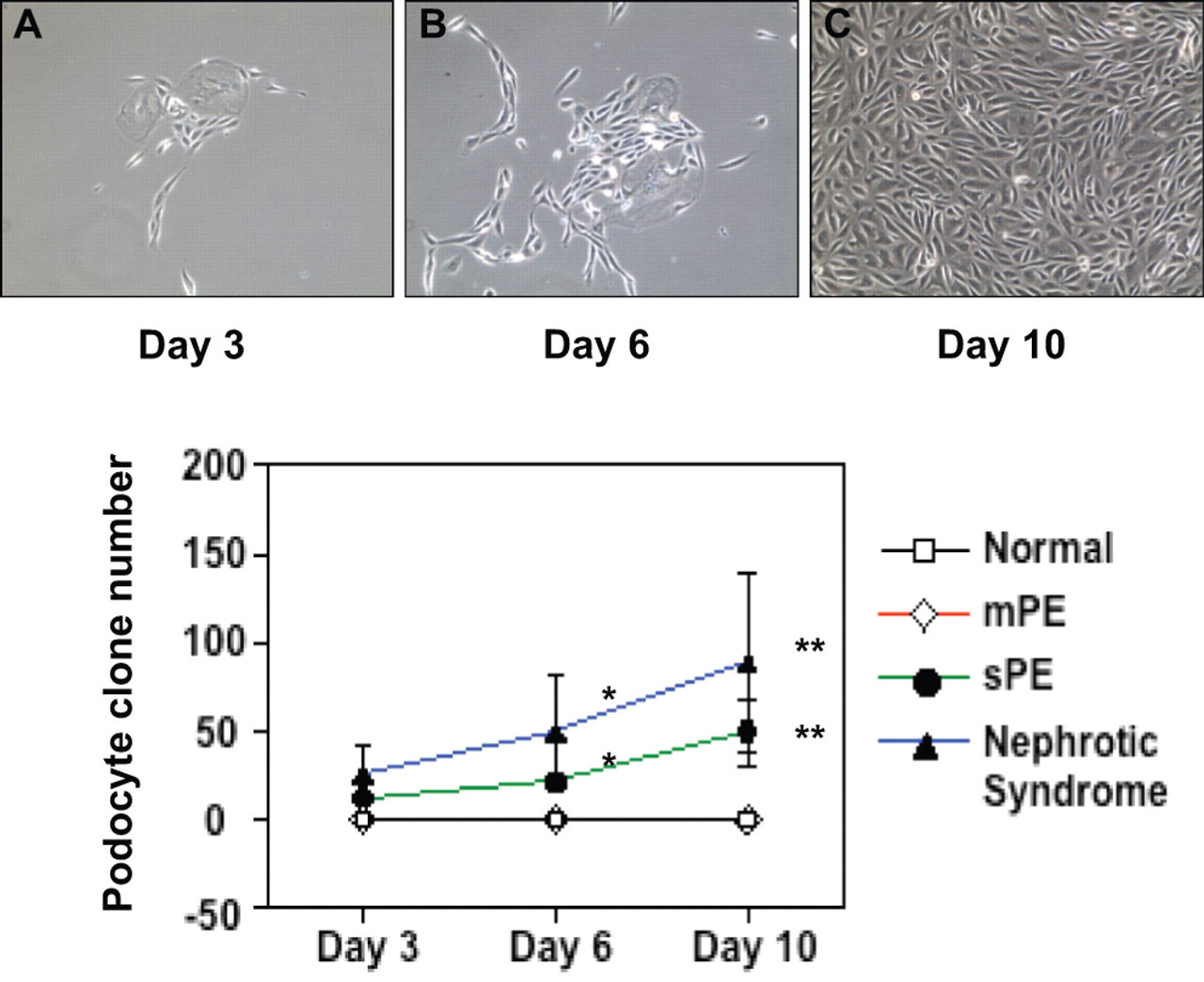
Because podocyte was negative in urine samples from normal pregnant women, we used AB 8/13 podocytes as control to determine whether altered distributions of specific podocyte protein nephrin and podoplanin were associated with changes of polarity protein PARD-3 and PARD-6 expressions in podocytes from preeclampsia. In each culture set, AB 8/13 cells were cultured at 37°C for more than 10 days after they were switched from 33°C. Inactivation of SV-40 antigen was confirmed by Western blot before cells were stained with nephrin, podoplanin, PARD-3, and PARD-6. As shown in Figure 3 , SV-40 is strongly expressed in AB 8/13 cells cultured under 33°C, the permissive temperature, whereas SV-40 expression becomes undetectable 3 days after switching the cells from 33°C to the nonpermissive temperature at 37°C. In contrast, increased nephrin expression was observed in AB 8/13 cells after switching to the nonpermissive temperature. These results confirm that AB 8/13 cells differentiate when cultured under 37°C and differentiated cells strongly express podocyte specific slit protein nephrin.
Nephrin expression is increased when AB 8/13 podocytes grown at 37°C in vitro. AB 8/13 podocytes were proliferative when they were cultured under permissive temperature at 33°C and differentiated when cultured under 37°C. SV-40 was strongly expressed when AB 8/13 podocytes were cultured at 33°C. SV-40 expression was not detectable when cells were cultured under 37°C, indicating SV-40 was inactivated. In contrast, nephrin expression is increased steadily after thermoswitching.
Altered Distribution and Expression of Podocyte-specific Protein Nephrin and Podoplanin, and Polarity Protein PARD-3 and PARD-6 in Podocytes from Preeclampsia
Using differentiated AB 8/13 cells as control, we examined podocyte-specific protein nephrin and podoplanin, and polarity protein PARD-3 and PARD-6 distribution and expression by immunofluorescent staining of urine podocytes from severe preeclampsia. Representative of 2-D and 3-D images are shown in Figure 4 . Nephrin distribution and expression: on control cells (Figure 4A series), nephrin immunoreactions are mainly localized and evenly distributed at the cell periphery in a punctuated pattern. On shed urine podocytes (Figure 4B series), staining signal was unevenly distributed at the cell periphery and also revealed as a diffused pattern in the cytoplasm. Altered nephrin distribution is more pronounced when Z-stacking was used to reconstruct the 3-D image. Nephrin is primarily located at the basal side of the cell (Figure 4A-3), while on the urine podocytes, the peripheral immunoreaction exhibited as a linear pattern. More importantly, nephrin seems to shift away from the basal side of the cell (Figure 4B-3). Podoplanin distribution and expression: on control cells, podoplanin is expressed throughout the cell surface observed on both 2-D and 3-D reconstructed images (Figure 4C series). On shed urine podocytes, podoplanin expression is greatly reduced on the cell surface (Figure 4D series). PARD-3 distribution and expression: on control podocytes, PARD-3 is localized at the cell periphery (Figure 4E series), a pattern similar to that of nephrin. The similarity is confirmed by the 3-D reconstruction of the Z-stacking images (Figure 4E-3). While on shed urine podocytes, PARD-3 reactivity is much less at the cell periphery, but exhibited as a diffused pattern on the cell surface (Figure 4F series). PARD-6 distribution and expression: PARD-6 is expressed at the cell periphery (Figure 4G series) in control cells. The basal and peripheral distribution of PARD-6 on control podocytes is more evident on 3-D reconstruction (Figure 4G-3). In contrast, PARD-6 exhibited as a diffused pattern on shed urine podocytes and lack of signal at the cell periphery or foot process region (Figure 4H series).
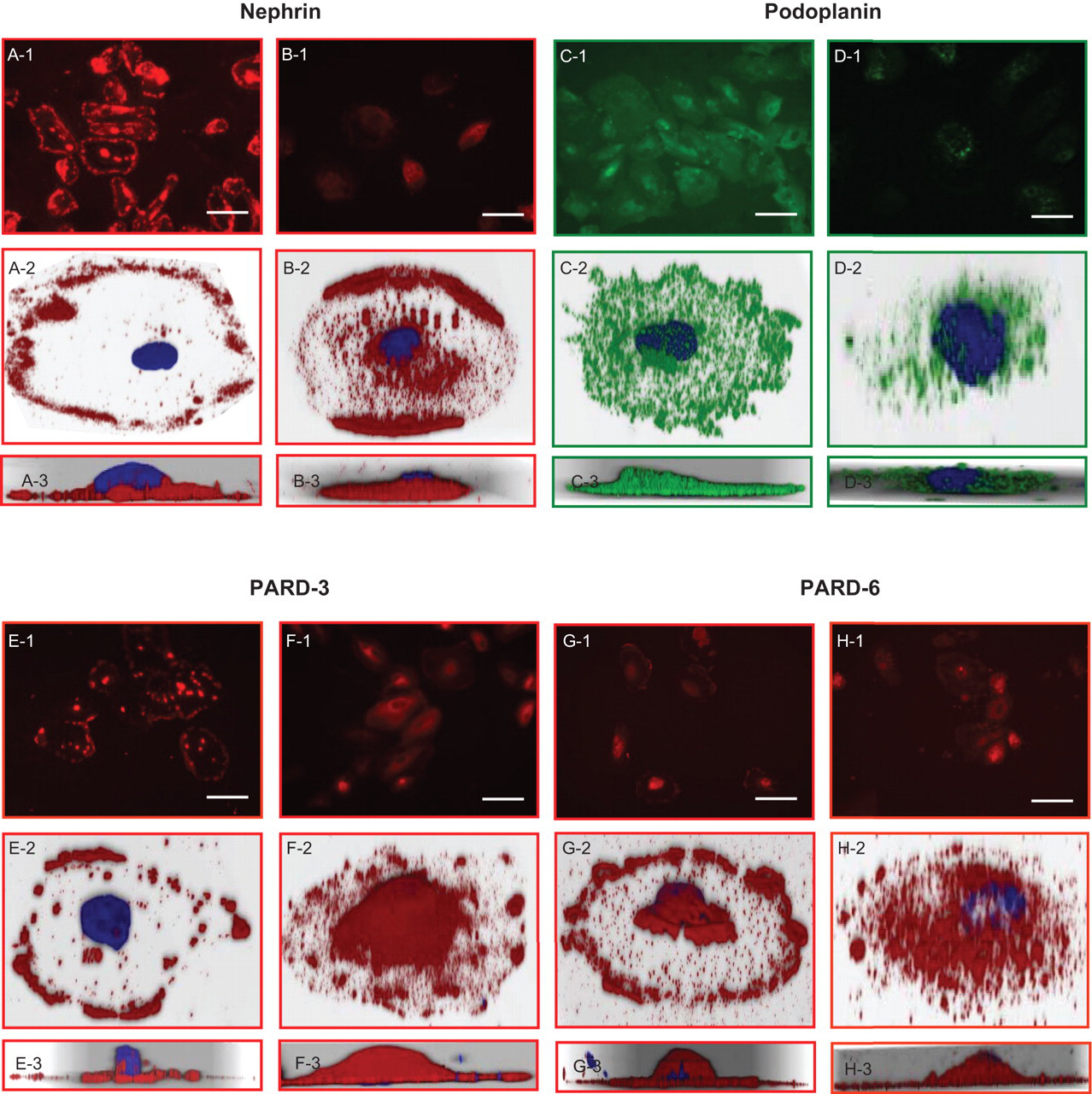
Expression and distribution of podocyte markers nephrin and podoplanin, and polarity proteins PARD-3 and PARD-6 on shed urine podocytes compared to AB 8/13 cells. Nephrin: A and B series; podoplanin: C and D series; PARD-3: E and D series; and PARD-6: G and H series. AB 8/13 podocytes: A, C, E and G. Shed podocytes from severe preeclampsia patients: B, D, F, and H. “1” series show low magnification of cells stained with each marker detected; “2” series show high magnification for stained cells in 3-D reconstruction; “3” series show Z-stacking of the 3-D reconstructed cells, respectively. On control podocytes (AB 8/13 cells), nephrin, PARD-3 and PARD-6 expressions are localized in peripheral regions (slit) and around cell borders, but expressions of these proteins were reduced and distribution pattern was markedly altered in shed urine podocytes from severe preeclampsia. Podoplanin is expressed evenly on the surface membrane on control podocytes (AB 8/13 cells), but reduced and/or lost in podocytes from severe preeclampsia, bar = 100 μM.
Compared to shed podocytes from preeclampsia, similar pattern for nephrin, podoplanin, PARD-3 and PARD-6 immunofluorescent staining, but very weak signal, was observed in shed podocytes from nephrotic syndrome (data not shown). We examined, nephrin expression by Western blot in shed podocytes from 2 nephrotic syndrome and 2 severe preeclampsia patients compared to control AB 8/13 cells. The result is shown in Figure 5 . Two major bands were detected in control cells, one around 180 kDa and one about 105 kDa. The 180 kDa band is the full-length nephrin protein and the 105 kDa band is likely a soluble form of nephrin. 7 The 180 kDa band was neither detected in shed podocytes from nephrotic syndrome nor from severe preeclampsia (Figure 5). The 105 kDa band was detected in podocyted from severe preeclampsia, but not the cells from nephrotic syndrome. However, a band approximately at 70 kDa was detected in shed podocytes from both nephrotic syndrome and severe preeclampsia.
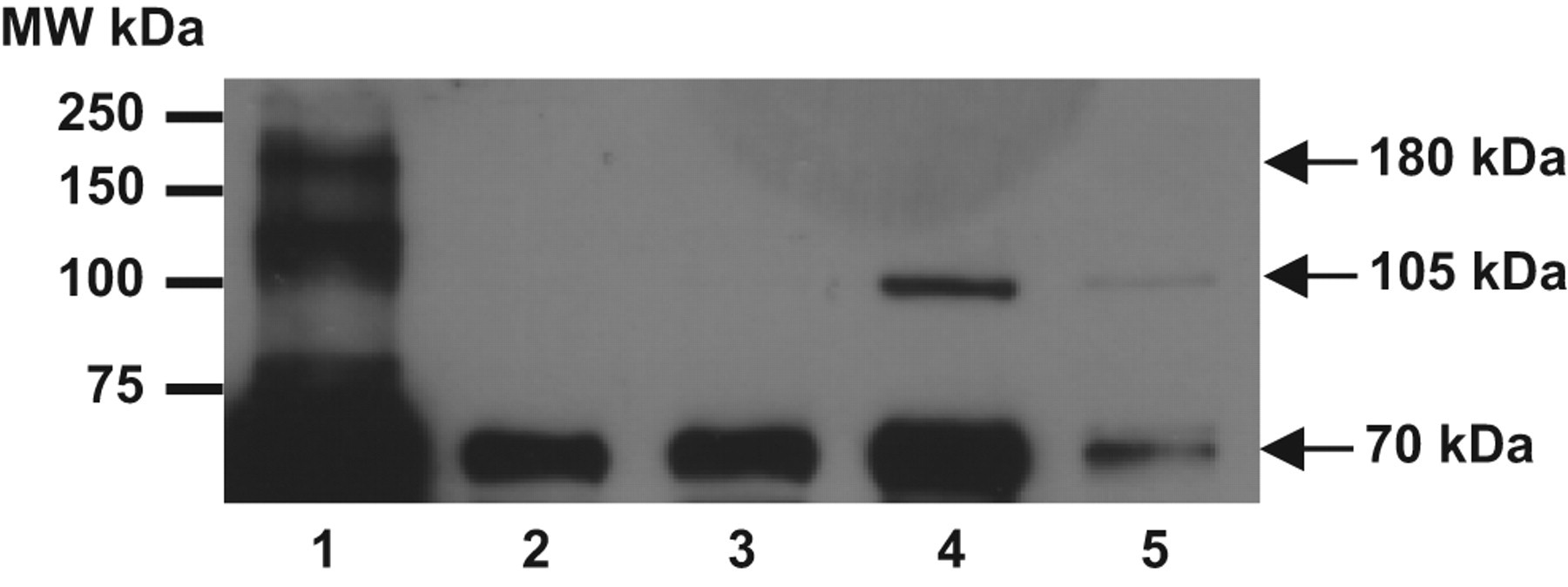
Nephrin expression by Western blot. Lane 1: differentiated AB 8/13 podocytes; lane 2-3: shed podocytes from nephrotic syndrome; and lane 4-5: shed podocytes from severe preeclampsia. Molecular weight marker is given on the left of the blot. Arrows on the right show band size (180 kDa, 105 kDa, and 70 kDa) detected in testing samples. Different molecular weight proteins were detected in shed podocytes from nephrotic syndrome and severe preeclampsia compared to the control differentiated AB 8/13 cells.
Discussion
In this study, we found that podocytes were shed from all severe preeclamptic cases. In contrast, no podocyte shedding was observed in women with normal pregnancy or mild preeclampsia. The shed cells in the urine can be cultured up to 3 passages in vitro. The identity of the shed cells as podocytes was confirmed by immunofluorescent staining of specific podocyte foot process slit protein nephrin and surface protein podoplanin. These findings support the notion that shed podocytes are viable, which indicates that the shed podocytes are neither necrotic nor apoptotic. In addition, podocyte shedding was also confirmed in pregnant women complicated by nephrotic syndrome. It is well accepted that podocytes are shed in certain renal diseases, including nephrotic syndrome. The cell morphology and immunofluorescent staining pattern of nephrin are similar in these 2 conditions based on our observation as well as that was described previously. 5
Since there is no podocyte shedding in normal pregnancy, we utilized the immortalized human normal podocytes cell line (AB 8/13 cells) as control to test our hypothesis of the phenotypic alteration of shed podocytes in preeclampsia. The AB 8/13 cells have been confirmed to represent the normal human podocyte phenotype in vitro and express podoplanin, nephrin, and many other molecules known to be located at the slit diaphragm. 6 Compared to the AB 8/13 cells, we found that shed podocytes from preeclampsia exhibited significant phenotypic changes, including nephrin, podoplanin, and polarity protein PARD-3 and PARD-6.
Nephrin is a specific podocyte foot process slit protein. In control cells, nephrin is expressed as a punctuated pattern localized on the cell periphery, corresponding to the place where foot processes are developed. On preeclampsia podocytes, nephrin is found as a linear pattern at the periphery and a diffused pattern throughout the cytoplasm, losing the typical punctuated pattern with reduced protein expression. Nephrin is a central component of the glomerular ultrafilter with both structural and signaling functions. 8,9 The extracellular domains of nephrin from adjacent podocytes form a meshwork acting as a possible size-selective barrier. 10 The short intracytoplasmic tail of nephrin contains tyrosine residue, which can be phosphorylated by Src kinase that initiates the intracellular transduction cascade. 11,12 Nephrin has also been demonstrated to interact with the cytoskeleton through diverse pathways/molecules including Nck, ILK, and CD2AP. 13,14 In congenital nephrotic syndrome, the nephrin gene is mutated and foot processes fail to develop. 15 In primary acquired nephrotic syndrome, nephrin redistribution has also been found. 7 These clinical observations strongly suggest the indispensable role of nephrin in the development and maintenance of renal barrier integrity. This notion is also supported by a mouse model for congenital nephrotic syndrome (NPHS1) where inactivation of nephrin gene causes massive proteinuria, absence of a slit diaphragm, and neonatal death. 16 Therefore, altered nephrin distribution found in podocytes from preeclampsia implicates that the integrity of podocyte slit diaphragm is disrupted in this pregnancy disorder.
Podoplanin is a negatively charged glycoprotein on the apical membrane domain in podocytes. 17 We found that podoplanin is evenly distributed on the cell surface and in the foot process area (Figure 4, C2 and C3). However, this pattern was lost and podoplanin expression is greatly reduced in podocytes shed from severe preeclampsia and pregnancy complicated with nephrotic syndrome. Since slit diaphragm of podocytes has been claimed as a signaling platform exemplified by nephrin, 9 it raises the question as to what the relationship is between signaling molecule nephrin at slit diaphragm and surface glycoprotein podoplanin in normal podocytes. Nonetheless, reduced negatively charged glycoprotein on podocytes would be expected to contribute to the increased protein leakage in preeclampsia.
It is well accepted that podocytes are differentiated polarized epithelial cells with 3 distinct domains: apical, basal, and slit diaphragm. However, little is known about the polarity signals required to regulate and maintain podocyte function and its foot process networks. In order to dissect the possible involvement of apical-basal polarity of podocytes in the phenotypic changes of shed podocytes in preeclampsia, we further examined the distribution and expression of polarity protein PARD-3 and PARD-6 on shed urine podocytes. As shown in Figure 4, we found both PARD-3 and PARD-6 exhibit a similar pattern as nephrin and are expressed at the peripheral and basolateral side in well differentiated AB 8/13 podocytes. While in shed podocytes, both PARD-3 and PARD-6 are reduced in the peripheral and basolateral region, but more signals were noticed on cell surface in shed podocytes. These observations clearly indicate that the cellular localization of PARD-3 and PARD-6 plays a critical role in maintaining the polarity and integrity of podocytes and their expressions are essential for the foot process slit diaphragm development. In fact, a study reported by Hartleben et al supports this notion. They found that PARD-3, PARD-6, and atypical protein kinase C (aPKC) polarity proteins are nephrin-associated proteins and colocalize at the slit diaphragm in HEK 293T cells transiently transfected with Neph1 and Nephrin gene. 19 They also showed that polarity protein PARD-3 and tight junction protein ZO-1 are colocalized during glomerular development in Wistar rats. 19 Thus, we have demonstrated that altered distribution of polarity protein PARD-3 and PARD-6 is associated with altered slit protein nephrin expression in shed podocytes from preeclampsia.
Our study also demonstrated that the degree of podocyte injury and increased podocyte shedding are associated with severity of the diseases. This notion is supported by our observations that no podocyte was found in urine from mild preeclamptic pregnancies and more podocytes shed from nephrotic syndrome patients than from severe preeclamptic patients (Figure 2). Negative podocyte shedding in mild preeclampsia is based on the observation of no podocyte growth in culture, not based on nephrin staining. However, it is possible that shed podocytes from mild preeclampsia lack of proliferative potential and they fail to grow in culture, which might account for the negative podocyte in culture/shedding observed in our study. More podocyte clone formation in nephrotic syndrome than in severe preeclampsia suggests that kidney injury is worse in nephrotic syndrome. We did not specifically analyze the difference of shed podocytes between preeclampsia and nephrotic syndrome since other than less staining signals, the pattern for nephrin, podoplanin, PARD-3, and PARD-6 staining in shed podocytes is similar in both groups. However, the difference in marker protein expression may exist at the molecular levels between shed podocytes from preeclampsia and nephrotic syndrome, since different molecular weight bands for nephrin expression were detected by Western blot between cells from preeclampsia and nephrotic syndrome (Figure 5). The reason for this is not known. Defective nephrin synthesis and/or proteolytic effect resulted in truncated fragment might be an explanation. Although the question cannot be answered in the view of the paucity of data currently available, the finding of acute and transient podocyte loss in preeclampsia 18 support the expectation. Future studies that focus on molecular levels shall provide insightful information as to the specificity of podocyte shedding in preeclampsia.
Dedifferentiation and transdifferentiation are the 2 phenotypic changes in kidney podocytes underlying renal injury processes. 20 –23 The phenomena of proliferative potential and the cobblestone appearance of shed podocytes in culture suggest that dedifferentiation is likely the phenotype of shed podocytes from severe preeclampsia. Immature-looking podocytes with cobblestone appearance and loss of their characteristic markers are described as dedifferentiation phenotype, whereas podocytes co-expressing podocyte marker such as podocalyxin with CD68 are described as transdifferentiation phenotype for podocytes. 20 In our study, we did not specifically examine dedifferentiation or transdifferentiation phenotypes in shed podocytes from preeclampsia. However, it is very likely that dedifferentiation is probably the phenotypic changes in shed podocytes from severe preeclampsia, since cobblestone appearance (Figures 1 and 2) and loss of their specific functional marker protein nephrin and podoplanin expressions (Figure 4) point out this possibility, which warrants further investigation.
The mechanism underlying podocyte shedding in preeclampsia remains largely unknown. It has been proposed that vascular endothelial growth factor (VEGF) deficiency within the glomerulus in women with preeclampsia may lead to the disruption of podocyte and glomerular endothelium symbiosis. 24 In vitro studies also provided evidence that preeclamptic sera could induce podocyte alteration either directly or indirectly through glomerular endothelial cells. 1,25,26 Thus, it can be anticipated that both glomerular endothelial cells and adjacent podocytes contribute significantly to the renal barrier dysfunction in preeclampsia. 27
In conclusion, we have successfully cultured shed podocytes in urine specimen from women with severe preeclampsia. The shed podocytes are viable which provides a valuable tool for studying kidney podocyte injury in preeclampsia. Most significant finding of our study is that we demonstrated altered distribution and expression of slit specific protein nephrin, apical surface glycoprotein podoplanin, and polarity protein PARD-3 and PARD-6 in shed podocytes from severe preeclampsia. Although the mechanism underlying podocyte injury/shedding is not known, our data clearly indicate that podocyte integrity is severely disrupted in women with preeclampsia. Loss of foot process slit protein nephrin associated with the polarity protein changes found in shed podocytes further demonstrates that these functional proteins are critical for the kidney barrier function. At present, there is scarce information as to whether defective nephrin at podocyte foot process/slit is an early event to the loss of cell polarity of podocytes or vice versa. Further studies are warranted to dissect the molecular and cellular mechanisms responsible for the phenotypic changes and may provide useful markers for prediction and prevention of renal injury in preeclampsia.
Footnotes
Acknowledgment
This study was funded by a fellowship from the Malcolm Feist Cardiovascular Research Endowment, Louisiana State University Health Sciences Center – Shreveport, LA, USA.
The authors declared no potential conflicts of interests with respect to the authorship and/or publication of this article.