
Abstract
In this study, we show that myometrial transcriptional complexes consisting of Sp1, Sp3, histone deacetylase (HDAC)1/2, RbAp48, and mSin3A are recruited to 4 out of the 6 Sp1-4 sites within the Gαs promoter. Moreover disruption in the binding of these complexes via mithramycin administration results in a substantial decrease in expression of Gαs proteins in myometrial cell cultures. In many instances, these transcriptional regulatory complexes repress expression of genes having a high CG content within their promoter region. This repression can be attenuated by inhibition of HDAC activity by the class I/II HDAC inhibitor trichostatin A (TSA) resulting in increased gene transcription. However, although a substantial increase in Gαs protein levels was observed upon administration of TSA to primary cultures of human myometrial cells, this was not preceded by an increase in messenger RNA (mRNA) and thus an elevation in gene transcription. Importantly the increase in Gαs protein levels occurred via ubiquitination and inhibition of proteasomal activity, indicating that this pathway is also involved in regulating Gαs protein expression during pregnancy and parturition.
Keywords
Introduction
G-proteins play an integral part in transmitting the effects of external signals to the cell via G-protein coupled receptors (GPCRs). G-proteins form heterotrimers which consist of αβγ subunits that dissociate into a single Gα subunit and a Gβγ dimer upon agonist binding to receptors and subsequent conformational changes in receptor structure. 1 Both Gα and Gβγ subunits are then able to activate a variety of downstream intracellular second messenger pathways.
In the myometrium, G-proteins play an active role in the processes that control both quiescence and activation of myometrial smooth muscle cells during pregnancy and labour. 2 Binding of pro-quiescent hormones to their receptors, for example human chorionic gonadotrtopin (hCG), illicit a relaxant effect on the myometrium via a Gαs-dependent increase in adenylate cyclase (AC) activity followed by an increase in intracellular cyclic adenosine monophosphate (cAMP) levels and activation of protein kinase A (PKA). 3 In contrast, binding of oxytocin to its receptor leads to Gαq/11 activation and an increase in phospholipase C (PLC) activity and intracellular Ca2+ levels leading to uterine contractions. 4 Therefore, the balance between levels of Gα proteins which control relaxatory or contractile signalling pathways is believed to have a key impact on the regulation of myometrial activity during pregnancy and at the onset of labor. 2 In human myometrium, an increase in Gαs protein levels has been observed during the early stages of gestation and is maintained until the onset of labor, thereafter dropping to the levels observed in the nonpregnant (NP) myometrium. 3 , 5 This increase is functionally coupled to an increase in intracellular cAMP formation and therefore is thought to play a major role in maintaining myometrial relaxation during pregnancy. 5 Moreover, during pregnancy, specific spliced isoforms of Gαs containing additional serine residues are preferentially expressed in the myometrium, 6 which may be subject to phosphorylation by PKA and PKC and thus modify Gαs activity. 7 , 8
Although fluctuations in Gαs protein levels during pregnancy and labor have been well characterized in the human myometrium, the mechanisms controlling gene expression of Gαs are not fully understood. The Gαs promoter region has a high content of CG nucleotides (85%), with putative binding sites for CREB, Erg1, WT-1, and the specificity proteins Sp1-4 family 9 (www.generegulation.com/cgi-bin/pub/programs/alibaba). With respect to the latter, there are 4 canonical 5′-CCCGCCC-3′ and 2 noncanonical 5′-CCCTCCC-3′ Sp1-4 transcription factor binding sites present in the proximal region of the promoter. 9 The Sp1-4 proteins modulate gene expression by directly binding to the promoter via zinc fingers and serve as a binding platform for the formation of regulatory transcriptional complexes. Components of these complexes include HDACs 1 and 2, mSin3A, and RbAp46/48, and these regulators often act as repressors of gene expression. 10 In the myometrium, the human chorionic gonadotropin/luteinizing hormone receptor (hCG/LHR) and PKA regulatory subunit (RIIα) genes, which are components of the cAMP/PKA relaxatory pathway, appear to be regulated by Sp1-4 transcriptional complexes. Furthermore, inhibition of HDAC activity by trichostatin A (TSA) leads to de-repression of these genes, resulting in an increase in the expression levels of both messenger RNA (mRNA) and protein. 11 , 12 With respect to Gαs, phosphorylation of recombinant Sp1 protein by PKA appears to be required for binding to Sp1-4 elements in the promoter, 13 thus indicating that increased PKA activity may modulate Gαs gene expression by affecting the binding capacities of Sp1-4 transcriptional regulators.
The purpose of this study was to further investigate the role of Sp1-4 proteins and their transcriptional epigenetic coregulators in controlling the expression of the Gαs gene in the human myometrium. Using primary human myometrial cell cultures treated with the HDAC inhibitor TSA, we demonstrate that indeed transcriptional complexes consisting of Sp1-4 and HDACs 1/2 proteins bind to the proximal promoter region of the Gαs gene and that inhibition of HDAC activity by TSA resulted in an increase in Gαs protein expression. However, this increase was not preceded by an elevation in mRNA levels, suggesting that other molecular mechanisms were involved in regulating Gαs protein expression. In this context, we provide novel evidence to indicate that in human myometrial smooth muscle cells TSA inhibits proteosomal degradation of Gαs proteins, resulting in increased ubiquitinated isoforms and a subsequent rise in Gαs protein levels.
Materials and Methods
Tissue Collection
Written consent was obtained from all women, and ethical approval was granted by the North Tyneside Health Authority Ethics Committee.
Preparation of Primary Myometrial Cell Cultures
Primary cell cultures were established from NP myometrial tissue samples as described previously. 12 , 14 In all instances, cultures were determined to be greater than 95% myometrial smooth muscle cells as defined by Phaneuf et al. 14 Tissue samples were taken from uteruses removed from women undergoing hysterectomy due to benign gynecological conditions (38-58 years).
Protein Lysate Preparation
Lysates from myometrial cell cultures were prepared by mechanical homogenization in Western blotting (WB) buffer (62.5 TRIS-Cl pH 6.8, 2% sodium dodecyl sulphate [SDS], 10% sucrose) supplemented with proteases, and phosphatase inhibitors (Sigma, UK).
Western Blotting
Western blotting was carried out as described previously. 11 , 12 Total protein, 10 µg, was separated by 10% SDS-polyacrylamide gel electrophoresis (PAGE) and transferred by semi-dry blotting (Bio-Rad, Hertfordshire, UK) onto nitrocellulose membranes. Membranes were blocked in 5% fat-free milk phosphate-buffered saline (PBS)/0.1%Tween and then incubated with the specific antibodies (Table 1) in 1% fat free milk in PBS/Tween for 1 hour at room temperature. After washing, the membranes were incubated with the respective secondary horseradish peroxidase (HRP)-conjugated antibodies (DAKO, Glostrup, Denmark, 1:5000) for 1 hour at room temperature. Enhanced chemiluminescent reagent (GE, UK) was used to detect the bands. Immunodetected bands were scanned and quantified using the intelligent quantifier software package (BioImage, Ann Arbor, Michigan). Membranes were re-probed with an antibody against β-actin as a loading control.
Antibodies Used for Western Blotting, DAPAs, and Immunoprecipitation Assays
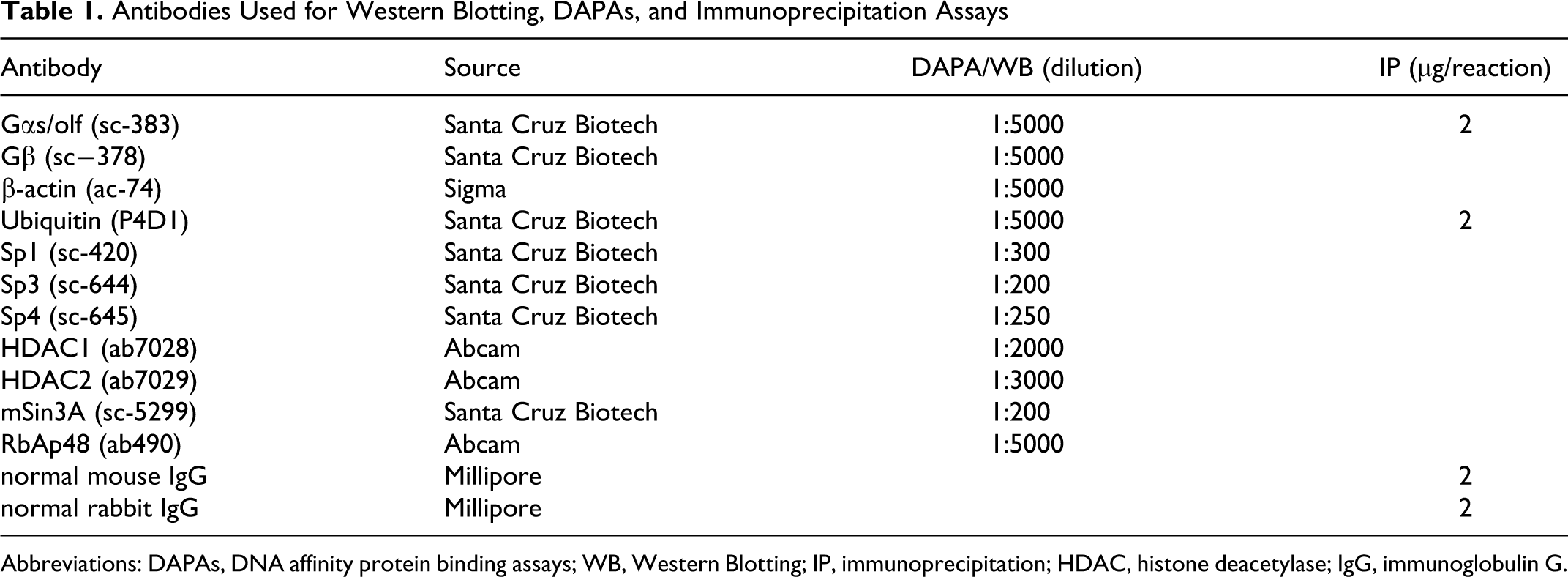
Abbreviations: DAPAs, DNA affinity protein binding assays; WB, Western Blotting; IP, immunoprecipitation; HDAC, histone deacetylase; IgG, immunoglobulin G.
DNA Affinity Protein Binding Assays
Cytoplasmic and nuclear extracts for DNA Affinity Protein Binding Assays (DAPAs) were prepared using a nuclear extraction kit (Chemicon, UK), according to the manufacturer’s instructions. Briefly, myometrial cells were collected and centrifuged in ice-cold PBS. Cell pellet was resuspended in ice-cold cytoplasmic lysis buffer and incubated for 15 minutes on ice and centrifuged (250g, 5 minutes, 4°C). The cell pellet was resuspended in cytoplasmic lysis buffer and drawn several times through a small gauge needle. After centrifugation (8000g, 20 minutes, 4°C), the resulting supernatant was kept as the cytosolic fraction. The remaining pellet was resuspended in nuclear extraction buffer and gently agitated at 4°C for 1 hour. The nuclear fraction was prepared by centrifugation (16 000g, 5 minutes) from which the supernatant was used. The DAPAs were performed as described in 12 . Cytoplasmic or nuclear protein extracts, 80 µg, were incubated with 35 pmol of double-stranded (ds), 5′-biotin-labeled oligonucleotides (MWG, AG, Ebersberg, Germany) for each of the respective 6 SpI-III (CG) elements (Table 2) in 500 µL binding buffer (12 mmol/L HEPES, pH 7.9, 4 mmol/L TRIS- HC, 60 mmol/L KCl, 5% glycerol, 0.5 mmol/L EDTA, 1 mmol/L DTT, 0.5% poly dI-dC) for 1 hour at 4°C to form protein/DNA complexes. These complexes were pulled down using 30 µL of streptavidin-coated magnetic beads (Active Motif, Rixensart, Belgium). Resins were then washed 5 times in binding buffer. Proteins were recovered from the complex by incubation in loading buffer containing SDS and β-mercaptoethanol, separated by SDS-PAGE and detected by immunoblotting using specific antibodies (see Table 1). To determine the binding specificity of proteins, 50× excess of the respective ds nonbiotinylated oligonucleotides was used as a specific competitor or 50× excess of the ds OCT1 oligonucleotides was used as a nonspecific competitor in the DAPA reactions.
Sequences of the Oligonucleotides Used for DAPAs
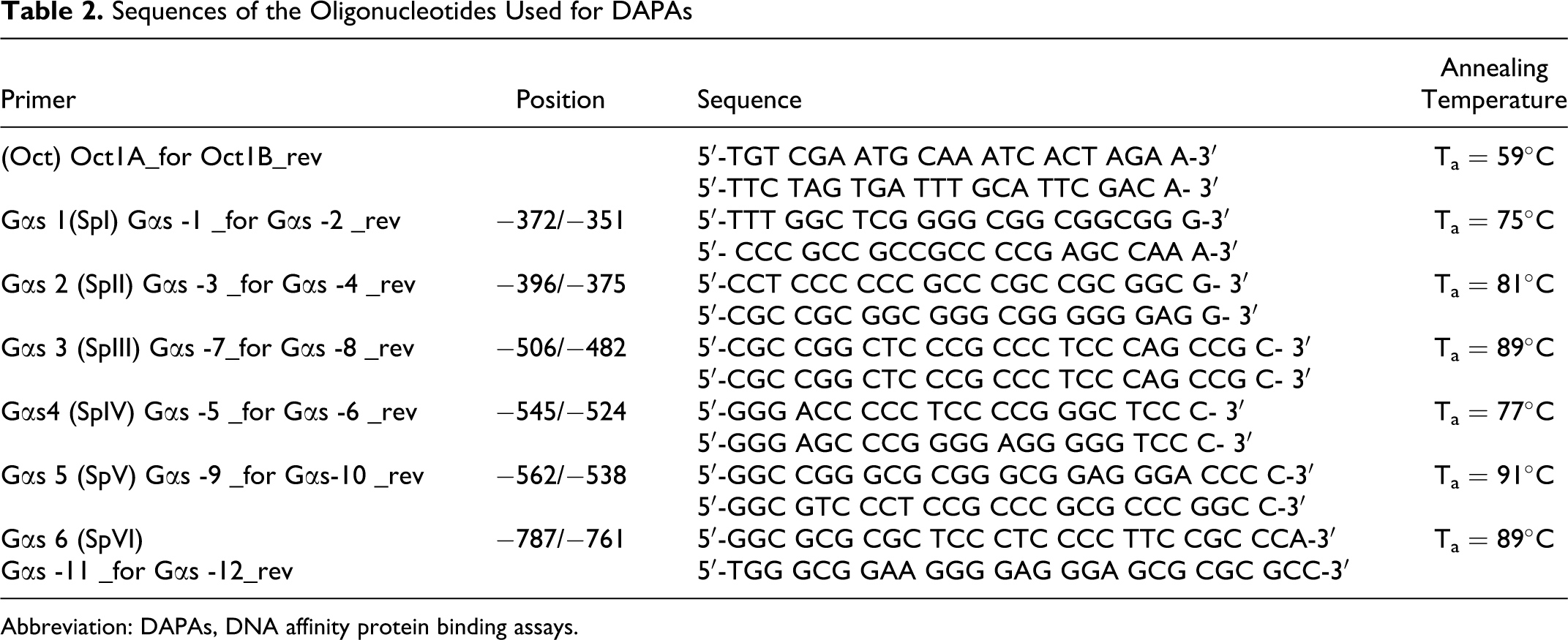
Abbreviation: DAPAs, DNA affinity protein binding assays.
Immunoprecipitation Assays
Cultured myometrial cells were washed and collected in ice-cold PBS. Cells were centrifuged at 1500 rpm for 5 minutes at 4°C. The cell pellet was homogenized in 500 µL of sucrose buffer (25 mmol/L Tris-Cl pH 7.6, 25 mmol/L sucrose, 1 mmol/L EDTA) using a plastic pestle, centrifuged for 10 minutes at 10 000 rpm at 4°C, and the resulting supernatant was used for subsequent experiments. Proteins, 500µg, were precleared with the rabbit or mouse immunoglobulin G ([IgG] Abcam, Cambridge, UK or Upstate, Watford, UK, respectively) in 5 volumes of HEPES buffer (25 mmol/L HEPES, 75 mmol/L KCl, 2.5 mmol/L MgCl2) with proteases inhibitors (Active Motif) and 15 µL of protein A coated magnetic beads (Invitrogen, UK) for 30 minutes at 4°C. In all cases, antibodies (Table 1) were added to the precleared proteins and allowed to form complexes overnight at 4°C. Protein/antibodies complexes were then recovered with 25 µL of protein A-coated magnetic beads and washed 4 times with HEPES buffer (150 mmol/L NaCl/0.5% detergent provided in the kit). Proteins were then recovered from the magnetic beads by boiling for 5 minutes in 20 µL of loading buffer and subjected to WB.
Transfection and Dual-Luciferase Reporter Assay
A pGαs2GL3 plasmid DNA promoter luciferase construct containing a 461-bp fragment of the Gαs gene promoter from −789 to −329 in relation to the translation initiation site of the Gαs promoter was used in the DNA transfection as described previously. 11 Myometrial cells grown in 35-mm culture plates to 70% confluence were transiently transfected with 8 µL of Trans IT-LT-1 (Mirus, Cambridge, UK) and 2 µg of sterile plasmid and 1 µg of control Renilla luciferase plasmid used as internal transfection control (Promega, Madison, WI, USA). Transfection with the empty pGL3-E vector was used as a control. Cultures were then incubated for 24 hours at 37°C to allow transfection to occur. After 24 hours, the medium supplemented with the respective treatments (330 nmol/L TSA or 1 µmol/L, 2 µmol/L, and 10 µmol/L mithramycin) for the times indicated. Data from 3 independent transfections, each in triplicate were collected using the dual-luciferase reporter assay system (Promega).
RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction
Total RNA was isolated from myometrial cell cultures harvested in 1 mL Tri Reagent (Sigma, UK)/10 cm2 culture plates according to the manufacturer’s instructions. Total RNA, 1 µg, was reverse transcribed using murine leukemia virus (MMLV) reverse transcriptase (Promega) and random hexanucleotide primers. Real-time analysis was used for quantitative analysis of Gαs mRNA expression levels. The Gαs primers and probes were designed using the File-builder 3.0 primer design software and ordered as an on-demand assay from Applied Biosystems, UK (Table 3). Changes in the Gαs mRNA expression levels after treatments were normalized to the 18S ribosomal RNA (rRNA) housekeeping gene. Real-time–polymerase chain reaction (RT-PCR) was performed using an ABI Prism 7000 sequence detector (Applied Biosystems, UK). All experiments were performed in 25 µL volume containing 2 µL of complementary DNA (cDNA) template, 1× TaqMan Universal PCR Master Mix (Applied Biosystems, UK), 1.25 µL of each 20x primers and TaqMan probe. Each sample was run in triplicates using following protocol: 2 minutes incubation at 50°C, an initial denaturation for 10 minutes at 95°C, and the subsequent 40 cycles (denaturation at 95°C for 15 seconds, primer annealing and elongation at 60°C for 1 minute). Each experiment included a negative control, water replacing the cDNA template. The experiment was carried out with 4 independent tissue cultures isolated from 4 individual patient samples. Expression levels for each individual sample were calculated by the ΔΔCt method. 15
The Sequences of the Primers and the Probes Used in Real-Time Polymerase Chain Reaction

Abbreviations: FAM/NON, 6-carbonylfluorescein; FAM/TAMRA, dihydrocyclopyrroloindole tripeptide.
Data Analysis
Data are presented as mean ± standard error of the mean (SEM). Analysis of variance (ANOVA)-Bonferroni was used with P < .05 considered statistically significant.
Results
Sp1/Sp3, HDACs 1/2, RbAp48, and mSin3A Form Transcriptional Complexes Within the Proximal Gαs Promoter
In vitro binding of recombinant Sp1 protein to cis elements within the Gαs gene promoter has been previously described. 13 However, binding of Sp1-4 proteins and their transcriptional coregulators which bind the CG-rich region of the Gαs gene in human myometrial cells has not been investigated. In order to determine the nature of these Sp1-4 complexes, DAPA analysis was performed for all 6 Sp1-4 sites present within the Gαs gene promoter employing nuclear and cytoplasmic extracts obtained from NP myometrial cell cultures: These cis elements included Sp1-4 (I): −372 to −351 bp, Sp1-4 (II): −396 to −375 bp, Sp1-4 (III): −506 to −482 bp, Sp1-4 (IV): −545 to −524 bp, Sp1-4 (V): −562 to −538 bp, and Sp1-4 (VI): −787 to −761 bp (see Figure 1A). Figure 1B shows DAPA analysis for HDAC1, HDAC2, RbAp48, and mSin3A binding to the Sp1-4 I Gαs binding domain. Supplementary Figure 1A shows further results for the Sp1-4, I, II, III, and VI binding sites within the Gαs gene promoter. Specific Sp1 and Sp3 protein binding was only evident for Sp1-4 I, II, III, and VI binding domains (see Figure 1B and Supplementary Figure 1A). The specificity of this binding was confirmed by a decrease in the binding of these proteins in the presence of specific competitors (see Figure 1B and Supplementary Figure 1A). Note that no specific binding of Sp4 was observed for any of the Gαs Sp1-4 binding cis elements (see Supplementary Figure 1A and B) and that Sp2 is poorly expressed in myometrial cells. 11 , 12 Recruitment of HDAC1, HDAC2, RbAp48, and mSin3A was observed in each instance for the Sp1-4 I, II, III and VI sites (see Figure 1B and supplementary Figure 1A), thus suggesting that these form transcriptional complexes with Sp1 and Sp3 proteins within the myometrial Gαs promoter region. No detectable complexes were observed when cytoplasmic extracts were used, suggesting specificity of these complexes to the nuclear compartment of cells. Furthermore, no specific binding of Sp1 and Sp3 proteins was observed for the Sp1-4 elements IV and V (see Supplementary Figure 1B).
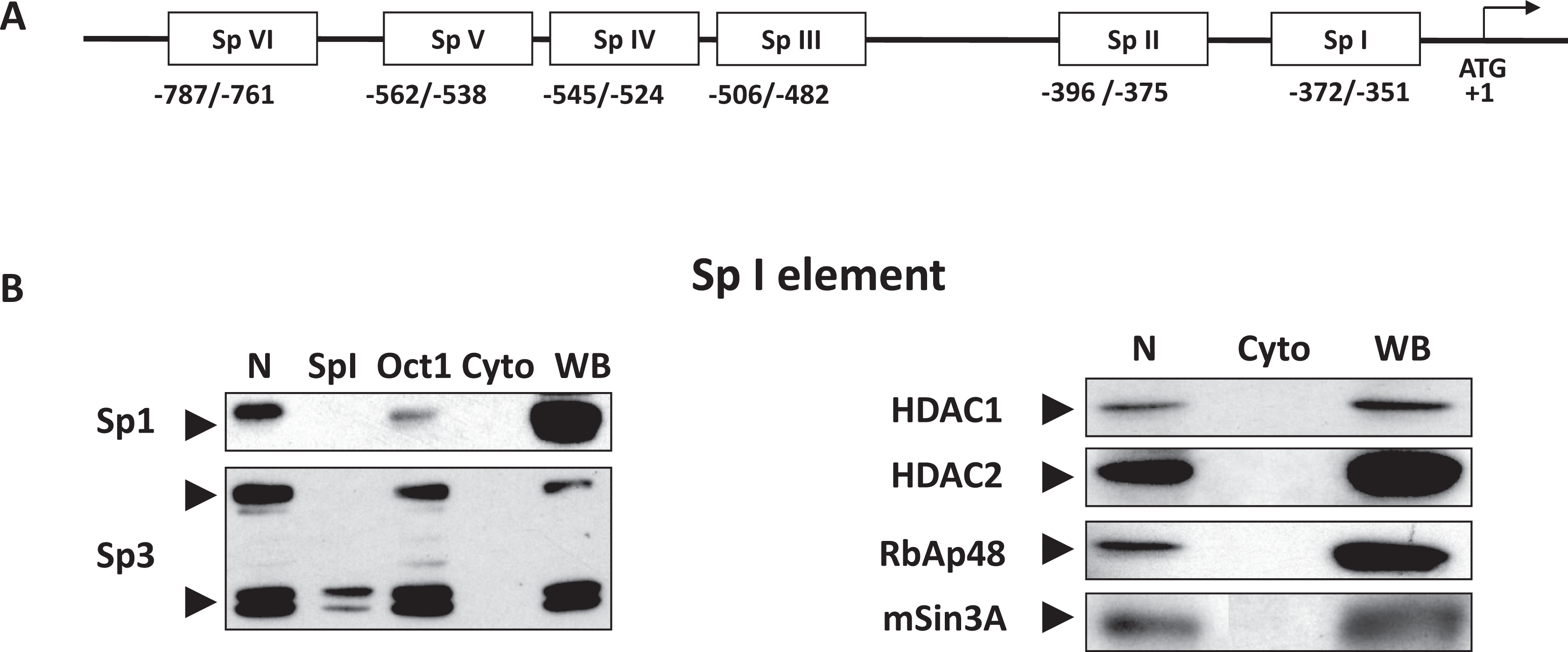
In vitro DAPA analysis indicates that the Sp1-4 binding sites I, II, III, and VI within the Gαs promoter specifically bind multiprotein complexes containing Sp1, Sp3, HDACS 1/2, RbAp48, and mSin3A. A, Schematic representation of the Gαs proximal promoter region showing the putative transcriptional regulatory CG rich cis elements. The transcriptional initiation site (+1) and the nucleotide position of the six Sp1-4 (I-VI) binding elements in the promoter region are as shown. B, DAPA analysis of binding of transcriptional regulators to the Sp1-4 I element within the Gαs gene promoter.
Binding of Sp1 and Sp3 Proteins Affects the Basal Expression of Gαs Proteins
We have previously shown that in human myometrial cells Sp1/Sp3 protein complexes regulate basal expression of the PKA regulatory subunit RIIα gene. 12 In order to define that this also occurred for basal expression of Gαs, myometrial cells were treated with mithramycin and Gαs protein levels monitored. Here, mithramycin can reversibly bind to CG-rich stretches of dsDNA, containing Sp1-4 binding sites and selectively interfere with the binding of these proteins to these elements to affect gene expression. Western blotting analysis of proteins from NP myometrial cells treated for 6 hours with 1, 2, and 10 µmol/L mithramycin showed a significant decrease in Gαs protein levels at all concentrations (Figure 2A). The concentration of mithramycin used in this study were based on earlier reports showing decreased Sp1 binding in the myometrium 12 as well as in the macrophage cell line RAW264 after 6 hours of treatments with this reagent. 16 Employing a Gαs luciferase construct, detailed in 13 in transient transfection of myometrial cell cultures treated with 1, 2, and 10 µmol/L mithramycin, for 6 hours also resulted in a significant decrease in luciferase activity (Figure 2B).
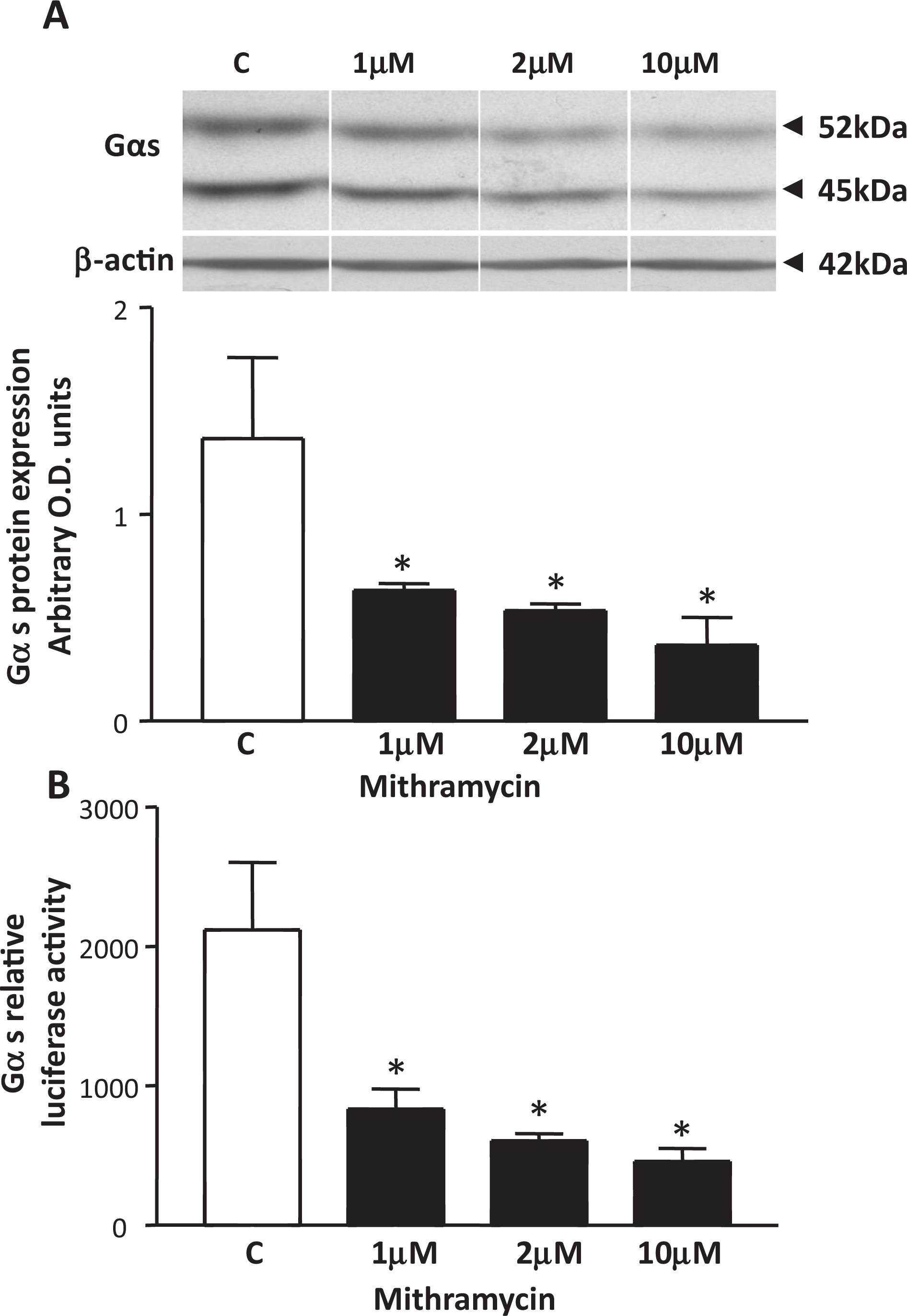
The effect of mithramycin on Gαs protein expression and Gαs luciferase promoter activity in myometrial cell cultures. A, Western blotting analysis indicated that mithramycin treatment of myometrial cells at 1, 2, and 10 µmol/L concentrations for 6 hours significantly decreases Gαs protein levels. Data are presented as means ± SEM (n = 3 individual patient samples repeated in triplicate), *p<0.05 (ANOVA Bonferroni) control C versus mithramycin. β-actin staining was used as loading control. Samples were run on the same gel but were not contiguous (lines). B, Primary human myometrial cell cultures transiently transfected with the Gαs luciferase plasmid and treated with 1, 2, and 10 µmol/L mithramycin, respectively, for 6 hours significantly decreased luciferase activity. Data are presented as means ± SEM (n = 3 individual patient samples repeated in triplicate). *P < .05 (ANOVA Bonferroni) control C versus mithramycin. SEM indicates standard error of the mean; ANOVA, analysis of variance.
Trichostatin A Increases Gαs Protein Expression
Inhibition of HDACs 1 and 2 by the pan class I/II HDAC inhibitor TSA has previously been shown to lead to an increase in mRNA and protein expression of both the hCG/LH receptor and the PKA regulatory subunit RIIα genes in human myometrial cell cultures. 11 , 12 Since HDACs 1 and 2 are recruited to the myometrial Gαs promoter, the effect of TSA on Gαs protein expression was investigated. Initially acetylation of histone H3 was monitored to verify that TSA after 1, 6, and 24 hours could inhibit HDAC activity; this was found to be the case as described by us in. 12 Subsequently the effect of TSA on Gαs protein expression was then analyzed. Levels of both 52- and 45-kDa isoforms of Gαs protein in myometrial cell cultures were significantly increased after TSA treatment for 1 and 6 hours (Figure 3A). β-actin staining was used as internal loading control.
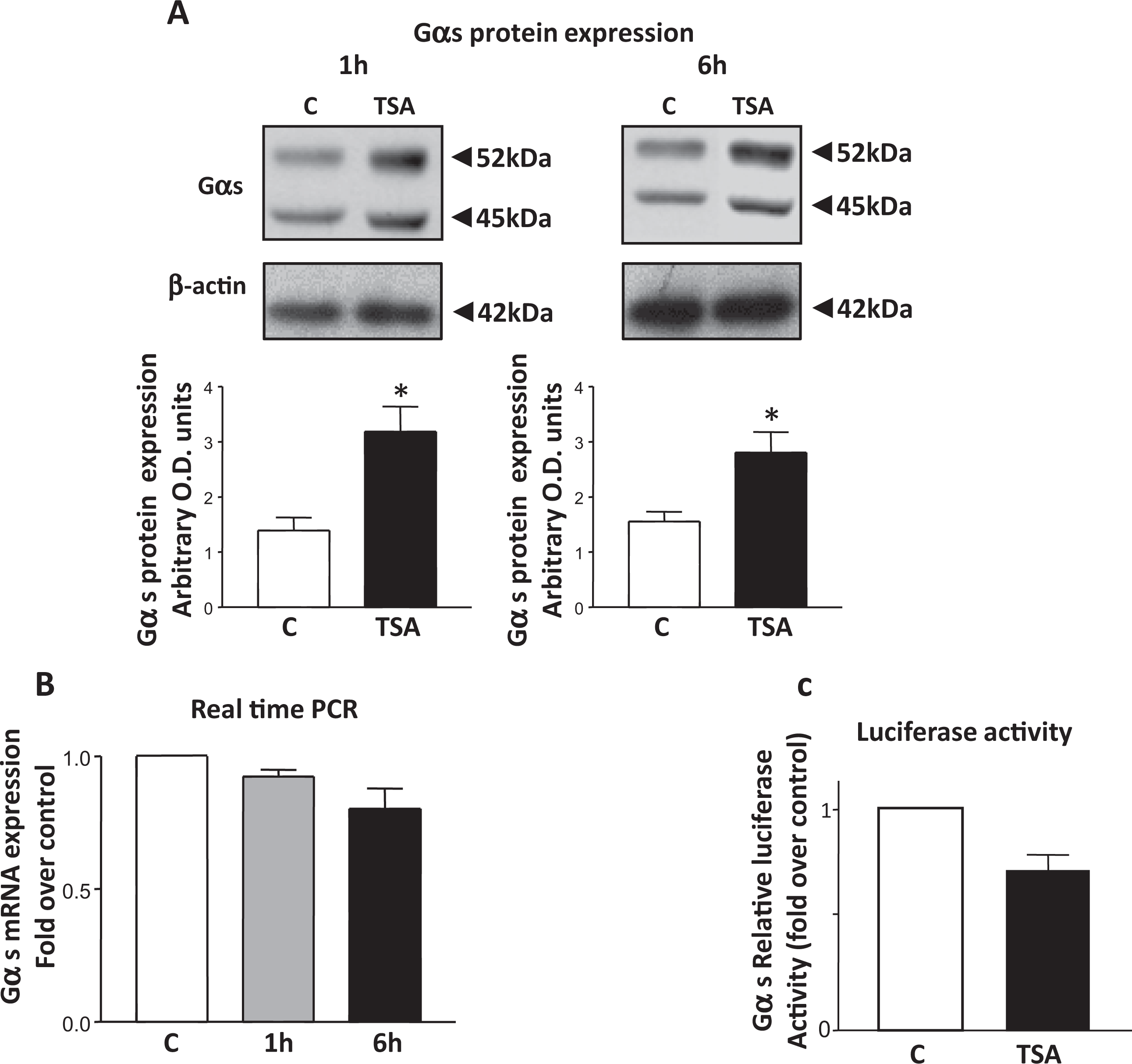
The effect of TSA treatment on Gαs protein/mRNA expression and Gαs luciferase promoter activity in primary human myometrial cell cultures. A, Western blotting analysis indicated that TSA (330 nmol/L) treatment of myometrial cells for 1 hour and 6 hours significantly increased Gαs protein expression. Data are presented as means ± SEM (n = 3 individual patient samples repeated in triplicate) *P < .05 (ANOVA Bonferroni) control C versus TSA. β-actin staining showed equal lane loading. B, Real-time PCR analysis detected no significant (ANOVA Bonferroni, P > .05) change in Gαs mRNAlevels after 1 hour or 6 hours of treatment with TSA (330 nmol/L) compared to control C. Data are presented as means ± SEM (n = 4 independent tissue cultures isolated from four individual patient samples).C, Myometrial Gαs luciferease assays showed no significant change (ANOVA Bonferroni P > .05) in luciferase activity after 6 hours of treatment with TSA (330 nmol/L) compared to control C. Data are presented as means ± SEM (n = 3 individual patient samples repeated in triplicate). SEM indicates standard error of the mean; ANOVA, analysis of variance; TSA, trichostatin A.
Trichostatin A Does Not Increase Gαs mRNA Levels
To determine whether the increase in Gαs protein levels after TSA treatment was due to increased transcription and a subsequent rise in mRNA levels, Gαs mRNA levels were determined via real-time RT-PCR after 1 and 6 hours of treatment with TSA. Here no change in Gαs mRNA expression levels was observed (Figure 3B), indicating that inhibition of HDAC 1/2 activity was not associated with an increase in the transcription of the Gαs gene. Employing the Gαs luciferase construct in transient transfection of myometrial cells no change in luciferase activity was also observed when the cells were treated with TSA for 6 hours (Figure 3C), suggesting no change in Gαs transcriptional activity after inhibition of HDACs. Together these findings indicated that the observed increase in Gαs protein levels was not due to chromatin remodeling and changes in the gene expression.
The Effect of TSA and MG132 on Ubiquitination and Proteasomal Degradation of Gαs Proteins
The lack of correlation of increased Gαs protein levels with a rise in Gαs mRNA expression after TSA treatment implied that other processes could be involved in the observed increase in the expression of Gαs proteins. In human osteosarcoma cells, TSA treatment has been shown to result in the appearance of polyubiquitinated forms of Gαs and accumulation of Gαs proteins due to the inhibition of proteasomal-mediated degradation. 17 Consequently, regulation of the myometrial ubiquitin/proteasome pathway by TSA and its effect on Gαs protein levels was investigated. MG132 (Z-Leu-Leu-Leu-al), a potent and specific proteasome inhibitor that reduces degradation of ubiquitinated protein via specific inhibition of the 26S proteasome, 18 was also used to treat myometrial cells, and Gαs protein levels monitored by WB. Similar to TSA, MG132 treatment resulted in an increase in Gαs protein expression after 6 hours of treatment (Figure 4A), indicating that proteasomal degradation plays a role in regulating the expression of Gαs proteins. Additionally, 6 hours of treatment of myometrial cells with either MG132 or TSA resulted in the accumulation of high-molecular-weight polyubiquitinated proteins (Figure 4A), suggesting also that TSA inhibits proteasomal activity in these smooth muscle cells.
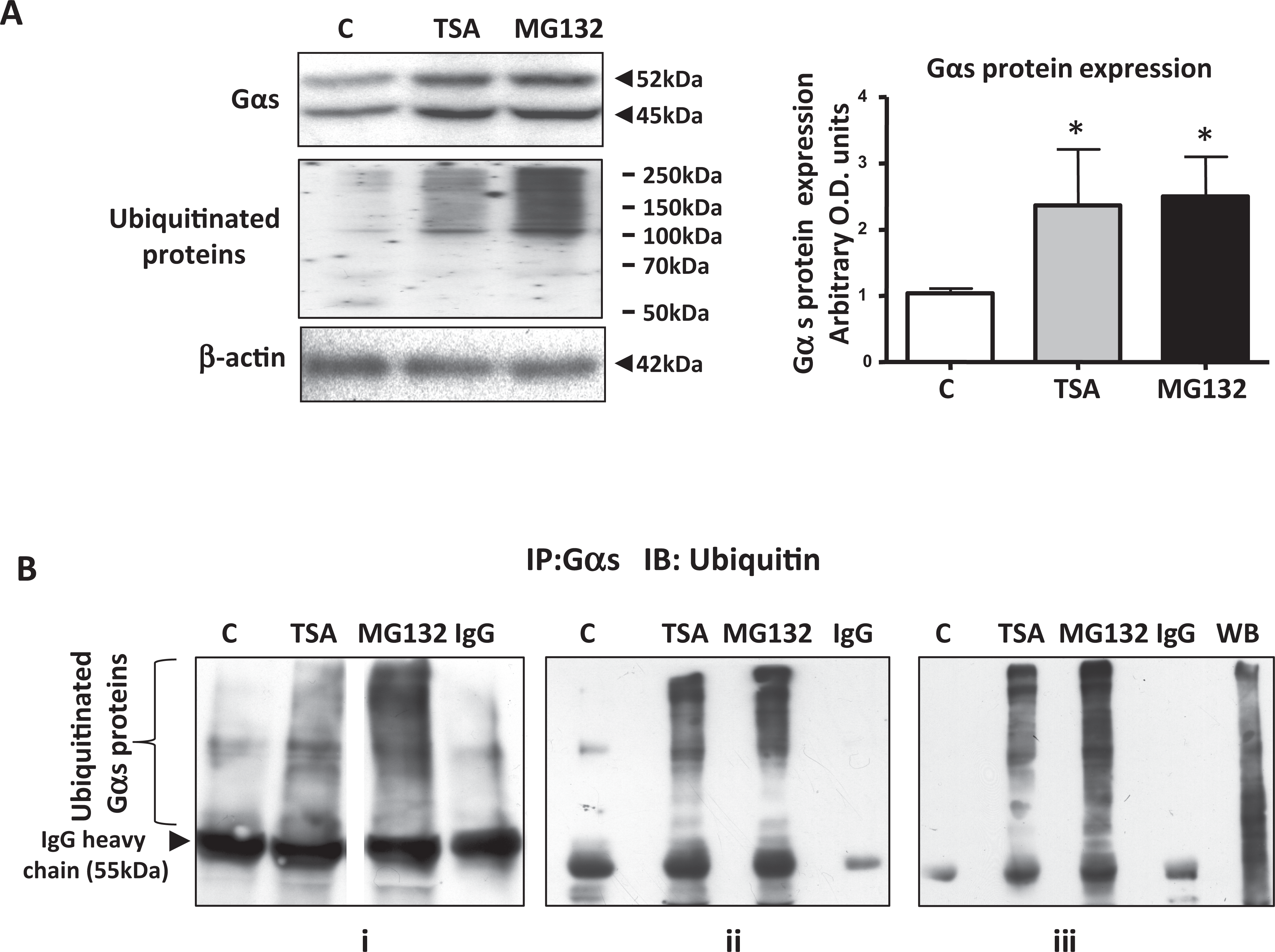
Both TSA and MG132 increase Gαs protein expression and its ubiquitination in human myometrial cells. Western blotting analysis of total protein extracts from smooth muscles cells indicated an increase in Gαs protein levels and total levels of ubiquitinated proteins (A) after 6 hours of treatment with both TSA (330 nmol/L) and the specific proteasome inhibitor MG132 (10 µmol/L) compared to vehicle treated control C. Data are presented as means ± SEM (n = 3 individual patient samples repeated in triplicate), *P < .05 (ANOVA Bonferroni) control C versus TSA and MG132. β-actin was used a loading control. B, Immunoprecipitation (IP) with the Gαs antibody and immunoblotting (IB) with the ubiquitin-specific antibody showed an increase in high-molecular-mass polyubiquitinated Gαs proteins after 6 hours of treatment with TSA (330 nmol/L) and MG132 (10 µmol/L) compared with vehicle treated control (C). Rabbit total IgG was used as a negative control in the IP reaction. Triplicate IPs from separate cell cultures (panels i-iii) are shown. In (i) samples were run on the same gel but were not contiguous (line). In (iii) WB indicates Western blotting control of ubiquitinated proteins in myometrial cell lysates (10 μg). SEM indicates standard error of the mean; ANOVA, analysis of variance; TSA, trichostatin A.
The presence of ubiquitinated forms of Gαs protein was further investigated by employing immunoprecipitation (IP) assays. Immunoprecipitation with the Gαs antibody and subsequent WB with a ubiquitin antibody of myometrial protein lysates prepared from cells treated with TSA or MG132 for 6 hours indicated that these treatments were associated with the appearance of high-molecular-weight polyubiquitinated Gαs isoforms (Figure 4B). Since inhibition of proteasome activity has been shown to be due to a decrease in the expression of either the 1 or 5β subunits of the proteasome by TSA. 19 Western blotting with a β1-proteasome subunit antibody was performed with protein lysates from cells treated for 6 hours with TSA. No decrease in the expression of the β1 subunit was observed (see Supplementary Figure 2).
Discussion
In this study, we have shown that transcriptional complexes consisting of Sp1, Sp3, HDAC1/2, RbAp48, and mSin3A appear to be recruited to 4 of the 6 Sp1-4 cis elements within the Gαs promoter. Moreover, disruption of binding of these complexes via mithramycin administration resulted in a substantial decrease in the expression of Gαs proteins in myometrial cell cultures. These events are similar to that observed for the hCG/LH receptor and RIIα genes in the human myometrium. 11 , 12 Here, Sp1, Sp3, HDAC1/2, and mSin3A complexes not only play an important role in regulating basal expression of these genes, but inhibition of HDAC activity by TSA leads to a substantial increase in both mRNA and protein expression. 11 , 12 This latter effect is in contrast to that observed here for Gαs expression where only an increase in protein expression is seen which is not preceded by elevated mRNA levels (nor activation of a Gαs promoter as measured by an in vitro luciferase assay). Consequently attenuation of HDAC activity by TSA at Sp1-4 binding domains within the Gαs gene promoter does not lead to an increase in transcription of the gene.
However, the data presented in this study indicates that the increase in Gαs protein levels observed after TSA treatment arises due to the inhibition of proteasomal activity. Importantly we show that myometrial Gαs protein isoforms are not only ubiquitinated as consequence of TSA treatment but also by the specific proteasomal inhibitor MG132. It is now well evidenced that the ubiquitination of proteins is an essential step that is required for labeling proteins for proteasomal degradation and that the accumulation of ubiquitinated proteins is an indicator of proteasome inhibition. 17 , 20 Ubiquitination and proteasomal degradation have been shown to take part in trafficking and degradation of a number of membrane proteins and receptors. 21 Furthermore, the protein stability of several G protein species has been reported to be regulated by ubiquitination. 21–24 With respect to Gαs, pharmacological inhibition of proteasome activity with MG132 in human osteosarcoma cells and Gαs transfected COS-7 results in increased Gαs protein levels and AC activity as well as in the prevention of β-adrenergic receptor-mediated downregulation of Gαs proteins. 17 However, the possibility exists that, since ubiquitination can change protein activity, the observed increase in AC activity 17 might be due to a change in Gαs activity as a result of ubiquitination. In this context increased myometrial Gαs expression is also associated with increased AC activity. 5 In contrast to ubiquitination affecting Gαs protein stability and thus its level of expression Kou et al 25 have recently shown that simvastatin treatment of vascular endothelial cells significantly decreases Gαs abundance by specifically inhibiting Gαs translation mediated by Akt/mTOR/eIF4/4EBP.
It is thought that the activity of the proteasome can be inhibited by TSA treatment in at least 2 ways. First, TSA may negatively regulate the expression of the catalytic β-subunits of the proteasome at both mRNA and protein levels 19 or second by increased acetylation that may affect stability and maturation of proteasomal subunits. 26 Additionally, acetylation may directly impair the performance of enzymes of the ubiquitination pathway. 26 In this present study, the levels of the β1-proteasomal subunit remained unchanged after TSA treatment. Consequently further experiments are required to elucidate the exact mechanism by which TSA causes inhibition of proteasomal activity in myometrial smooth muscle cells.
In conclusion, our data indicates that Gαs proteins are a proteasomal substrate in the human myometrium and thus liable to be degraded via the ubiquitin/proteasome pathway. Consequently proteasomal activity would also appear to be involved in regulating myometrial Gαs protein expression during pregnancy and parturition.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Work in this laboratory is funded by grants (G0800202 and G0900525) made available from the MRC (UK).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.